





(C) 2013 Anne I. Gondim. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
We provide the first annotated checklist of ophiuroids from the continental shelf of the State of Paraíba, northeastern Brazil. Identification keys and taxonomic diagnoses for 23 species, belonging to 14 genera and 8 families, are provided. The material is deposited in the Invertebrate Collection Paulo Young, at the Federal University of Paraíba. Ophiopsila hartmeyeri represents the first record for the northeastern region of Brazil, while Ophiolepis impressa, Ophiolepis paucispina, Amphiura stimpsoni, Amphiodia riisei, Ophiactis quinqueradia, Ophiocoma wendtii and Ophionereis olivaceae are new records for the State of Paraíba. The number of species known for the state was increased from 16 to 23, representing approximately 17% of the species known for Brazil and 54% of the species known for northeastern Brazil. The recorded fauna has a large geographical and bathymetrical distribution.
Echinoderms, Ophiurida, checklist, Brazilian coast, distribution
The class Ophiuroidea includes the most agile and diverse animals within the phylum Echinodermata (
The study of ophiuroids in Brazil started with Lyman (1875), who described the results of the Hassler Expedition, off Bahia and Rio de Janeiro (
The coast of the State of Paraíba (Fig. 1) has 138 km and its continental shelf is narrow, shallow and relatively plane, with a mean width of 30 km, becoming broader in a north-south direction (
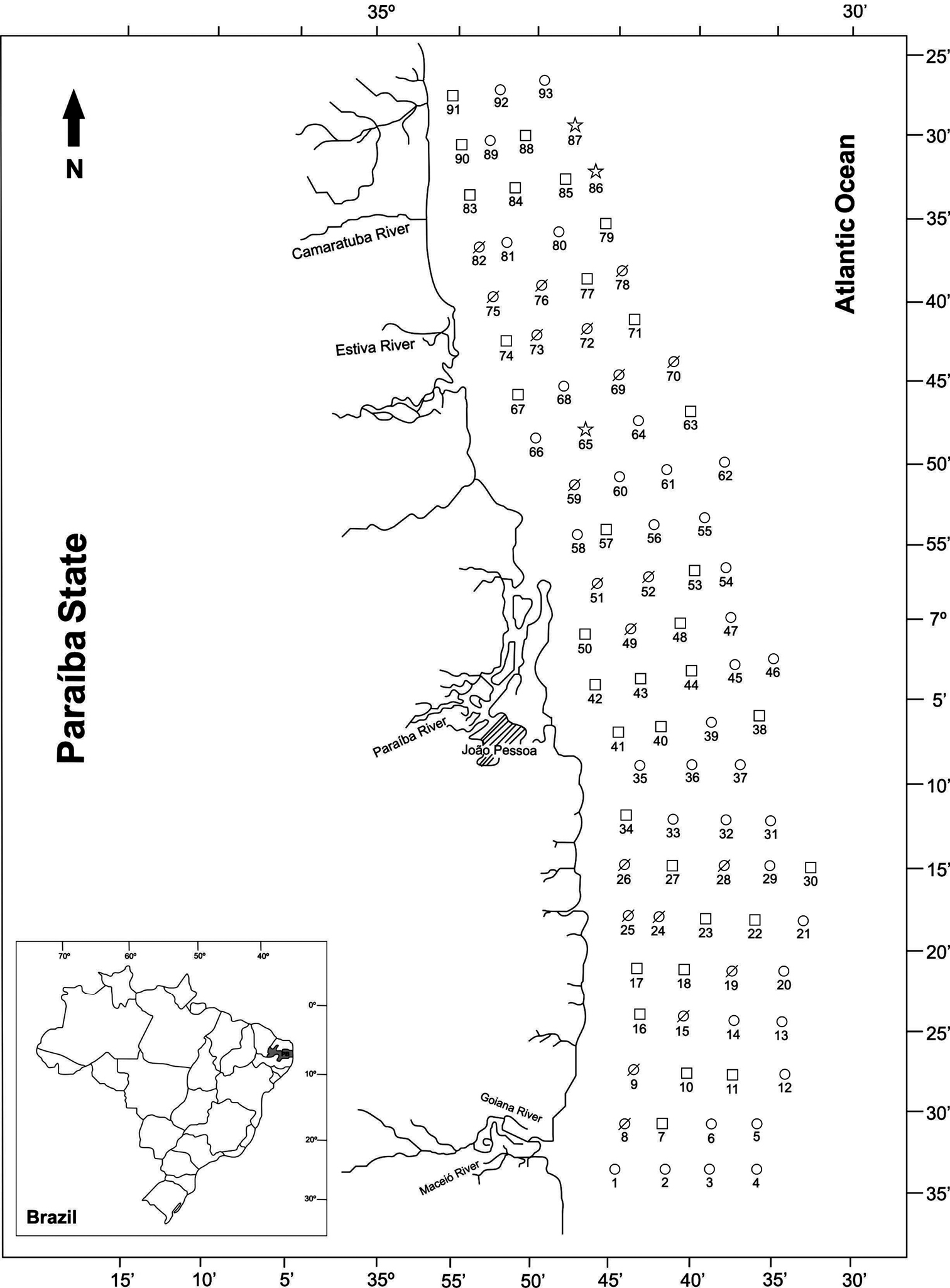
Collecting stations of Project Algae along the coast of the State of Paraíba, northeastern Brazil, with indication of the abundance of specimens in each site. Ø = absent in the collection point, □ = occurrence of 1 to 10 specimens, ○ = occurrence of 11 to 50 specimens, and ☆ = occurrence of more than 50 specimens.
Along the continental shelf of the Paraíba State, bottoms of calcareous algae prevail beyond the isobath of 20 m, with a predominance of the ramified corallinacean alga Halimeda Lamoroux, 1812 (
During the 1980s the Superintendance for the Development of the Northeast (SUDENE) developed the Algae Project along the continental shelfs of the States of Rio Grande do Norte and Paraíba, in order to map and collect banks of calcareous algae in this region. The thoroughest coverage of this project was along the State of Paraíba, and representative samples of ophiuroids were obtained and deposited in the Federal University of that state.
Our aim is to provide a checklist and identification keys for the species of ophiuroids that inhabit the continental shelf of the State of Paraíba, describing species from based on the collected material and summarizing data on their ecology and distribution.
The studied material belongs to the Invertebrate Collection Paulo Young, Department of Systematics and Ecology, Federal University of Paraíba (CIPY/DSE–UFPB). Collections were made in 1981 during the Algae Project, on the continental shelf of the State of Paraíba, between coordinates 6°58'S, 34°46'W and 7°34'S, 34°45'W, between the isobaths of 10 and 35 m. The animals were captured with dredges at 93 stations positioned along 23 transects perpendicular to the coast (Fig. 1).
For taxonomic identifications, specimens were dried, observed with a dissecting microscope Olympus SZ40 and identified with the works of
Abbreviations: dd–disk diameter. Spec–specimens.
Acronym: UFPB.Ech.–Echinodermata Collection of Federal University of Paraíba.
The fauna of ophiuroids recorded along the shelf of the State of Paraíba contains species known to have a wide geographical and bathymetrical distribution, occurring, in general, along a considerable extension of the coast of Brazil. The number of known species in the State of Paraíba increased from 16 to 23, corresponding to 17% of the Brazilian species and 54% of the northeastern species.
We examined 647 samples (totaling 1.379 specimens), identifying 23 species, 14 genera, 8 families, and 1 order of Ophiuroidea (Tab. 1, Tab. 2, Supplementary Material).
Class Ophiuroidea Gray, 1840
Order Ophiurida Müller & Troschel, 1840
Family Ophiomyxidae Ljungman, 1867
Ophiomyxa flaccida (Say, 1825)
Family Ophiolepididae Ljungman, 1867
Ophiolepis impressa Lütken, 1859
Ophiolepis paucispina (Say, 1825)
Family Amphiuridae Ljungman, 1867
Amphiodia planispina (von Martens, 1867)
Amphiodia riisei (Lütken, 1859)
Amphipholis januarii Ljungman, 1866
Amphipholis squamata (Delle Chiaje, 1828)
Amphiura stimpsoni Lütken, 1859
Ophiocnida scabriuscula (Lütken, 1859)
Ophiophragmus brachyactis H.L. Clark, 1915
Ophiostigma isocanthum (Say, 1825)
Family Ophiotrichidae Ljungman, 1867
Ophiothrix (Ophiothrix) angulata (Say, 1825)
Family Ophiactidae Matsumoto, 1915
Ophiactis quinqueradia Ljungman, 1872
Ophiactis savignyi (Müller & Troschel, 1842)
Family Ophionereididae Ljungman, 1867
Ophionereis reticulata (Say, 1825)
Ophionereis squamulosa Koehler, 1914
Ophionereis dolabriformis John & A.M. Clark, 1954
Ophionereis olivacea H.L. Clark, 1900
Family Ophiocomidae Ljungman, 1867
Ophiocoma echinata (Lamarck, 1816)
Ophiocoma wendtii Müller & Troschel, 1842
Ophiopsila hartmeyeri Koehler, 1913
Family Ophiodermatidae Ljungman, 1867
Ophioderma appressa (Say, 1825)
Ophioderma cinerea Müller & Troschel, 1842
| 1 | Presence of a clump of dental papillae at the apex of the jaw (Fig. 2h, 10c) | 3 |
| – | Without dental papillae | 2 |
| 2 | One pair of infradental oral papillae on apex of jaw (Fig. 4c) | Amphiuridae |
