





(C) 2012 Pavel Valdés. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Comparisons of structural features (principally mouthparts, elytral-abdominal locking mechanism, and female genitalia) of the ardistomine genera (Aspidoglossa Putzeys, Ardistomis Putzeys, and Semiardistomis Kult) with those features of members of the subtribe Clivinina (Clivina Latreille, Oxydrepanus Putzeys, Schizogenius Putzeys, Ancus Putzeys, Nyctosyles Putzeys, and Obadius Burmeister) confirm the taxonomic validity of the subtribe Ardistomina. Based on morphological features, the ardistomine genera are postulated to be related as follows: [Aspidoglossa [Ardistomis + Semiardistomis]]. Knowledge of this subtribe is further extended by taxonomic treatment of the genus Semiardistomis Kult. Of the 30 valid names of Semiardistomis previously recognized, four were proposed as variety names, and are listed as junior synonyms: Ardistomis labialis picipes Bates, 1881, Ardistomis labialis nanus Bates, 1881, and Ardistomis labialis dilatatus Bates, 1881; and Ardistomis pallipes caerulea Putzeys, 1846. Eight names, treated as specific epithets, are junior synonyms, as follows: Ardistomis (Semiardistomis) balthasari Kult, 1950 = Semiardistomis glabratus (Putzeys, 1866); Ardistomis (Semiardistomis) emdeni Kult, 1950 = Semiardistomis deletus (Putzeys, 1846); Ardistomis aenea Putzeys, 1866, Ardistomis (Semiardistomis) brittoni Kult, 1950, and Ardistomis (Semiardistomis) marani Kult, 1950 = Semiardistomis flavipes (Dejean, 1831); Ardistomis tuspanensis Putzeys, 1846 = Semiardistomis labialis (Chaudoir, 1837); Ardistomis (Semiardistomis) vlastae Kult, 1950 = Semiardistomis subglabra (van Emden, 1949); and Ardistomis striga Putzeys, 1866 = Semiardistomis pallipes (Dejean, 1831). Two new species described are Semiardistomis exspectatus sp. n. (type locality PERU, Madre de Dios, Rio Manu, 11°56'47"S, 071°17'00"W), and Semiardistomis major sp. n. (type locality PERU, Loreto, Rio Samiria, 05°12'S, 75°20'W). The 20 species of Semiardistomis are arranged intwo species–groups here proposed: the puncticollis group, including 12 species; and labialis group, including eight species. The species recognized are keyed, described or redescribed, and notes are provided about their Geographical distribution, habitat and activity. Distribution maps show known geographical ranges, from which are inferred patterns of speciation from a center of radiation in northern South America of both lineages.
Resumen
La comparación de las características estructurales (principalmente partes bucales, mecanismo de cierre élitro- abdominal y genitalia femenina) de los géneros ardistominos (Aspidoglossa Putzeys, Ardistomis Putzeys, y Semiardistomis Kult) con sus similares de miembros de la subtribu Clivinina (Clivina Latreille, Oxydrepanus Putzeys, Schizogenius Putzeys, Ancus Putzeys, Nyctosyles Putzeys, y Obadius Burmeister) confirma la validez taxonómica de la subtribu Ardistomina. Basado en características morfológicas, se postula una relación entre los géneros ardistominos de forma: [Aspidoglossa [Ardistomis + Semiardistomis]]. El tratado taxonómico del género Semiardistomis Kult extiende el conocimiento de esta subtribu. De los 30 nombres válidos previamente reconocidos, cuatro estaban propuestos como variedades, y se tratan como sinónimos: Ardistomis labialis picipes Bates, 1881, Ardistomis labialis nanus Bates, 1881, y Ardistomis labialis dilatatus Bates, 1881; y Ardistomis pallipes caerulea Putzeys, 1846. Ocho nombres usados como epítetos específicos resultan los siguientes sinónimos: Ardistomis (Semiardistomis) balthasari Kult, 1950 = Semiardistomis glabratus (Putzeys, 1866); Ardistomis (Semiardistomis) emdeni Kult, 1950 = Semiardistomis deletus (Putzeys, 1846); Ardistomis aenea Putzeys, 1866, Ardistomis (Semiardistomis) brittoni Kult, 1950, y Ardistomis (Semiardistomis) marani Kult, 1950 = Semiardistomis flavipes (Dejean, 1831); Ardistomis tuspanensis Putzeys, 1846 = Semiardistomis labialis (Chaudoir, 1837); Ardistomis (Semiardistomis) vlastae Kult, 1950 = Semiardistomis subglabra (van Emden, 1949); y Ardistomis striga Putzeys, 1866 = Semiardistomis pallipes (Dejean, 1831). Las dos especies nuevas descritas son Semiardistomis exspectatus sp. n. (localidad tipo PERU, Madre de Dios, Rio Manu, 11°56'47"S, 071°17'00"W), y Semiardistomis major sp. n. (localidad tipo PERU, Loreto, Rio Samiria, 05°12'S, 75°20'W). Las 20 especies de Semiardistomis se distribuyen en dos grupos de especies aquí establecidos: el grupo puncticollis con 12 especies y el grupo labialis con 8. Las especies reconocidas son incluidas en una clave de identificación, se describen o redescriben, y se dan notas a cerca de su distribución, hábitat y actividad. Los mapas de distribución muestran los rangos geográficos conocidos, de los cuales se infieren patrones de especiación a partir de un centro de radiación en el Norte de Suramérica para ambos linajes.
Coleoptera, Carabidae, scaritids, new species
N’ayant vu qu’un seul individu, j’ignore si ces différences sont constantes et si l’insecte ne constitue pas une variété de l’A. pallipes.
(Putzeys, 1846: 646)
Between these extremes, however, there is, either in the same or other localities, every grade of variation
(Bates, 1881: 35)
The species of this group are very difficult to determine without preparing a key; they form some slightly different subspecies in some localities.
(Kult, 1950: 317)
Introduction
Although the major part of this paper treats the species of Semiardistomis, a more general background is provided by a preliminary treatment of the subtribe Ardistomina and comparison with other taxa of the tribe Clivinini, principally the nominotypical subtribe Clivinina. Many scaritid genera were included in the Ardistomina, but
The quotations above provide an idea of past taxonomic difficulties regarding members Semiardistomis. Interpretations made using non-diagnostic characters from short series of specimens have led to a taxonomic system, mostly useless, in order to identify and correlate beetles of this genus. The first arrangement of the group was made by
Whitehead [in
Study is based on 1038 specimens examined, provided by, or checked in, the following collections (names of curators or owners at the time of the loan in parentheses).
MHNP Muséum National d’Histoire Naturelle, Paris, France. (T. Deuve and A. Tagavian).
IRSNB Institut Royal des Sciences Naturelles de Belgique, Bruxelles, Belgique. (A. Drumont).
CMN Canadian Museum of Nature, Aylmer, Québec, Canada. (R. S. Anderson and F. Génier).
IES Instituto de Ecología y Sistemática, Habana, Cuba. (A. Lozada).
MNHNCu Museo Nacional de Historia Natural de Cuba, Habana, Cuba. (E. Gutiérrez)
PVCCu P.Valdés Collection, Habana, Cuba.
PBPC P. Bulirsch Collection. Prague, Czech Republic.
BMNH British Museum of Natural History. London, UK. (M. Barclay and C. Taylor)
HECO Hope Entomological Collection, Oxford University, UK. (J. Hogan).
UASM University of Alberta, Strickland Museum, Edmonton, Alberta, Canada. (G. E. Ball and D. Shpeley).
ADVA A. Dostal Collection (includes Kult Collection), Vienna, Austria.
CAS Department of Entomology, California Academy of Sciences, Golden Gate Park, San Francisco, U.S.A. (D. H. Kavanaugh).
FSCA Florida State Collection of Arthropods, Florida Department of Agriculture, Gainesville, Florida, U.S.A. (P. Skelley).
USNM National Museum of Natural History, Washington, D.C., U.S.A. (T. L. Erwin).
