





(C) 2013 Mikhail Potapov. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
In this paper, we describe the new genus Pauropygus gen. n. which includes three minute species, blind and unpigmented, living in interstitial littoral habitats in tropical or subtropical countries. Two of these species are new to science (type species Pauropygus projectus sp. n. from New Caledonia and Pauropygus pacificus sp. n. from China); the third one, originally described in the genus Cryptopygus (Cryptopygus caussaneli Thibaud, 1996), has a larger pantropical distribution. We synonymize here Cryptopygus riebi Barra, 1997 from South Africa with Pauropygus caussaneli. Two paratypes of the Mexican species Cryptopygus axayacatl Palacios & Thibaud, 2001 turned also to be Pauropygus caussaneli, while the holotype and remaining paratypes of this species support its placement in Proisotomodes. Among the Cryptopygus complex, Pauropygus gen. n. is easily recognized by characters of mouthparts (presence of two large projections on pleural fold, basolateral field with 6 chaetae, modified mouthparts) and reduced sensillar chaetotaxy (tergal sensilla 2-3, 0-1/0-1, 0-1, 1-2, 1-2, 1-3, microsensilla reduced in number: 00/0-100, with sensilla situated in p-row on the abdomen). Small size, absence of eyes and pigment are also shared by all its species. The three species belonging to the genus differ by sensillar chaetotaxy.
New genus, modified mouthparts, sensillar chaetotaxy, littoral habitat
The species of the subfamily Anurophorinae with furca and two last abdominal segments fused are widely distributed in the world. If special remarkable detail had not been found in such species, they were usually assigned to genera Isotomina Börner, 1903 (mostly in Northern Hemisphere) or Cryptopygus Willem, 1902 (mostly in Southern Hemisphere). In the course of the development of taxonomy in the family Isotomidae the question of division of these two artificial taxa became important. So far Isotomina, Cryptopygus and several formally similar genera have been given some discussions and historical reviews (
urn:lsid:zoobank.org:act:FD85013E-80AF-46F9-9BC9-564339BF37E5
Pauropygus projectus sp. n.
Blind small-sized Anurophorinae with two last abdominal segment fused, modified mouthparts including remarkably modified pleural fold, and first segments of antenna set together on frontal part of head.
Without pigment and eyes, Abd.V and VI fused. Body size small, with primary granulation only. Antennal bases set close together on frontal side of the head, almost touching each other (Figs 1, 2). Sensilla on three first antennal segments thickened. Sensilla of Ant.IV moderately thickened. Maxillary palp simple, with 3 sublobal hairs set together (Fig. 3). Pleural fold with two high projections (Figs 3, 20, 23). Basolateral field of the labium with 6 chaetae (Fig. 20). Labium with three papilla projected forward, number of guards not reduced. Papillate sensilla reduced in size. Labrum swollen in central part, labral chaetae set on wide papilla. Two prelabral chaetae. Maxillary head with four enlarged lamellae, three of them ciliated; claw reduced to small finger-like process, not dentate. Mandible head reduced and thin, molar plate with 2 strong basal teeth. Tergal sensilla on abdomen situated in p-row of chaetae, their number reduced (2-3, 0-1/0-1, 0-1, 1-2, 1-2, 1-3, depending on species), number of microsensilla 00/100 or 00/000 (Figs 8, 15, 21). Th.I-III without ventral chaetae. Body macrochaetae differentiated. Tibiotarsal tenent chaetae present (1-2-2), not clavate. VT with 4+4 laterodistal chaetae. Tenaculum with 4+4 teeth. Furca slender, manubrium with a pair of chaetae on anterior side, dens with crenulation and wide swelling on posterior side, mucro bidentate.
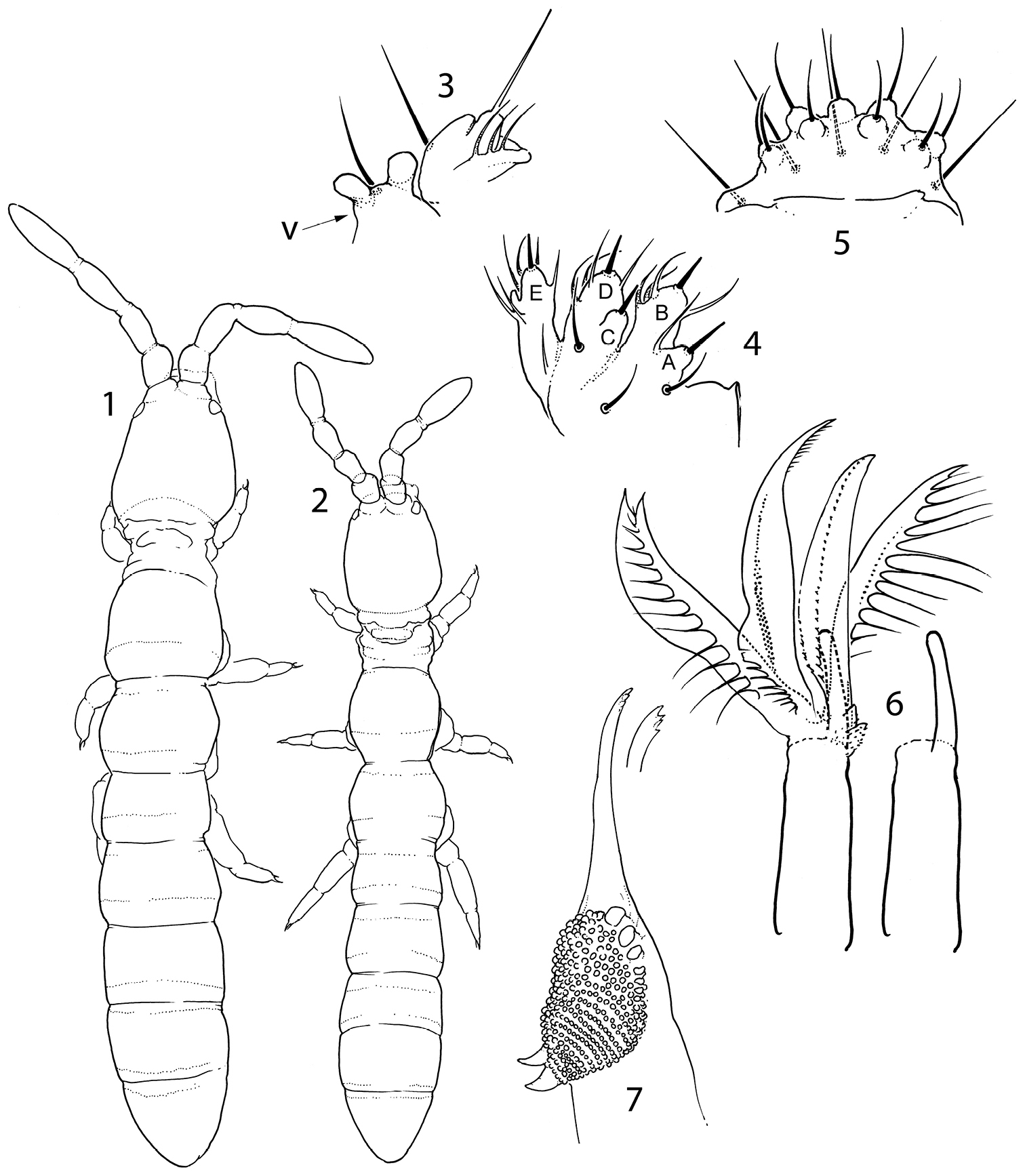
Pauropygus projectus sp. n. (1, 3–7) and Pauropygus caussaneli (2). 1–2 general habitus (dorsally) 3 maxillary outer lobe and pleural fold (v: v-shaped process.) 4 labial palp 5 labrum 6 maxillary head (on right, basal part and claw shown) 7 mandible.
