Research Article |
|
Corresponding author: Madhava Meegaskumbura ( madhava_m@mac.com ) Academic editor: Sven Kullander
© 2019 Hiranya Sudasinghe, Rohan Pethiyagoda, Kalana Maduwage, Madhava Meegaskumbura.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Sudasinghe H, Pethiyagoda R, Maduwage K, Meegaskumbura M (2019) The identity of the Sri Lankan Amblypharyngodon (Teleostei, Cyprinidae). ZooKeys 820: 25-49. https://doi.org/10.3897/zookeys.820.29632
|
Abstract
Morphological and molecular analyses of specimens representative of the geographic range of the cyprinid genus Amblypharyngodon in Sri Lanka suggest the presence of only a single species in the island, for which the name Amblypharyngodon grandisquamis Jordan & Starks, 1917, is available. Amblypharyngodon grandisquamis is a species endemic to Sri Lanka, distributed across the lowlands of both of the island’s main climatic zones. It is distinguished from all other species of Amblypharyngodon, including the three species recorded from peninsular India (A. mola, A. microlepis, and A. melettinus), by a suite of characters that includes a body depth of 26.9–31.2% of the standard length (SL), 42–56 scales in the lateral series (of which usually 8–16 are pored), 20–24 circumpeduncular scales, 14–17 scale rows between the origins of the dorsal and pelvic fins, a dorsal-fin height of 21.1–27.6% SL, 18–19 caudal vertebrae and an eye diameter of 22.7–30.5% of the head length. Amblypharyngodon grandisquamis differs from A. melettinus and A. mola by uncorrected pairwise genetic distances of more than 9% and 6%, respectively, for the mitochondrial cytochrome oxidase subunit 1 (COI) gene.
Keywords
Amblypharyngodon grandisquamis , Amblypharyngodon melettinus , biodiversity hotspot, DNA barcoding, integrative taxonomy, island endemism
Introduction
The cyprinid genus Amblypharyngodon Bleeker, which has a range extending from the Indian subcontinent to Southeast Asia, is considered to contain just five valid species: A. atkinsonii (Blyth), A. chulabhornae Vidthayanon & Kottelat, A. melettinus (Valenciennes), A. microlepis (Bleeker), and A. mola (Hamilton) (
The genus was first reported from Sri Lanka by
Here we review Sri Lankan Amblypharyngodon using an integrative taxonomic approach (
Materials and methods
Metrics and meristics
Measurements and counts follow
Comparative data for A. chulabhornae and (in part) A. atkinsonii and A. microlepis are from
Material
Specimens referred to in the text are deposited in the collections of Muséum national d’Histoire naturelle, Paris, France (
Comparative material
Amblypharyngodon melettinus (all from India):
A. cf. mola:
A. mola: WHT30316, 5, 43.2–47.5 mm SL, India, 24 Parganas, West Bengal.
A. microlepis:
A. atkinsonii:
Morphometric analysis
A Principal Component Analysis (PCA) in a covariance matrix was carried out to identify variables that best discriminate among the species. In view of one of us (HS) having measured the recent material and another (RP) having measured the historical material (the types of Leuciscus melettinus and Amblypharyngodon grandisquamis), and in order to account for the deforming of specimens through long-term storage in preservative, the respective datasets were analysed separately. Prior to the PCA, all measurements were size-corrected by using the equation
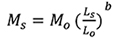
where Ms is the size corrected measurement, Mo is the measured character, Ls is the overall (arithmetic) mean standard length for all individuals from all populations of all putative species, and Lo is the standard length of the specimen, while b is calculated for each character from the observed data by using the allometric-growth equation M = aLb in which b is the gradient of regression of log Mo on log Lo (
Molecular analysis
The protocols for DNA extraction, PCR amplification and PCR product purification for the mitochondrial cytochrome oxidase subunit 1 (COI) and 16s rRNA subunit (16s) follow
Sequences were checked and assembled in ChromasPro v1.34 (Technelysium Pty Ltd) and contig sequences of the two strands were constructed using MEGA v.7.0 (
Species included in the phylogenetic analysis, with their localities, voucher references, and GenBank accession numbers.
