





(C) 2012 Gioele Tropea. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
A new species of the genus Euscorpius Thorell, 1876is described based on specimens collected from Dilek Peninsula (Davutlar, Aydın) in Turkey. It is characterized by an oligotrichous trichobothrial pattern (Pv= 7, et= 5/6, eb= 4) and small size. Euscorpius (Euscorpius) avcii sp. n. is the first named species of the subgenus Euscorpius from Turkey.
Scorpion, Euscorpius, new species, Turkey
The genus Euscorpius Thorell, 1876 is one of the most studied taxa of scorpions. According to the present taxonomy, it includes 18 species grouped in four subgenera (Alpiscorpius
A number of 79 specimens collected at Dilek Peninsula, in Turkey, were examined. Furthermore, 56 specimens from MZUF (Euscorpius tergestinus (C.L. Koch, 1837): 132/5856, 84/5847, 5848, 5861, 5862, 5863, 131/5838, 5839, 5840, 5841, 5842, 5843, 132/5854, 5856, 5857, 5860, 135/5699, 161/5850, 5851, 162/5864, 5865, 5866, 5867, 163/5987, 5988, 5889, 5990, 5991, 5992, 5993, 5994, 5995, 5996, 5997, 5998, 180/5852, 1417/5999, 6000, 6001, 6002, 6003, 6004, 6005, 6006, 6007, 6008, 6009, 6010, 6011, 6012, 165/6226, 73/6032, 1149/6238; Euscorpius oglasae Caporiacco, 1950 lectotype 122/5974, paralectotypes 123/5975) and 13 specimens of the private collection of Gioele Tropea (10 Euscorpius tergestinus (C.L. Koch, 1837) from Italy (Abruzzo, Latium and Umbria) and 3 Euscorpius carpathicus sensu stricto(Linnaeus, 1767) from Romania) were included in this study as comparison material.
Abbreviations: V: trichobothria on ventral pedipalp chela manus; Pv: trichobothria on patella ventral surface; Pe: trichobothria on the pedipalp patella external surface; et: external terminal; est: external sub-terminal; em: external medium; esb: external suprabasal; eba: external basal a; eb: external basal; DPS: dorsal patellar spur; DD: distal denticle; MD: median dentition; OD: outer dentition; ID: inner dentition; IAD: inner accessory denticles; AMNH: American Museum of Natural History, New York, USA; MZUF: Museo Zoologico ‘La Specola’ dell’Università di Firenze, Florence, Italy; GTC: private collection of Gioele Tropea; MTAS: Museum of the Turkish Arachnological Society; ZMSU: Zoology Museum of Sinop University; KUAM: Arachnological Museum of Kırıkkale University; ARC: private collection of Andrea Rossi.
The trichobothrial notations follow
urn:lsid:zoobank.org:act:B6799900-DF03-488A-B675-F0195AEB9825
Holotype: 1 ♂, Dilek Peninsula National Park, Canyon, Dilek Peninsula, near Davutlar Town, Kuşadası, Aydın, Turkey, 07.10.2005, leg. H. Koç (MTAS).
Paratypes: 1. 2 ♀♀, 3 ♂♂, Dilek Peninsula National Park, Canyon, Dilek Peninsula, near Davutlar Town, Kuşadası District, Aydın Province, Turkey, 06.11.2004, leg. H. Koç (ZMSU); 2 ♀♀, 1 sub♂, 3 sub♀, Dilek Peninsula National Park, Canyon, Dilek Peninsula, near Davutlar Town, Kuşadası District, Aydın Province, Turkey, 07.10.2005, leg. H. Koç (MZUF); same data but 1 ♂, 2 ♀ (GTC); 4 ♂♂, 3 ♀♀, Dilek Peninsula National Park, Canyon, Kuşadası District, Aydın Province, Turkey, 04.05.2011. 37°41'37"N, 27°09'37"E, 82 m, leg. E.A. Yağmur, A. Avcı and F. Yeşilyurt (MTAS); 5 ♂♂, 10 ♀♀, Dilek Peninsula National Park, Canyon, Dilek Peninsula, near Davutlar Town, Kuşadası District, Aydın Province, Turkey, 07.10.2005, leg. H. Koç (ZMSU); 3 ♀♀, Dilek Peninsula National Park, Canyon, Dilek Peninsula, near Davutlar Town, Kuşadası District, Aydın Province, Turkey, 18.06.2005, leg. H. Koç (ZMSU).
