





(C) 2012 Wilfredo A. Matamoros. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
A new species of Profundulus, Profundulus kreiseri (Cyprinodontiformes: Profundulidae), is described from the Chamelecón and Ulúa Rivers in the northwestern Honduran highlands. Based on a phylogenetic analysis using cytochrome b and the presence of synapomorphic characters (dark humeral spot, a scaled preorbital region and between 32-34 vertebrae), this new species is placed in the subgenus Profundulus, which also includes Profundulus (Profundulus) oaxacae, Profundulus (Profundulus) punctatus and Profundulus (Profundulus) guatemalensis. Profundulus kreiseri can be distinguished from other members of the subgenus Profundulus by having less than half of its caudal fin densely scaled. Profundulus kreiseri can further be differentiated from Profundulus (Profundulus) oaxacae and Profundulus (Profundulus) punctatus by the absence of rows of dark spots on its flanks. The new species can further be differentiated from Profundulus (Profundulus) guatemalensis by the presence of fewer caudal- and pectoral-fin rays. The new species is distinguished from congeners of the profundulid subgenus Tlaloc (viz., Profundulus (Tlaloc) hildebrandi, Profundulus (Tlaloc) labialis, Profundulus (Tlaloc) candalarius and Profundulus (Tlaloc) portillorum) by having a scaled preorbital region and a dark humeral spot. Profundulus kreiseri and Profundulus portillorum are the only two species of Profundulus that are endemic to the region south of the Motagua River drainage in southern Guatemala and northwestern Honduras.
Central America, nuclear Middle America, Cyprinodontiformes, Kreiser’s Killifish, phylogeny, systematics
The genus Profundulus Hubbs, 1924, is a depauperate lineage of northern Central American and southern Mexican (Fig. 1) cyprinodontiforms that belong to Profundulidae. The family comprises a single genus, Profundulus, which has seven valid species (
Taxonomic relationships within Profundulidae were inferred by
Among neotropical members of Cyprinodontiformes, profundulids are the least studied. Although some progress has been made over the past two decades in advancing our understanding of the genetics (
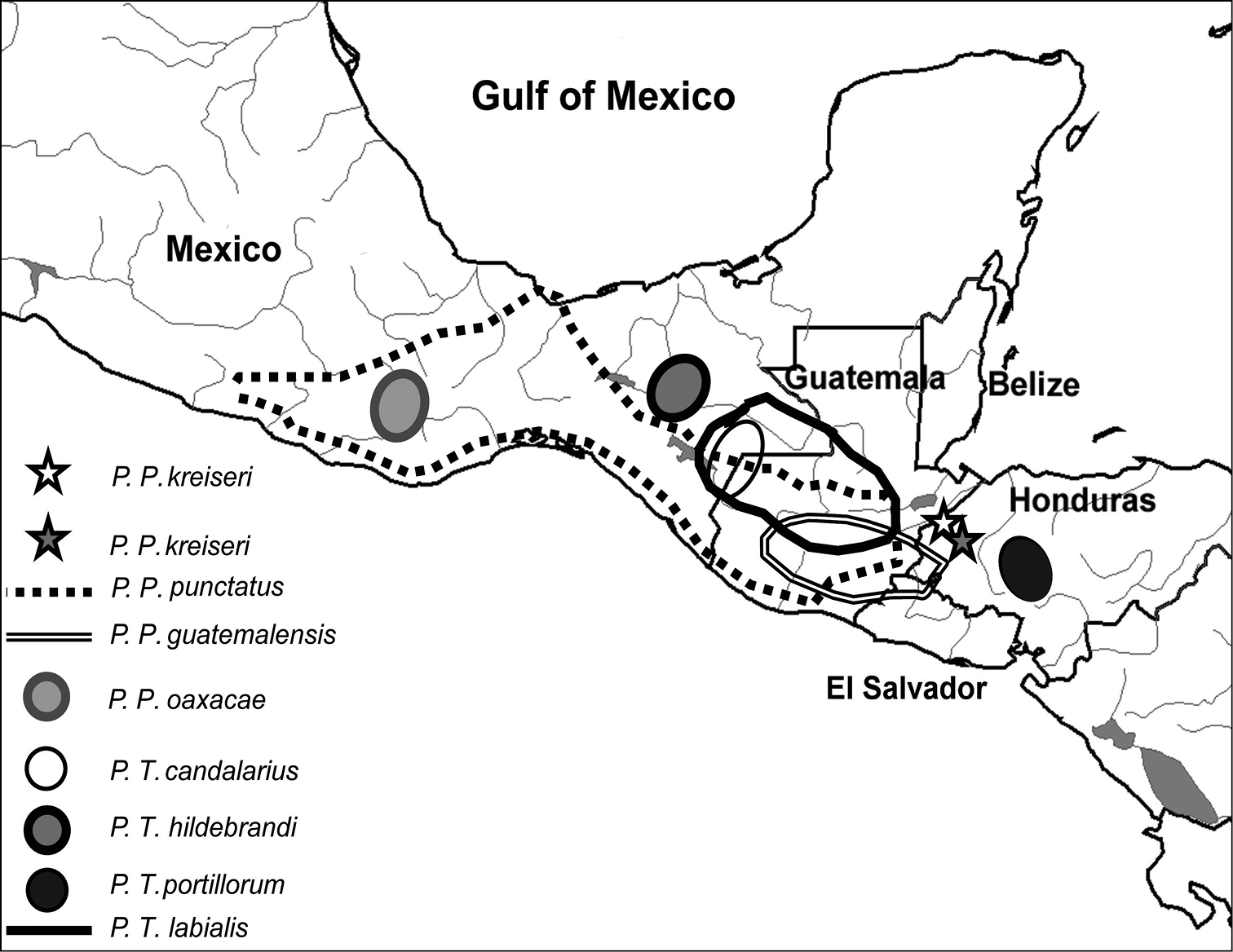