| Species | Voucher | Location | GPS coordinates | Source | 16s | COI | cytb |
|---|---|---|---|---|---|---|---|
| Amblypharyngodon grandisquamis | DZ 1504 | Sri Lanka: Mahaweli River basin, Ulhitiya | 07.38N 81.09E | This study | MH884614 | MH884634 | MH884638 |
| DZ 3175 | Sri Lanka: Mahaweli River basin, Polonnaruwa | 07.87N 80.96E | This study | MH884615 | MH884633 | MH884639 | |
| DZ 3176 | Sri Lanka: Mahaweli River basin, Polonnaruwa | 07.87N 80.96E | This study | MH884616 | MH884632 | MH884643 | |
| DZ 3434 | Sri Lanka: Mahaweli River basin, Naula, Matale | 07.52N 80.64E | This study | MH884617 | MH884628 | NA | |
| WHT 81 | Sri Lanka: Mahaweli River basin, Elahera | NA | This study | MH884618 | NA | MH884645 | |
| WHT 92 | Sri Lanka: Mahaweli River basin, Minneriya | NA | This study | MH884619 | NA | MH884646 | |
| DZ 3376 | Sri Lanka: Walawe River basin, Samanala reservoir | 06.69N 80.78E | This study | MH884620 | MH884630 | MH884640 | |
| DZ 3377 | Sri Lanka: Walawe River basin, Samanala reservoir | 06.69N 80.78E | This study | MH884621 | MH884629 | MH884641 | |
| DZ 3292 | Sri Lanka: Kalu River basin, Remuna, Horana | 06.69N 80.07E | This study | MH884622 | MH884631 | MH884642 | |
| DZ 3854 | Sri Lanka: Attanagalu Oya basin, Yakkala | 07.07N 80.07E | This study | MH884623 | MH884637 | NA | |
| DZ 3855 | Sri Lanka: Attanagalu Oya basin, Yakkala | 07.07N 80.07E | This study | MH884624 | MH884636 | NA | |
| DZ 3856 | Sri Lanka: Attanagalu Oya basin, Yakkala | 07.07N 80.07E | This study | MH884625 | MH884635 | NA | |
| WHT 38 | Sri Lanka: Menik River basin | NA | This study | MH884626 | NA | MH884644 | |
| WHT 101 | Sri Lanka: Kala Oya basin, Nocchiyagama | NA | This study | MH884627 | NA | MH884647 | |
| Amblypharyngodon melettinus | Am2 | India: Kerala | NA | GenBank | FJ751268 | NA | NA |
| Am1 | India: Kerala | NA | GenBank | FJ751267 | FJ751272 | NA | |
| ACI | India: Kerala | NA | GenBank | FJ751266 | FJ751271 | NA | |
| A2 | India: Kerala | NA | GenBank | FJ751265 | FJ751270 | NA | |
| A1 | India: Kerala | NA | GenBank | FJ751264 | FJ751269 | NA | |
| Amblypharyngodon mola | NBFGRMU AM8004L | India | 23.32N 91.28E | GenBank | KT878048 | NA | NA |
| NBFGRMU AM8004K | India | 23.32N 91.28E | GenBank | KT878047 | NA | NA | |
| NBFGRMU AM8004H | India | 23.32N 91.28E | GenBank | KT878046 | NA | NA | |
| NBFGRMU 8004S | India | 24.39N 93.58E | GenBank | KT878045 | NA | NA | |
| NBFGRMU 8004M | India | 24.30N 93.46E | GenBank | KT878044 | KT896674 | NA | |
| PUMNH 28/2013 | India | NA | GenBank | NA | KX266827 | NA | |
| 8004Q | India | 24.34N 93.46E | GenBank | NA | KX245049 | NA | |
| 8004P | India | 24.34N 93.46E | GenBank | NA | KX245048 | NA | |
| 8004o | India | 24.34N 93.46E | GenBank | NA | KX245047 | NA | |
| 8004N | India | 24.34N 93.46E | GenBank | NA | KX245046 | NA | |
| DUZM031 | Bangladesh: Tanguar Haor, Sunamganj | 25.08N 91.33E | GenBank | NA | KT364774 | NA | |
| AM-1005 | India | 25.76N 89.95E | GenBank | NA | KJ936819 | NA | |
| AM-1004 | India | 25.76N 89.95E | GenBank | NA | KJ936818 | NA | |
| AM-1003 | India | 25.76N 89.95E | GenBank | NA | KJ936817 | NA | |
| AM-1002 | India | 25.76N 89.95E | GenBank | NA | KJ936816 | NA | |
| Amblypharyngodon mola | AM-1001 | India | 25.76N 89.95E | GenBank | NA | KJ936815 | NA |
| AM-2005 | India | 24.29N 92.63E | GenBank | NA | KJ936763 | NA | |
| AM-2004 | India | 24.29N 92.63E | GenBank | NA | KJ936762 | NA | |
| AM-2003 | India | 24.29N 92.63E | GenBank | NA | KJ936761 | NA | |
| AM-2002 | India | 24.29N 92.63E | GenBank | NA | KJ936760 | NA | |
| AM-2001 | India | 24.29N 92.63E | GenBank | NA | KJ936759 | NA | |
| NF770 | India: Gujarat, Bharuch, Estuary, Bharuch | 21.68N 73.45E | GenBank | NA | JX983212 | NA | |
| NF771 | India: Gujarat, Bharuch, Estuary, Bharuch | 21.68N 73.45E | GenBank | NA | JX983211 | NA | |
| PRUM01 | India: Maharashtra, Vidarbha, Umarkhed Painganga | 19.59N 77.68E | GenBank | NA | JX260822 | NA | |
| GF675 | India: Maharashtra, Marathwada, Kaigaon | 19.62N 75.01E | GenBank | NA | JX260821 | NA | |
| BESH02 | India: Maharashtra, Shahagad River Pool Cad | 19.36N 75.71E | GenBank | NA | JX260820 | NA | |
| PSSH02 | India: Maharashtra, Shahagad River Pool Cad | 19.36N 75.71E | GenBank | NA | JX260819 | NA | |
| GF679 | India: Maharashtra, Marathwada, Kaigaon | 19.62N 75.01E | GenBank | NA | JX260818 | NA | |
| GDK025-11 | India: Maharashtra, Shahagad River Pool Cad | 19.36N 75.71E | GenBank | NA | JX260817 | NA | |
| DOF16 | India | 24.46N 93.01E | GenBank | NA | JN815278 | NA | |
| DOF15 | India | 24.46N 93.01E | GenBank | NA | JN815277 | NA | |
| CTOL01907 | NA | NA | GenBank | NA | HM224137 | HM224256 | |
| AM2 | India | NA | GenBank | NA | NA | KX389702 | |
| AM1 | India | NA | GenBank | NA | NA | KX389701 | |
| Amblypharyngodon chulabhornae | NA | NA | NA | GenBank | AP012114 | AP012114 | AP012114 |
| NA | NA | NA | GenBank | U21380 | NA | NA | |
| CTOL01544 | NA | NA | GenBank | NA | NA | HM224255 | |
| Barilius vagra | CTOL03301 | NA | NA | GenBank | NA | HM224140 | HM224259 |
| Barilius gatensis | 4BG | India | NA | GenBank | NA | KF853168 | NA |
| 6BG | India | NA | GenBank | KF853144 | NA | NA | |
| 5BG | India | NA | GenBank | KF853143 | NA | NA | |
| Salmostoma phulo | CTOL03316 | NA | NA | GenBank | NA | HM224248 | HM224379 |
| Salmostoma bacaila | WL-F38 | India | NA | GenBank | NA | EU417789 | NA |
| NBFGRMU 8020G | India | 24.42N 93.04E | GenBank | KT878279 | NA | NA | |
| NBFGRMU 8020F | India | 24.42N 93.04E | GenBank | KT878278 | NA | NA |
Haplotype network reconstruction for the 16s, COI, and cytb genes of the various populations of Amblypharyngodon was inferred by TCS network (
Results
Molecular analysis
The HKY+G model was chosen as the best-fit nucleotide-substitution model under BIC in the jModelTest for the COI dataset. In the Bayesian phylogenetic inference for the COI dataset, A. mola was recovered as the sister species of A. grandisquamis, though weakly supported (PP = 66, Figure

A, C Phylogram based on Bayesian phylogenetic inference of A the COI and C16s dataset for species of Amblypharyngodon. Numbers above nodes represent Bayesian Posterior Probabilities. The scale bar represents the number of changes per site B, D TCS haplotype network for species of Amblypharyngodon based on the analysis of B a 654-bp fragment of the COI gene and D a 497-bp fragment of the 16s gene. The sizes of the circles are proportional to the number of individuals sharing a given haplotype. The number of mutational steps is indicated by hatch marks. The black circles represent hypothetical nodes.
