





(C) 2013 Valentina Iannilli. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Ingolfiella arganoi sp. n. from Abd al Kuri Island in the Arabian Sea is described from two specimens, a male and a female. The western shore of the Indian Ocean was hitherto a vacant spot in the distribution of circumtropical shallow marine interstitial ingolfiellids and therefore the location of the new species fills a meaningful gap in the geography of the family. Morphologically, the new species shows close affinities with Ingolfiella xarifae from the Maldives.
Taxonomy, meiofauna, subterranean thalassoid amphipods, beach environment, mesopsammon, Arabian Sea
During sampling of aquatic fauna from the Socotra Archipelago conducted by Roberto Argano and co-workers (
The late Sandro Ruffo, curator of the natural history museum of Verona, started the work on the two specimens from Abd al Kuri, helped by one of us (V.I.), but did not finish it because he was not confident about the status of a new species based on so little material and the presence of only few available distinctive morphological characters. A few years later we decided to take up this work and bring it to fruition because it provides relevant new information on the geographic range of ingolfiellids and on new combinations of character states as shown by these specimens from an anchialine environment in the Arabian Sea.

a Abd al Kuri, beach, with gully in foreground, scoured in the sand by rainwater during the wet season (photograph taken while standing at the collection site by R. Argano). Pointed hills on the white beach sand are Ocypode crab burrows b position of Abd al Kuri Island (red color) in the Arabian Sea.
The two specimens were collected on a beach on the north coast of Abd al Kuri Island, Republic of Yemen, (12°11.988'N, 52°15.943'E), close to a little village called Bait Eesa. Sampling was done with a micro-creel in coarse sand at the bottom of a pool in an otherwise dry wadi, running as a slight depression to the sea (pers. comm. R. Argano). The approximate position of the pool with unknown salinity from which the ingolfiellids were collected on 7 February 2000 is shown in Fig. 1a.
urn:lsid:zoobank.org:act:807BB2C1-62F5-4930-A55B-74AF21C033EB
http://species-id.net/wiki/Ingolfiella_arganoi
Figs 2–5Two specimens: one male holotype, 1.4 mm, dissected and mounted in Faure’s liquid on slide MSNVRCr nr. 434; one preparatory female paratype on two slides MSNVRCr nr. 470 in Museo di Storia Naturale di Verona, Italy.
Lateral lobes on frontal margin of head developed. Maxillule, basal endite (= outer lobe) left and right with asymmetrical seta. Gnathopods 1 and 2 carpochelate with oblique palm, dactyli with a serrated inner margin with four teeth. Female with extra palmar margin robust seta. Oöstegites on pereiopod 3 and 4, with three regularly placed small button-like processes. Gills present on P3-5. Dactylus of P3 and P4 with slender trifid unguis; P5-7 with thicker bifid unguis, not clearly separated from dactylus. Pleopods 1-3 subtrapezoidal and similar, except first pleopod in male which is flexed and has a broadened tip. Uropod 1 with inner ramus about 1.5 times as long as outer ramus; uropod 2 peduncle without basofacial spine and with two diagonal rows of sturdy rectangular setae, three rows in female, individual setae mostly bifid but with some of them trifid at the tip.
The new species is named after Roberto Argano (University of Rome “la Sapienza”) who collected the specimens and gave them to the Verona Museum for study.
Body elongate, without coloration, all segments laterally compressed. Head (Fig. 2a) with lateral margin rounded; lateral or ‘ocular lobes’ present on frontal margin, well developed, suboval. Pleonites I-III with diffusely developed posteriorly rounded epimeral plates adorned with simple seta, a superficial marginal edge slightly visible. Urosomite III subcilindrical, slightly longer than deep, enclosing base of telson and uropod III.
Antennule (Fig. 2a), peduncular article slightly shorter than head; article ratio 1:0, 42:0, 42; flagellum of 4 articles, half the length, articles 2-4 with 1 aesthetasc; accessory flagellum slightly shorter than flagellar articles 1+2, three articles.
Antenna (Fig. 2a) subequal in length to antennule; flagellum of 5 articles, slightly shorter than half the length of peduncle, the last article bearing one aesthetasc (antenna drawn by S. Ruffo but not present in mounted slides).
Mandibles with non-triturative molar process, spiniform. Left mandible (Fig. 5d) with broad incisor, right mandible (Fig. 5c) with fine serrations on lacinia and molar process margin.