Map of Honduras and neighboring countries showing the distribution of all species of Profundulus. The type locality of Profundulus kreiseri (15.197667°N, 88.616°W) is represented by a white star, and a second known locality for the species (15.029520°N, 88.508°W) is represented by a dark star.
All specimens examined are housed at the University of Southern Mississippi Ichthyology Collection (USM), Louisiana State University Museum of Natural Science (LSUMZ), and University of Michigan Museum of Zoology (UMMZ). Measurements and counts were taken following
Genomic DNA was extracted from ethanol-preserved fin tissue using a DNeasy Tissue Kit (QIAGEN Inc.). We amplified the mitochondrial cytochrome b gene using the GluF and ThrR primers described by
Sequence divergence (uncorrected p-distance) was estimated in PAUP* (
GenBank accession numbers and locality information of data used in the molecular analysis. Gu= Guatemala, Me= Mexico, Ho= Honduras.
| Taxa | Accession No. | Locality | Source |
|---|---|---|---|
| Profundulus guatemalensis | AY155568 | Gu. Blanco River, San Miguel Duenas |
|
| Profundulus labialis | AY155567 | Gu. Jeronimo River, San Jeronimo |
|
| Profundulus punctatus | AY155566 | Me. Manialtepec Basin, San Juan Lachao |
|
| Profundulus candalarius | JQ254931 | Me. Chiapas, Grijalva River basin | this study |
| Profundulus hildebrandi | JQ254932 | Me. Chiapas, San Cristobal de las Casas | this study |
| Profundulus kreiseri n. sp. | JQ254935 | Ho. Chamelecón River | this study; USM 39024 |
| Profundulus kreiseri n. sp. | JQ254934 | Ho. Ulúa River | this study; USM 39028 |
| Profundulus oaxacae | JQ254933 | Me. Oaxaca, Verde River basin | this study |
| Profundulus portillorum | JQ254929 | Ho. Nacaome River | this study; USM 31628 |
| Profundulus portillorum | JQ254930 | Ho. Ulúa River | this study; USM 31597 |
| Allodontichthys hubbssi | AF510835 | Me. Jalisco, El Trampolin |
|
| Alloophorus robustus | AF510809 | Me. Michoacán, Uruapan |
|
The “genetypes” nomenclature was proposed by
The genetype terminology is not used here in a strict nomenclatural sense, as it is not formally accepted by the International Code for Zoological Nomenclature. However, we consider this terminology useful and expect increased application in the future. The present paper is one of the first uses of the genetype terminology.