2. 3 ♂♂, 6 ♀♀, 5 km south of Güzelçamlı Village, Davutlar Town, Kuşadası District, Aydın Province, Turkey, 07.06.2011, 37°41'22"N, 27°13'31"E, 311 m, leg. F. Yeşilyurt and E.A. Yağmur (KUAM). Same data but 1 ♂, 1 ♀ (ARC). 3 ♂♂, 8 ♀♀. 5 km south of Güzelçamlı Village, Davutlar Town, Kuşadası District, Aydın Province, Turkey, 13.07.2010, 37°41'25"N, 27°13'53"E, 428 m, leg. F. Yeşilyurt and T. Danışman (KUAM).
3. 1 ♂, 8 ♀♀, Dilek Peninsula, 2 km south of Davutlar Town, pine forest, Kuşadası District, Aydın Province, Turkey, 02.07.2011, leg. E.A. Yağmur and A. Avcı (MTAS).
4. 6 ♂♂, 2 ♀♀, Dilek Peninsula National Park, picnic area, laurel forest, Kuşadası District, Aydın Province, Turkey, 13.08.2009, leg. E.A. Yağmur, N. Tezcan and V. Ülgezer (MTAS).
The specific epithet refers to Dr. Aziz Avcı who is a Turkish herpetologist and the new species is named after him for his kind contributions to collecting scorpion species and his friendship.
A small Euscorpius species, total length 24–28 mm. Color of adults is light brown to brown-reddish with the carapace and pedipalps darker brown-reddish, legs and telson lighter, yellowish colored. Euscorpius avcii sp. n. is oligotrichous; the number of trichobothria on the pedipalp manus ventral surface is 4 (3 V + Et 1); the number of trichobothria on the pedipalp patella ventral surface is 7 (of 78.5% of examined specimens and of 88% of pedipalps). The number of trichobothria on pedipalp patella external surface is: eb = 4, eba = 4, esb = 2, em = 4, est = 4, et = 5/6 (generally 5). The pectinal teeth count is: 7-9 (generally 8) in males, 6-7 (generally 7) in females. The telson vesicle in males is more swollen than in females, but only slightly more swollen if compared to other species of the subgenus Euscorpius. The pedipalps are stocky with a notch on fixed finger and scalloping of the movable finger well developed in adult males, obsolete in females. The dorsal patellar spur is weakly developed. Carinae on the metasomal segments are strongly reduced, almost smooth. Average value of the length from center median eyes to anterior margin of the carapace is equivalent to 39.20±2.0% of the carapace length. Average value of the length from center median eyes to posterior margin of the carapace is equivalent to 60.80±2.0% of the carapace length.
Coloration: Light brownish with carapace and pedipalps darker, brownish-reddish, legs, telson and chelicerae are lighter, yellowish-orange. Carapace slightly marbled. The coxal region is distinctly brownish-orange colored. The sternites, pectines and genital operculum are very light brownish-white (Fig. 3, 4 and 5).
Carapace: Length 3.70 mm; posterior width 3.75. Very slightly and finely granulated in laterally. All the furrows are shallow, only the posterior lateral furrows are slightly more marked. Distance from the center of the median eyes to the anterior margin of the carapace is equivalent to 39.62% of the prosoma; the length from the center of the median eyes to the posterior margin of the carapace is equivalent to 60.38% of the prosoma (Fig. 1A).
Mesosoma: Tergites veryslightly and finely granulated, almost smooth; sternites smooth. The area of overlap between the sternites is lighter in color. Pectinal teeth count is 8-9. The spiracles are very small, oval shaped and it is inclined to about 45° downwards towards outside.