| – | One apical papilla on apex of jaw (Fig. 9c, 13c) | 4 |
| 3 | A continuous series of oral papillae (Fig. 11c) | Ophiocomidae |
| – | Without oral papillae (Fig. 2h) | Ophiotrichidae |
| 4 | Disk and arms covered with a thick and naked tegument | Ophiomyxidae |
| – | Disk covered with scales, granules or spines | 5 |
| 5 | Two pairs of bursal slits in each interradius (Fig. 13b, g) | Ophiodermatidae |
| – | One pair of bursal slits in each interradius (Fig. 3b, 8b, 9b) | 6 |
| 6 | Disk covered only with scales. Oral papillae in continuous series | 7 |
| – | Disk covered by scales and spines. Presence of a diastema separating the lateral oral papillae from the apical papillae (Fig. 8b, i) | Ophiactidae |
| 7 | Dorsal scales of disk thin and imbricating (Fig. 9a, f, 10a, f). Genital papillae present or absent (Fig. 9b, 10b) | Ophionereididae |
| – | Dorsal scales of disk thick and imbricating (Fig. 3a). Dorsal arm plate present or absent | Ophiolepididae |
| 1 | accessory dorsal arm plates small, restricted to the first segments (Fig. 3a, f). Four to five arm spines | Ophiolepis impressa |
| – | All arm segments with accessory dorsal plates (Fig. 3g, j), except near tip. Two arm spine | Ophiolepis paucispina |
| 1 | Distal oral papilla never opercular | 4 |
| – | Distal oral papilla opercular (Fig. 5i, 7j) | 2 |
| 2 | Disk covered by scales. Two perpendicular tentacle scales | 3 |
| – | Disk covered by small papillae (Fig. 7f, h). Two almost parallel tentacle scales | Ophiostigma isocanthum |
| 3 | Three or four arm spines, the second and third with two hyaline denticles at tip (Fig. 5f). Radial shields narrow and long (Fig. 5a) | Amphipholis januarii |
| – | Three arm spines with tip tapering (Fig. 5j). Radial shields slightly longer than broad (Fig. 5g) | Amphipholis squamata |
| 4 | Oral papillae continuous (Fig. 6g). Two tentacle scales | 5 |
| – | Oral papillae separated from infradentals by a gap (Fig. 6c). Single reduced tentacle scale (Fig. 6e) | Amphiura stimpsoni |
| 5 | Radial shields divergent (like a sheep hoof) (Fig. 6e). Three arm spines slightly flattened | Ophiocnida scabriuscula |
| – | Radial shields contiguous, separated only proximally | 6 |
| 6 | With a fence of broadened papillae on margin of disk (Fig. 7a). Radial shields small and rounded | Ophiophragmus brachyatis |
| – | Without spines on margin of disk | 7 |
| 7 | Primary scales not evident (Fig. 4a). Three arm spines compressed, flattened and blunt | Amphiodia planispina |
| – | Primary scales very evident (Fig. 4f). A well marked row of large scales in interbrachial region. Three short and blunt arm spines, compressed in median and distal region | Amphiodia riisei |
| 1 | With six arms and some spines scattered over the dorsal disk (Fig. 8g). One or two oral papillae (Fig. 8i) | Ophiactis savignyi |
| – | With five arms and some small spines usually restricted to the ventral and lateral sides of the disk (Fig. 8a). Two or three oral papillae | Ophiactis quinqueradia |
| 1 | Genital papillae present (Fig. 9b) | 2 |
| – | Genital papillae absent (Fig. 10b) | 3 |
| 2 | A distinct line forming a reticulated pattern on the aboral surface of the disk (Fig. 9a). Arm spines markedly compressed, with apex blunt. With dark bands on arm segment separated by three-six light bands (Fig. 9a, d) | Ophionereis reticulata |
| – | Dark blotches on aboral surface of disk. Arm spines markedly compressed. Dark bands occupying two or three arm segments, separated by one light bands | Ophionereis squamulosa |
| 3 | Sometimes several superposed accessory dorsal arm plate are observed. Three arm spines longer than arm segment (Fig. 10i), match-shaped | Ophionereis olivacea |
| – | Accessory dorsal arm plates small (Fig. 10a, d). Three long, thin spines on arm (Fig. 10d, e) | Ophionereis dolabriformis |
| 1 | Disk covered by scales and granules (Fig. 11a, e) | 2 |
| – | Disk covered only by scales (Fig. 12a). Four to 6 arm spines, ventral spines longest and slightly curved | Ophiopsila hartmeyeri |
| 2 | Dorsal arm spines robust and broadened (Fig. 11d). Two tentacle scales | Ophiocoma echinata |
| – | Dorsal arm spines long and slender (Fig. 11h). One tentacle scale on first arm segments | Ophiocoma wendtii |
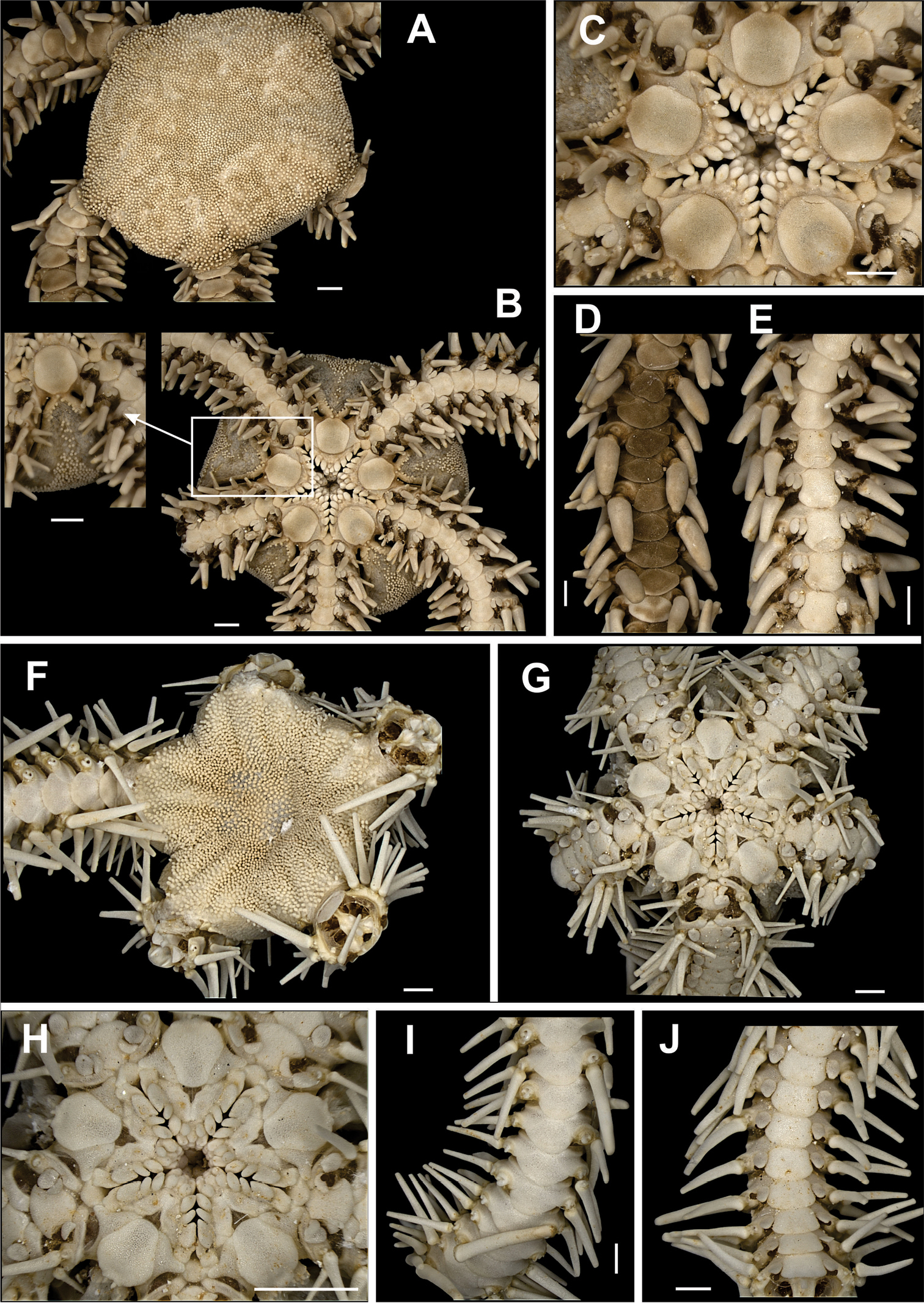
Disk pentagonal (dd = 5.67 to 12.16 mm). Covered by thick and naked tegument (Fig. 2a). Radial shields enlarged along internal margin. Marginal interradius with a row of 8 to 10 large and overlapping scales (Fig. 2a). Bursal slits short and narrow (Fig. 2b). Oral shields triangular to circular, distal margin rounded. Adoral shields narrow and enlarged laterally. Three enlarged oral papillae on each side of jaw angle, distal free end totally dentate, the two proximal papillae being longer and wider than distal papilla (Fig. 2c). Dorsal arm plate long, narrow, fragmented into two (Fig. 2d). Ventral arm plates small, pentagonal, with a small notch on the distal margins. Four to five small, compressed, arm spines, with denticles on tips. Without tentacle scale (Fig. 2e).
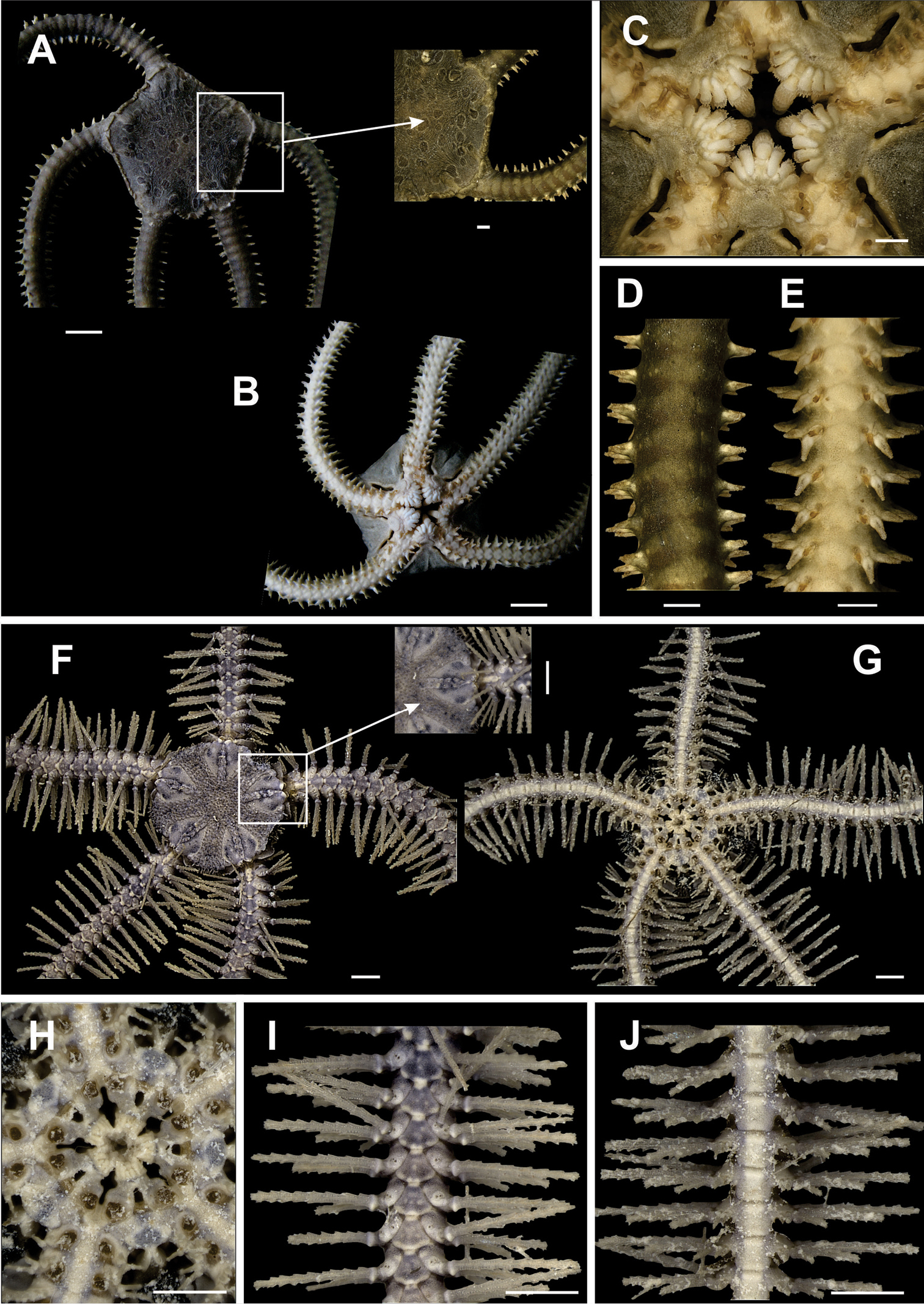
Species of the families Ophiomyxidae (A–E) and Ophiotrichidae (F–J). Ophiomyxa flaccida. A dorsal view, in detail the marginal interradius with a row of large scales B ventral view C jaw D dorsal view of the arm E ventral view of the arm. Ophiothrix (Ophiothrix) angulata F dorsal view, in detail the radial shields G ventral view H jaw I dorsal view of the arms J ventral view of the arms. Scale bar = 1 mm.