The SYM+G model was chosen as the best-fit nucleotide-substitution model under BIC in the jModelTest for the Bayesian phylogenetic inference of the 16s dataset. Here, Amblypharyngodon melettinus was recovered as the sister species of A. grandisquamis (PP = 85, Figure
In the combined 16s+COI (1151 bp) Bayesian phylogenetic inference, A. mola was recovered as the sister species of A. grandisquamis (PP = 89) while A. melettinus was recovered as the sister species of [A. grandisquamis + A. mola] (PP = 100).
The uncorrected pairwise genetic distances obtained for the 16s, COI, and cytb genes for the species of Amblypharyngodon is given in Table
Intraspecific and interspecific percentage uncorrected pairwise genetic distances, for the 16s, COI, and cytb genes, between species of Amblypharyngodon.
| A. grandisquamis | A. melettinus | A. mola | A. chulabhornae | |||||||||
| % | 16s | COI | cytb | 16s | COI | cytb | 16s | COI | cytb | 16s | COI | cytb |
| A. grandisquamis | 0.0–0.4 | 0.0–1.9 | 0.0–4.2 | |||||||||
| A. melettinus | 4.4–5.2 | 9.9–11.0 | NA | 0.0–0.4 | 0.8–1.1 | NA | ||||||
| A. mola | 4.0–4.6 | 6.6–8.3 | 7.0–9.2 | 5.6–6.3 | 9.6–10.7 | NA | 0.0–0.6 | 0.0–0.8 | 0.0–0.4 | |||
| A. chulabhornae | 6.3–6.9 | 10.2–10.5 | 14.8–15.1 | 7.3–7.9 | 10.7–11.6 | NA | 5.9–6.7 | 10.7–11.0 | 14.4–14.8 | 0.8 | NA | 0.0 |
In the COI haplotype network, the dry zone and wet zone samples of A. grandisquamis form a shared a haplotype (H1) (Figure
Only three haplotypes occur in the 16s haplotype of A. grandisquamis (Figure
In the cytb haplotype network for A. grandisquamis, the only representative of the wet zone population (DZ 3292) in our molecular analysis formed a unique haplotype, while the dry zone samples formed four unique haplotypes. Populations of A. grandisquamis (ten sequences) included 22 segregating sites and three parsimony-informative sites. The nucleotide diversity and haplotype diversity for A. grandisquamis were 0.0101 and 0.667, respectively. Although Tajima’s D test was positive (1.7016) and Fu and Li’s F* test statistic was negative (-2.0688), neither was significant (p > 0.05, p > 0.02, respectively).
Statistical analysis
The morphometric PCA of the syntypes of A. melettinus (

Component loadings in the principal component analysis of the size-adjusted morphometric measurements of the paratypes of Amblypharyngodon grandisquamis (SU 22868) and syntypes of A. melettinus (
| Component 1 | Component 2 | Component 3 | |
| Eigenvalue | 5.4435 | 0.9590 | 0.6933 |
| Variance explained | 73.725 | 12.989 | 9.3901 |
| Body depth | 0.8028 | 0.5239 | -0.1882 |
| Caudal peduncle length | 0.5863 | -0.7560 | 0.1355 |
| Caudal peduncle depth | -0.0145 | -0.0398 | -0.4534 |
| Head length | 0.0500 | 0.1730 | 0.8129 |
| Snout length | -0.0563 | 0.1567 | -0.1712 |
| Eye diameter | -0.0286 | -0.0125 | 0.0303 |
| Inter-orbital width | 0.0698 | 0.3125 | 0.2225 |
Component loadings in the principal component analysis of the size-adjusted morphometric measurements of the recent material of Amblypharyngodon examined in the study.