Dissections of adults were made using standard techniques. The genitalia were preserved in glycerin in a microvial, pinned beneath the specimen from which they were removed. Mouthparts were glued on small cards pinned beneath the specimens. Observations were made using a stereobinocular microscope and a compound microscope. All line drawings were made from digital microphotographs using Corel Draw 13X.
The following measurements were made using an ocular micrometer: head length (HL): linear distance from apical margin of clypeus to posterior margin of right eye; length of pronotum (PL): linear distance from anterior to posterior margin along the midline; pronotal width (PW): greatest linear transverse distance; elytral length (EL): linear distance from basal ridge to apex along the suture; elytral width (EW): greatest linear transverse distance across both elytra. The standardized body length (SBL) is the sum of the lengths of head, pronotum and elytra. Body ratios (PW/EW, PW/PL, PL/EL and EW/EL) are given for each species. Mandible proportion is interpreted from ratio of the length of the transverse line at outer molar point (width) and length of the perpendicular distance from that line to apical edge (length).
Most terms used for structural features are found in previous works on Carabidae:
Within the Clivinini. the subtribe Ardistomina is characterized as follows: Elytron latero-distally (Fig. 22) with a plica (ep) and an adjacent abdominal (segment VII) pleural projection (app) that together form an elytral-abdominal locking mechanism; ovipositor (Figs 27–30) with an asetose quadranguloid laterotergite, and unsegmented gonocoxa bearing at apex few long setae.
This subtribe includes Aspidoglossa Putzeys, Semiardistomis Kult, and Ardistomis Putzeys.
Mandibles, labium, and female genitalia (source of many diagnostic features within the tribe Clivinini) were compared within the ardistomines and between that subtribe and selected members of the nominotypic Clivinina. The latter taxon was represented by Western Hemisphere members of six exemplar genera: Clivina Latreille, Oxydrepanus Putzeys, Schizogenius Putzeys, Ancus Putzeys, Nyctosyles Putzeys, and Obadius Burmeister. Also, mandibles of Dyschiriodes Jeannel (subtribe Dyschiriina) were included.
Mandibles: Compared to those of Dyschiriina (Figs 12A–12D), the mandibles of the clivinines (Figs 9A–9D to 11A–11D) and ardistomines (Figs 6A–6D to 8A–8D) are relatively straight.
For dentition, Dyschiriodes exhibits the least number of occlusal teeth. The clivinine genera Clivina (Figs 9A–9D)and Oxydrepanus (Figs 10A–10D), and the ardistomine genera Aspidoglossa (Figs 8A–8D) and Ardistomis (Figs 7A–7D)have a full complement of occlusal teeth, Semiardistomis (Figs 6A–6D) and Schizogenius (Figs 11A–11D) lack the premolar tooth.
In values for L/W (a measure of relative length), the mandibles of clivinines, dyschiriines, and the ardistomine Aspidoglossa are relatively short (L/W 1.44–1.68), whereas the mandibles of the ardistomine genera Semiardistomis (L/W 1.86) and Ardistomis (L/W 2.25) are relatively long, with a slender terebra.
Labium: In form, the mentum (m) is markedly to slightly transverse (L/W 0.43–0.56) among the clivinine genera (Figs 17A–21A); among the ardistomine genera (Figs 13A–16A) the mentum is slightly transverse to slightly elongate (L/W 0.57–0.95), a morphocline being Aspidoglossa—Semiardistomis—Ardistomis.
In the Ardistomina, the apex of the lateral lobes (ll) is extended distally beyond the apex of the mental tooth (mt). This feature is varied among the clivinines: lateral lobes not extended beyond the apex of the mental tooth (Clivina, Oxydrepanus, and Nyctosyles, Figs 17A, 18A, 21A), to slightly extended beyond that line (Schizogenius and Ancus, Figs 19A, 20A).
The paraglossae (pg) are elongate, extended distinctly beyond the apex of the glossal sclerite (gs) in most genera of Ardistomina and Clivinina. In the ardistomine Ardistomis (Fig. 15B) and clivinine Nyctosyles (Fig. 21B) the paraglossae are much shorter than the glossal sclerite.