Bermuda, the islands off southern Florida, the Bahamas, the Antilles, Mexican Caribbean, Belize, Guatemala, Honduras, Panama, islands off Caribbean Colombia, Venezuela, and Brazil (
According to
Disk circular to pentagonal (dd = 4.26 to 9.82 mm). Covered by large, imbricating scales, surrounded by smaller scales of different sizes and irregular shapes (Fig. 3a). Primary plates conspicuous, central primary plate rounded. Interradius with three rows of large scales. Radial shields triangular, separated distally by three large scales disposed in a triangle, and proximally by a large scale (Fig. 3a). Ventral interradius covered by imbricating scales, slightly smaller and narrower than dorsal scales (Fig. 3b). Bursal slits long and narrow. Oral shields pentagonal, elongate, distal margin convex. Adoral shields broad, enlarged laterally and contiguous along internal median line of jaw. Four to five oral papillae on each side of jaw angle, the three proximal of which are pointed and subequal, the penultimate one is longest and broadest (Fig. 3c). Dorsal arm plate wider than long. Accessory dorsal arm plate reduced and restricted to the first arm segments (Fig. 3f). Ventral arm plate on first segments as large as long, on last segments slightly broader than long, tending to become pentagonal in shape, with lateral margins concave and distal margin rounded (Fig. 3e). Three or four arm spines short and conical, blunt, the two dorsal ones smaller. Tentacle pore large. Two large tentacle scales, the outer one slightly broader than the inner one (Fig. 3e).
Species of the family Ophiolepididae. Ophiolepis impressa A dorsal view B ventral view C jaw D detail of the radial shield E ventral view of the arms F dorsal view of the arms. Ophiolepis paucispina G dorsal view H ventral view I jaw J dorsal view of the arms (adp: accessory dorsal arm plate) L ventral view of the arms. Scale bar = 1 mm.
Bermuda, the Bahamas, the islands off southern Florida, Texas, the Antilles, Mexican Caribbean, Belize, Honduras, Costa Rica, Panama, Colombia, Venezuela, and Brazil (
According to
Disk circular (dd = 2.74 to 3.55 mm). Covered by large scales, and surrounded by smaller ones of similar size. Central primary plate circular, well defined, surrounded by five small primary plates, and intercalated by two smaller scales (Fig. 3g). Radial shield triangular, separated distally by three large scales disposed in a triangle. Ventral interradius covered by scales, slightly smaller and narrower than dorsal scales (Fig. 3h). Bursal slits long and narrow. Oral shields pentagonal, elongate, distal margin convex (Fig. 3i). Adoral shields broad, enlarged laterally. Four oral papillae on each side of jaw angle, the three proximal of which are pointed and subequal, the last one being longest and broadest (Fig. 3i). Dorsal arm plate fan-shaped (Fig. 3j). Arm segments with accessory dorsal plate, except near the tip (Fig. 3j). Ventral arm plate pentagonal, with lateral margins concave and distal margin rounded. Two tentacle scales oval, outher one larger. Two arm spines small (Fig. 3l).
Bermuda, the Bahamas, Florida, Caribbean Sea, Panama, Colombia, Brazil, and off Africa (
This species is known from shallow, sandy reef flats, mangrove, lagoonal, and seagrass environments, under coral rubble on sand, in calcareous algae such as Halimeda, and among plant debris (
http://species-id.net/wiki/Amphiodia_planispina
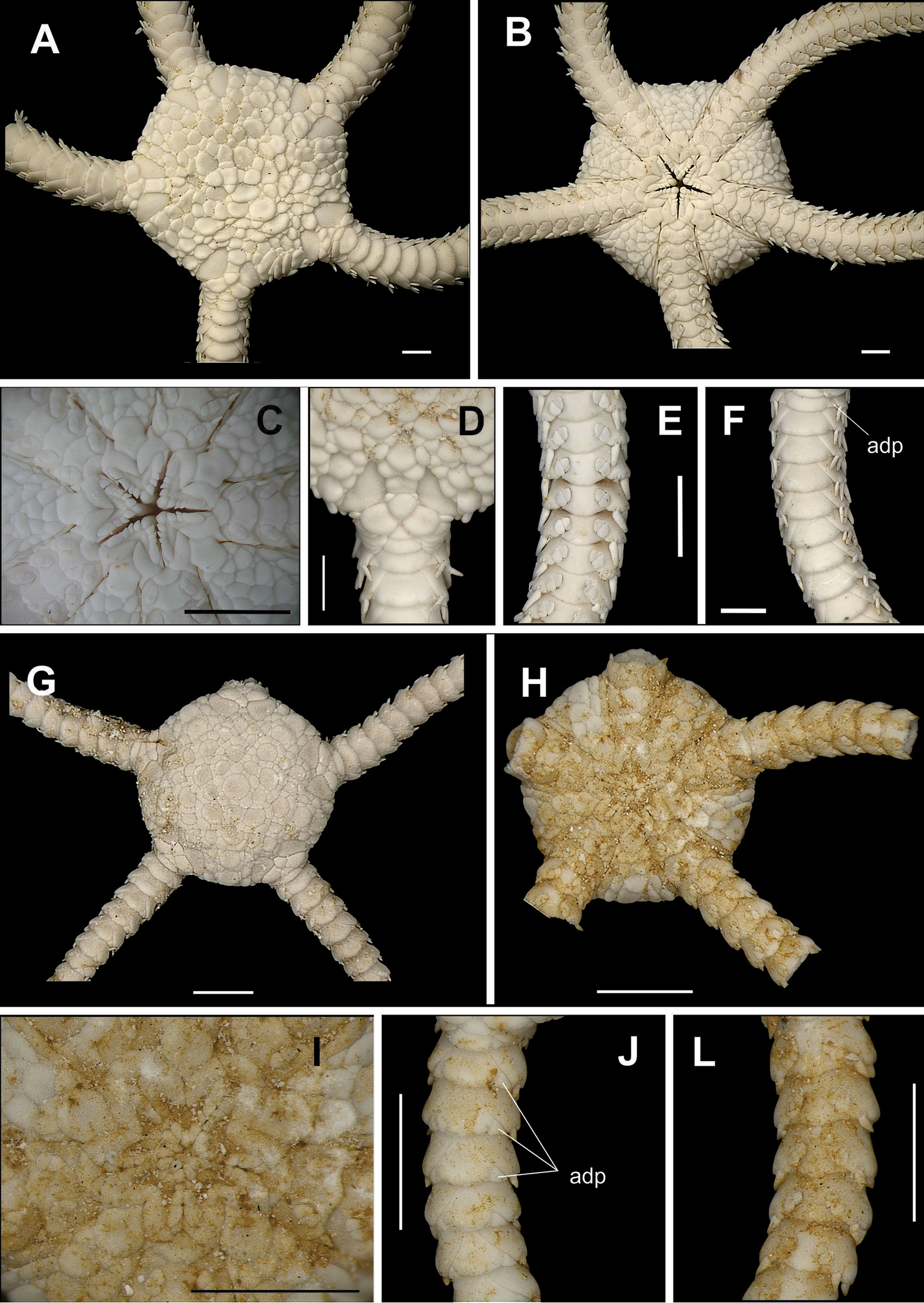
Figure 4a–eDisk circular (dd= 4.46 to 5.80 mm). Covered by numerous small and imbricating scales (Fig. 4a). Radial shields slightly longer than wide, with external margin convex and internal margin straight, united except at proximal ends, where one or two small triangular and elongate scales separate them (Fig. 4a). Ventral interradius covered by scales slightly smaller than dorsal scales (Fig. 4b). Bursal slits narrow. Oral shields arrow-shaped (Fig. 4c). Madreporite with one or two pores at distal margin. Adoral shields narrow and enlarged laterally. Two oral papillae at each side of jaw angle, distal one longer and broader (Fig. 4c). Infradental papillae small. Dorsal arm plate broader than long, rectangular (Fig. 4d). Ventral arm plate pentagonal, wider than long, with a slight notch in distal margin. Three arm spines enlarged, compressed and blunt, the ventral one being the longest (Fig. 4e). Two small, perpendicular, tentacle scales.

Species of the family Amphiuridae. Amphiodia planispina A dorsal view, detail of the radial shields B ventral view C jaw D dorsal view of the arms E ventral view of the arms. Amphiodia riisei F dorsal view, detail of the primary plates G ventral view H jaw I dorsal view of the arms J ventral view of the arms. Scale bar = 1 mm.
Florida, the islands off southern Florida, the Antilles, Panama Brazil, and off Mar del Plata, Argentina (
This species is known from bottoms of sand, mud, gravel and algae (
Disk circular (dd = 4.16 mm). Covered by numerous imbricating scales of irregular shapes (Fig. 4f). Primary plates very conspicuous (Fig. 4f). Central primary plate slightly pentagonal, surrounded by the radial primary plates. Radial shields enlarged distally, contiguous, except at proximal end, where there is a small, elongate, triangular scale (Fig. 4f). Ventral interradius covered by small, imbricating scales (Fig. 4g). Bursal slits long and broadened. Oral shields diamond-shaped (Fig. 4h). Adoral shields enlarged laterally. Two oral papillae on each side of jaw angle, the distal one a little larger than the proximal one (Fig. 4h). Infradental papillae rectangular and robust. Dorsal arm plate broader than long, rectangular, with the distal border rounded (Fig. 4i). Ventral arm plate pentagonal. Three arm spines slightly bigger than one arm segment, which is laterally flattened, with blunt tip (Fig. 4j). Two tentacle scales subequal and perpendicular.
Off Florida, the Antilles, possibly Panama and Puerto Rico, and Brazil (
This species is known from sand and mud bottoms (
Disk circular or pentagonal (dd = 1.90 to 2.70 mm). Covered by small and imbricating scales (Fig. 5a). Radial shields long, narrow, contiguous, separated proximally by a scale (Fig. 5a). Ventral interradius covered by slightly smaller scales, but similar to those on dorsal surface (Fig. 5b). Oral shields diamond-shaped (Fig. 5c). Adoral shields broadened laterally, almost united medially. Two elongated and broadened oral papillae on each side of jaw angle (Fig. 5c). Infradental papillae rectangular and robust. Dorsal arm plate broader than long, with proximal margin rounded and distal margin almost straight (Fig. 5d). Ventral arm plates pentagonal (Fig. 5e). Three to four elongate and blunt arm spines, the second or third with hyaline denticles on tip (Fig. 5f). Two perpendicular tentacle scales, inner scale slightly larger than outer.
Species of the family Amphiuridae. Amphipholis januarii A dorsal view, detail of the radial shields B ventral view C jaw D dorsal view of the arms E ventral view of the arms F detail of the arm spine. Amphipholis squamata G dorsal view H ventral view I jaw J dorsal view of the arms L ventral view of the arms. Scale bar = 1 mm.
South Carolina, Florida, the islands off southern Florida, Texas, the Antilles, Belize, Panama, and Brazil (
Species known from bottoms of mud, sand, shells (
http://species-id.net/wiki/Amphipholis_squamata
Figure 5g–lDisk circular (dd = 1.08 to 2.47 mm). Covered by large, irregular, and only slightly imbricating scales (Fig. 5g). Sometimes with the central primary plate evident. Radial scales slightly longer than broad, contiguous, separated proximally by a small scale, with outer margin rounded and inner margin straight. Ventral interradius covered by strongly imbricating scales, which are smaller than dorsal scales (Fig. 5h). Distinct line of demarcation between the scales of the dorsal and ventral surface. Bursal slits long and broad (Fig. 5h). Oral shields fan-shape, distal margin enlarged and convex, slightly longer than wide (Fig. 5i). Adoral shields large, united proximally. Two oral papillae on each side of jaw angle, distal long and opercular (Fig. 5i). A pair of infradental papillae. Dorsal arm plate broader than long, proximal margin rounded and distal margin straight (Fig. 5j). Ventral arm plate pentagonal, twice as long as wide. Three arm spines conical, erect, serrate at tip (Fig. 5l). Two tentacle scales small, narrow and elongated.