Results Systematic Accounturn:lsid:zoobank.org:act:97D8525F-9BDF-48EF-A7C5-21AD7917558B
http://species-id.net/wiki/Profundulus_kreiseri
Figure 2Holotype: USM 39022, field number WAM09-28, Honduras, Department of Santa Barbara, Municipality of Macuelizo. Drainage: Chamelecón, System: Chamelecón. Locality: small creek that drains to the Chamelecón River, near the Chamelecón Hydroelectric Dam 15.197667°N, 88.616°W; Collectors: W.A. Matamoros, M. Medina and J.C. Carrasco, 3 July 2009 (Fig. 2).
Paratypes: LSUMZ 14851 (n=9), field number WAM08-141, Honduras, Department of Santa Barbara, Drainage: Ulúa, small creek that drains into the main river, 15.029520°N, 88.508°W, Collectors: W.A. Matamoros, F. Elvir and H. Vega, 7 August 2008; LSUMZ 14852 (n=4), same data as the holotype; USM 39024 (n=9), same data as the holotype; USM 39025 (n=7), same data as LSUMZ 14851; USM 39026 (n=5), same data as LSUMZ 14851.
Profundulus kreiseri is a new member of the subgenus Profundulus and shares with other members of that subgenus (viz., Profundulus (Profundulus) punctatus, Profundulus (Profundulus) guatemalensis and Profundulus (Profundulus) oaxacae) the following set of characters: dark humeral spot, a scaled preorbital region and between 32-34 vertebrae. It differs from all members of the subgenusby having less than half of its caudal fin densely covered with scales. It can further be distinguished from Profundulus (Profundulus) oaxacae and Profundulus (Profundulus) punctatus by the absence of conspicuous brown spots on the sides of the body; from Profundulus (Profundulus) guatemalensis by having fewer pectoral-fin rays (17-20 in Profundulus (Profundulus) guatemalensis versus 13-16 in Profundulus kreiseri) and caudal-fin rays (19-23 in Profundulus (Profundulus) guatemalensis versus 13-18 in Profundulus (Profundulus) kreiseri); and from Profundulus (Profundulus) guatemalensis and Profundulus (Profundulus) punctatus by having a golden blotch that covers most of the operculum and reaches the base of the pectoral fin. Profundulus (Profundulus) kreiseri is distinguishable from all members of the subgenus Tlaloc (viz., Profundulus (Tlaloc) candalarius, Profundulus (Tlaloc) hildebrandi, Profundulus (Tlaloc) labialis, Profundulus (Tlaloc) portillorum) by having a dark humeral spot and a scaled preorbital. Profundulus (Profundulus) kreiseri can further be differentiated from Profundulus (Tlaloc) candalarius, Profundulus (Tlaloc) hildebrandi and Profundulus (Tlaloc) labialis by having between 32-34 vertebrae (versus 35-39).
Morphometric and meristic data for type material are summarized in Table 2. The largest specimen is 81.2 mm SL. The body is elongate with the dorsal and ventral profiles nearly symmetrical. The narrowest point on the body is the tip of the snout, with the body expanding gradually dorsally and ventrally to the deepest point slightly anterior to the verticals through the dorsal- and anal-fin origins. The vertical through the origin of the dorsal fin is slightly anterior to the origin of the anal fin. The body depth narrows in the region of these two unpaired fins, and the dorsal and ventral body margins are straight and parallel on the caudal peduncle before diverging out slightly at the origin of the caudal fin.