The glossal sclerite (gs) varies appreciably in both subtribes, being very large and rotund in Semiardistomis (Figs 13B, 14B4), broad with a broad apex in Nyctosyles (Fig. 21), and narrower and shorter in the remaining ardistomines and clivinines.
The glossal sclerite in the genus Semiardistomis has an extra pair of preapical setae (Figs 13B, 14B, pas). All other ardistomine and clivinine genera have a single pair of glossal setae (as).
Labial palpomere 3 (lp3) is elongate and fusiform in the Ardistomina (Figs 13A–16A). This sclerite varies in the clivinines: fusiform in Clivina (Fig. 17A), and variously widened in the remaining clivinine exemplar genera (Figs 18A–21A).
Female genitalia: The ovipositor sclerites come in two types, either ardistomine (Figs 27–30) or clivinine (Figs 31A–36A). For clivinine genera Clivina, Nyctosyles and Obadius, the gonocoxa is segmented, each segment designated as a gonocoxite (gc1, gc2). Gonocoxite 2 is more or less falcate. In ardistomine females, in contrast, the gonocoxa (gc) is unsegmented, and slightly curved or essentially straight. The laterotergites (lt) of clivinines are more or less triangular, whereas those of ardistomines are rectanguloid. Further, the laterotergites and gonocoxites of clivinines are setose along their margins, but the apex of gonocoxite 2 is glabrous. In contrast, for ardistomines, the ovipositor sclerites are essentially glabrous, but the apex of the gonocoxa bears a few setae. An unsegmented gonocoxa with reduced number of setae is also seen in Dyschiriina (Fedorenko, 1996). Within the ardistomines, the gonocoxae of Aspidoglossa and Ardistomis are moderately broad (Figs 29, 30), but those of Semiardistomis (Figs 27, 28) are slender, virtually rod-like.
Within the Clivinina, the reproductive tract (Figs 31A–36A) is strikingly varied, but the range of variation is about as extensive as, and similar to, that of the Ardistomina, and thus uninformative from a diagnostic perspective at subtribal level. Within each subtribe, the genera are clearly distinguishable from one another. Here, I treat only the ardistomine genera.
In Aspidoglossa females (Fig. 30), the spermathecal duct (spd) is narrow, elongate, with few loose coils proximally; the spermathecal gland (spgd) is relatively short. The female tract of Ardistomis (Fig. 29) is narrow, relatively short; the spermatheca (sp) is moderately long and markedly expanded distally. For Semiardistomis females (Figs 27, 28) the reproductive tracts of the two species-groups are sufficiently different from one another to require separate descriptions. They also differ markedly from the reproductive tracts of Aspidoglossa and Ardistomis. For details, see species-group treatments, below.
The members of the Ardistomina are hygrophilous or mesophilous, living in riparian situations, lowland swamp forests, or wet montane tropical forest.
As indicated above, of the three ardistomine genera, Aspidoglossa is most similar to members of subtribe Clivinina in mandibular and mental proportions and is more remote from Ardistomis and Semiardistomis in structure of the female reproductive tract than the latter two genera are from each other. In turn, Ardistomis and Semiardistomis share elongate mandibles and mentum. Based on these observations, I conclude that relationships of these three genera may be summarized using brackets as follows: [Aspidoglossa [Ardistomis + Semiardistomis]]
This subtribe is a Western Hemisphere indigenous group. Its range extends in South America east of the Andes mountain range from central Argentina northward through Colombia, Middle America, and the West Indies to temperate North America, principally east of the Mississippi drainage basin.
http://species-id.net/wiki/Semiardistomis
Features of adults used in recognition of this genus include: antennomere 2 shorter than antennomere 3 (Fig. 1, a2, a3); glossal sclerite of ligula (Figs 13B, 14B) with a secondary pair of setae; mentum with median carina (Figs 13C, 14C) extended distal in form of an appendiform keel; protibia (Figs 23, 24) with basal half of ventral surface with a group of setae.
Adult. Body shape, color and size (Figs 1, 2, 37–41, 44–52): Body pedunculate. Body size small, ranging from 4.0 to 7.5 mm. Body monochromous, dark with metallic greenish and/ or brassy luster, shiny in species without microsculpture; appendages reddish brown. One species, Semiardistomis laevistriatus (Fleutiaux & Sallé), is dark brown a color also found in monticolous brachypterous species of the genus Ardistomis.