Traditionally considered cosmopolitan, except for the extreme polar regions (but see remarks). Western Atlantic from Canada, United States, Mexico, the Antilles, Belize, Costa Rica, Panama, Colombia, Brazil, Uruguay, and Santa Cruz Province, Argentina (
Viviparous polychromatic species, presenting simultaneous hermaphroditism (
Disk pentagonal (dd = 2.63 to 3.03 mm). Covered by imbricating scales of different sizes (Fig. 6a). Radial shields narrow, three times as long as wide, almost completely separated by two or three broad and elongated scales (Fig. 6a). Ventral interradius covered by scales similar to dorsal, but slightly smaller (Fig. 6b). Bursal slits narrow. Oral shields longer than wide, tending to be diamond-shaped (Fig. 6c). Adoral shields enlarged laterally. Two oral papillae on each side of jaw angle, distal spatuliform and proximal spiniform, the latter positioned more internally on jaw (Fig. 6c). Dorsal arm plate slightly wider than long, proximal angle acute and distal margin rounded, tending to be fan-shaped (Fig. 6d). Ventral arm plate rectangular and narrow (Fig. 6e). Four to five subequal arm spines, crown of denticles on tip. One small tentacle scale (Fig. 6e).

Species of the family Amphiuridae (A–I). Amphiura stimpsoni. A dorsal view, detail of the radial shields; B ventral view C jaw D dorsal view of the arms E ventral view of the arms. Ophiocnida scabriuscula F dorsal view, detail of the radial shields G ventral view H jaw I dorsal view of the arms J ventral view of the arms. Scale bar = 1 mm.
The Bahamas, the islands off southern Florida, west coast of Florida, Texas offshore reefs, the Antilles, Belize, islands off Caribbean Colombia, and Brazil (
Hermaphroditic and viviparous species. It lives in bottoms of mud, sand, calcareous algae (
http://species-id.net/wiki/Ophiocnida_scabriuscula
Figure 6e–iDisk circular with slight indentations in interradius (dd = 4.45 mm). Dorsal and ventral surfaces covered by numerous small spines, also in between the radial shields (Fig. 6e). Scales numerous on disk, imbricating and of different sizes, the largest surrounding the radial shields. Radial shields longer than wide, divergent and with two small accessory plates (Fig. 6e). Bursal slits long. Oral shields arrowhead-shape (Fig. 6g). Adoral shields enlarged laterally. Two oral papillae on each side of jaw angle, small, rounded, and slightly elongated (Fig. 6g). Dorsal arm plate rectangular, narrow, with distal margin rounded (Fig. 6h). Ventral arm plate pentagonal (Fig. 6i). Two small tentacle scales. Three arm spines slightly flattened, the dorsal one slightly longer and wider than the other two.
Bermuda, Florida, the islands off southern Florida, the Antilles, Mexican Caribbean, Panama, Colombia, Venezuela, and Brazil (
This littoral species has a limited bathymetric distribution (
http://species-id.net/wiki/Ophiophragmus_brachyactis
Figure 7a–eDisk circular (dd = 4.45 mm). Covered by imbricating scales of irregular shape (Fig. 7a). Margin of interradial field with 12-13 small, enlarged, blunt spines that decrease in size in the direction of the radial shields (Fig. 7a). Radial shields slightly longer than wide, united, except at proximal end, where a small triangular scale occurs between the pair of radial shields. Ventral interradius covered by scales similar to dorsal scales (Fig. 7b). Oral shields arrow-shaped (Fig. 7c). Adoral shields enlarged laterally, contiguous in proximal region. Two papillae on each side of jaw angle, the distal one longer and wider (Fig. 7c). Infradental papillae robust, and rectangular. Dorsal arm plate wider than long, rectangular, with rounded borders (Fig. 7d). Ventral arm plate pentagonal, with small notch on distal margin (Fig. 7e). Two small, perpendicular, tentacle scales, inner scale slightly longer than outer scale. Three small, compressed, blunt, arm spines, the ventral one largest (Fig. 7e).

Species of the family Amphiuridae. Ophiophragmus brachyactis A dorsal view, detail of the fence of papillae B ventral view C jaw D dorsal view of the arms E ventral view of the arms. Ophiostigma isocanthum F dorsal view G ventral view H detail of the radial shields I jaw J dorsal view of the arms L ventral view of the arms. Scale bar = 1 mm.
Florida and Dry Tortugas, the Antilles, Gulf of Mexico, and Brazil (
Species known from bottoms with sand, in which bryozoans predominate (
Disk circular to pentagonal (dd = 1.58 to 3.31 mm). Covered by small blunt tubercles (Fig. 7f). Some large and blunt tubercles distributed in the interradius, usually near the radial shields. Radial shields small (Fig. 7f, h). Ventral interradius covered by short and blunt tubercles similar to the dorsal ones (Fig. 7g). Bursal slits narrow and long. Oral shields fan-shape (Fig. 7i). Adoral shields united proximally and almost touching the adoral shield of the neighbouring jaw along median arm line (Fig. 7i). Two oral papillae on each side of jaw angle, distal operculate, closing oral slit (Fig. 7i). Dorsal arm plate with proximal margin rounded and distal margin straight (Fig. 7j). Ventral arm plate pentagonal, long. Three conical arm spines (Fig. 7l). Two elongate tentacle scale (Fig. 7l).
Bermuda, North Carolina to Florida and the island off southern Florida, Texas offshore reefs, the Bahamas, the Antilles, Mexican Caribbean, Panama, Colombia and islands off Caribbean, Colombia, Venezuela, and Brazil (
This species rarely exceeds 7 mm in disk diameter (
http://species-id.net/wiki/Ophiothrix_angulata
Figure 2f–j, 14cDisk circular (dd = 0.63 to 4.79 mm). Covered by small, hyaline bifid or trifid spines, also on the radial shields (Fig. 2f). Radial shields longer than wide, separated by a row of scales (Fig. 2f). Ventral interradius covered by spines similar to dorsal (Fig. 2g). Bursal slits short and wide. Oral shields enlarged laterally, triangular, with distal margin (Fig. 2h). Adoral shields united proximally. No oral papillae, but jaws bear terminal clump of dental papillae (Fig. 2h). Dorsal arm plate fan-shaped (Fig. 2i). Ventral arm plate slightly longer than wide, hexagonal, with distal margin long and slightly concave (Fig. 2j). Nine long arm spines, vitreous and denticulate, the one but last smallest and the last modified into a hook. Single tentacle scale small.
Bermuda, North Carolina to Texas coast and offshore reefs, Dry Tortugas, the Bahamas, the Antilles, Mexican Caribbean, Honduras, Belize, Panama, islands off Caribbean, Colombia, Venezuela, Brazil, Uruguay, and off La Plata river, Argentina (
Associated with seaweeds, such as the brown alga Sargassum spp. (
Five arms. Disk circular to pentagonal (dd = 2.48 to 7.62 mm). Covered by imbricating scales of irregular sizes. Scales at center of disk smaller, the largest on interradial field and mainly on lateral margins of radial shields (Fig. 8a). Sparsely distributed small spines on aboral region of disk. Radial shields long and separated over almost full length by three long scales (Fig. 8f). Ventral interradius covered by small spines on small and imbricating scales (Fig. 8b). Bursal slits long and wide. Oral shields long, diamond-shaped, proximally enlarged and distally narrow (Fig. 8c). Adoral shields large, laterally wide. Two or three small, spatulate, oral papillae, distal papilla broader and proximal papilla slightly curved towards the interior of the mouth (Fig. 8c). Dorsal arm plate wider than long, rectangular, with proximal margin rounded (Fig. 8d). Ventral arm plate hexagonal (Fig. 8e). Five or seven serrated arm spines, with a crown of denticles at tip. First dorsal spine small, second largest, and remaining decrease in size in the ventral direction. Single tentacle scale spatulate (Fig. 8e).

Species of the family Ophiactidae. Ophiactis quinqueradia A dorsal view B ventral view C jaw D dorsal view of the arms E ventral view of the arms F detail of the radial shields. Ophiactis savignyi G dorsal view, detail of the radial shields H ventral view I jaw J dorsal view of the arms L ventral view of the arms. Scale bar = 1 mm.
The Bahamas, the islands off southern Florida, off Mississippi, Texas offshore reefs, the Antilles, Mexican Caribbean, Cuba, Belize, Panama, and Brazil (
This species lives on bottoms of mud, sand, gravel or corals, being very common in sponges (
http://species-id.net/wiki/Ophiactis_savignyi
Figure 8g–lSix arms. Disk circular (dd = 1.3 mm). Covered by numerous imbricating scales. Some small spines on scales at margin of disk (Fig. 8g). Radial shields large, triangular, contiguous (Fig. 8g). Ventral interradus with some spines on the scales (Fig. 8h). Bursal slit enlarged. Oral shields oval (Fig. 8i). Adoral shields widened laterally. One oral papilla on each side of jaw angle (Fig. 8i). Apical papilla well developed. Dorsal arm plate wider than long, sometimes subdivided into two plates (Fig. 8j). Ventral arm plate octogonal. Six arm spines, with denticles along margin and at tip (Fig. 8l). Single tentacle scale semi-elliptical (Fig. 8l).
Cosmopolitan, in warm waters throughout the western Indo-Pacific, eastern Pacific, including Malpelo Island off western coast of Colombia, and both sides of the Atlantic, including Ascension island in the South Atlantic. Western Atlantic from South Carolina, Bermuda, Mexican Caribbean, Honduras, and Brazil (
Species found in all reef zones, seagrass beds, mangroves, and in contaminated communities (
Diskcircular to pentagonal (dd = 1.94 to 6.59 mm). Covered by numerous small and imbricating scales (Fig. 9a). Radial shields small, triangular, elongated and largely separated (Fig. 9a). Aboral surface of disk finely reticulated by fine brownish lines (Fig. 9a). Bursal slits large and with genital papillae (Fig. 9b). Oral shields diamond-shape, longer than wide (Fig. 9c). Adoral shields distally flaring. Three to five oral papillae on each side of jaw angle (Fig. 9c). Distal oral papilla slightly larger, other papillae diminishing progressively in size towards the mouth. Two to three apical papillae. Dorsal arm plates as wide as long, with rounded borders (Fig. 9d). Accessory dorsal arm plates well developed, not touching the neighbouring dorsal plates. Three slightly flattened spines on lateral arm plates (Fig. 9e). Single large, rounded, tentacle scale. Dark brown band (same colour as disk reticulation) along one arm segment, alternated by 3-6 light bands (Fig. 9d).