Metasoma: Medium to small size with respect to body length. Dorsal carinae from segment I–IV are almost smooth, exhibit a few distanced fine granules, obsolete or almost obsolete on the segment V; ventromedian carinae from segment I–IV absent; ventromedian carinae on segment V are formed by very fine granules. Ventrolateral carinae from segment I–IV are obsolete; on segment V they are formed by a few spaced granules (Fig. 2E, F).
Telson:Vesicle weakly swollen (Fig. 2A); smooth, with ventral setae of different sizes; telson height 1.37, telson length 3.65, vesicle length 2.65, vesicle width 1.40.
Pectines: Pectinalteeth count 8-9; middle lamellae count 5-4.
Genital operculum: Partially divided with genital papillae protruding; a few microsetae present.
Sternum: Pentagonal shape, type 2. Length similar to width, deep posterior emargination.
Pedipalp: Coxa and trochanter with strong granulation. Femur: dorsal internal carinae tuberculate; dorsal external carinae formed by low spaced tubercles, their size increases from distal to proximal. Intercarinal spaces bears scattered small granules, larger in the posterior proximal area. Ventral external carina is granulated in the proximal half. External median carinae serrulate, anterior median crenulate and tuberculate distally. Patella length 3.25; patella width 1.20; dorsal internal carinae crenulate. Dorsal external carinae from rough to smooth and are crenulate proximally. Ventral external carinae from smooth to rough. Ventral internal carinae serrulate. Intercarinal tegument smooth or rough. Dorsal patellar spur weakly developed (Fig. 1E).
Chelal carina D1 isdistinctly strong, dark and from smooth to rough; D4 is formed from scattered granules; V1 isdistinctly strong, crenulate and dark; V3 is formed from granules on 2/3 of length.External carina with granules on distal half. Intercarinal tegument rough or smooth except between carinae D4 and V3. Movable finger dentition: MD like a straight line formed from very small denticles closely spaced and an DD on the distal tip; OD formed from 7 denticles on movable finger and 6 denticles on fixed finger, immediately outside of MD, their size increases progressively but the terminal denticle is not very pronounced; ID formed from 7 denticles on movable finger and 6 denticles on fixed finger, spaced from MD, their size increases progressively but the terminal denticle is not very pronounced; IAD on both movable and fixed finger formed from 4 small denticles.
Trichobothria: Chela trichobothria series V standard: V = 4-4 (3 V+ Et1); patella ventral (Pv): 8-7; Patella external (Pe): et = 5-5, est = 4-4, em = 4-4, esb = 2-2, eba = 4-4, eb = 4-4.
Legs: legs with two pedal spurs. Tarsal ventral row with 10-12 stout spinules; 3 tarsal setae flanked pairs adjacent to the ventral spinules row. Basitarsus with 6 prolateral stout spinules on leg pair I; 7 prolateral stout spinules on leg pair II; 1 prolateral stout spinules on leg pair III; absent on leg pair IV. Granulation on the leg femora II and III is more marked both dorsally and ventrally, and only ventrally on leg I. Granulation is formed from dark granules; while the granulation on the dorsal surface of the femur of leg I and on the femur of leg IV both dorsally and ventrally is weakly marked and of lighter colored granules.
Chelicerae: movable finger: The dorsal distal tooth is smaller than the ventral distal tooth; Ventral edge is smooth with brush-like setae on the inner part; dorsal edge has five teeth: one distal, two small subdistal, one big median and a small basal; fixed finger has four teeth: one distal, one subdistal, one median and one basal. The median and the basal are in a fork arrangement. The internal edge has brush-like setae.
Variation: The variation observed in 79 studied specimens (29 males, 50 females) is the follows: pectinal teeth in males: 7-7 (1/29), 8-8 (23/29), 8-9 (4/29), 9-9 (1/29); females: 6-6 (5/50), 6-7 (11/50), 7-7 (34/50); pedipalp patella trichobothria Pv: 8-8 (2/79), 8-7 (9/79), 7-7 (62/79), 6-7 (6/79); pedipalp patella trichobothria Pe: et = 5-5(41/79), 5-6 (19/79), 6-6 (19/79); est = 4-4 (79/79), em = 4-4 (79/79), esb= 2-2 (79/79), eba = 4-4 (79/79), eb = 4-4 (79/79). The variation in the trichobothrial pattern is within the standard values of variability and shows the stability of diagnostic characters.