Pauropygus projectus sp. n. 8 sensillar chaetotaxy 9 posterior part of abdomen (s – sensillum) 10 distal half of Leg III 11 PAO and Ant.I-III, dorsal view 12 subapical organite and microchaeta.
The name is derived from the Greek suffix –pygus which points to the fusion of the abdominal segments and from the Greek prefix pauro- which refers to the reduced chaetotaxy, particularly reduced number of sensilla on the body.
Apart from the type species Pauropygus projectus sp. n., the new genusincludes Cryptopygus caussaneli Thibaud, 1996 and Pauropygus pacificus sp. n.
The new genus is established mainly due to the unusual position of antennae on head and the strongly modified mouthparts. In mouthparts, the V-shaped pleural folds and the presence of 6 chaetae on basolateral fields of the labium are especially important; these characters were never seen in the family before, except for the latter one that was mentioned for part of the Algerian population of Isotominella geophila sensu Jordana et al., 2009; normally, the pleural fold looks like a weak swelling proximal to the maxillary outer lobe, and basolateral field of mouth cone has 5 chaetae in species of the Isotomidae family (
As an unusual feature for the group, the species of the new genus show considerable reduction of sensillar chaetom. In Pauropygus projectus sp. n. all medial sensilla on Th.II-Abd.IV and lateral sensilla on Th.III-Abd.II are lost, while in Pauropygus caussaneli and Pauropygus pacificus sp. n. it is the posterior and lateral parts of body that lost sensilla. In spite of differences between number of sensilla on body (2, 0/0, 0, 1, 1, 3) and (3, 1/1, 1, 2, 2, 3) among Pauropygus species, the general pattern of their distribution and differentiation is kept.
Pauropygus is closely related to Isotominella Delamare Deboutteville, 1948 after the redescription of
urn:lsid:zoobank.org:act:4747ED35-9352-4F40-B567-598C47E6BF71
http://species-id.net/wiki/Pauropygus_projectus
Figs 1, 3–14Holotype female on slide and 13 paratypes (6 on slides, 7 in alcohol): New Caledonia: Iles Loyautés: Ouvéa island: Gossanah: plage de Hoony, collected by flotation from beach sand, 13.11.2000 (sample # NC00-252), leg L. Deharveng & A. Bedos. Coordinates: 166.632°E, 20.4365°S. Material is deposited in Museum National d’Histoire Naturelle, Paris (holotype and 9 paratypes, no male) andin Moscow State Pedagogical University (4 paratypes, including one adult male).
Other material. New Caledonia: Iles Loyautés: Tiga island: Toka village, collected by flotation from beach sand, 31.10.2000 (sample # NC00-145: two on slides; sample # NC00-146: one on slide), leg L. Deharveng & A. Bedos. Coordinates: 167.795704°E, 21.098038°S.
Size 0.5–0.6 mm. White, without eyes. Cuticle with thin hardly visible primary hexagonal granulation (“smooth”). PAO flat, roundish, not constricted, about 1.5 as long as inner edge of U.III and shorter than width of Ant.I (Fig. 11). Sublobal plate of maxillary outer lobe small with 3 hairs grouped together, palp simple. Pleural fold with one chaeta (as common for the family) and two high projections (Fig. 3). Labral chaetotaxy as 2/554, middle part of labrum swollen, chaetae in two apical rows set on wide papillae, edge of labrum weakly developed (Fig. 5). Labium with 3 proximal, 6 basolateral and 4 basomedian chaetae and a complete set of papillae (A-E) and guards (16). Papillae B, D and E projected considerably forward, papillae A and C partly reduced and fused with B and D, respectively (Fig. 4). Ventral side of head with 6-7+6-7 postlabial chaetae. Ventrolateral chaetae of head and postlabial chaetae delimit an almost unbroken unsetaceous area. Maxillary head elongated, with four well visible enlarged lamellae of which two have long cilia and two have fine serration. Two remaining lamellae possibly as small weakly serrated projections set in a common cluster at base of claw (Fig. 6). Maxillary claw reduced, single-tipped, finger-shaped, with some weak teeth at the head which are visible in lateral view only (not shown on figure). Pars incisiva of mandible slender, apically with four weak teeth, basal part of pars molaris with two strong hooks (Fig. 7).
Ant.I with many chaetae (more than 20), 1 ventro-basal microchaeta (bms; dorsal bms not differentiated), and 2 thick ventral sensilla (s). Ant.II with 3 rather large bms and 1 thick laterodistal s. Ant.III without bms and with 5 distal s of which two inner as thick and short as outer ones. Male antennal “spurs” unknown. Sensilla on Ant.IV weakly differentiated, as common for the family, subapical organite pin-like and small, subapical microsensillum short and curved (Figs 11, 12).