| Component 1 | Component 2 | Component 3 | |
| Eigenvalue | 4.4466 | 3.2916 | 1.8519 |
| Variance explained | 30.144 | 22.314 | 12.554 |
| Predorsal length | 0.0175 | 0.3200 | -0.0137 |
| Postdorsal length | 0.2933 | -0.2495 | -0.1115 |
| Preanal length | -0.4055 | 0.2789 | 0.2023 |
| Prepelvic length | -0.2027 | 0.2117 | 0.1323 |
| Caudal peduncle length | 0.3304 | 0.0390 | -0.3040 |
| Caudal peduncle depth | 0.1731 | -0.0041 | 0.0432 |
| Body depth | 0.5663 | -0.0706 | 0.1794 |
| Dorsal-fin height | 0.0970 | -0.2158 | 0.5209 |
| Dorsal-fin base length | 0.0238 | -0.1490 | 0.1537 |
| Anal-fin height | 0.0730 | -0.0733 | 0.3317 |
| Anal-fin base length | 0.0095 | -0.1455 | 0.1407 |
| Pelvic-fin height | 0.0025 | 0.0514 | 0.3114 |
| Pelvic-fin base length | 0.0096 | 0.0160 | 0.0913 |
| Pectoral-fin length | 0.0484 | -0.2036 | 0.3004 |
| Pelvic-anal distance | -0.1957 | 0.0167 | 0.1557 |
| Head length | 0.2971 | 0.6690 | 0.0630 |
| Head depth | 0.2368 | 0.3131 | 0.3509 |
| Snout length | 0.0013 | 0.0617 | 0.0177 |
| Eye diameter | -0.0671 | 0.0275 | 0.0929 |
| Post-orbital head length | 0.1730 | 0.1212 | -0.01411 |
| Inter-orbital width | 0.1131 | 0.0311 | -0.0267 |
| Inter-narial width | -0.0175 | 0.0100 | 0.0304 |
Amblypharyngodon grandisquamis
Amblypharyngodon
melettinus
, Non-Valenciennes, 1844 (from Sri Lanka):
Material examined (all from Sri Lanka)
SU 22868, 8 paratypes, 56.4–65.4 mm SL, river at Colombo (presumably the Kelani, which is the only river in the vicinity of the city); WHT 30275, 6, 65.0–76.4 mm SL, Bolgoda River basin, Bellanwila-Attidiya; WHT 64, 3, 45.4–47.5 mm SL, Kalu River basin, Ingiriya, Dombagaskanda; WHT 22, 2, 37.2–38.7 mm SL, Kalu River basin, Walandure, Kuruwita; WHT 225, 2, 30.2–34.3 mm SL, Kalu River basin, Ekneligoda, Kuruwita; WHT 30790, 3, 49.3–52.8 mm SL, Kelani River basin, Deraniyagala; WHT 32, 3, 49.6–60.1 mm SL, Attanagalu Oya basin, Weliweriya; WHT 30260, 58.2 mm SL, Nilwala River basin, Godapitiya, Akuressa; WHT 30167, 3, 54.2–74.7 mm SL, Malwathu Oya basin, Anuradhapura; DZ 3193, 7, 57.2–74.0 mm SL, Mahaweli River basin, Polonnaruwa; DZ 3564, 2, 57.2–62.3 mm SL, Mahaweli River basin, Badulu Oya, Kandeketiya; WHT 1694, 3, 42.7–45.1 mm SL, Mahaweli River basin, Wasgamuwa; WHT 45, 4, 54.6–71.9 mm SL, Mi Oya basin, Puttlam; WHT 1810, 4, 40.8–52.4 mm SL, Kirindi Oya basin, Weerawila; WHT 1851, 41.2 mm SL, Gal Oya basin, Rathugala near Bibile. Cleared and stained: WHT 11075, 64.2 mm SL, Bolgoda River basin, Bellanwila-Attidiya; WHT 11046, 54.2 mm SL, Mi Oya basin, Puttlam; WHT 11081, 45.1 mm SL, Mi Oya basin, Puttlam; WHT 11052, 45.9 mm SL, Kelani River basin, Deraniyagala; WHT 11070, 40.0 mm SL, Kelani River basin, Deraniyagala. Other material (identified for distribution data, but not measured): WHT 30789, Mahaweli River basin, Wasgamuwa; DZ 3434, Mahaweli River basin, Naula; DZ 1504, Mahaweli River basin, Ulhitiya; WHT 30263, Deduru Oya basin, Wewagama, Kuliyapitiya; WHT 30679, Deduru Oya basin; WHT 2158, Kirindi Oya basin, Tissamaharama; DZ 4423, Kirindi Oya basin, Lunugamwehera; WHT 30723, Heda Oya, Lahugala; WHT 30310, Kala Oya basin, Eluwankulam; WHT 30167, Malwathu Oya basin, Anuradhapura; DZ 4042, Attanagalu Oya basin, Yakkala; DZ 3577, Mi Oya basin, Galgamuwa; DZ 3292, Kalu River basin, Remuna.