Well developed lamina with well visible basal constriction, tapered distally; truncal flexure present and well developed; capsular lobe complex well developed, with acuminate process; ental channel spinose distally, exhibiting six delicate variable sized spines (Fig. 6).
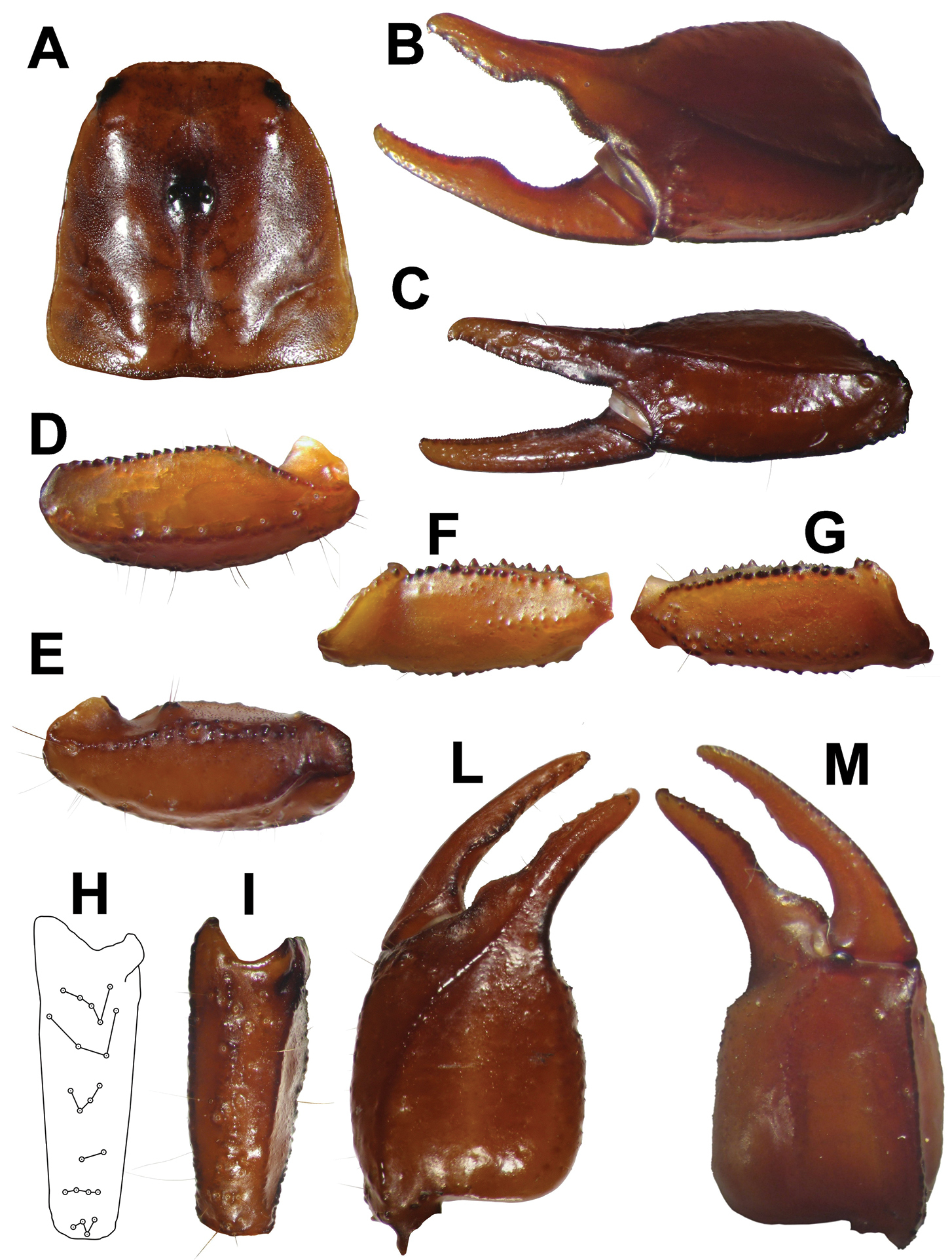
A carapace B external view of chela of the adult male C external view oh chela of the adult female D ventral view of pedipalp patella E dorsal view of pedipalp patella F ventral view of pedipalp femur G dorsal view of pedipalp femur H schematic view of trichobothrial pattern on external surface of pedipalp patella I view of external surface of pedipalp patella L dorsal view of chela M ventral view of chela.