Microsculpture: Frontoclypeus mostly smooth; supraantennal lobes smooth; vertex with mesh pattern isodiametric; gena with mesh pattern isodiametric; gula with mesh pattern transverse. Mandibles smooth; submentum and mentum with mesh pattern isodiametric. Pronotal disc generally smooth or with mesh pattern isodiametric, microlines very shallow, marginally and smooth at the center; proepisternum smooth or with isodiametric mesh pattern, submarginal band of microsculpture absent; prosternum smooth or with mesh pattern transverse. Metasternum smooth or with mesh pattern transverse. Abdominal sterna smooth or with mesh pattern transverse. Elytra smooth or with isodiametric mesh pattern covering entirely or only part of the disc.
Chaetotaxy: Two pair of supraorbital setae (except for Semiardistomis puncticollis); pronotal disc with two pair of marginal setae (except for Semiardistomis puncticollis and Semiardistomis viridis); except for hirsute species, setae on elytral disc located in interval 3 and Semiardistomis darlingtoni (Kult) also in interval 5; abdominal sterna IV-VII with accessory setae.
Head capsule: Clypeus with anterior margin concave medially; lateral lobes distinct, projected at the same level or below anterior margin. Antennal lobes prominent, sometimes partially sulcate medially in basal half. Frontoclypeus smooth except in Semiardistomis rugosus (Putzeys). Frontal impressions deep and wide from base of antennal lobes to anterior margin of clypeus. Gena with sides subparallel. Eyes: Hemispherical, prominent. Antennae (Figs 1, 2): Filiform to submoniliform. Antennomere 1 (a1) with single preapical seta. Antennomere 2 (a2) with few setae, shorter than antennomere 3 (a3). Antennomeres 4–11 densely setose, setae short.
Mouthparts: Labrum (Figs 3, 4): anterior margin angulate, dentiform medially (dp); dorsal setae (ds) 7, as for most Clivinini; apical portion of epipharyngeal setae (es) serrulate anteriolaterally. Maxillae (Fig. 5) with lacinia (lac) apical tooth sharp and curved; galeomere 1 (g1) slender, a little longer than galeomere 2 (g2), the latter flat, broad and sinuate in outline; palpomere 2 (mp2) thick and triangular in outline; palpomere 3 (mp3) subequal in length to palpomere 4 (mp4); palpomere 4 apically acuminate. Stipes (st) with 4 setae, palpifer (pf) with one seta. (This structure shows little difference from other clivinines being most similar to that of Ardistomis. Mandibles (Figs 6A–6D): elongate, about 1.9 times longer than wide; ventral groove (vg) moderate in length, with short microtrichia (mt); left mandible with terebral ridge curved, the terebral tooth (tt) blunt and moderate in size, visible from ventral view, retinaculum (rt) broad, premolar tooth (pm) absent, the molar tooth (m) broad and rounded; right mandible with terebral ridge more or less straight, the terebral tooth small, not visible from ventral view, the retinaculum with acute anterior tooth, premolar tooth absent, and the molar tooth broad and rounded. Labium (Figs 13A–C, 14A–C) with mental–submental suture complete; submentum with paramedian projections distinct, two pair of setae; mentum (m) trapezoidal, about 1.5 times wider than long; two pair of setae; lateral lobes with anterior margins angulate; mental tooth (mt) with distal margin almost straight, about 1/3 length of lateral lobes (ll); median carina (mc)(13C, 14C) extended distad, or not, beyond anterior margin of lateral lobes; pit organs (po) distinct, opened through oval orifices in basal part of mentum or on mental–submental suture; palpomere 3 (lp3) subequal in length to palpomere 2, palpomere 2 (lp2) bisetose; ligula (Figs 13B, 14B) with glossal sclerite (gs) notably wide, its anterior margin with acuminate median projection and two long setae (as), two additional small setae (pas) located preapically; paraglossae (pg) divergent from glossal sclerite and covered with microtrichia dorsally, lobes thin and acuminate, extended distally beyond distal margin of glossal sclerite, apices serrulate.