Maxillule (Figs 5a, e) coxal endite (= inner lobe) with 3 simple setae; basal endite (= outer lobe) with six robust setae of which the second one on the medial side has four teeth in the left maxillule and three teeth in the right one. Endopod (= palp) two-segmented, distal segment with two setae.
Maxilla (Fig. 5b) with short, equally long plates, each bearing four distal setae.
Maxilliped (Fig. 5f) basal endite slender, with one simple seta; ischium with two setae; merus and carpus without setae; propodus with one seta; dactylus with one lateral robust seta and distally two long setae, unguis not discernible.
Oöstegites on pereiopods III-IV (Figs 3d, e), suboval, without setae and with 3 button-like processes.
Coxal gills on pereiopods III-V.
Gnathopod I (Figs 2b, 3b) carpo-subchelate, palm strongly oblique, carpus 2.4 times as long as wide, palm margin smooth, not serrated, and with three short, bifid flagellate setae along lateral side of margin, and one simple seta on palm angle in male. In female two of such setae of which one placed closer to the row of three bifid setae. Just posterior to the palmar angle seta is a broad triangular spine on the medial side in the male, and three spines in the female: two smaller ones and a larger, more pointed one. Dactylus with four long spines along posterior margin and thin setules or grooves at the base of the unguis.
Gnathopod II (Figs 2c, d; 3c) Carpo-subchelate, palm oblique, carpus stronger than in gnathopod I, subtrapezoidal, carpal index = 4.6, palm angle defined by one large seta and one smaller spine in female (Fig. 3c), one seta in male (Figs 2c, d), and with triangular tooth proximal to the palmar angle seta, palm margin with irregular serrations; propodus strong with lobe on lateral side ending in a setule, less pronounced in female; dactylus with four strong teeth enforced with thick margins on lateral side and a groove or bundled setules at the base of the unguis.
Pereipods III-IV (Figs 3d, e) with two distal setae on dactylus at the base of the unguis, and three distal setae on propodus, one of them long and apically bifid, unguis apically trifid. Oöstegites with in both pereiopods regularly placed series of 3 button-like processes.
Pereiopods V – VII (Figs 4a, b, c) progressively longer towards P7; basis of P5 broad, that of P7 slender; carpus of P5 with two long and stout distal setae, others shorter; carpus of P7 with broad, curved and modified comb setae; merus of P7 with long distal seta; dactyli with two small setae distally; unguis bifid.
Pleopods I-III (Fig. 2a) subtrapezoidal, without setae. Pleopod I in male deformed or broadened distally.
Uropod I (Figs 2e, 4e) male: protopod with one seta and a row of fine setules on anterolateral margin; exopod with very feeble segment suture and one seta placed at two-thirds the length; endopod with terminal row of spines and four long setae laterally. In female protopod with three setae; endopod with six long setae laterally.
Uropod II (Figs 3a, 4d), protopod with two oblique comb rows in male, and three in female; setae of rows more or less rectangular with bifid or trifid, or even comb-like tips; endopod slightly longer than exopod, sharper, and with four setae.
Uropod III (Fig. 2a) short, 2 segmented, with one ramus, protopod with 2 distal setae, ramus short with 1 distal seta.
Telson (Fig. 2a) globose, with 1 pair of long dorsal setae.
Differences between male and female: gnathopods without extra palmar seta in male, and uropod II without a third comb row in male. Pleopod I in the male has a broadened tip.