A telson of adult male B telson of adult female C sternopectinal area of adult male D sternopectinal area of adult female E latero-dorsal view of the metasomal segment V F ventral view of the metasomal segment V G tarsus and basitarsus H leg femur I I leg femur II L leg femur III M leg femur IV.

Dorsal and ventral views of Euscorpius avcii sp. n.male.

Dorsal and ventral views of Euscorpius avcii sp. n.female.

Euscorpius avcii sp. n. in its natural habitat.
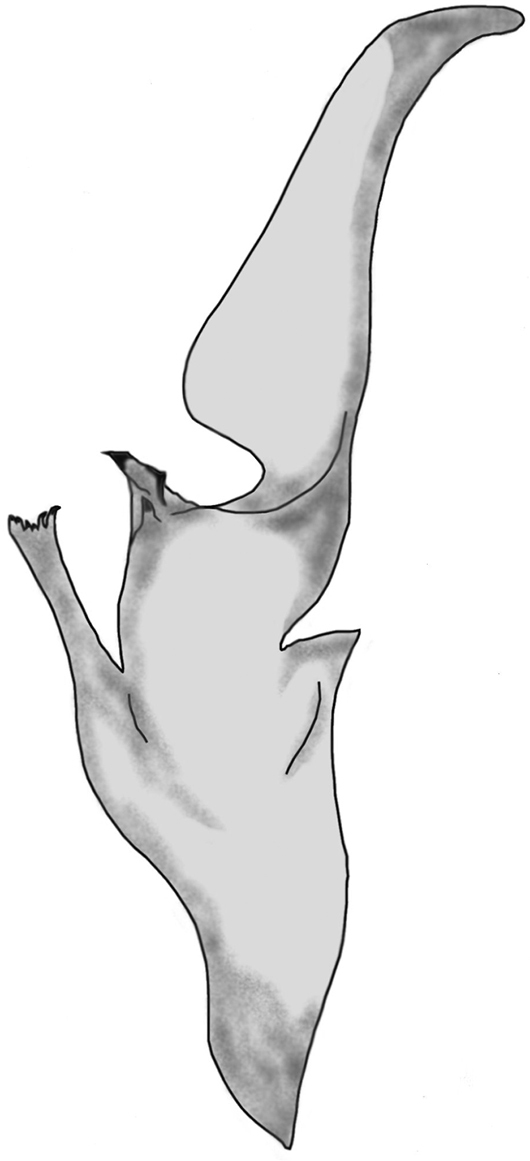
Left hemispermatophore of Euscorpius avcii sp. n.