Pronotum. Ovate or cordate, anterior transverse and median longitudinal impressions distinct, proepisternum visible from above, lateral border not extended to base in some specimens. Anterior transverse and median longitudinal impressions distinct. Proepipleura visible from above.
Elytra. Oval, convex; striae complete or obliterated to various degrees or absent (stria 1 always complete), punctate or not, punctures shallowly to deeply impressed. Preapical epipleural plica (Fig. 22, ep) displaced outward, interrupting outline of elytra.
Hind wings. Macropterous or brachypterous.
Underside (Fig. 2). Metasternum with a row of setiferous punctures along margin of coxal cavity, median contact with abdomen dividing metacoxae. Suture between abdominal sterna III and IV obliterated at middle. Pleurite VII with small projection(Fig. 22, app) fitting into elytral epipleural plica (ep, ).
Legs. Slender, protibia (Figs 23, 24) with basal half of ventral surface with a group of setae, protarsomeres in males slightly dilated.
Male genitalia (Figs 25, 26). Phallus lightly sclerotized; basal bulb with basal orifice (bo) oval, wide; basal projection (bp) distinct, apical margin curved; basal apophysis (ba) moderate; median portion tubular; apical portion formed by two lamellae (rl and ll). Endophallus pubescent in apical portion; basal sclerite (bs) clearly developed. Parameres subequal in length, the right one wider, both with 4-5 apical setae.
Female genitalia (Figs 27, 28). Ovipositor: gonocoxa (gc) unsegmented, very slender, with a pair of setae at apex, surface with micropores; laterotergites (lt) thinly sclerotized, flat and asetose, sclerotized border thin and folded in basal portion with a small anterior part; mediotergite (mt) wide, its width delimited by external borders of laterotergites. Reproductive tract: bursa copulatrix (bc) elongate; spermathecal duct (spd) =distance from apex of bursa copulatrix to insertion point of spermathecal gland duct (spgd) relatively short or long, spermathecal gland duct elongate; spermatheca (sp) various in thickness, length, and conformation; see description of this structure for each of the two species-groups.
Cuticular sculpture: The absence of a submarginal band of microsculpture on the proepisternum has been used to differentiate Semiardistomis from Ardistomis but the character state is not diagnostic since the band has been lost in at least two lineages of Ardistomis (Valdes, 2009).
The microsculpture on the elytral disc has a diagnostic value, the absence of microsculpture being the apotypic state.
Vestiture: Hirsuteness is a common phenomenon in this genus. In the two species groups, this apotypic character state appears, its origin independently developed. In the puncticollis group, three species have hirsute structures: Semiardistomis puncticollis from southern U.S.A. (supraorbital area of the head capsule, pronotal disc, elytral disc, profemur and abdominal sternum VII), Semiardistomis pilosellus (Kult) from southern South America (elytral disc and profemur) and Semiardistomis subglabra (van Emden) from South America (profemur). In the labialis group, two species have hirsute structures: Semiardistomis viridis from southern U.S.A. (pronotal disc and elytral disc) and Semiardistomis propinquus (Putzeys) from Mexico (elytral disc),
Elytra: In this genus, I regard the state of having the striae complete and impunctate as plesiotypic, being present in both species groups.
Hind wings: Macropterous except Semiardistomis laevistriatus brachypterous.
Legs: Protarsomeres in males so slightly dilated that this feature is useless for sex recognition.
Male genitalia: Phallic structure enhances definition of the two species groups of Semiardistomis. See details, provided in the species-group diagnoses. The differences in the phallic structures are too small to separate the species within each group, a situation also known in other clivinine genera, such as Schizogenius Putzeys (see
The first instar of Semiardistomis was described by
Based primarily on male and female genitalic features, the species of Semiardistomis are arranged in two species-groups designated as the puncticollis and labialis group, respectively.