The head (including cheek, infraorbital and preorbital regions) is covered with scales that are deeply embedded in the skin. The mouth is terminal, the lower jaw protruding slightly beyond the upper. The posterior portion of the maxilla extends ventrally to a vertical through the anterior region of the orbit.
The number of dorsal-fin rays ranges from 10-12 (holotype=11). The number of pectoral-fin rays ranges from 13-16 (holotype=14). The posterior edge of the pectoral fin does not reach the pelvic-fin origin. The number of anal-fin rays ranges from 9-14 (holotype=12; mode=12). The caudal fin is rounded, the number of fin rays ranging from 13-18 (holotype=17). All specimens examined have six pelvic-fin rays. The number of scales along the midline of the body ranges from 33-34 (holotype=33). The number of scales around the caudal peduncle ranges from 9-10 (holotype=10). The number of vertebrae ranges from 31-33 (holotype=32).
In life this species is brown, with a golden-yellow blotch that covers most of the operculum and reaches the base of the pectoral fin. An inconspicuous dark stripe is present along the midline of the body starting at a vertical between the dorsal- and anal-fin origins and terminating at the origin of the caudal fin.
Thebody is a uniform dusky brown with a prominent dark humeral spot posterior to the upper insertion of the pectoral fin. A dark stripe is present along the midbody; this stripe is more conspicuous in preservation than in life. The distal margins of the unpaired fins are opaque, but the basal ¾ of the fins are covered with scattered melanophores.
Profundulus kreiseri is only known from the middle reaches of the Chamelecón and Ulúa rivers in Honduras (Figs. 1, 3).
Ecological notes. The only known localities of Profundulus kreiseri are both characterized as small tributaries ranging in width from 0.8 to 4 meters with stones (from pebbles to boulders) as the dominant substrate. The canopy cover of both localities is estimated to be 70–80%. Both creeks feature a variety of run, pool, riffle, rapid and small waterfall habitats (Fig. 3).
The limited range of this species makes it vulnerable to extinction via habitat loss. The creation of a new hydroelectric dam on theChamelecón River will likely drastically impact populations of this new species.
The specific epithet is in honor of Dr. Brian R. Kreiser, the doctoral advisor and friend of the first author.
Suggested English name: Kreiser’s Killifish
Suggested Spanish name: El Escamudo de Kreiser
Morphological data and counts from the holotype and 33 paratypes of the new species Profundulus kreiseri. Measurements are presented as % of standard length (%SL) or % of head length (%HL). SD=standard deviation.
| Morphological data | Holotype | Range | Mean | SD |
|---|---|---|---|---|
| Standard length | 53.2 | 46.1–81.2 | 60.3 | 9.7 |
| Snout length (%HL) | 30.1 | 20.4–36.1 | 28.9 | 3.6 |
| Head length (%SL) | 26.0 | 24.0–28.8 | 25.8 | 1.2 |
| Predorsal length (%SL) | 70.9 | 20.4–74.9 | 68.9 | 2.2 |
| Anal-fin origin to caudal-peduncle base (%SL) | 32.0 | 29.6–35.7 | 31.9 | 1.7 |
| Anal-fin length (% SL) | 32.0 | 28.5–34.9 | 31.9 | 1.8 |
| Eye diameter (%HL) | 23.0 | 20.4–29.5 | 23.4 | 2.1 |
| Head depth (%HL) | 75.6 | 42.8–78.1 | 69.4 | 6.6 |
| Maximum body depth (%SL) | 24.9 | 18.1–25.7 | 23.2 | 2.0 |
| Caudal-peduncle depth (%SL) | 14.0 | 12.0–15.5 | 13.8 | 0.8 |
| Head width (%HL) | 66.9 | 36.2–68.1 | 61.2 | 6.6 |
| Maximum body width (%SL) | 17.4 | 10.2–19.5 | 16.0 | 1.8 |

Photographs of preserved specimens of A Profundulus kreiseri sp. n., USM 39022, holotype, 53.2 mm SL, Río Chamelecón, Honduras B Profundulus punctatus, UMMZ 194154, 47.0 mm SL, Rio Nahualete, Guatemala; and C Profundulus guatemalensis, UMMZ 190542, 43.7 mm SL, Río Maria Linda, Guatemala. The diagnostic characters of each species (A–C) are shown in D–F the white bar shows the extent of squamation on the caudal fin D Profundulus kreiseri sp. n., USM 39022, holotype (note that scales cover less than half of the caudal fin, and there are no rows of dark spots on the caudal peduncle and caudal-fin base) E Profundulus punctatus, UMMZ 194154 (note that the entire anterior half of the caudal fin is covered with scales, and rows of dark dots occur on the caudal peduncle and caudal-fin base; and F Profundulus guatemalensis, UMMZ 190542 (note that scales extend beyond the midpoint of the caudal fin, and there are no rows of dark spots on the posterior portion of the caudal peduncle and caudal-fin base).