Dorsal axial chaetom of Th.II–Abd.III as 12-13, 7-9/5(4), 5(4), 5(4). Macrochaetae erect, smooth and rather long, more differentiated laterally on Th.II–III and on posterior half of abdomen, with the whole number as 1, 1/3, 3, 3, 4 (Th.II-Abd.IV). Medial macrochaetae on Abd.V about 0.4 as long as dens. Sensilla on tergites clearly differentiated, significantly shorter and slightly thinner than ordinary chaetae. Sensillar formula 20/00113 (s), 00/100 (ms) (Fig. 8). Lateral sensilla on Abd.III, IV in posterior position. Sensilla on Th.II and Abd.III much longer than on Abd.V, sensilla on Abd.IV of medium size (Fig. 9).
Unguis of normal shape, without inner tooth, two lateral teeth forming a weak tunica partly covering dorsal edge of unguis (Fig. 10). Upper subcoxa of Leg I-III with 1, 1, 4 chaetae, lower subcoxa with 1, 10, 11 (one individual studied). Ti.I-III with basic set of chaetae (21-22, 22-23, 28), T-chaetae absent. Chaetae on tibiotarsi with irregular distribution, outer side with higher number of chaetae than inner side. Chaetae C7 (inner part of tibiotarsi) either lost or migrated laterally. Modification of chaetae x and B5 on Ti. III in males unknown (no males). Distal tibiotarsal tenent chaetae on Ti.I-III (1-2-2) well developed, not clavate, about 1.5 as long as U.III. Each tibiotarsus with one (sometimes two on Leg II) additional tenent chaeta at middle part. Ti.III with two stick-like chaetae (A6, A7) in distal ring. Ti.I, II with similar thickened chaetae but much less developed. One or two inner chaetae of distal ring of Ti.I-III shorter than others. Three chaetae of distal ring A (two tenent chaetae and one nearby) set apart from distal edge of tibiotarsus, unlike other chaetae of the ring (Fig. 10). Pretarsus with two chaetae, inner one shorter. Ventral tube with 4+4 laterodistal and 2 posterior chaetae. Tenaculum with 4+4 teeth and 1 chaeta. Anterior furcal subcoxa with 13–15 chaetae, posterior with 6-7. Furca of medium size. Anterior side of manubrium with a pair of distal chaetae, posterior one with 8+8 chaetae on the main part and 3+3 on the laterobasal lobes (one individual studied). Manubrial thickening with a pair of additional inner teeth. Lateral parts of manubrium with 1+1 chaetae. Dens slender, anteriorly with 12 (more rarely 11) chaetae. Posterior side of dens slightly crenulated in the medial part, with 5 chaetae of which 3 basal and 2 at the medial part (short inner and long outer) set together on wide papillum. Mucro slender with two teeth of which the subapical one is larger, lamellae not differentiated (Figs 13, 14). Ratio of manubrium: dens: mucro = 2.8–3.4: 4.5–4.6: 1. Anal lobes without microchaetae.
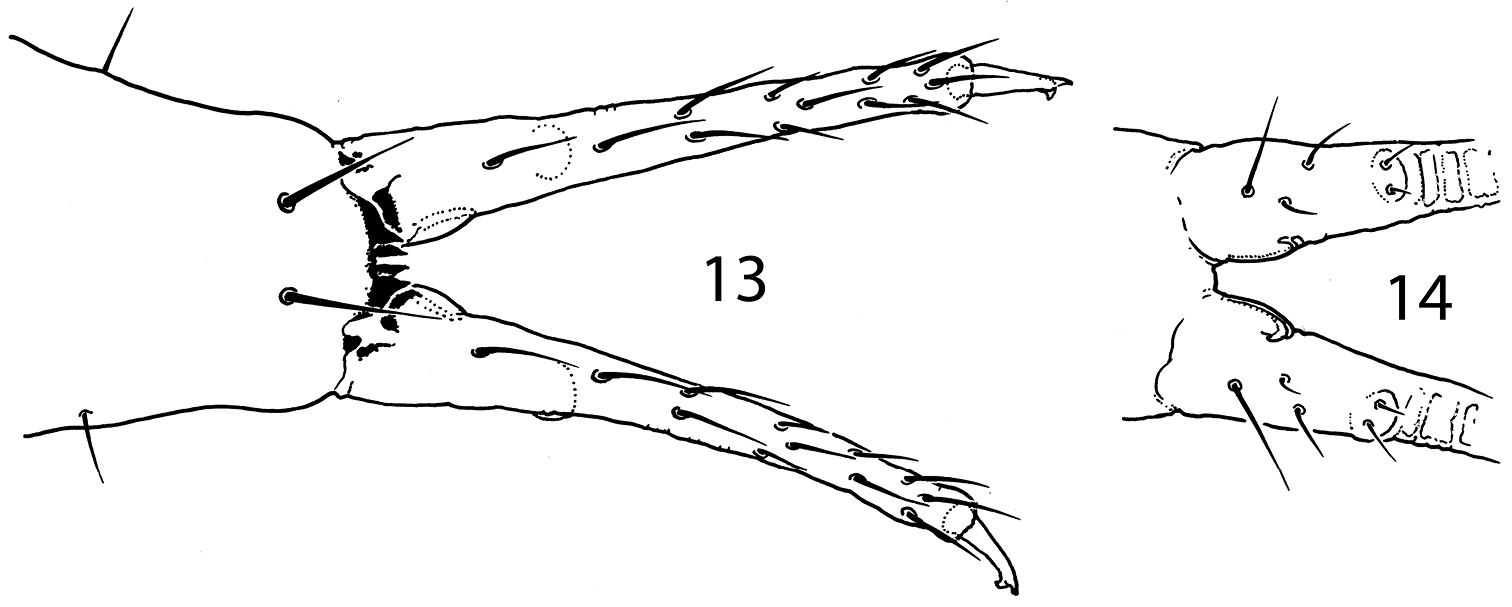
. Pauropygus projectus sp. n. anterior (13) and posterior (14, part) sides of furca.
Pauropygus projectus sp. n. differs from Pauropygus caussaneli in number of sensilla on body (20/00113 vs. 31/11221, see also the genus affinity section), relative size of inner and outer sensilla in AIIIO (outer ones smaller than inner ones in Pauropygus caussaneli), number of posterior chaetae on ventral tube (2 vs. 4), serrated chaetae on upper and lower subcoxae (smooth in Pauropygus caussaneli), more differentiated chaetal equipment of legs. Besides, Pauropygus projectus sp. n. is larger in size, which may explain higher number of chaetae on Ant.I, parts of legs, axial group of tergites and postlabial area than in Pauropygus caussaneli (see descriptions for details).