Euscorpius avcii sp. n. is an oligotrichous form with Pv = 7 and Pe-et = 5/6. Most of the species belonging to the subgenus Euscorpius have generally higher trichobothrial numbers, with some exceptions e.g. Euscorpius oglasae Di Caporiacco, 1950 (Pv = 7, Pe-et = 5) (
Euscorpius oglasae has a trichobothrial pattern that is almost identical to Euscorpius avcii sp. n., but the morphology and geographic distribution (Euscorpius oglasae is endemic to the island of Montecristo in the Tyrrhenian Sea in Tuscany, Italy) make easy to separate these two species. Euscorpius oglasae is larger than Euscorpius avcii sp. n. (up to 43 mm) (
Samos is a Greek island inhabited by an unnamed oligotrichous form, similar to Euscorpius avcii sp. n. The Samos population is characterized by small size, stocky pedipalps and trichobothrial pattern Pv = 5 and et = 5 (
Euscorpius tergestinus s. str. is easily distinguished from Euscorpius avcii sp. n., even if the color and the trichobothrial pattern eb = 4, eba = 4, em = 4 may suggest that Euscorpius avcii sp. n. is a species belonging to the “tergestinus complex”, but these are the only similar characters, in fact the morphology and the chaetotaxy reveal the great differences between these two species. Euscorpius tergestinus is larger in size (30-40 mm), it has a slender habitus with elongated pedipalps and DPS strongly developed, among the largest in the entire genus Euscorpius.
Its telson is very swollen, above average in both male and female. The metasomal carinae are much more pronounced, granulated and Euscorpius avcii sp. n. has a less swollen telson and the metasomal carinae almost smooth. The pedipalpal chela of Euscorpius tergestinus is slender and long, especially the fingers. In this species, trichobothrium db on the fixed finger is much more distal than in Euscorpius avcii sp. n. that has it in proximal position.
Euscorpius tergestinus has a more granulated carapace, and body, and developed furrows on the carapace whereas Euscorpius avcii sp. n. has almost smooth carapace, and body, with weak furrows, causing the appearance of a fairly flat carapace. The trichobothrial pattern of the pedipalp patella of Euscorpius tergestinus is reported as Pv = 7/11 (9), Pe-et = 5/8 (6 +) in
Other species and subspecies of subgenus Euscorpius s.str. that are relatively geographically close, from the Aegean area: Euscorpius sicanus (C. L. Koch, 1837), Euscorpius koshewnikowi Birula, 1900, Euscorpius carpathicus candiota Birula, 1903, Euscorpius carpathicus ossae Di Caporiacco, 1950, Euscorpius carpathicus aegaeus Di Caporiacco, 1950 and Euscorpius carpathicus scaber Birula, 1900. Euscorpius sicanus has never been reported in Turkey; furthermore, it is easy to separate because of its particular trichobothrial pattern; Pe: eb=5 and eba = 4/5 (
Euscorpius carpathicus scaber is a scorpion from the northern Aegean area, has a dark coloration with a high number of pectinal teeth, a higher trichobothrial pattern, and in addition, its whole body is covered with granules of various size, as also the name suggests, whereas Euscorpius avcii sp. n. has a light coloration, and its granulation is very little accentuated, almost smooth.
In our opinion, Euscorpius avcii sp. n. is well divided from all described Euscorpius forms including those that await taxonomic clarification. At present there are no described species or subspecies that corresponds to the morphology and to the trichobothrial pattern of this new species. We are confident that these data are enough to describe this form as a new species of the genus Euscorpius, and the first described species of the subgenus Euscorpius in Turkey.
Measurements (in mm) of male holotype and female and male paratype of Euscorpius avcii sp. n.
| Holotype | Paratype female | Paratype male | ||
|---|---|---|---|---|
| Total | Length | 26.18 | 23.65 | 27.70 |
| Carapace | Length | 3.70 | 3.60 | 3.90 |
| Posterior width | 3.75 | 3.70 | 3.80 | |
| Metasoma | Length | 9.78 | 8.20 | 10.10 |
| Segment I | Length | 1.25 | 1.10 | 1.30 |
| Width | 1.40 | 1.30 | 1.40 | |
| Segment II | Length | 1.50 | 1.35 | 1.60 |
| Width | 1.20 | 1.10 | 1.20 | |
| Segment III | Length | 1.75 | 1.45 | 1.80 |
| Width | 1.15 | 1.05 | 1.10 | |
| Segment IV | Length | 2.05 | 1.70 | 2.20 |
| Width | 1.07 | 1.00 | 1.05 | |
| Segment V | Length | 3.23 | 2.60 | 3.20 |
| Width | 1.10 | 1.00 | 1.10 | |
| Telson | Length | 3.65 | 2.85 | 3.70 |
| Vesicle | Length | 2.65 | 2.10 | 2.80 |
| Width | 1.40 | 0.75 | 1.40 | |
| Height | 1.37 | 0.95 | 1.40 | |
| Aculeus | Length | 1.00 | 0.75 | 0.90 |
| Femur | Length | 3.20 | 3.10 | 3.10 |
| Width | 1.25 | 1.20 | 1.20 | |
| Patella | Length | 3.25 | 3.20 | 3.45 |
| Width | 1.20 | 1.25 | 1.30 | |
| Chela | Length | 6.70 | 6.35 | 7.05 |
| Width | 3.00 | 2.70 | 3.05 | |
| Movable finger | Length | 3.85 | 3.10 | 4.00 |
| Pectines teeth | 8–9 | 7–7 | 8–8 |
Trichobothrial counts of Euscorpius species discussed in this paper.