Photographs of the habitat of Profundulus kreiseri. A Type locality of Profundulus kreiseri, 15.197667°N, 88.616°W. A small creek that drains directly into the Chamelecón River, near the new hydroelectric dam. Department of Santa Barbara, Municipality of Macuelizo B Small creek that drains into the main river. Department of Santa Barbara, 15.029520°N, 88.508°W.
The phylogenetic analysis included sequences from 12 taxa (10 in the ingroup and 2 outgroups). Novel sequences were deposited in GenBank under accession numbers JQ254929-JQ254935 (Table 1). A total of 990 bp of cytochrome b were analyzed, of which 280 sites (28.3%) were parsimony informative. The GTR + I + G model was selected as the best fit for the dataset by jModelTest using the AIC. The optimized parameters were: A=0.2692, C=0.2640, G=0.2334, T= 0.3335, Rmat= (8.4428, 56.7004, 8.4428, 1.0000, 56.7004, 1.0000), the gamma distribution was 1.4610, and the proportion of invariable sites was 0.4970.
The same tree topology was recovered by MP (a single tree, length: 585, CI: 0.735; RI: 0.796) and BA (Fig. 4). Two monophyletic groups corresponding to the two subgenera were recovered. Node support was generally high throughout the tree (Fig. 4). The new species was recovered as the sister group of Profundulus guatemalensis with high bootstrap and posterior probability values. P- distances between the members of the two subgenera ranged from 17.8 to 19.3%. Within the subgenus Tlaloc, sequence divergence ranged from 1 to 9.2%. The shortest genetic distance was found between Profundulus labialis and Profundulus candalarius (1%). Sequence divergence between the new species (two specimens) and each of the three other species in the subgenus Profundulus ranged from 4.1 to 6.5% (Table 3).
Uncorrected pairwise genetic divergence based on cytochrome b sequences. C= chamelecón river, u= ulúa river, n= nacaome river.