The species has well developed “projections” on the body as: swollen labrum, v-shaped pleural folds, extended labial palp, and antennae projected ahead.
Only from Loyalty Islands in New Caledonia.
http://species-id.net/wiki/Pauropygus_caussaneli
Figs 2, 15–20Holotype and paratypes of Cryptopygus caussaneli: Mauritania (Aftout-es-Sahel); holotype and paratypes of Cryptopygus riebi: South Africa (Sodwana Bay, Natal Province); two paratypes of Cryptopygus axayacatl Palacios-Vargas & Thibaud, 2001: Mexico (Guerrero, Acapulco); material of J.-M.Thibaud identified by him as Cryptopygus caussaneli from Senegal, Morocco, Congo, Madagascar, Maurice and Mayotte Islands. More precise labels in the associated publications (
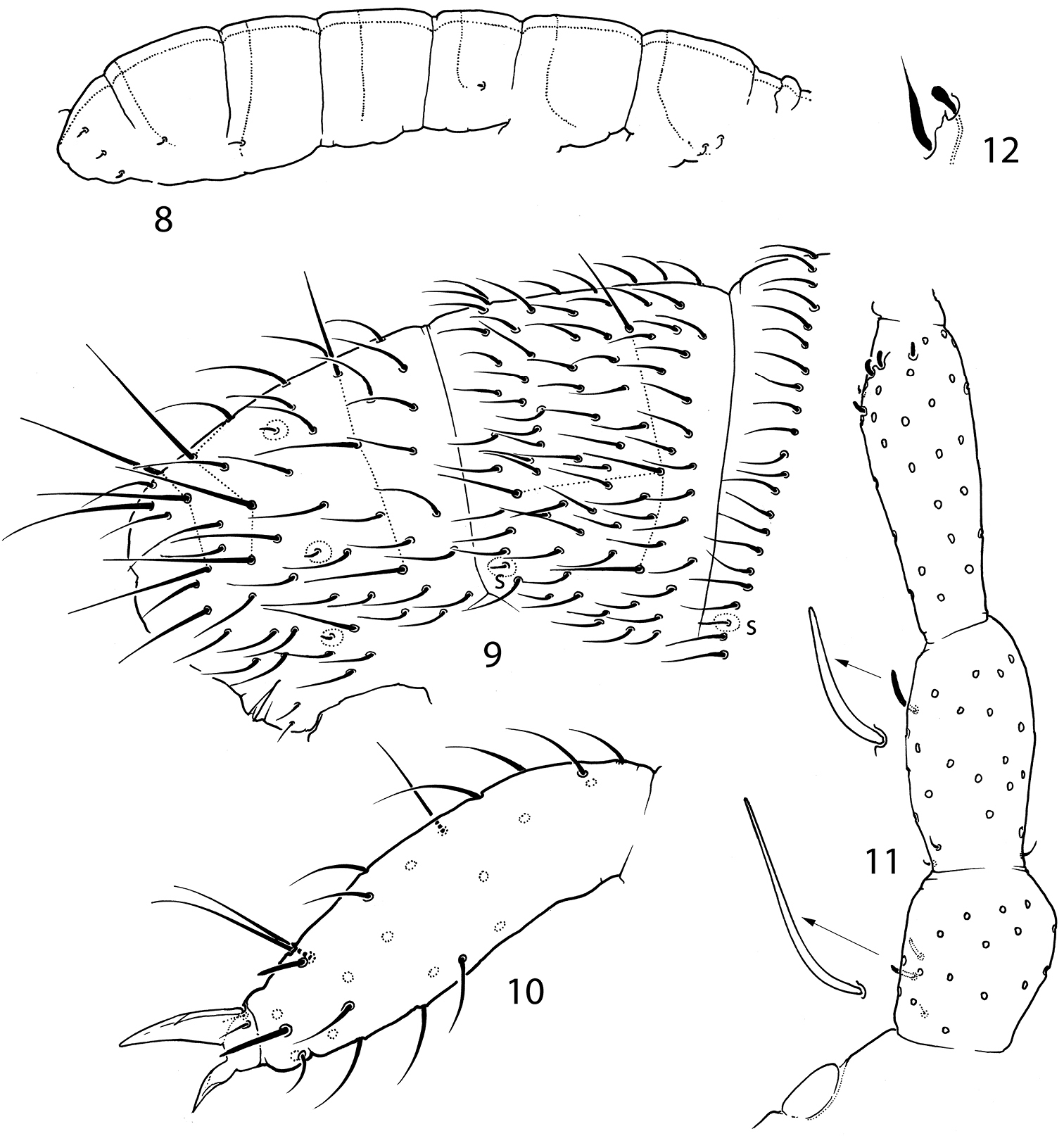
White, without eyes. Size up to 0.4 mm. Cuticle smooth. PAO about 1, 9 as long as inner edge of U.III and shorter (0.7) than width of Ant.I. Outer (Fig. 20) and inner mouthparts principally as in Pauropygus projectus. Ventral side of head with 5+5 postlabial chaetae. Ant.I with 14-15 chaetae, 1 ventro-basal microchaeta (bms; dorsal bms not differentiated), and 2 thick ventral sensilla (s), short and long. Ant.II with 3 bms and 1 thick laterodistal s. Ant.III without bms and with 5 distal s of which two inner thicker and longer than outer ones. Male antennal “spurs” present on Ant.II, III and basal part of Ant.IV.
Dorsal axial chaetom of Th.II–Abd.III as 10, 6/4, 4(3), 4(3). Thorax without ventral chaetae. Macrochaetae smooth, with the whole number as 1(2), 1(2)/3, 3, 3, 4 (Th.II-Abd.IV). Medial macrochaetae on Th.II-Abd.III hardly differentiated. Medial macrochaetae on Abd.V about 0.4 as long as dens. All sensilla of thorax and medial sensilla of abdomen nearly as long as ordinary chaetae and hardly visible, lateral sensilla on Abd.III-V shorter than ordinary chaetae. Microsensilla short but well visible. Sensillar formula 31/11221 (s), 00/100 (ms) (Fig. 15-18). Sensilla in posterior position. (Fig. 16).