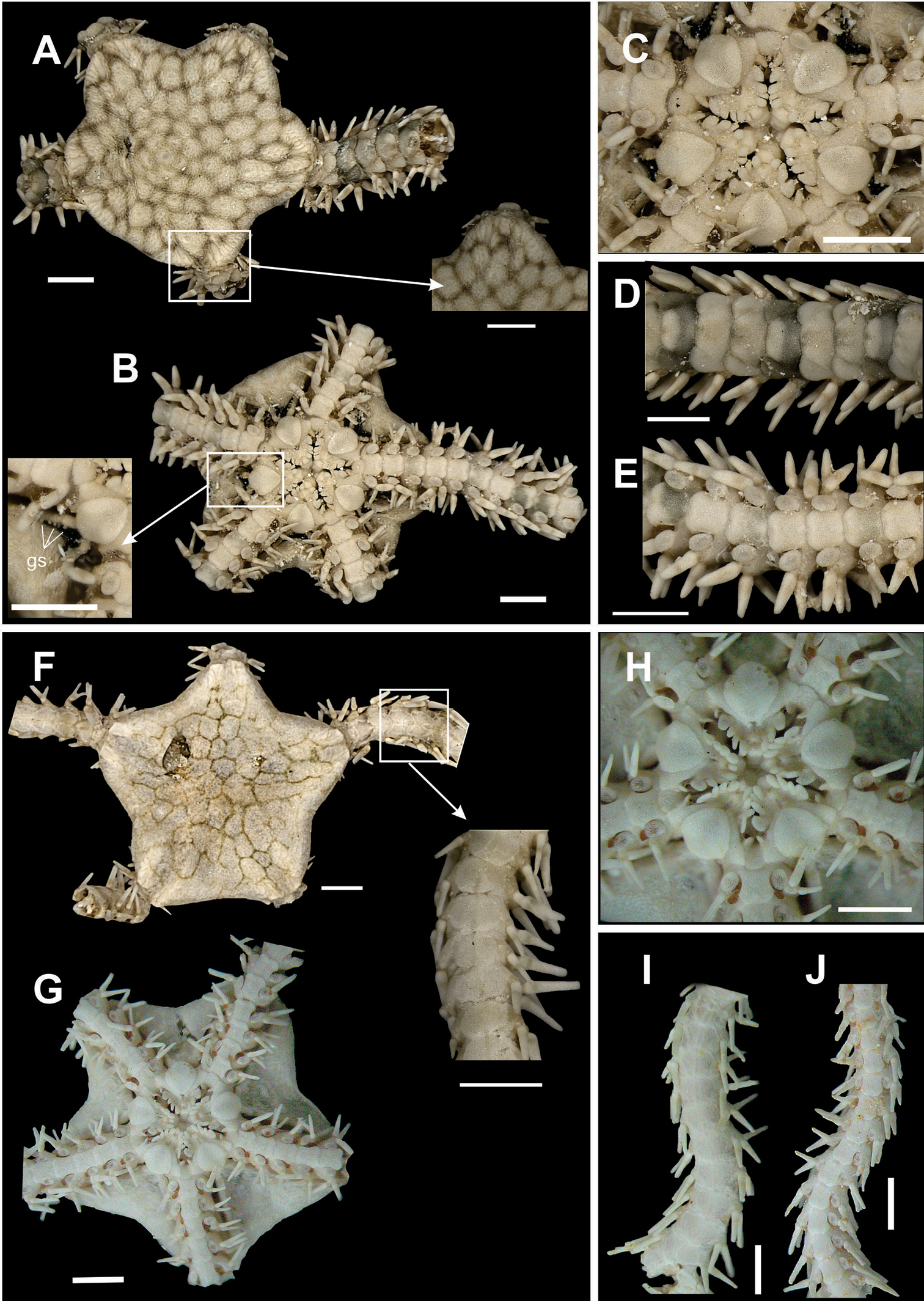
Species of the family Ophionereididae. Ophionereis reticulata A dorsal view, detail of the radial shields B ventral view, detail of the genital scale (gs) C jaw D dorsal view of the arms E ventral view of the arms. Ophionereis squamulosa F dorsal view, detail of the accessory dorsal arm plate (adp) G ventral view H jaw I dorsal view of the arms J ventral view of the arms. Scale bar = 1 mm.
Bermuda, North Carolina, South Carolina offshore reefs, Florida and the islands off southern Florida, the Bahamas, Texas offshore reefs, the Antilles, Mexican Caribbean, Belize, Honduras, Costa Rica, Panama, coast and islands off Caribbean Colombia, Venezuela, and Brazil (
Occurs in moderate densities in seagrass beds, and on sand with pebbles (
Disk circular (dd = 2.46 to 5.33 mm). Covered by numerous small and imbricating scales (Fig. 9f). Radial shields small, narrow and widely separated. Dark blotches on aboral surface of disk. Disk scales extending onto first brachial segment. Ventral interradius covered by scales similar to dorsal ones (Fig. 9g). Oral shields oval (Fig. 9h). Adoral shields enlarged laterally. Four oral papillae on each side of jaw angle (Fig. 9h). One pair of apical papillae. Dorsal arm plates longer than wide, proximal margin enlarged and distal margin narrow. Accessory dorsal arm plates well developed (Fig. 9f, i). Ventral arm plates slightly longer than wide (Fig. 9j). Three arm spines slightly flattened, with blunt tip (Fig. 9i, j). Single large tentacle scale.
The Bahamas, the islands off southern Florida, the Antilles, Mexican Caribbean, Belize, Panama, and Brazil (
Known from bottoms of sand, gravel, dead shells (
http://species-id.net/wiki/Ophionereis_dolabriformis
Figure 10a–eDisk circular to pentaradial (dd = 3.20 to 5.09 mm). Covered dorsally by imbricating scales of different sizes (Fig. 10a). Radial shields small, narrow, triangular (Fig. 10a). Pentaradial olive-green colour pattern on dorsal surface of disk (Fig. 10a). Bursal slits long and without genital papillae (Fig. 10b). Oral shields arrowhead-shaped, partially covering the adoral shields (Fig. 10c). Adoral shields united proximally and enlarged distally. Four oral papillae on each side of jaw angle (Fig. 10c). Dorsal arm plate longer than wide, distal region strongly convex (Fig. 10d). Ventral arm plate rectangular, lateral margins concave, distal margin enlarged and slightly convex (Fig. 10e). Single large, oval, tentacle scale. Three elongate arm spines, needle-shaped (Fig. 10d, e), fully denticulate. Olive-green stripe on 1 to 2 ½ dorsal arm segments.
Species of the family Ophionereididae. Ophionereis dolabriformis A dorsal view, detail of the radial shields B ventral view, detail of the genital scale (gs) C jaw D dorsal view of the arms E vental view of the arms. Ophionereis olivacea F dorsal view, detail of the radial shields F ventral view, detail of the accessory dorsal arm plate (adp) H jaw I dorsal view of the arms J ventral view of the arms. Scale bar = 1 mm.
Caribbean coast of Mexico and Colombia, Venezuela (
Ophionereis dolabriformis seems to be a rare species, with a high tolerance of river influence (
Disk usually pentagonal (dd = 3.15 to 3.75 mm). Covered by small and imbricating scales (Fig. 10f). Radial shields small, triangular, narrow, elongated and broadly separated (Fig. 10f). Largest scales surrounding and in between radial shields. Ventral interradius covered by imbricating scales similar to dorsal ones (Fig. 10g). Oral shields tending to heart-shape (Fig. 10h). Adoral shields enlarged laterally. Four oral papillae on each side of jaw angle (Fig. 10h). Dorsal arm plate slightly longer than wide (Fig. 10i). Accessory dorsal arm plate small, sometimes with several overlapping plates (Fig. 10f, i). Ventral arm plate slightly longer than wide (Fig. 10j). Three arm spines slightly larger than arm segment. Single large tentacle scale.
The Florida Keys, the Antilles, the Mexican Caribbean, Belize, Panama, the Colombian Caribbean (
Known from bottoms of quartz sand, corals, coral fragments, mangroves, and phytal communities (
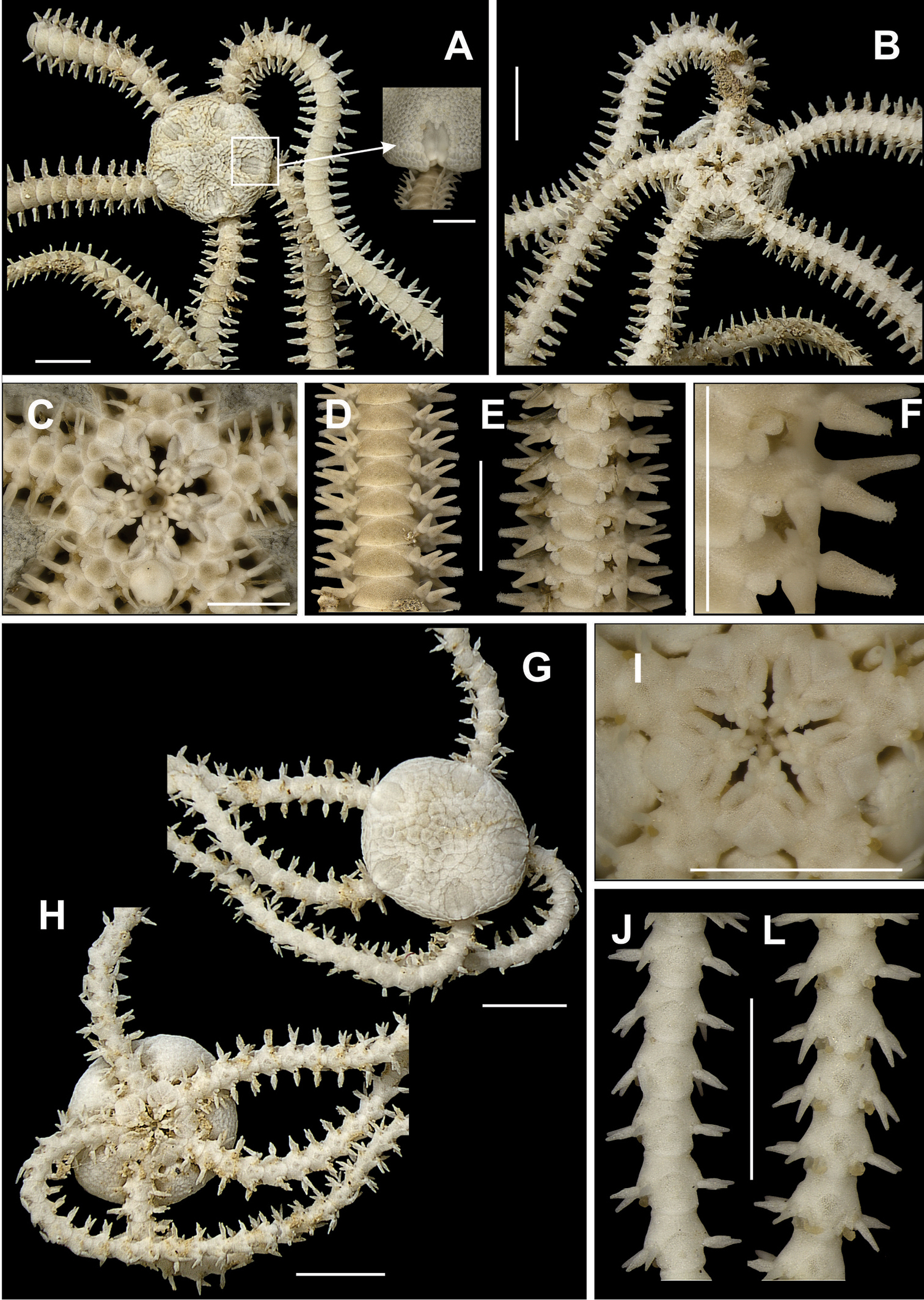
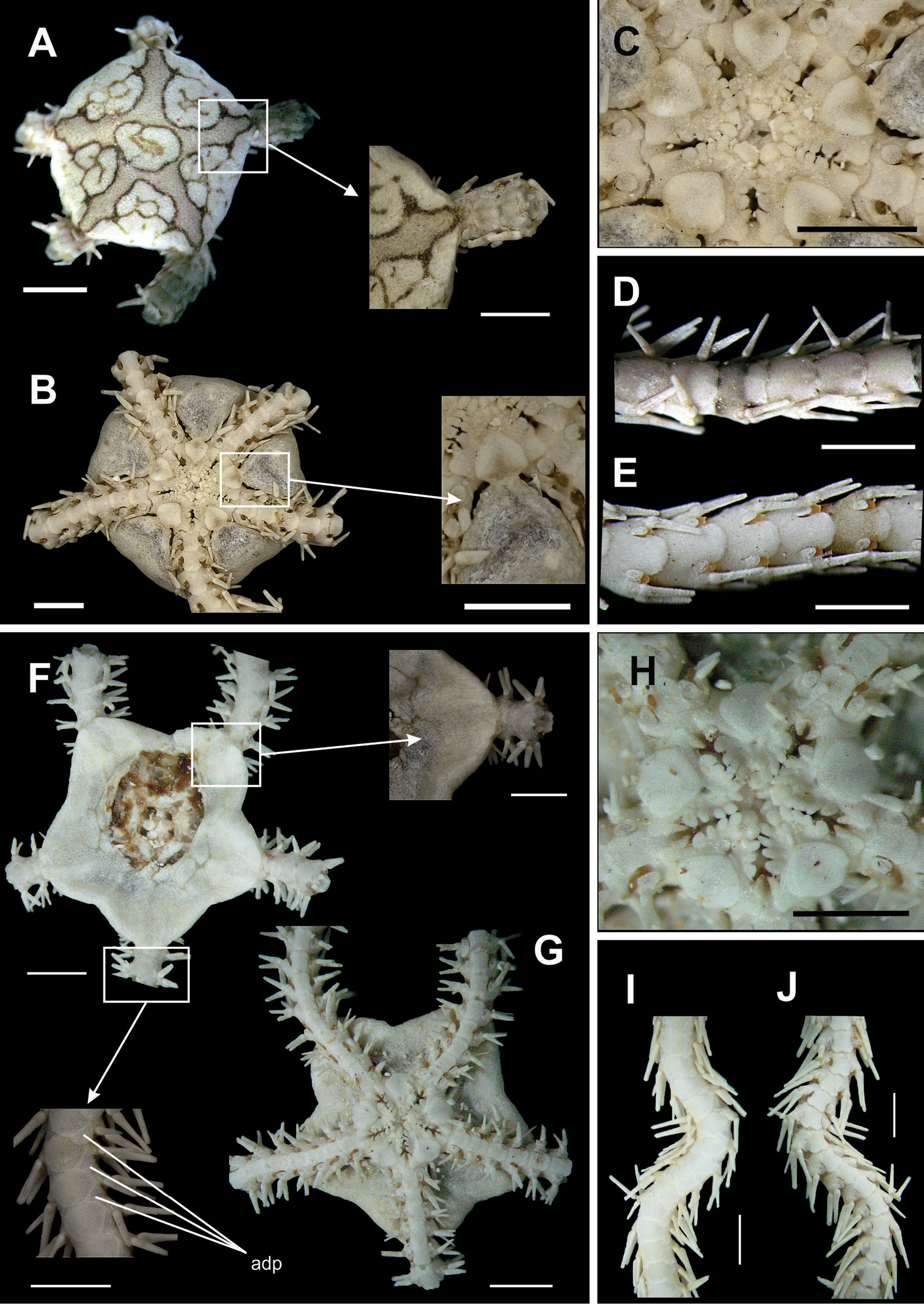
Disk circular to pentagonal (dd = 3.06 to 16.68 mm). Uniformly covered by small granules (Fig. 11a), which are smaller in central region than in marginal region. These granules occupy a v-shaped area on the ventral interradius (Fig. 11b). In the areas without granules there are large and imbricating scales. Bursal slits enlarged, with well developed genital scales in margins (Fig. 11b). Oral shields large and rectangular, proximal margin slightly rounded (Fig. 11c). Adoral shields small, almost totally covered by oral shield. Four oral papillae on each side of jaw angle (Fig. 11c). Two proximal papillae slightly cylindrical and subequal, two distal papillae longer and broader. Cluster of well developed dental papillae on apex of jaw (Fig. 11c). Dorsal arm plate longer than wide, fan-shaped (Fig. 11d). Ventral arm plate longer than wide, octogonal, with distal margin slightly convex (Fig. 11e). Two tentacle scales, internal one slightly larger than external one. Three or four arm spines alternating on arm segments. Dorsal spine longer and broader (bottle-shaped) (Fig. 11d), median ones of equal size and ventral one smaller and slightly flattened.