Of the 20 known species of Semiardistomis only those occurring in U.S.A. (Semiardistomis viridis (Say) and Semiardistomis puncticollis (Dejean)) are represented by large series of specimens in collections (
Members of this genus are difficult to identify in part because of a shortage of useful external features, and in part because the male and female genitalia, so useful in distinguishing between species groups, are not useful within each group. Moreover, some of the characters traditionally used to separate the species have been found in large samples to vary extensively. Some samples are a mixture of combinations from extreme patterns considered as different species; being impossible to demonstrate if those populations are hybrids or simply polymorphs, while in other putative species, allopatric populations exhibit no evident differences. In the labialis group in southern South America, the species limits are especially difficult to assess probably because the group is young and composed of species with a great power of dispersal. Without direct evidence of the state of gene flow between known populations, it is almost impossible to testify in favor of the reproductive isolation of each species described or in other cases to designate subspecies. I will follow a criterion with a practical value, so maybe for some species, taxonomic determination will denote unintentionally a combination of extreme characters shared by more than one biological species.
(Figs 57–59)The range of this genus is co-extensive with the range of the Ardistomina.
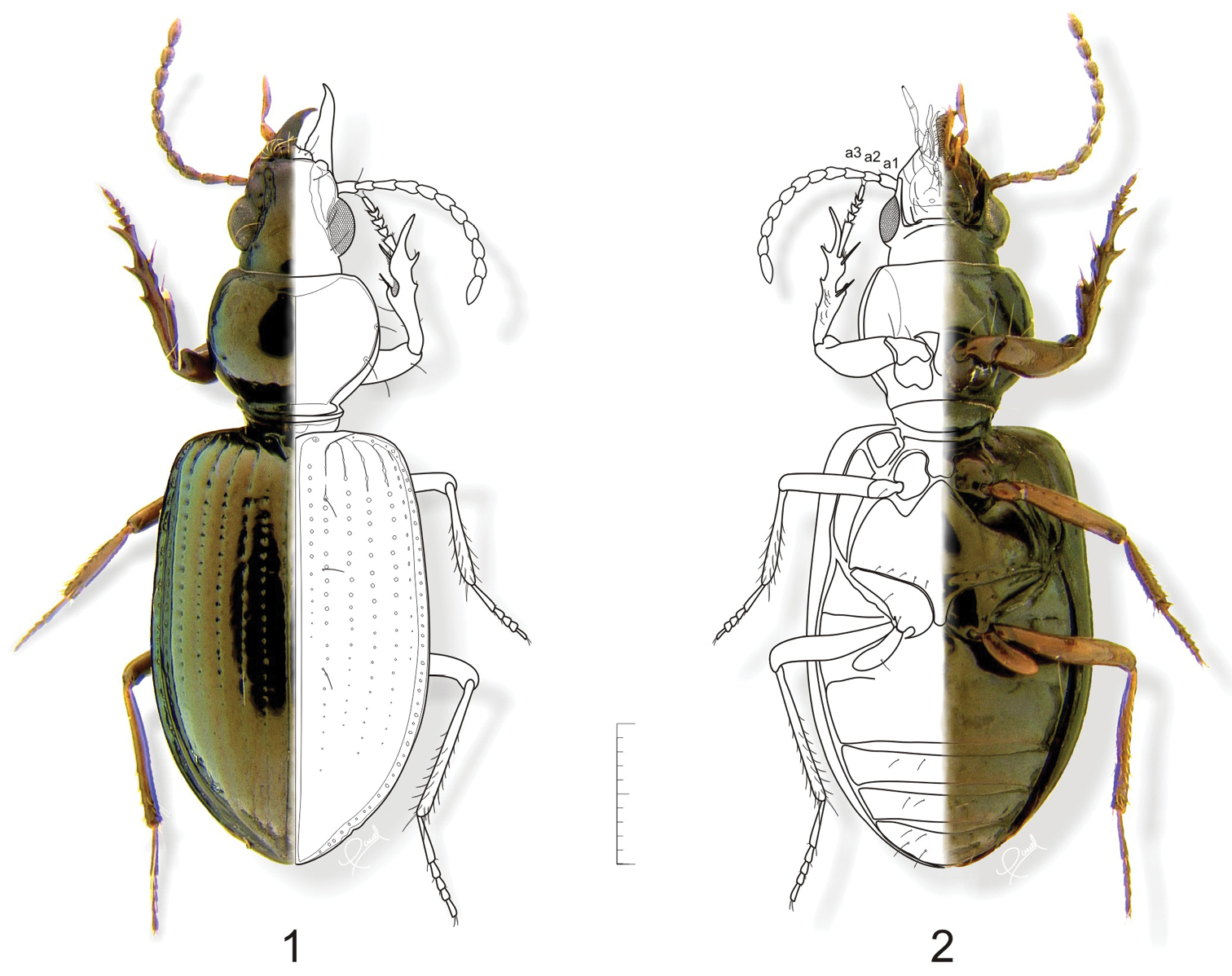
Semiardistomis labialis (Chaudoir). 1 Dorsal aspect 2 Ventral aspect. Legend: a1 a2 a3 antennomeres 1–3, respectively. Scale bar 1 mm.