Unguis of normal shape, without inner tooth, two broad unequal lateral teeth. Upper subcoxa of Leg I-III with 1, 1, 4 chaetae, lower subcoxa with 1, 6-7, 7-8. Ti.I-III with one chaeta lost in basic set (20, 21, >25). Chaetae x and B5 on Ti. III in males unmodified. Distal tibiotarsal tenent chaetae on Ti.I-III (1-2-2) well developed, not clavate, about 1.1-1.2 as long as U.III. Each tibiotarsus with one additional tenent chaeta at middle part. Ti.III with one stick-like chaetae (A7) in distal ring, its shape slightly varies (Fig. 19). Tenent hairs (1, 2, 2) about 1.1–1.2 as long as U.III (Fig. 19). Pretarsus with two chaetae. Ventral tube with 4+4 laterodistal and 4 posterior chaetae in one transversal row. Tenaculum with 4+4 teeth and 1 chaeta. Anterior furcal subcoxa with 12–15 chaetae, posterior with 6 ones. Manubrium principally as in previous species. Dens slender, anteriorly normally with 11 chaetae. Posterior side of dens slightly crenulated in the medial part, with 5 chaetae of which 3 basal and 2 at the medial part (short inner and long outer) set together on low papillum. Mucro slender with two teeth of unequal size. Ratio of manubrium: dens: mucro = 3.2–3.6: 4.1–4.3: 1. Anal lobes without microchaetae.
Pauropygus caussaneli. 15 sensillar chaetotaxy 16 posterior part of abdomen 17 lateral part of Abd.I 18 lateral part of Th.III. s sensillum, ms microsensillum.
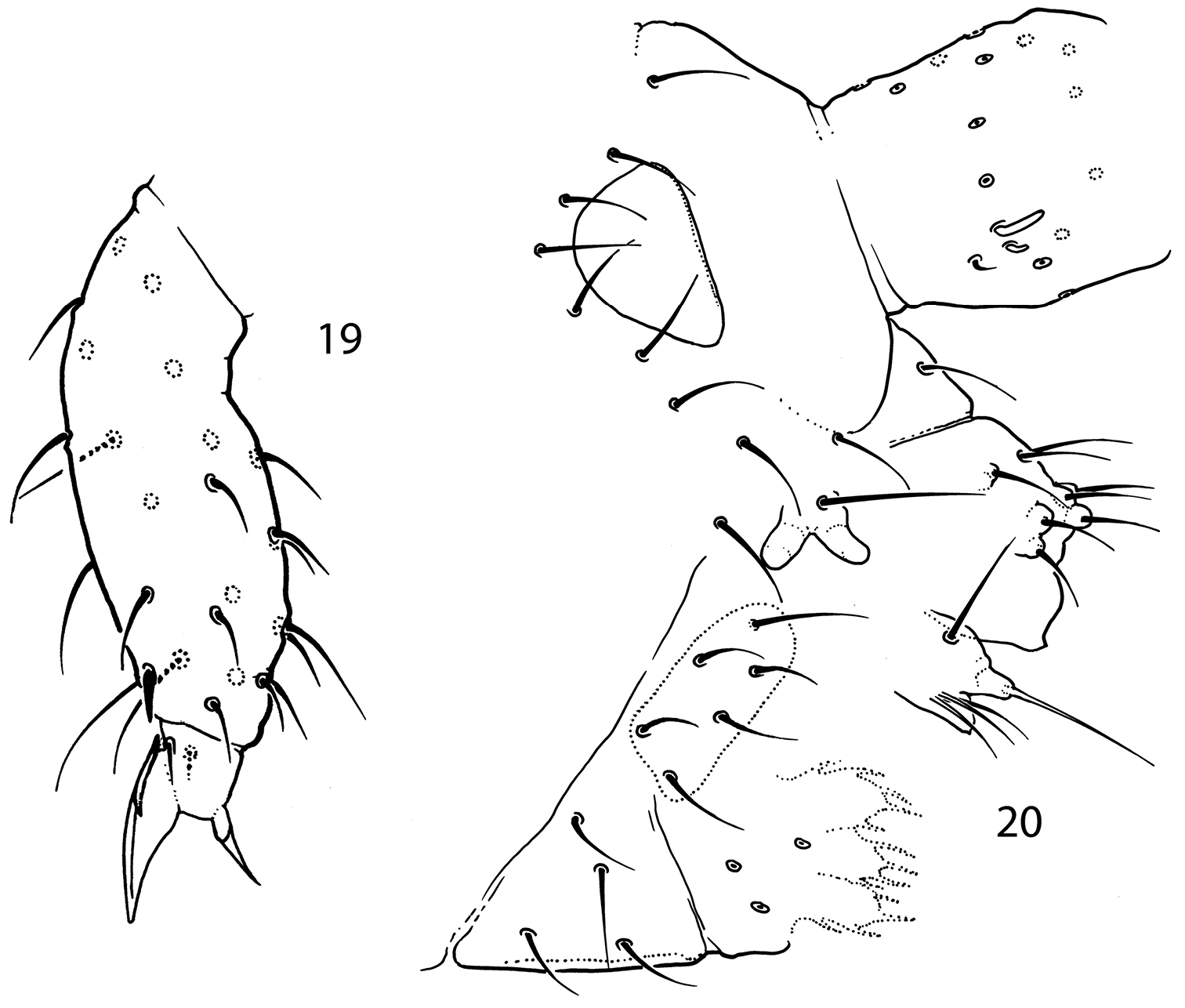
Pauropygus caussaneli. 19 distal half of Leg III 20 anterior part of head, lateral view (group of basolateral chaetae marked).
See above for differences with Pauropygus projectus sp. n. After our study of type material of Cryptopygus riebi (South Africa) and two paratypes of Cryptopygus axayacatl (Mexico) we concluded that these species only complement the wide distribution area of Pauropygus caussaneli. Holotype and other paratypes of Cryptopygus axayacatl support its species status, but in the genus Proisotomodes.
Probably widely distributed on sandy seashores of tropics and subtro-pics. Recorded from the coast of Indian and Atlantic Oceans (Africa, Central America).
urn:lsid:zoobank.org:act:AA6C41A0-CEAE-4961-B9EB-CC8AAB3B813C
http://species-id.net/wiki/Pauropygus_pacificus
Figs 21–24Holotype: China, Shandong Province, Yantey City, sea coast near Tuchengzi Hills, sandy beach, collected by flotation of thin sand under sparse vegetation on dunes, 22.04.2011, leg. Huang C.-W., Luan Y., and Potapov M.B.; 3 paratypes: at the same place; 2 paratypes: China, Hainan, western coast, nearby Changhua town and ca 5 km NE from the town, in both locations by flotation of sand with plant roots in dunes of sea coast, leg. Bu Y., Huang C.-W., Kuznetsova N.A., Potapov M.B. 06.04.2011. Material is deposited in Institute of Plant Physiology & Ecology, Shanghai (holotype and two paratypes) andin Moscow State Pedagogical University (three paratypes).