| Species | vP | tPe - e | tPe - es | mPe - e | bPe - es | aPe - eb | bPe - e |
|---|---|---|---|---|---|---|---|
| Euscorpius avcii sp. n. | 7 | 5–6 | 4 | 4 | 2 | 4 | 4 |
| Euscorpius oglasae | 7 | 5 | 4 | 4 | 2 | 4 | 4 |
| Euscorpius koschewnikowi | 8 | 5–6 | 4 | 4 | 2 | 4 | 4 |
| Euscorpius sp. from“Samos” | 5 | 5 | 4 | 4 | 2 | 4 | 4 |
| Euscorpius carpathicus aegaeus | 7–8(8) | 5/6(6) | 4 | 4 | 2 | 4 | 4 |
| Euscorpius carpathicus ossae | 6–8(7/8) | 5 | 4 | 4 | 2 | 4 | 4 |
| Euscorpius carpathicus scaber | 7–10(8/9) | 6 | 4 | 4 | 2 | 4 | 4 |
| Euscorpius carpathicus candiota | 9–10 | 6–7 | 4 | 4 | 2 | 4 | 4 |
| Euscorpius tergestinus | 8–11(9) | 6–8(6) | 4 | 4 | 2 | 4 | 4 |
| Euscorpius carpathicus s.str. | 7–9 (8) | 5–7 (7) | 4 | 3 | 2 | 4 | 4 |
Specimens of Euscorpius avcii sp. n. were collected from the northern side of Dilek Peninsula (Fig. 7). Vegetation in this area is composed of both deciduous forest (Quercus cerris, Tilia rubra subsp. caucasica, Tilia argentea and Castanea sativa) and evergreen forest (which are Pinus brutia and Laurus nobilis). Coastal areas include scrub vegetation. Furthermore, northern side of Dilek Peninsula has a humid climate and in both summer and winter, flowing streams and wetlands exist. Specimens of Euscorpius avcii sp. n. were collected during the day under bark of decomposed wood, under stones and in rock crevices and at night with UV light from rocky places, roadsides and under pine forests (Fig. 8 and 9). Euscorpius avcii sp. n. specimens are sympatric with Mesobuthus gibbosus Brullé, 1832 and Iurus kinzelbachi Kovarik, Fet, Soleglad, Yagmur, 2010. We report an example of intraguild predation, we witnessed Mesobuthus gibbosus feeding on Euscorpius avcii sp. n. during one of our night trips (Fig. 10).