Diagnosis
Amblypharyngodon grandisquamis is distinguished from A. melettinus by having a deeper body (26.9–31.2% SL in A. grandisquamis, vs. 22.9–26.3 in A. melettinus); fewer pored lateral-line scales (8–16 (25), 19 (1), vs. 15(1), 17–21 (13) in A. melettinus: Figure

Description
For general appearance, see Figure
Morphometric data for Amblypharyngodon grandisquamis (SU 22868, WHT 30275, WHT 64, WHT 30790, WHT 32, WHT 30260, WHT 30167, WHT 45, DZ 3193, DZ 3564), A. melettinus (
| A. grandisquamis | A. melettinus | A. mola (n = 5) | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| paratypes of A. grandisquamis (n = 8) | recently collected material of A. grandisquamis (n = 23) | syntypes of A. melettinus (n = 5) | recently collected material of A. melettinus (n = 14) | |||||||||||||||||
| min | max | mean | s.d. | min | max | mean | s.d. | min | max | mean | s.d. | min | max | mean | s.d. | min | max | mean | s.d. | |
| Standard length (mm) | 56.4 | 65.4 | – | – | 45.4 | 76.4 | – | – | 60.7 | 65.7 | – | – | 51.4 | 77.3 | – | – | 43.2 | 47.5 | – | – |
| As percentage of standard length | ||||||||||||||||||||
| Total length | – | – | – | – | 125 | 128 | 127 | 0.9 | – | – | – | – | 126 | 131 | 128 | 1.4 | 126 | 132 | 130 | 2.6 |
| Predorsal length | – | – | – | – | 52.8 | 57.7 | 55.2 | 1.3 | – | – | – | – | 53.8 | 56.7 | 55.2 | 0.8 | 51.5 | 56.3 | 54.7 | 1.9 |
| Postdorsal length | – | – | – | – | 47.4 | 53.1 | 50.4 | 1.5 | – | – | – | – | 46.9 | 50.0 | 48.6 | 0.9 | 48.9 | 52.6 | 50.3 | 1.5 |
| Preanal length | – | – | – | – | 64.3 | 69.4 | 67.4 | 1.3 | – | – | – | – | 68.9 | 72.6 | 70.6 | 1.3 | 67.8 | 69.7 | 68.5 | 0.8 |
| Prepelvic length | – | – | – | – | 47.6 | 52.3 | 49.7 | 1.2 | – | – | – | – | 48.8 | 53.1 | 50.9 | 1.4 | 48.6 | 53.9 | 50.3 | 2.1 |
| Caudal peduncle length | 19.9 | 24.2 | 22.7 | 2.1 | 21.1 | 25.7 | 23.9 | 1.1 | 17.8 | 21.1 | 18.9 | 1.3 | 20.4 | 23.8 | 21.7 | 0.8 | 19.4 | 22.1 | 21.3 | 1.1 |
| Caudal peduncle depth | 10.6 | 13.3 | 12.0 | 0.9 | 11.4 | 13.9 | 12.5 | 0.6 | 12.2 | 13.2 | 12.7 | 0.4 | 10.3 | 12.4 | 11.3 | 0.7 | 10.7 | 11.8 | 11.2 | 0.5 |
| Body depth | 29.3 | 33.6 | 31.8 | 1.6 | 26.9 | 31.2 | 29.0 | 1.2 | 23.2 | 28.8 | 26.6 | 2.3 | 22.9 | 26.3 | 24.7 | 1.0 | 28.0 | 29.7 | 28.7 | 0.7 |
| Dorsal–fin height | – | – | – | – | 21.1 | 27.6 | 24.5 | 2.0 | – | – | – | – | 21.9 | 26.1 | 23.8 | 1.3 | 27.8 | 29.2 | 28.2 | 0.6 |
| Dorsal–fin base length | – | – | – | – | 10.6 | 13.6 | 12.0 | 0.8 | – | – | – | – | 10.0 | 13.5 | 11.7 | 1.0 | 12.0 | 14.1 | 13.2 | 0.8 |
| Anal–fin height | – | – | – | – | 14.6 | 19.8 | 17.1 | 1.5 | – | – | – | – | 15.5 | 19.4 | 16.8 | 1.0 | 17.3 | 21.3 | 18.8 | 1.5 |
| Anal–fin base length | – | – | – | – | 10.1 | 13.3 | 11.7 | 0.8 | – | – | – | – | 10.0 | 12.6 | 11.6 | 0.9 | 13.0 | 15.1 | 14.1 | 0.9 |
| Pelvic–fin height | – | – | – | – | 14.7 | 19.4 | 16.8 | 1.2 | – | – | – | – | 15.8 | 18.6 | 16.9 | 0.9 | 17.5 | 19.6 | 18.7 | 0.8 |
| Pelvic–fin base length | – | – | – | – | 4.2 | 5.7 | 4.9 | 0.4 | – | – | – | – | 3.7 | 6.8 | 5.0 | 0.9 | 3.7 | 5.3 | 4.6 | 0.7 |
| Pectoral fin height | – | – | – | – | 15.6 | 21.5 | 18.6 | 1.5 | – | – | – | – | 17.2 | 19.9 | 18.2 | 0.7 | 20.0 | 22.3 | 21.4 | 1.0 |
| Pelvic–anal distance | – | – | – | – | 17.5 | 20.9 | 19.7 | 0.9 | – | – | – | – | 19.7 | 23.9 | 21.4 | 0.9 | 18.7 | 22.0 | 20.0 | 1.4 |
| Head length | 26.9 | 30.9 | 28.7 | 1.2 | 24.6 | 39.7 | 28.5 | 2.7 | 25.8 | 28.9 | 27.7 | 1.3 | 26.1 | 29.0 | 27.4 | 1.0 | 25.9 | 29.4 | 27.7 | 1.4 |
| Head depth | – | – | – | – | 18.2 | 31.5 | 21.1 | 2.4 | – | – | – | – | 18.5 | 20.3 | 19.6 | 0.6 | 20.4 | 22.5 | 21.4 | 0.8 |
| As percentage of head length | ||||||||||||||||||||
| Snout length | 20.1 | 26.0 | 23.5 | 1.8 | 19.3 | 29.2 | 24.3 | 2.6 | 25.4 | 29.8 | 27.0 | 1.8 | 23.4 | 28.6 | 25.8 | 1.6 | 23.7 | 27.5 | 24.8 | 1.5 |
| Eye diameter | 24.1 | 26.9 | 26.2 | 1.2 | 22.7 | 30.5 | 27.0 | 2.6 | 26.8 | 30.4 | 28.2 | 1.5 | 25.2 | 33.5 | 28.8 | 2.3 | 31.3 | 36.6 | 33.7 | 2.5 |
| Post–orbital head length | – | – | – | – | 39.6 | 57.5 | 52.9 | 3.5 | – | – | – | – | 45.6 | 53.6 | 50.3 | 2.5 | 48.9 | 52.8 | 50.2 | 1.5 |
| Inter–orbital width | 31.9 | 36.6 | 34.7 | 1.6 | 24.4 | 42.3 | 36.6 | 3.5 | 28.3 | 38.5 | 34.9 | 3.9 | 31.8 | 38.0 | 34.8 | 1.5 | 32.8 | 37.1 | 34.9 | 1.7 |
| Inter–narial width | – | – | – | – | 14.7 | 28.6 | 21.8 | 2.7 | – | – | – | – | 19.5 | 25.3 | 22.1 | 1.9 | 22.1 | 24.4 | 23.3 | 1.2 |