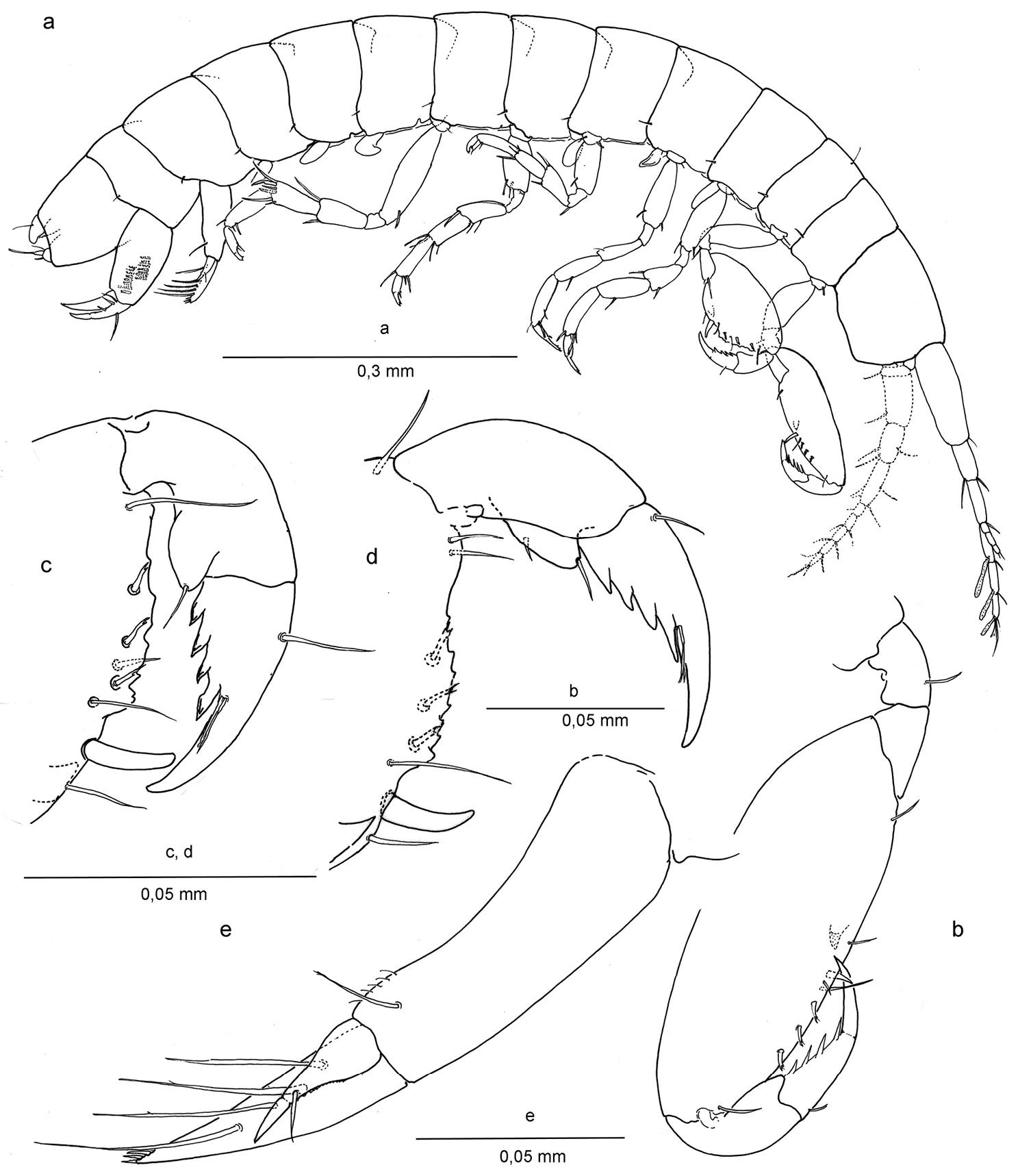
Ingolfiella arganoi sp. n., male holotype 1.4 mm. a habitus, male 1.4 mm b left gnathopod I, lateral c right gnathopod II, lateral d left gnathopod II, medial e right uropod I, lateral.