Species of the family Ophiocomidae. Ophiocoma echinata A dorsal view B ventral view, detail of the genital scale (gs) C jaw D dorsal view of the arms E ventral view of the arms. Ophiocoma wendtii F dorsal view G ventral view H jaw I dorsal view of the arms J ventral view of the arms. Scale bar = 1 mm.
Bermuda, Florida and the islands off southern Florida, the Bahamas, the Antilles, the Mexican Caribbean, Belize, Nicaragua, Guatemala, Honduras, Costa Rica, Panama, Colombia, Venezuela, and Brazil (
This species has diurnal habits. It lives in reef zones, seagrass beds, mangroves, being particularly abundant under rocks (
Disk pentagonal with small notches on interradius (dd = 2.71 to 15.07 mm) (Fig. 11e). Covered by small, imbricating scales, totally covered by small granules that extend over the first three dorsal arm segments (Fig. 11e). Ventral interradius covered by granules, that form a V-shaped area (Fig. 11f). Bursal slits long and enlarged. Oral shields triangular (Fig. 11g). Adoral shields distally flaring, not touching medially. Four oral papillae on each side of jaw angle (Fig. 11g), the one but last largest and partially covering last papilla. A cluster of papillae on jaw apex (Fig. 11g). Dorsal arm plate wider than long, fan-shaped (Fig. 11h). Ventral arm plate pentagonal, with distal margin rounded (Fig. 11i). Single large tentacle scale, but the six first arm segments may present two scales. Three or four long, pointed, arm spines, with blunt tip, the dorsal one biggest, four or five arm segments long (Fig. 11h). They decrease in size in the direction of the ventral spine, which is slightly curved. Arm segments with three or four alternating spines.
Bermuda, the Bahamas, the islands off southern Florida, Texas offshore reefs, the Antilles, Mexican Caribbean, Belize, Honduras, Costa Rica, Panama, islands off Caribbean Colombia, Venezuela, and Brazil (
Lives in bottoms of coral, dead shells (
Disk circular (dd = 1.30 to 6.90 mm). Covered by imbricating scales of different sizes, largest between radial shields and on interradial margin of disk (Fig. 12a). Radial shields narrow, long, broadly separated, distal end slightly broader, distinct from remaining disk because of their white coloration (Fig. 12a). Numerous olive-green patches on the dorsal and ventral sides of the disk (Fig. 12a). Ventral interradius with scales similar to dorsal disk surface (Fig. 12b). Bursal slits broad and elongated. Oral shields large, diamond-shaped, laterally broadened, some with a dark spot on distal margin. Two spatulate oral papillae (Fig. 12c), borders slightly denteate, on each margin of jaw, the outer one slightly longer. A cluster of dental papillae on apex of jaw (Fig. 12c). Dorsal arm plate slightly wider than long, distal border slightly wider than anterior border (Fig. 12d). Ventral arm plate longer than wide, pentagonal, posterior margin concave (Fig. 12e). Two tentacle scales (Fig. 12b, e), outer smaller and inner longer, flattened and overreaching median ventral plane of arm. Tentacle pore large. Four to six arm spines, ventral one longest and slightly curved (Fig. 12d, e). Remaining spines decrease in size ventralwards, with small denticles at apex. Two close brown stripes on lateral arm plate, and a lighter median band on the dorsal arm plate (Fig. 12a, d). Ventral surface sometimes with two brown stripes close to insertion of spines.
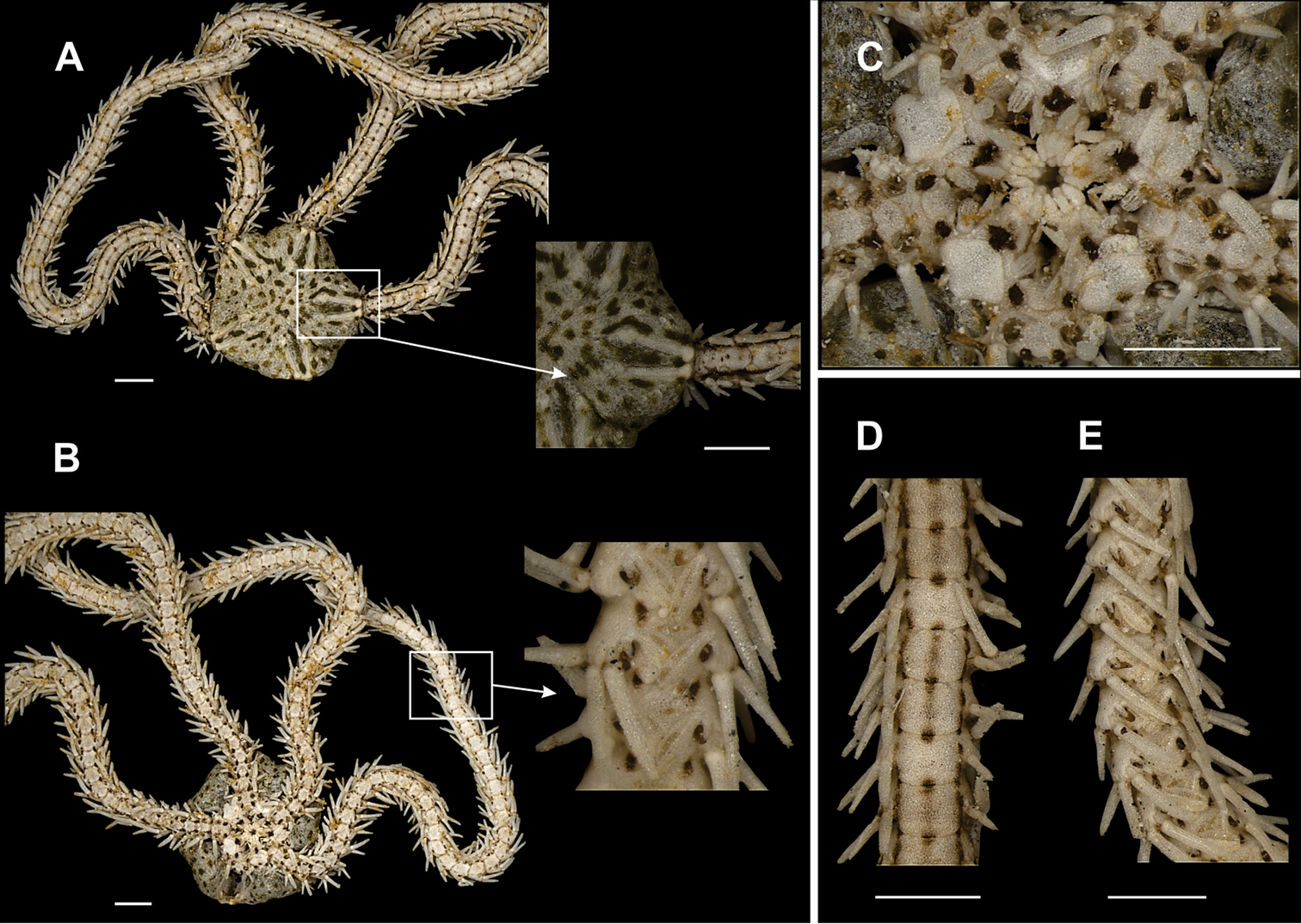
Specimen of the family Ophiocomidae (a–e). Ophiopsila hartmeyeri A dorsal view, detail of the radial shields B ventral view, detail of the tentacle scale C jaw D dorsal view of the arms E ventral view of the arms. Scale bar = 1 mm.
Florida Keys, the Mexican Caribbean, the Antilles, Costa Rica, Colombia, Venezuela, and Brazil (
Typically, this is a coralline bottom species (
Disk circular to pentagonal (dd = 4.18 to 7.89 mm), covered by small granules (Fig. 13a). Radial shields oval and covered with granules (Fig. 13a). Ventral interradius covered by similar granules. Four short bursal slits (Fig. 13b). Oral shields oval (Fig. 13c). Adoral shields broadened laterally, not covered by granules (Fig. 13c). Seven or eight oral papillae on each side of jaw angle (Fig. 13c), the three proximal ones small and elongated the last one narrow and partially covered by the previous papilla. Single apical papillae. Dorsal arm plate longer than wide, with distal margin rounded (Fig. 13e). Seven small, compressed, arm spines, the ventralmost one partially covered by outer tentacle scale. Two tentacle scales, the inner one longest (Fig. 13d).