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |
|---|---|---|---|---|---|---|---|---|---|
| 1. Profundulus (Profundulus) kreiseri (C) | 0 | ||||||||
| 2. Profundulus (Profundulus) kreiseri (U) | 0.016 | 0 | |||||||
| 3. Profundulus (Profundulus) guatemalensis | 0.041 | 0.047 | 0 | ||||||
| 4. Profundulus (Profundulus) punctatus | 0.062 | 0.064 | 0.064 | 0 | |||||
| 5. Profundulus (Profundulus) oaxacae | 0.063 | 0.065 | 0.063 | 0.040 | 0 | ||||
| 6. Profundulus (Tlaloc) portillorum (U) | 0.185 | 0.184 | 0.182 | 0.181 | 0.178 | 0 | |||
| 7. Profundulus (Tlaloc) portillorum (N) | 0.187 | 0.186 | 0.183 | 0.180 | 0.178 | 0.008 | 0 | ||
| 8. Profundulus (Tlaloc) hildebrandi | 0.191 | 0.191 | 0.186 | 0.183 | 0.184 | 0.080 | 0.080 | 0 | |
| 9. Profundulus (Tlaloc) labialis | 0.192 | 0.193 | 0.183 | 0.187 | 0.186 | 0.087 | 0.087 | 0.078 | 0 |
| 10. Profundulus (Tlaloc) candalarius | 0.192 | 0.193 | 0.183 | 0.183 | 0.187 | 0.090 | 0.092 | 0.083 | 0.010 |
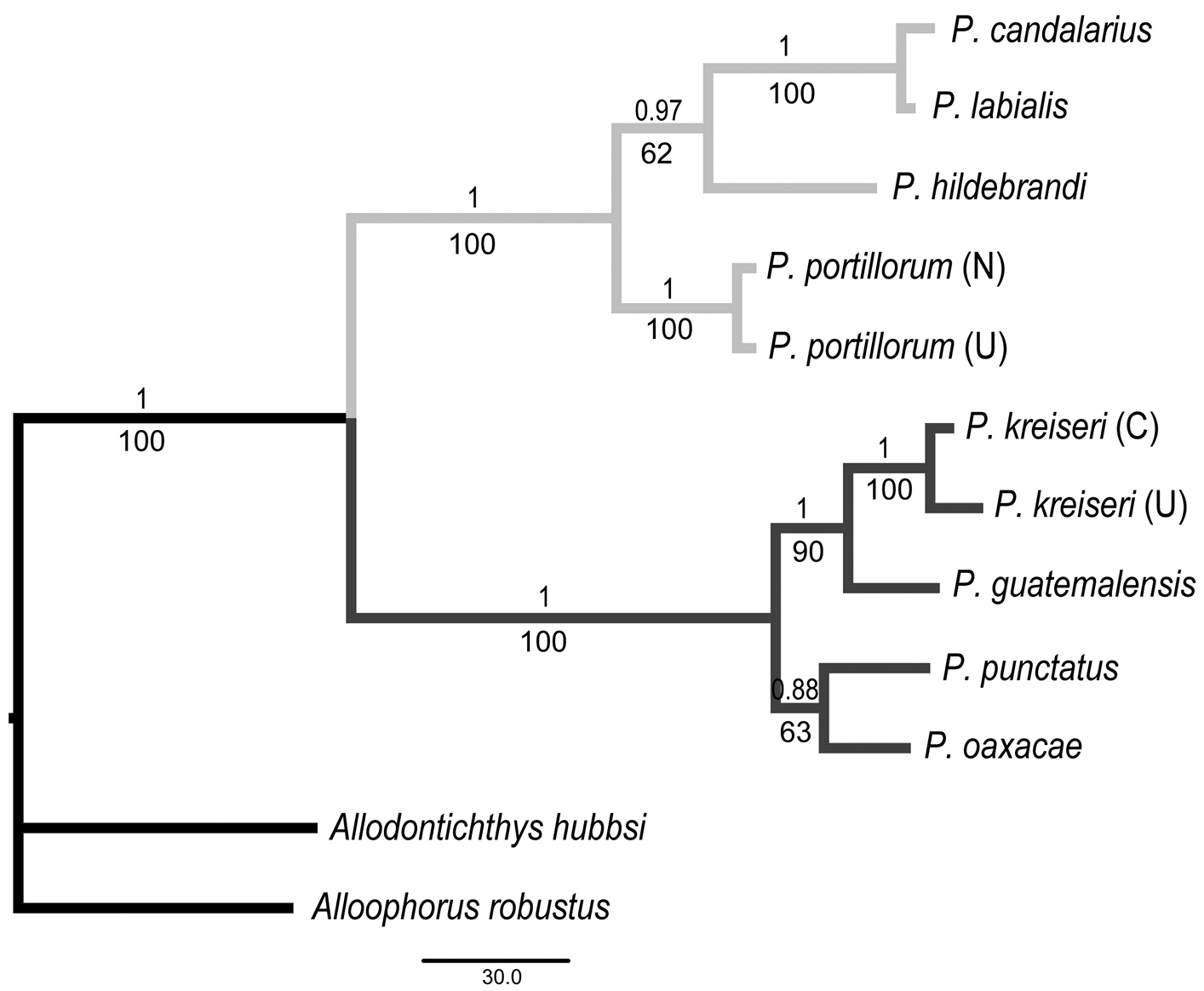
Bayesianand maximum parsimony trees based on cytochrome b sequences. Number above each node represents Bayesian posterior probabilities. Number below each node represents nonparametric bootstrap support. Subgenus membership is shown with either light gray branches (Tlaloc) or dark gray branches (Profundulus). C = Chamelecón River, U = Ulúa River, N = Nacaome River.