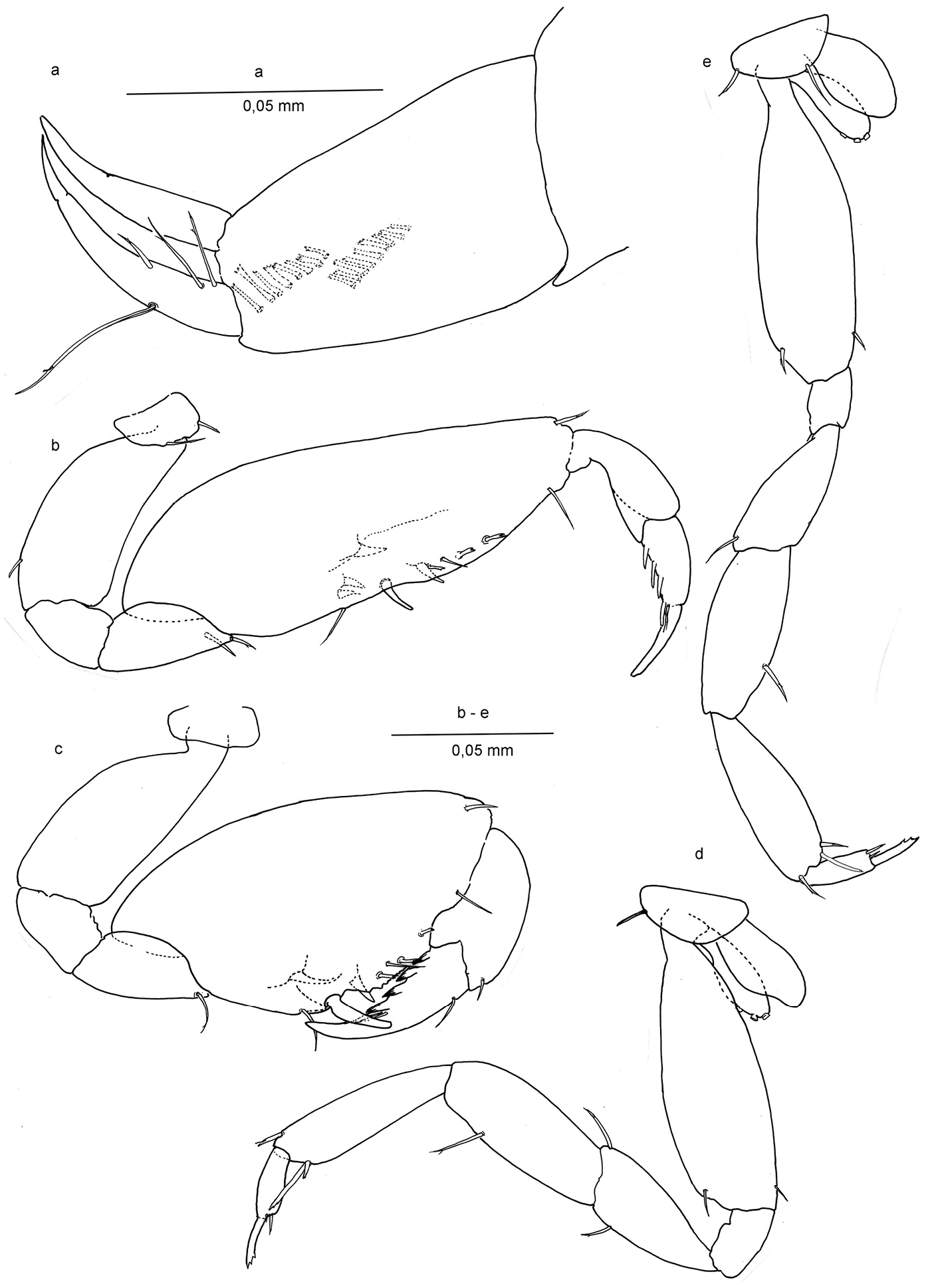
Ingolfiella arganoi sp. n., male holotype 1.4 mm. a right uropod II, lateral; female paratype 1.5 mm. b right gnathopod I, lateral c right gnathopod II, lateral d right pereiopod III, lateral e right pereiopod IV, lateral.