Size about 0.4 mm. White, without eyes. Cuticle smooth. PAO about 1.7 as long as inner edge of U.III and somewhat shorter (0.8-0.9) than width of Ant.I. Outer (Fig. 23) and inner mouthparts principally as in Pauropygus caussaneli. Ventral side of head with 5+5 postlabial chaetae. Ant.I with about 18 chaetae, 1 ventro-basal (bms) microchaeta (dorsal bms not differentiated), and 2 thick ventral sensilla (s), short and long. Ant.II with 3 bms and 1 thick laterodistal s. Ant.III without bms and with 5 distal s of which two inner thicker and longer than outer ones. Male antennal “spurs” present.
Thorax without ventral chaetae. Dorsal macrochaetae smooth, weakly differentiated, as 3, 3, 3, 4 on Abd.I-IV. Medial macrochaetae on Abd.V about 0.4 as long as dens. All sensilla of thorax and medial sensilla of abdomen nearly as long as ordinary chaetae and hardly visible, lateral sensilla on Abd.III-V shorter than ordinary chaetae. Microsensilla absent. Sensillar formula 31/11221 (s), 00/000 (ms) (Fig. 21). Sensilla in posterior position. Microsensilla on Abd.I absent (Fig. 22).
Unguis of normal shape, without inner tooth, two broad unequal lateral teeth. Upper subcoxa of Leg I-III with 1, 1, 4 chaetae, lower subcoxa with 1, 6-7, 7-8. Ti.I–III with 20, 21, >25 chaetae. Distal tibiotarsal tenent chaetae on Ti.I–III (1-2-2) well developed, not clavate, about 1.1-1.2 as long as U.III. Each tibiotarsus with one additional tenent chaeta at middle part. Ti.III with one stick-like chaetae (A7) in distal ring. Tenent hairs (1, 2, 2) slightly longer than U.III. Pretarsus with two chaetae. Ventral tube with 4+4 laterodistal and 4 posterior chaetae in one transversal row. Tenaculum with 4+4 teeth and 1 chaeta. Anterior furcal subcoxa with 12-14 chaetae, posterior with 6 ones (rarely 5). Manubrium principally as in previous species. Dens slender, anteriorly with 10-12 chaetae. Posterior side of dens slightly crenulated in the medial part, with 5 chaetae of which 3 basal and 2 at the medial part (short inner and long outer) set together on low papillum. Mucro slender with two teeth of unequal size (Fig. 24). Ratio of manubrium: dens: mucro = 3.7–3.8: 4.0–5.0: 1. Anal lobes without microchaetae.
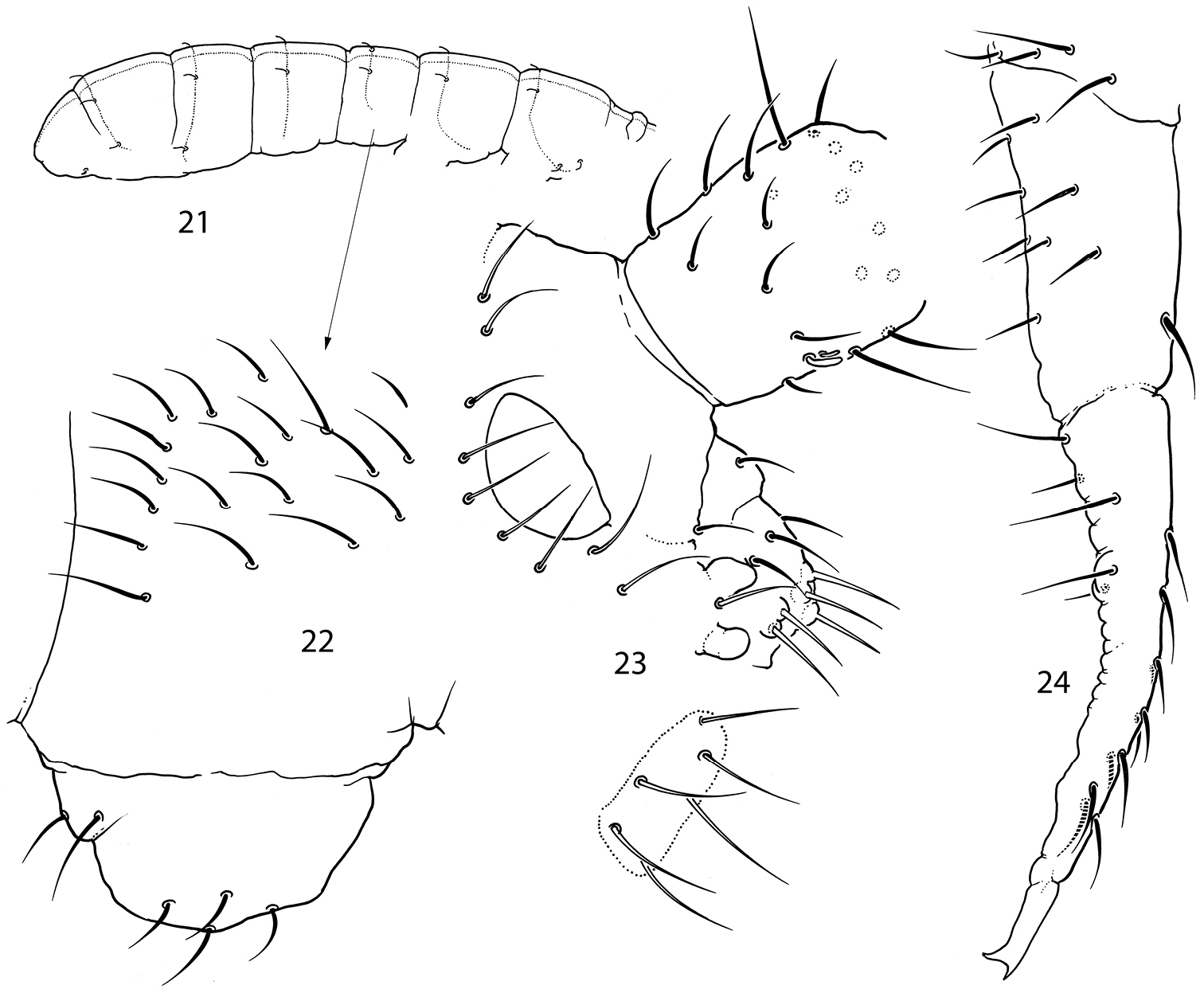
Pauropygus pacificus sp. n. 21 sensillar chaetotaxy 22 lateral part of Abd.I 23 anterior part of head, lateral view (group of basolateral chaetae marked, outer maxillary lobe and labial palp not shown) 24 furca, lateral view.