Amblypharyngodon grandisquamis, A in aquarium (~70 mm SL, not collected), Sri Lanka, Ulhitiya, Mahaweli River basin B in preservation (DZ 3193A, 74.0 mm SL), Sri Lanka, Polonnaruwa, Mahaweli River basin C illustration of the holotype of A. grandisquamis by
Lateral line incomplete, with 8 (1), 9 (2), 10 (4), 11 (4), 12 (6), 13 (5), 15 (1), 16 (2) or 19 (1) pored scales, 42 (1), 44 (1), 45 (4), 46 (3), 47 (4), 48 (4), 49 (2), 50 (1), 51 (1), 52 (2), 53 (2) or 56 (1) scales in lateral series plus 2–4 scales on base of caudal fin. Predorsal scales 27 (2), 28 (3), 29 (2), 30 (2) or 32 (1). Prepelvic scales 25 (1), 26 (1), 27 (1), 28 (1), 29 (1), 30 (2), 31 (1), 33 (1) or 34 (1). Lateral scale rows between origins of dorsal and pelvic fins ½8+1+5½ (1), ½8+1+6 (1), ½9+1+4½ (2), ½9+1+5½ (8), ½9+1+6 (1), ½9+1+6½ (2), ½10+1+4½ (1), ½10+1+5 (1), ½10+1+5½ (4), or ½10+1+6½ (5). Circumpeduncular scales 20 (2), 21 (7), 22 (10), 23 (5) or 24 (2).
Dorsal fin with two unbranched and seven branched rays, its origin just posterior to vertical through pelvic-fin origin, its distal margin straight. Anal fin with three unbranched and five branched rays, its origin slightly posterior to vertical through origin of dorsal fin, its distal margin slightly concave. Pectoral fin with a single unbranched and 11 (1), 12 (10), 13 (2), or 14 (1) branched rays, its origin anterior to posteriormost point of opercular opening, not reaching pelvic-fin origin when adpressed. Pectoral-fin axillary lobe rudimentary. Pelvic fin with one unbranched and 7 (3) or 8 (13) branched rays, its origin slightly closer to anal-fin origin than to origin of pectoral fin, its tip not reaching anal-fin origin when adpressed. Pelvic ‘axillary’ scale present. Caudal fin with 9 + 8 (14) branched rays, forked, lobes rounded distally, upper and lower lobes subequal.
Vertebrae 15 + 18 = 33 (1), 15 + 19 = 34 (4). Pharyngeal teeth 5 + 2 + 1 (4), 5 + 3 + 1 (1) (Figure

Coloration
In 70% alcohol (Figure
In life (Figure
Habitat, distribution, and natural history
Amblypharyngodon grandisquamis occurs in lotic habitats such as rivers and canals as well as in lentic habitats such as reservoirs and marshes. The species is recorded primarily from the lowland floodplain of Sri Lanka, in both the dry and the wet zones of the island (annual precipitation less than, and greater than, 2,000 mm, respectively), though much more frequently encountered in the dry zone (Figure

Discussion
While
Amblypharyngodon melettinus
The species we here refer to Amblypharyngodon grandisquamis was first reported from Sri Lanka by
The syntypes of A. melettinus are in quite poor condition. Jean-Jacques Dussumier probably collected these on his visit to Bombay 1827–30 (see

A syntype of Amblypharyngodon melettinus,
Our search for recent specimens of A. melettinus from the vicinity of Mumbai led to the examination of two specimens of Amblypharyngodon,
Amblypharyngodon microlepis
In differentiating A. microlepis from A. melettinus,
Bleeker was evidently misled by the illustration of A. melettinus that accompanied the original description (Valenciennes 1844: pl. 501, reproduced here as Figure
Amblypharyngodon mola etc
Amblypharyngodon grandisquamis is easily distinguished from A. mola by possessing fewer scales in the lateral series: 42–56, vs. 69–73 in A. mola from the putatively topotypical West Bengal specimens examined by us, 65–79 in the Orissa specimens examined by
There remain five additional nominal species of Amblypharyngodon reported from the Indian peninsula: A. chakaiensis Babu Rao and Nair (type locality the Chakai backwater near Trivandrum [Thiruvananthapuram], Kerala); Leuciscus chitul Sykes (‘Inderance River near Chakun, Deccan’); Rhodeus indicus Jerdon (Palghat [Palakkad] River, Kerala); Brachygramma jerdonii Day (Cochin [Kochi], Kerala); and Rhodeus macrocephalus Jerdon (Cavery [Kaveri] River and Carnatic [Karnataka], southern India). Except for A. jerdoni, which
Genetic analysis
While our genetic analysis clearly separates A. grandisquamis from its peninsular-Indian congeners A. melettinus and A. mola, it does not fully resolve the sister-group relationships of A. grandisquamis. Our COI and the COI+16s combined analyses recover A. melettinus as the sister group of [A. grandisquamis + A. mola], whereas our 16s phylogram recovers A. mola as the sister group of [A. grandisquamis + A. melettinus].