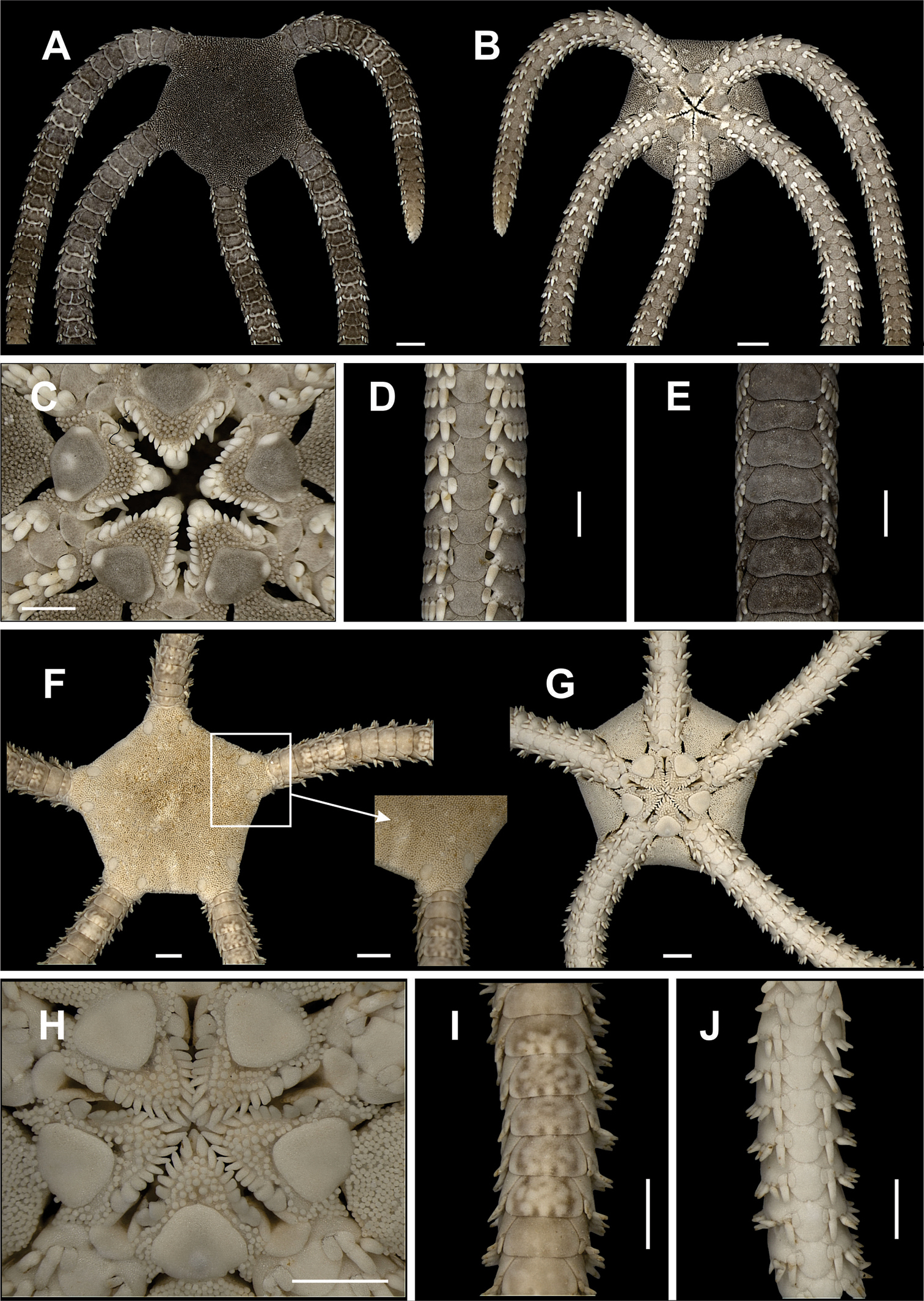
Species of the family Ophiodermatidae (a–d). Ophioderma appressa A dorsal view B ventral view C jaw D ventral view of the arms E ventral view of the arms. Ophioderma cinerea F dorsal view, detail of the radial shields G ventral view H jaw I dorsal view of the arms J ventral view of the arms. Scale bar = 1 mm.

Some ophiurans in their natural habitat. A Ophiomyxa flaccida B Ophiolepis impressa C Ophiothrix angulata D Ophiocoma echinata E Ophioderma cinerea F Ophioderma appressa. Photos by Thelma L. P. Dias.
Western Atlantic from Bermuda, South Carolina, the islands off southern Florida, Texas offshore reefs, the Bahamas, the Antilles, Mexican Caribbean, Belize, Honduras, Costa Rica, Panama, islands off Caribbean Colombia, Venezuela, and Brazil (
This cryptic species lives in reef environments, seagrass beds, on gravel and coral rubble. Usually found together with other ophiuroids, such as Ophioderma cinerea, Ophiocoma echinata, and Ophiocoma wendtii (
http://species-id.net/wiki/Ophioderma_cinerea
Figure 13f–j, 14eDisk circular to pentagonal (dd = 4.96 to 9.67 mm). Covered by small granules, except on radial shields (Fig. 13f). Radial shields oval. Ventral interradius covered by granules similar to dorsal ones (Fig. 13g). Four short bursal slits. Oral shields heart-shaped (Fig. 13h). Adoral shields small, laterally broadened, not covered by granules. Seven to nine oral papillae on each side of jaw angle (Fig. 13h), the three proximal ones small and elongate, the following ones becoming progressively wider, the last being elongate, narrow and partially covered by preceeding papilla. Single long and robust apical papilla. Dorsal arm plate wider than long (Fig. 13i). Ventral arm plate longer than large, with distal margin rounded (Fig. 13j). Seven to nine small and compressed arm spines, the ventral largest and partially covered by the outer tentacle scale. Two tentacle scales, the inner one long and narrow, the outer one small and subtriangular.
The Bahamas, the islands off southern Florida, the Antilles, Mexican Caribbean, Belize, Honduras, Costa Rica, Panama, coast and islands off Caribbean Colombia, Venezuela, and Brazil (
This is one of the most common and largest species in the genus, and differs from other Ophioderma from Brazil, such as Ophioderma appressa (Say, 1825), and Ophioderma januarii Lütken, 1856, by the following characteristics: 1. radial shields within granular covering; 2. dorsal arm plates partitioned. Tommasi (unpublished data) suggested that Ophioderma besnardi Tommasi, 1970 represents the young of Ophioderma cinerea Müller & Troschel, 1842 before the dorsal plates were divided. More detailed studies are currently being developed to elucidate the taxonomic status of these species. It lives in muddy bottoms, corals (
Of the 23 studied species of ophiuroids, 17 occur along practically the whole continental shelf of the State of Paraíba. Only five, Ophiactis savignyi, Ophiophragmus brachyactis, Ophiocnida scabriuscula, Amphipholis squamata, and Amphiodia riisei had punctual occurrence, being recorded from only one or two collection sites (Fig. 15, Tab. 1, Supplementary material). Ophiothrix angulata was the most common, occurring in 73% of the collection stations (Tab. 2, Fig. 16). The bathymetric range of three species, Ophiolepis impressa, Ophiocnida scabriuscula, and Ophiocoma echinata was extended. Ophionereis squamulosa was the most abundant species, representing 17.04% of the specimens studied, being more abundant in isobaths 20–24 m deep (Fig. 16). Figure 1 indicates the abundance of specimens in each collection point studied over the continental shelf of Paraíba state, in the isobaths studied.
Among the eight recorded families, Amphiuridae, Ophiactidae, Ophiomyxidae, and Ophiuridae had a patchy occurrence. On the other hand, Ophiocomidae, Ophiodermatidae, Ophionereididae, and Ophiotrichidae were well represented, being frequent over all the extent of the continental shelf (Fig. 17).
In general the species richness was constant along the different depth intervals, the highest species richness occurring between 26 and 30 m (n = 19 spp.) and the lowest species richness occurred between 31 and 35 m (n = 11 spp.) (Fig. 15, 16). Essentially the same pattern was observed for individual families, all families occurred with their greatest numbers at depths between 20 and 29 m (Tab. 2, Supplementary Material) (Fig. 17).
The taxa Ophiactidae and Ophiocomidae showed a similar bathymetric occurrence, being best represented in the intervals 20-29 m and 30–35 m (Fig. 17), and least represented in the interval 10–19 m (Fig. 17). Ophiodermatidae showed the same depth occurrence, but the number of specimens was similar at 10–19 m and 30–35 m (Fig. 16). Ophiomyxidae, Ophionereididae, Ophiotrichidae, and Ophiuridae formed another grouping, in which the interval with the smallest number of specimens was 30–35 m.
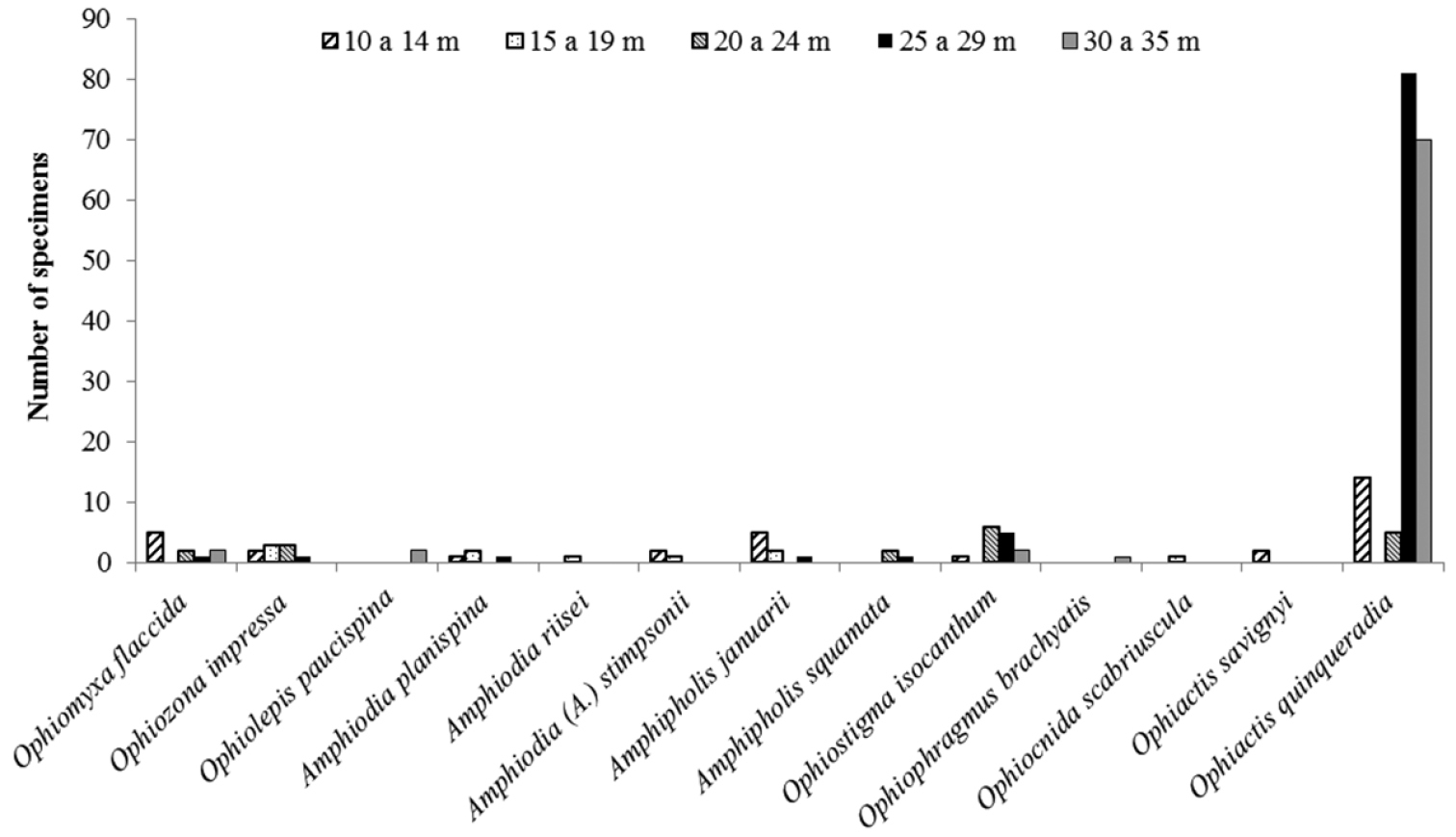
Number of individuals/species of the families Ophiomyxidae, Amphiuridae and Ophiactidae, according to different bathymetric ranges.