Profundulus portillorum has the southernmost range of any species in the family; it is found in the Ulúa River in the Atlantic slope of Honduras and the Nacaome River in the Pacific slope of Honduras (
Profundulus kreiseri has higher intraspecific genetic variation (0.016) than is found between some species (e.g., 0.010 in Profundulus labialis versus Profundulus candalarius). The low sequence divergence found between Profundulus labialis and Profundulus candalarius may be a reflection of a recent divergence. These two taxa are currently allopatric and can be differentiated based on morphological characters (e.g., fin ray counts, body depth). However, additional work may be needed to verify the taxonomic status of these taxa given the results from our molecular analysis. Intraspecific variation in Profundulus kreiseri is also twice as high as what was recovered for the only other species for which multiple individuals were sampled in our study, Profundulus portillorum (0.008). This variation in Profundulus kreiseri may reflect a relatively long period of isolation between the populations in the Chamelecón and Ulùa rivers.
Our molecular phylogenetic analysis recovered two distinct clades corresponding to the nominal subgenera and placed Profundulus kreiseri within the subgenus Profundulus as the sister taxon of Profundulus guatemalensis.The caudal-fin scales of Profundulus kreiseri do not extend beyond the anterior half of the fin (Fig. 2D), whereas they cover half or more of the caudal fin in other members of the subgenus Profundulus (e.g., Profundulus punctatus and Profundulus guatemalensis—Fig. 2E, F). Because the presence of squamation on at least half of the caudal fin is a defining character of the subgenus (
| 1a | Preorbital region usually naked, frequently with one or two isolated inconspicuous embedded scales; humeral spot absent | 2 |
| 1b | Preorbital region with at least three to four conspicuous (not deeply embedded under the skin) scales; humeral spot present | 4 |
| 2a | Lower jaw protruding beyond upper jaw | Profundulus labialis |
| 2b | Lower and upper jaws same length | 3 |
| 3a | Pectoral-fin rays 16 – 19. Endemic to the Grijalva-Usumacinta River basin | Profundulus candalarius |
| 3b | Pectoral-fin rays 13 – 16. Endemic to Honduras | Profundulus portillorum |
| 4a | Body of adults with conspicuous brown spots (distinctly within the scales) in longitudinal rows on the midline from the midpoint of the body to the caudal fin; dorsal surface of head concave to nearly flat | Profundulus punctatus |
| 4b | Body of young and adults lacking distinct brown spots (as described in 4a); dorsal surface of head convex (rounded) | 5 |
| 5a | Scales on the caudal fin covering at least the anterior half or more of the fin. Caudal-fin rays 19-23, very rarely 18 or 24; pectoral-fin rays 17-20, very rarely 16 or 21 | Profundulus guatemalensis |
| 5b | Scales on the caudal-fin covering less than half of the fin. Caudal-fin rays 13-18, pectoral-fin rays 13–16. | Profundulus kreiseri sp. n. |
Profundulus candalarius: UMMZ 1767317, n = 10, 23–44 mm SL, Guatemala, Huehuetenango, tributary of Río Lagartero in Ciénega Lagartero; UMMZ 2108276, n = 10, 41–66 mm standard length, LS, Mexico, Chiapas, Arroyo tributary to Río Comitan, 11.5 km east of Highway 190 onthe road to Montebello National Park. Profundulus guatemalensis: UMMZ 1905421, n = 15, 43–62 mm SL, Guatemala, Escuintla, Río Marinala at Finca Peña Blanca 11 km north-east Escuintla; UMMZ 2191352, n = 4, 37–49 mm SL, Honduras, Río Copan bridge crossing on Ca 11, 26.4 km east of Copan.