The uncorrected pairwise genetic distances for COI between A. grandisquamis and A. melettinus, A. mola and A. chulabhornae, of 9%, 6% and 10%, respectively, are consistent with the distances observed between independently validated species (
The non-significant results of the neutrality tests suggest that there has been no recent range expansion or population bottleneck in A. grandisquamis in Sri Lanka.
Conservation
The National Red List of Sri Lanka (
Conclusions
Since the description of A. grandisquamis by
Acknowledgments
HS and MM are grateful to the Director General of Wildlife Conservation and the Conservator General of Forests, Sri Lanka, for permits to carry out fieldwork. HS thanks Rahul Kumar, Rajeev Raghavan, and Unmesh Katwate for useful discussions, providing generous hospitality during a visit to India and allowing access to Indian material in their collections; Sanuja Kasthuriarachchi, Director of National Museums, and Lankani Somarathna and her staff, for access to specimens; the Wildlife Heritage Trust of Sri Lanka for providing financial support for this research; and finally Charana Widuranga, Dhanushka Lakshan, Kumudu Wijesooriya, and R.H. Tharindu Ranasinghe for their assistance in the field. RP thanks Patrice Pruvost and Zara Gabsi (
References
- Batuwita S, Maduwage K, Sudasinghe H (2015) Redescription of Pethia melanomaculata (Teleostei: Cyprinidae) from Sri Lanka. Zootaxa 3936: 575–583. https://doi.org/10.11646/zootaxa.3936.4.7
- Babu RM, Nair NB (1978) On a new species of Amblypharyngodon (Cyprininae – Cyprinidae) from Trivandrum (Kerala, India). Aquatic Biology 3: 121–125.
- Bauchot ML, Daget J, Bauchot R (1990) L’ichtyologie en France au début du XIXè siècle: L’histoire naturelle des poissons de Cuvier & Valenciennes. Bulletin du Muséum National d’Histoire Naturelle, Paris, Sect. A, Sér. 4, 12(1, Suppl.): 1–142.
- Bleeker P (1853) Nalezingen op de ichthyologische fauna van Bengalen en Hindostan. Verhandelingen van het Bataviaasch Genootschap van Kunsten en Wetenschappen 25: 1–164. https://doi.org/10.5962/bhl.title.60451
- Bossuyt F, Meegaskumbura M, Beenaerts N, Gower DJ, Pethiyagoda R, Roelants K, Mannaert A, Wilkinson M, Bahir MM, Manamendra-Arachchi K, Schneider CJ, Oommen OV, Milinkovitch MC (2004) Local endemism within the western Ghats-Sri Lanka biodiversity hotspot. Science 306: 479–481. https://doi.org/10.1126/science.1100167
- Clement M, Snell Q, Walke P, Posada D, Crandall K (2002) TCS: estimating gene genealogies. Proceedings of the 16th International Parallel and Distributed Processing Symposium (USA), April 2002. 2: 184. https://doi.org/10.1109/IPDPS.2002.1016585
- Collen E (1909) Imperial gazetteer of India: The Indian Empire; Vol. 4: Administrative. Clarendon Press, Oxford, 552 pp.
- Conway KW (2011) Osteology of the South Asian genus Psilorhynchus McClelland, 1839 (Teleostei: Ostariophysi: Psilorhynchidae), with investigation of its phylogenetic relationships within the order Cypriniformes. Zoological Journal of the Linnean Society 163: 150–154. https://doi.org/10.1111/j.1096-3642.2011.00698.x
- Cuvier G, Valenciennes A (1844) Histoire naturelle des poissons. Tome seizième. Livre dix-huitième. Les Cyprinoïdes 17: 1–497. [pls 487–519]
- Darriba D, Taboada GL, Doallo R, Posada D (2012) jModelTest 2: more models, new heuristics and parallel computing. Nature methods 9: 772. https://doi.org/10.1038/nmeth.2109
- Day F (1878) Fishes of India, Part 4. Quaritch, London, 553–778. [pls 134–195]
- Day F (1889) The Fauna of British India, including Ceylon and Burma. Fishes Vol. 1. Taylor & Francis, London, 548 pp.
- Dayrat B (2005) Towards integrative taxonomy. Biological Journal of the Linnean Society 85: 407–415. https://doi.org/10.1111/j.1095-8312.2005.00503.x
- De Silva MA, Hapuarachchi N, Jayaratne T (2015) Sri Lankan Freshwater Fishes. Wildlife Conservation Society, Galle, 391 pp.
- Deraniyagala PEP (1952) A coloured atlas of some vertebrates from Ceylon, Vol. 1. Fishes. National Museums of Ceylon, Colombo, 149 pp. [34 pls]
- Duncker G (1912) Die Süßwasserfische Ceylons. Mitteilungen aus dem Naturhistorischen (Zoologischen) Museum in Hamburg 29: 241–272. [1 pl.]
- Elliott NG, Haskard K, Koslow JA (1995) Morphometric analysis of orange roughy (Hoplostethus atlanticus) off the continental slope of southern Australia. Journal of Fish Biology 46: 202–220. https://doi.org/10.1111/j.1095-8649.1995.tb05962.x
- Fu YX, WH Li (1993) Statistical tests of neutrality of mutations. Genetics 133: 693–709.
- Guindon S, Gascuel O (2003) A simple, fast and accurate method to estimate large phylogenies by maximum-likelihood. Systematic Biology 52: 696–704. https://doi.org/10.1080/10635150390235520
- Günther A (1868) Catalogue of the fishes in the British Museum. Vol. 7. Catalogue of the Physostomi containing the families Heteropygii, Cyprinidae, Gonorhynchidae, Hyodontidae, Osteoglossidae, Clupeidae, Chirocentridae, Alepocephalidae, Notopteridae, Halosauridae, in the collection of the British Museum. British Museum, London, 512 pp.