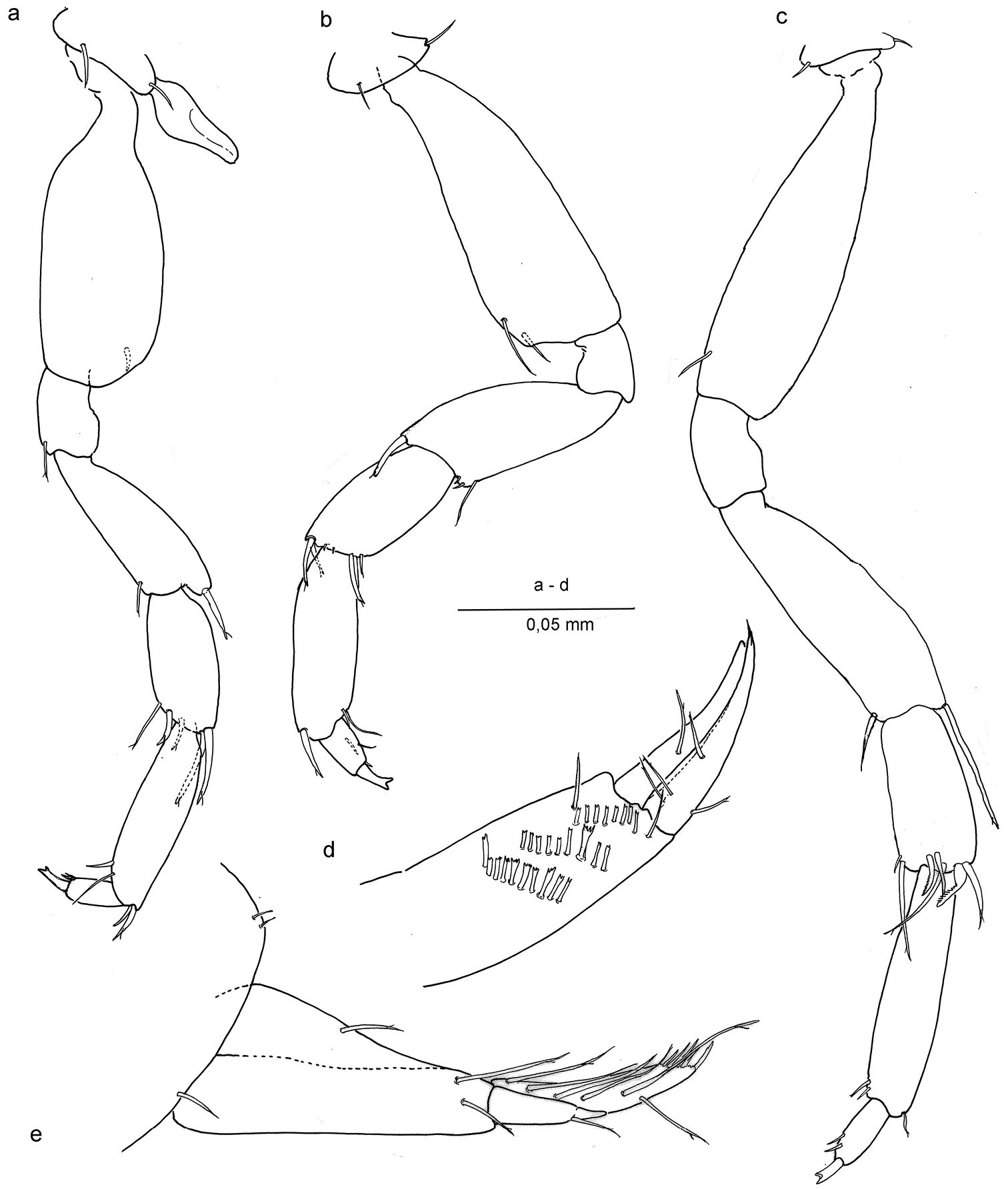
Ingolfiella arganoi sp. n., female paratype. a pereiopod V b pereiopod VI c pereiopod VII d right uropod II, medial e left uropod I, lateral.
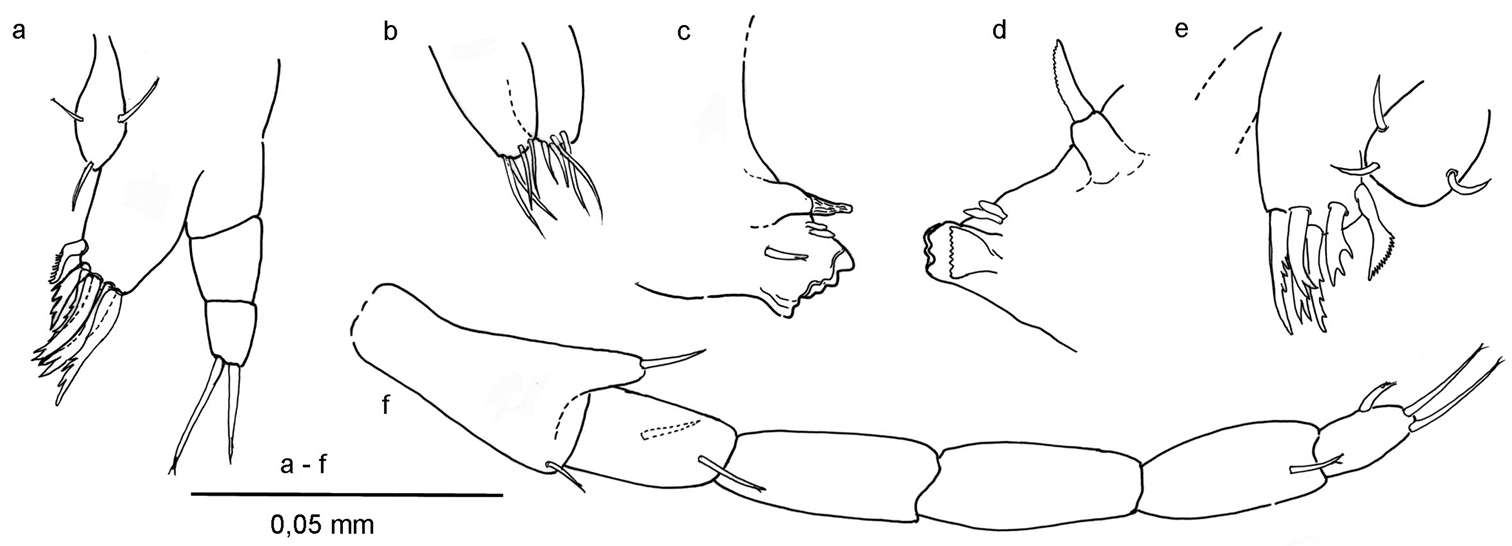
Ingolfiella arganoi sp. n., female paratype. a left maxillule b maxilla c right mandible d left mandible e right maxillule f right maxilliped.