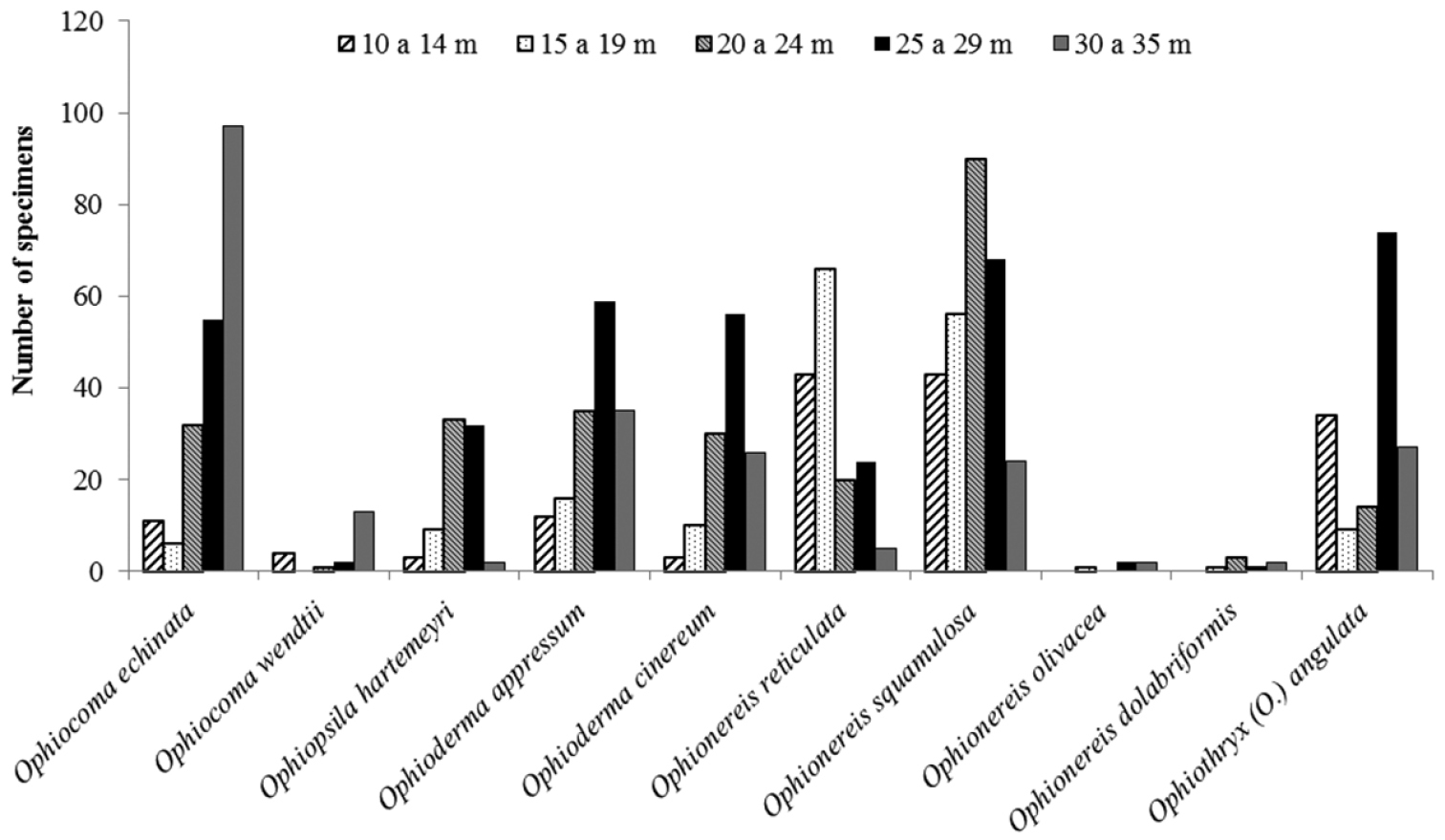
Number of individuals/species of the families Ophiocomidae, Ophiodermatidae, Ophionereididae, Ophiotrichidae and Ophiuridae, according to different bathymetric ranges.
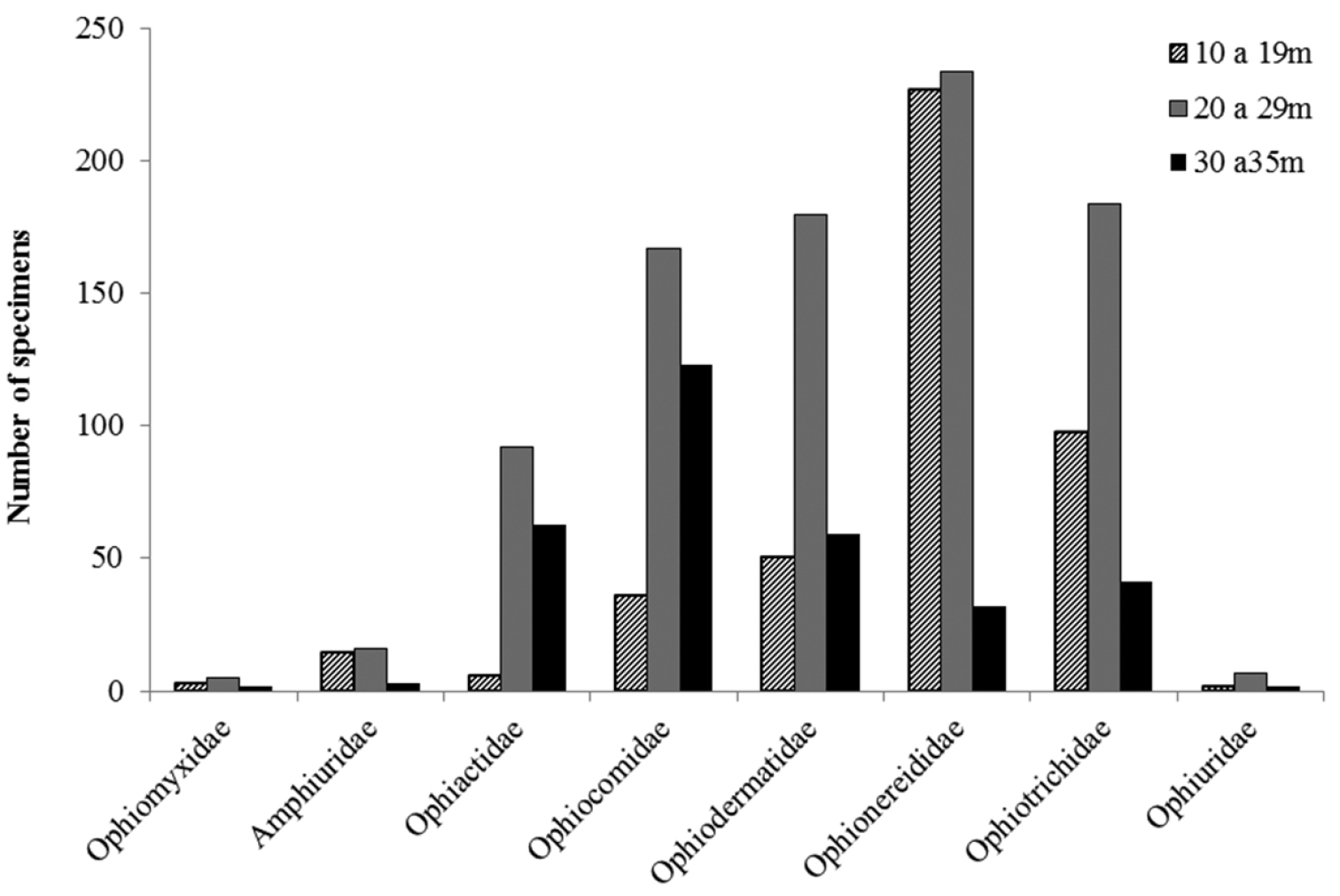
Number of individuals per family, according to different bathymetric ranges.
The ophiuroid fauna recorded for the continental shelf of the State of Paraíba is composed of species with a large bathymetric distribution, considering that most of them occur from shallow waters to depths greater than 50 m, as pointed out by
We recorded two supposedly cosmopolitan species, Amphipholis squamata and Ophiactis savignyi, and one amphiatlantic species, Ophionereis reticulata. The first two species are visually abundant in the intertidal zone of Paraíba coast, mainly associated with sponges and macroalgae, but were very scarce sampled on the continental shelf. Only two and one specimen of these species was sampled in this study. This may be due to scarcity in the biological material collected, which was composed mainly of calcareous unattached algae (rhodoliths), composed by are calcareous and rigid thallus. Another possibility relates to a possible low abundance of Ophiactis savignyi in deeper waters.
The occurrence of the fauna of ophiuroids on the continental platform of the State of Paraíba could be the result of the homogeneity of the bottom types along the sampled area (
In general the most diverse places in the Western Atlantic Ocean with respect to the ophiuran fauna are the Gulf of Mexico (182 spp.), and the Caribbean Sea (148 spp.) (
This suggests that, despite the existence of the barrier of the Amazon River, there is some connectivity between some Caribbean species and those found in the South Atlantic. Studying reef fishes,
Although it has long been postulated that diversity in marine species or communites may follow latitudinal gradients with diversity peaking at the equator and declining towards higher latitudes (
We are grateful to the Federal University of Paraíba, for providing the infrastructure enabling this research, to the National Science Foundation (CNPq) for the scholarships that contributed to this research. MLC is supported by a CNPq productivity research grant (Process number: 300198/2010-8). Special thanks are due to our Graduate Course (Programa de Pós-Graduação em Ciências Biológicas) through Project CT-Infra and Projeto Biota - Paraíba: Macrofauna de Praias com Substrato Consolidado da Zona de Entre-Marés ao Infralitoral, financed by Conselho Nacional de Desenvolvimento Científico e Tecnológico - CNPq, through Edital Universal (Process n° 484601/2007-5), with which the essential photographic equipment for the illustration of specimens was obtained. We also wish to express our sincere gratitude to two anonymous reviewers for their critical reading of the manuscript and constructive comments, and Carolina Nunes Liberal for help with photographs.
Collection data of voucher specimens (spec.) collected at Paraíba State continental shelf, Northeastern Brazil. (doi: 10.3897/zookeys.307.4673.app1) File format: Mircrosoft Word Document (doc).
Data on the size (measured by disk diameter), number of individuals and relative abundance of voucher specimens collected at Paraíba State continental shelf, Northeastern Brazil. SD = standard deviation. (doi: 10.3897/zookeys.307.4673.app2) File format: Mircrosoft Word Document (doc).