- Hammer Ø, Harper DAT, Ryan PD (2001) PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontologia Electronica 4: 1–9.
- Huelsenbeck JP, Ronquist F, Nielsen R, Bollback JP (2001) Bayesian inference of phylogeny and its impact on evolutionary biology. Science 294: 2310–2314. https://doi.org/10.1126/science.1065889
- Jayaram KC (2010) The Freshwater Fishes of the Indian region (2nd edn). Narendra Publishing House, Delhi, 616 pp.
- Jordan DS, Starks EC (1917) Notes on a collection of fishes from Ceylon with descriptions of new species. Annals of the Carnegie Museum 11: 430–460. [pls 43–47]
- Kumar S, Stecher G, Tamura K (2016) MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Molecular Biology and Evolution 33: 1870–1874. https://doi.org/10.1093/molbev/msw054
- Leigh JW, Bryant D (2015) PopART: Full-feature software for haplotype network construction. Methods in Ecology and Evolution 6: 1110–1116. https://doi.org/10.1111/2041-210X.12410
- Menon AGK (1999) Check list – Fresh water fishes of India. Records of the Zoological Survey of India, Occasional Papers 175: 1–366.
- MOE (2012) The National Red List 2012 of Sri Lanka; conservation status of the fauna and flora. Ministry of Environment, Colombo, 476 pp.
- Ng HH, Pethiyagoda R (2013) Mystus zeylanicus, a new species of bagrid catfish from Sri Lanka (Teleostei: Bagridae). Ichthyological Exploration of Freshwaters 24: 161–170.
- Pet JS, Gevers GJM, Van Densen WLT, Vijverberg J (1996) Management options for a more complete utilization of the biological fish production in Sri Lankan reservoirs. Ecology of Freshwater Fish 5: 1–14. https://doi.org/10.1111/j.1600-0633.1996.tb00032.x
- Pethiyagoda R (1991) Freshwater fishes of Sri Lanka. Wildlife Heritage Trust, Colombo, Sri Lanka, 362 pp.
- Pethiyagoda R (2006) Conservation of Sri Lankan freshwater fishes. In: Bambaradeniya CNB (Ed.) Fauna of Sri Lanka: status of taxonomy, research and conservation.The World Conservation Union, Colombo, 103–112.
- Rambaut A, Suchard MA, Xie D, Drummond AJ (2014) Tracer v1.6, https://tree.bio.ed.ac.uk/software/tracer
- Ronquist F, Teslenko M, van der Mark P, Ayres DL, Darling A, Höhna S, Larget B, Liu L, Suchard MA, Huelsenbeck JP (2012) MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Systematic Biology 61: 539–542. https://dx.doi.org/10.1093/sysbio/sys029
- Rozas J, Ferrer-Mata A, Sánchez-Del Barrio JC, Guirao-Rico S, Librado P, Ramos-Onsins SE, Sánchez-Gracia A (2017) DnaSP 6: DNA sequence polymorphism analysis of large data sets. Molecular Biology and Evolution 34: 3299–3302. https://doi.org/10.1093/molbev/msx248
- Senanayake FR (1980) The Biogeography and ecology of the inland fishes of Sri Lanka. PhD Thesis, Department of Wildlife and Fisheries Biology, University of California, Davis.
- Sudasinghe H, Meegaskumbura M (2016) Ompok argestes, a new species of silurid catfish endemic to Sri Lanka (Teleostei: Siluridae). Zootaxa 4158: 261–271. https://doi.org/10.11646/zootaxa.4158.2.7
- Sudasinghe H, Pethiyagoda R, Maduwage K, Meegaskumbura M (2016) Mystus nanus, a new striped catfish from Sri Lanka (Teleostei: Bagridae). Ichthyological Exploration of Freshwaters 27: 163–172.
- Sudasinghe H, Ranasinghe RHT, Goonatilake SDA, Meegaskumbura M (2018a) A review of the genus Labeo (Teleostei: Cyprinidae) in Sri Lanka. Zootaxa 4486: 201–235. https://doi.org/10.11646/zootaxa.4486.3.1
- Sudasinghe H, Herath J, Pethiyagoda R, Meegaskumbura M (2018b) . Undocumented translocations spawn taxonomic inflation in Sri Lankan fire rasboras (Actinopterygii, Cyprinidae). PeerJ 6: e6084. https://doi.org/10.7717/peerj.6084
- Tajima F (1989) Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 123: 585–595.
- Talwar PK, Jhingran AG (1991) Inland fishes of India and adjacent countries. Oxford & IBH Publishing Company, New Delhi, 2 vols, 1158 pp. [3 pls]
- Taylor WR, Van Dyke GC (1985) Revised procedures for staining and clearing small fishes and other vertebrates for bone and cartilage study. Cybium 9: 107–119.
- Vidthayanon C, Kottelat M (1990) Amblypharyngodon chulabhornae sp. nov., a new cyprinid fish from Thailand and Kampuchea. Natural History Bulletin of the Siam Society 38: 45–57.
- Wagh GK, Ghate HV (2002) Freshwater fish fauna of the rivers Mula and Mutha, Pune, Maharashtra. Zoos’ Print Journal 18: 977–981. https://doi.org/10.11609/JoTT.ZPJ.18.1.977-89
- Wahab MA, Alim MA, Milstein A (2003) Effects of adding the small fish punti (Puntius sophore Hamilton) and/or mola (Amblypharyngodon mola Hamilton) to a polyculture of large carp. Aquaculture Research 34: 149–163. https://doi.org/10.1046/j.1365-2109.2003.00784.x
- Ward RD (2009) DNA barcode divergence among species and genera of birds and fishes. Molecular Ecology Resources 9: 1077–1085. https://doi.org/10.1111/j.1755-0998.2009.02541.x