The complete absence of microsensilla on body of Pauropygus pacificus sp. n. is the second reported case among Isotomidae (so far only known in Proisotoma minima). This peculiarity remains the only difference between Pauropygus pacificus sp. n. and Pauropygus caussaneli. Taking into consideration also the different distribution ranges of these species we prefer to consider them as two independent species.
The species is probably distributed in the sands all over the Pacific coast of China.
Known from two distant localities on Pacific coast of China.
Examination of numerous species of the Cryptopygus complex revealed that six genera of this complex share the posterior position of sensilla on the three first abdominal segments. Like in other taxa of Isotomidae, as for instance in Proisotoma complex (Potapov et al., 2006) and in species groups of Folsomia (Grow & Christiansen, 1976;
To figure out the relation between these genera, we propose below a key to the genera of the Cryptopygus complex with posterior position of sensilla on three first abdominal segments. Pauropygus projectus sp. n. lost posterior sensilla and can be identified by pleural fold; the loss of mentioned sensilla is probably shared with several species of Arlea but descriptions do not give information on these characters. The position of the odd genus Appendisotoma is not fully decided: six species studied by us (Appendisotoma abiskoensis (Agrell), Appendisotoma bisetosa Martynova (types), Appendisotoma sibirica Stebaeva (types), and three species from Far East Russia and North America) have 11 apical chaetae on tibiotarsi while other four (Appendisotoma stebayevae (Grinbergs), Appendisotoma montana(Martynova), Appendisotoma juliannae (Traser et al.), and one species from Kazakhstan) show common set for the subfamily (7 chaetae). We include the genus Appendisotoma in the key since it formally matches the diagnosis of Cryptopygus complex.
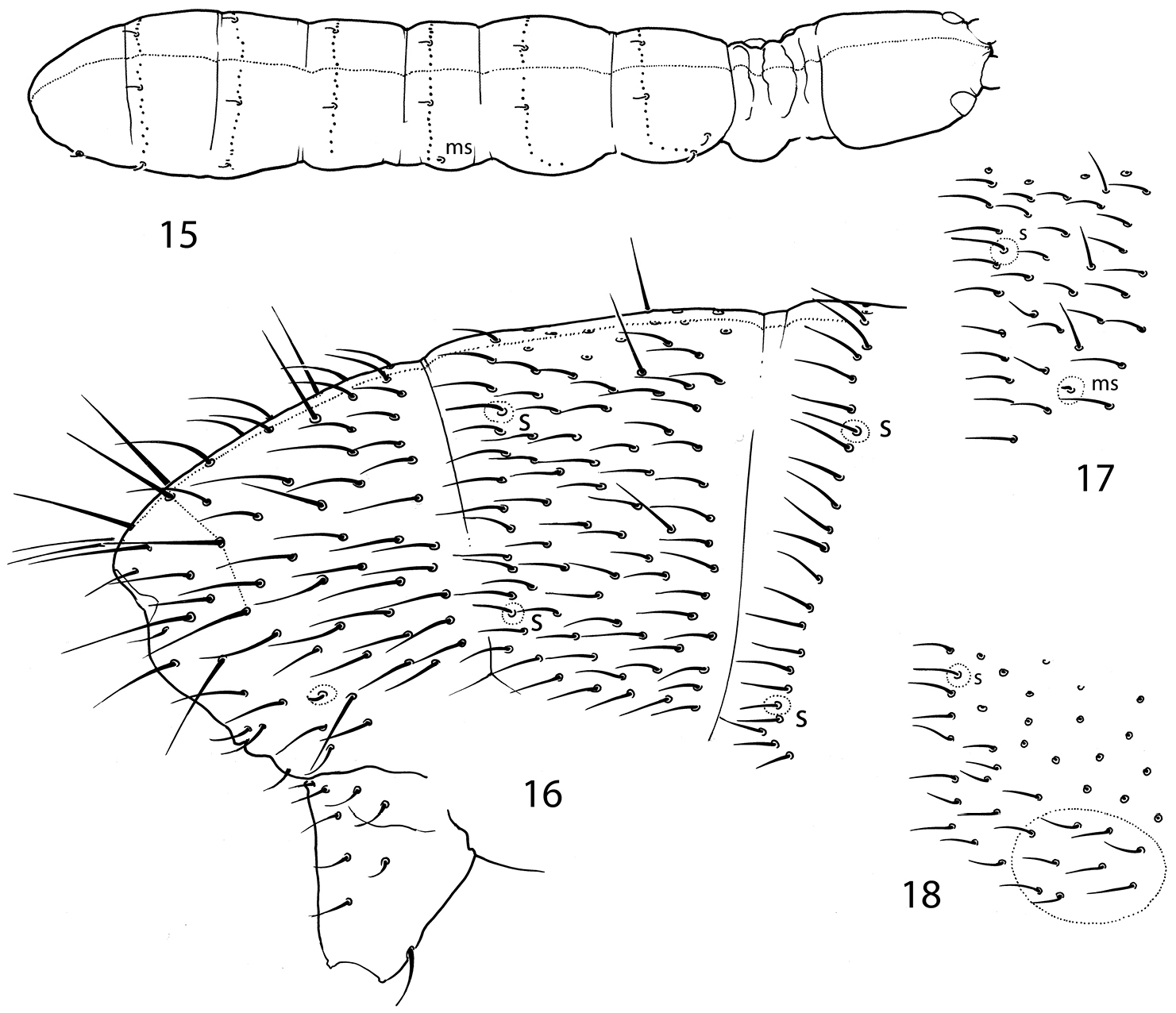
| 1 | At least 6+6 ommatidia on head. Sensilla on abdomen subequal, Abd.I - III and Abd.IV often with more than 3 and 4 sensilla on each side, respectively (Figs 25–26). Usually dark coloured | Appendisotoma Stach, 1947 |
| – | Usually blind, at most 2+2 ommatidia on head. Sensilla at the end of abdomen differentiated, Abd.I - III with no more than 2 sensilla on each side (Figs 27–29). White | 2 |
| 2 | Mucro falciform | Arlea Womersley, 1939 |
| – | Mucro bidentate | 3 |
| 3 | PAO complex, with lobes. Well-marked cylindrical foil-chaetae at the end of abdomen. Anterior medial sensilla on Abd.V present (Fig. 27) | Micrisotoma Bellinger, 1952 |
| – | PAO simple. Without foil-chaetae at the end of abdomen. Anterior medial sensilla on Abd.V absent (Figs 8, 28, 29) | 4 |
| 4 | Pleural fold on mouth cone with two finger-like processes (Fig. 20) | Pauropygus gen. n. |
| – | Pleural fold without such processes | 5 |
| 5 | Abd.IV with 3 sensilla on each side. Dorsal part of Abd.V with a pair of leaf-like sensilla (Fig. 28) | Proisotomodes Bagnall, 1949 |
| – | Abd.IV with 2 sensilla on each side. Dorsal part of Abd.V with two pairs of long and slender sensilla (Fig. 29) | Isotominella Delamare Deboutteville, 1948 |
The key is based on examination of three species of Pauropygus, four of Proisotomodes (one undescribed), two of Isotominella (one undescribed), eight of Appendisotoma (two undescribed), and one of Micrisotoma.