Ingolfiella arganoi sp. n. shares most morphological character states with a species found 2500 km southeastward across the Indian Ocean, on the Maldives, namely Ingolfiella xarifae (Ruffo, 1966), from washed-out broken coral pieces (Favites sp.). Species ranges of stygobionts have not been reported to exceed such large distances in the past and molecular work on cryptic lineage diversity of populations of groundwater crustaceans have even diminished existing ranges to distances of less than 1000 km (
For now, five clear morphological differences justify the designation of a new species that also is geographically quite far away from its nearest congeners. These differences can be observed in the four spines on the medial margin of the propodus of gnathopod I – three spines in Ingolfiella xarifae; subtrapezoidal carpus of gnathopod 2 – elongate oval in xarifae; palmar index of gnathopod 2 is 4.6 – against 6.4 in xarifae; palm of gnathopod 2 strongly serrate – almost smooth in xarifae; pereiopod VII with specialized, combed, robust setae distally on carpus – not present in xarifae.
The differences in the placement, form and number of setae have shown to be quite consistent in the case of the poorly setose ingolfiellids (
The oöstegites have 3 small button-like processes. The same character was described in Ingolfiella alba Iannilli et al., 2008, where also three small button-like processes are present, but here from P3 to P5. Re-examination of Ingolfiella xarifae typus by Sandro Ruffo and V.I. allowed them to observe also on the oöstegites of this species the button-like processes, as described for Ingolfiella alba.Something similar was described in Metaingolfiella mirabilis Ruffo, 1969, although the processes were smaller and more numerous. These structures are probably present in other Ingolfiella species but have yet to be observed and described (
In other amphipods with preparatory females (females in a moult stage in between two brooding periods), these structures are sometimes present on the oöstegites (pers. comm. D. Jaume).
In agreement with
However, the species-groups in sensu
The present discovery represents the westernmost record of ingolfiellids in the Indo-Pacific Oceans. The Red Sea shores, Gulf of Aden, and the shores of the Arabian sea are relatively unknown areas for stygobiont and marine interstitial crustaceans. Geographically it is an interesting region as it forms the corridor between the better known Mediterranean and south east Asian/Australian stygofaunas. Sampling by
We thank Leonardo Latella and Roberta Salmaso (Museo Civico di Storia Naturale, Verona) for help and access to the crustacean collection; Traudl Krapp (Zoologisches Forschungsmuseum Alexander Koenig, Bonn) for her indispensable work on amphipod identifications and cataloguing in the Verona collection; Damiá Jaume (Institut Mediterrani d’Estudis Avanzats, Mallorca) for his comments on peracarid development and taxonomy; Bert Hoeksema (Naturalis Biodiversity Center, Leiden) for close reading; Kees van den Berg (Naturalis Biodiversity Center, Leiden) for new microscopy tools and assistance; and Roberto Argano (University La Sapienza, Rome) for donating his expedition specimens. We dedicate this study, in the warmest remembrance, to the late Sandro Ruffo. The work at the Verona museum by RV was supported by Naturalis bench fee grants of 2011 and 2012.