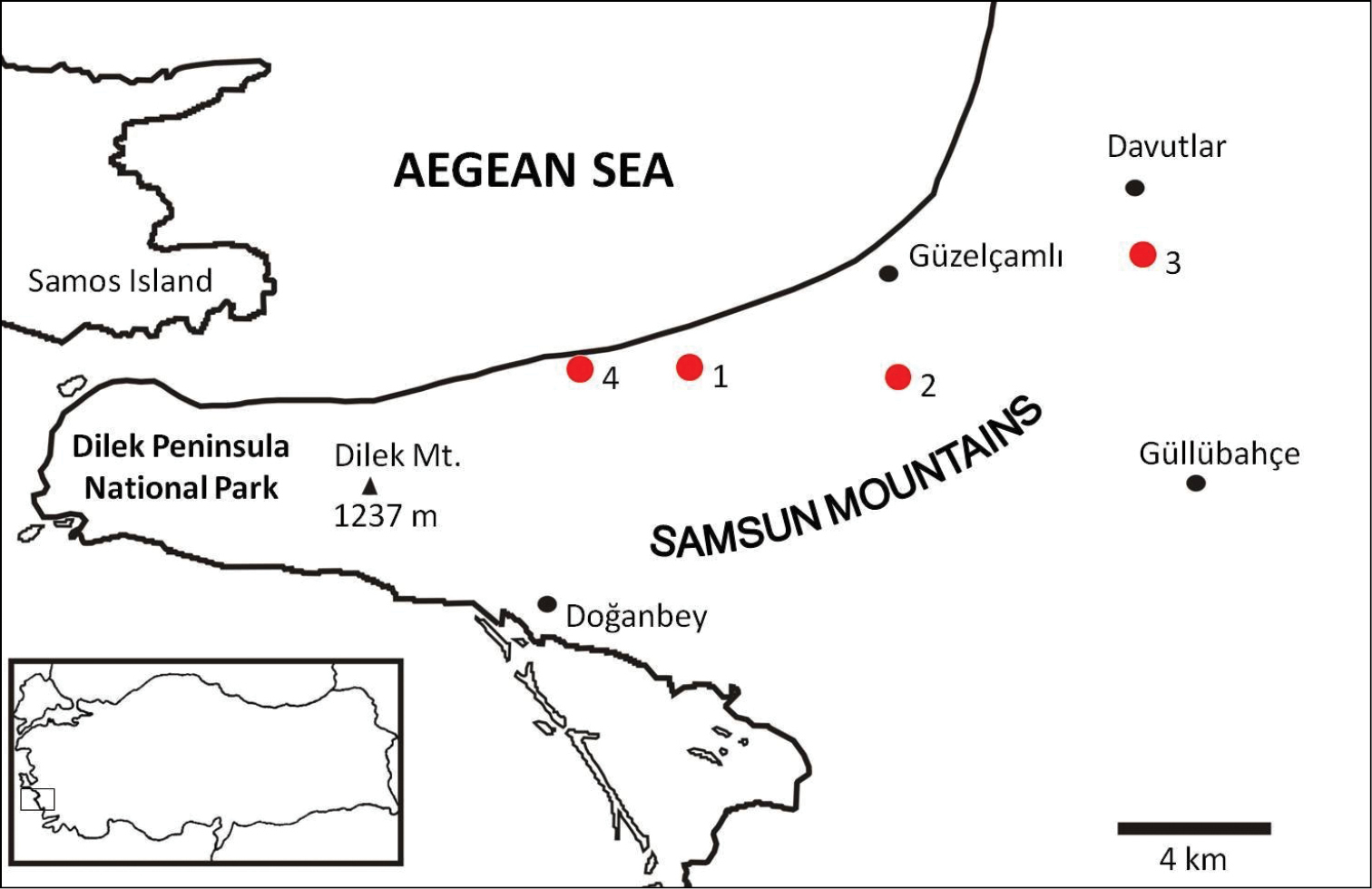
Map of Dilek Peninsula National Park

The habitat in Canyon in Dilek Peninsula National Park

The habitat in North of Güzelçamlı in Dilek Peninsula.

A Mesobuthus gibbosus which feeds on Euscorpius avcii sp. n.
We wish to thank Dr. Aziz Avcı, Dr. Tarık Danışman, Nurullah Tezcan and Volkan Ülgezer for their help during field trips and also Dr. Luca Bartolozzi, Dr. Sara Whitman, Dr. Jan Ove Rein, Mauro Domenici and Dr. Voltolino Voltolini for their kindness and for the material they provided. We would like to thank Dr. Michiel Cozijn and two anonymous reviewers for reviewing the English text and for their suggestions.