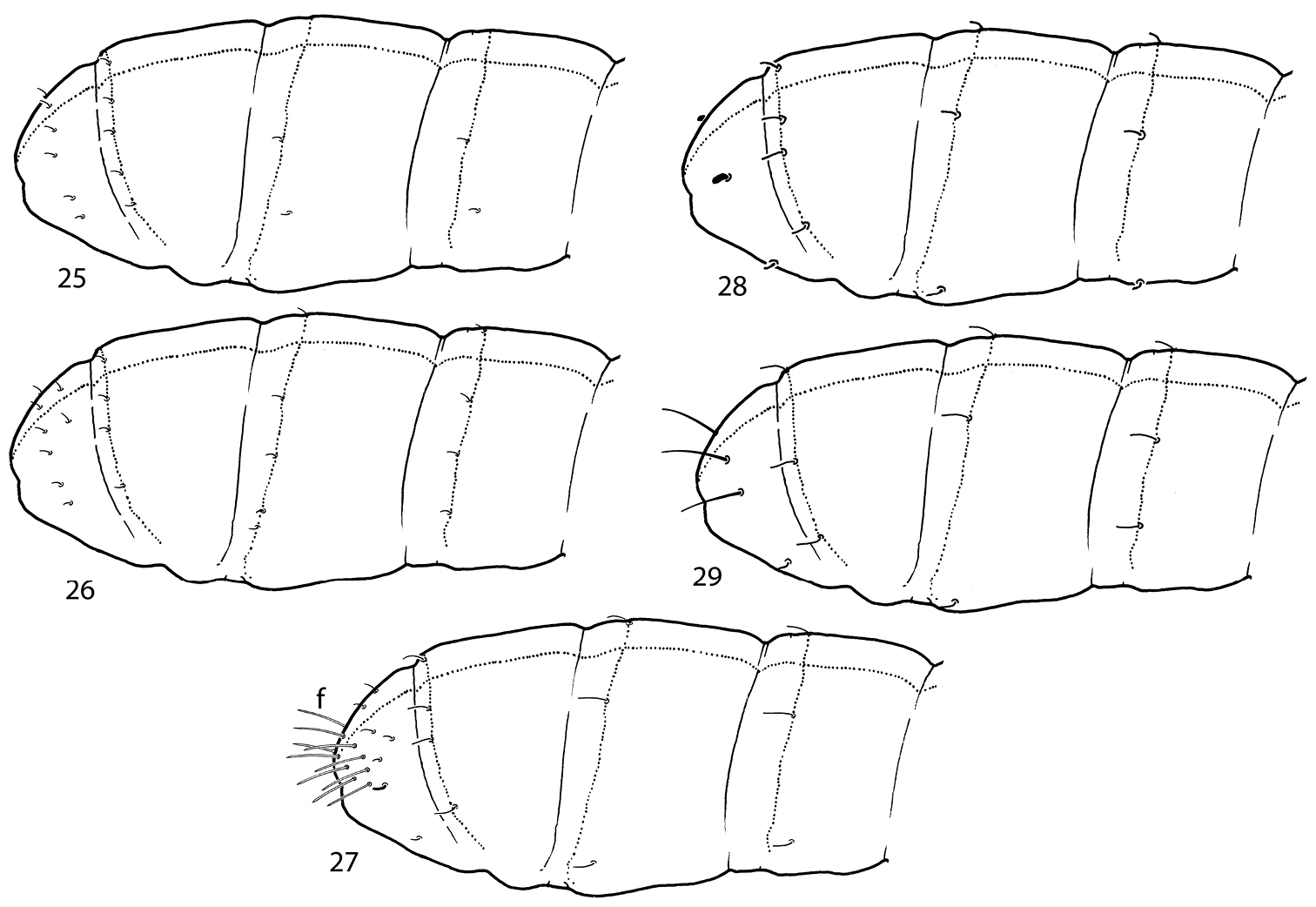
Sensillar chaetotaxy of genera of Cryptopygus complex with abdominal sensilla in posterior position. 25 Appendisotoma bisetosa Martynova, 1969 26 Appendisotoma abiskoensis (Agrell, 1939) 27 Micrisotoma achromata Bellinger, 1952 28 Proisotomodes bipunctatus (Axelson, 1903) 29 Isotominella geophila Delamare Deboutteville, 1948. f foil chaetae.
The work was supported by National Museum of Natural History (Paris). The field trips were partly financed by the National Natural Sciences Foundation of China (31071887) and NSFC-RFBR Cooperative Research Project (31111120077/11-04-91179-GFENa). We sincerely thank Y.X. Luan, Y. Bu, C.W. Huang (Shanghai) and N.A. Kuznetsova (Moscow) for their assistances during the collection of littoral Collembola in China. We also give our cordial thanks to Anne Bedos (Paris) for providing favorable working conditions, and to Jean-Marc Thibaud for access to his material from various countries. Arne Fjellberg and Lubomir Kovac significantly contributed to improve the quality of the manuscript.