Profundulus punctatus: UMMZ 1847303, n = 10, 42–51 mm SL, Mexico, Chiapas, Stream at Piedra Parada, 12.1 km west-north-west of Ocozocoautla; UMMZ 1941544, n = 4, 54–70 mm SL, Escuintla, Guatemala, Río Siguacan at Ca Highway 2, km 120, 8 km east of Río Bravo; UMMZ 1941605, n = 5, 55–66 mm SL, Escuintla, Guatemala, Río Aguna at CA 2, km 98, 15 km west-north-west.
Profundulus hildebrandi : UMMZ 1576348, 15, 46–103 mm SL, Mexico, Chiapas, Laguna de Maria Eugenia, San Cristobal de las Casas; UMMZ 1839149, n = 5, 33–47 mm SL, Mexico, Chiapas, Irrigation ditch of Laguna Maria Eugenia at Highway. 190, 2.4 km southwest of San Cristobal de las Casas. Profundulus labialis: UMMZ 16669811, n = 5, 50–71 mm SL, Guatemala, Rio Carchela c. 29·0 km north of Salama on road to Coban; UMMZ 17672610, n = 15, 43–64 mm SL, Guatemala, Huehuetenango, stream through pasture southwest of Jacaltenango.
Profundulus portillorum: USM 31597, n = 31, field number WAM07-03, Honduras, department of Comayagua, municipality of Potrerillos, in the town of Siguatepeque. Drainage: Ulúa. System: Ulúa: Río Calam. Quebrada de Potrerillos at Barrio San José, 14.53000°N, 87.84000°W, Collectors: Matamoros W.A. and H. Portillo; USM 31628, n = 21, field number WAM07-39, Honduras, department of Francisco Morazan, municipality of Lepaterique, Basin: Pacific. Drainage: Nacaome, System: Nacaome, Quebrada El Sapo, near the community of Lepaterique, 100 meters away from Catholic Church, 14.064275°N, 87.466850°W, Collectors: Matamoros W.A., A. Sanchez, E. Lopez and J. Hernandez.
Profundulus kreiseri: USM 39028, 9, field number WAM08-141, Honduras, department of Santa Barbara, Drainage: Ulúa, System: Ulúa, small creek that drains in to main river. 15.029520°N, 88.508°W. Collectors: W.A. Matamoros, F. Elvir and H. Vega; USM 39027, n = 3, same data as USM 39028.
Financial support for this project was provided by grants from the World Wildlife Fund, the United States Agency for International Development Project MIRA Honduras, and the Critical Ecosystems Partnership Fund to WAM, and a National Science Foundation grant # 0916695 to PC. Funds to survey the middle reaches of the Chamelecón River were provided by the Honduran company Generación de Energía Renovable (Gersa) in July 2009. We are grateful to J. I. Restrepo for the logistical support provided while we were in Honduras, and also to R. Jordan and P. Herrero of USAID–MIRA Honduras for logistical support. We also extend our gratitude to Melissa Medina, Juan Carlos Carrasco, Fausto Elvir, Hermes Vega, Alejandra Sanchez, Jonathan Hernandez and Edis Lopez for their assistance in the field. We are very thankful to Said Lainez, Iris Acosta and Claudia Carcamo from the Instituto de Conservacíon Forestal for their help in obtaining collection permits in Honduras. Many thanks to Ignacio Doadrio, Rocio Rodiles-Hernández and Alfonso González-Díaz for providing tissue for our DNA analysis.