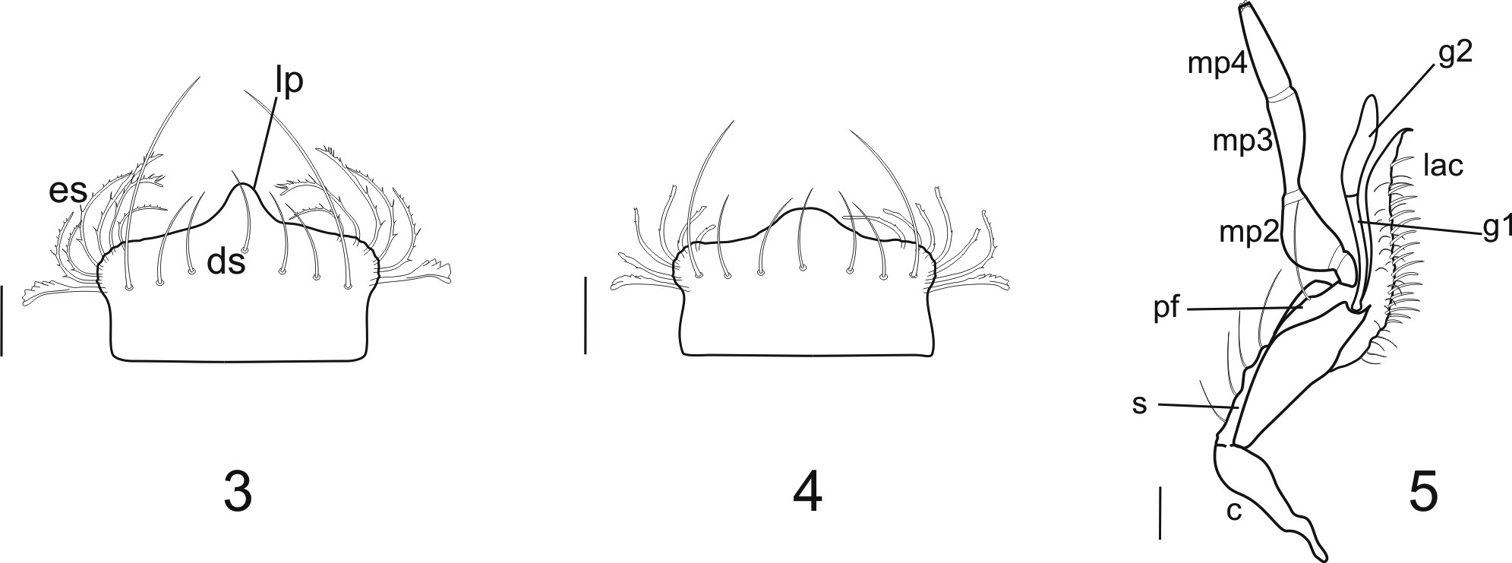
3–4 Labrum (dorsal aspect) 3 Semiardistomis glabratus (Putzeys) 4 Semiardistomis labialis (Chaudoir) 5 RightMaxilla (dorsal aspect) of Semiardistomis labialis (Chaudoir). Legend : Labrum: ds dorsal seta es epipharyngeal seta lp labral dentiform projection. Maxilla: c cardo g1 galeomere 1 g2 galeomere 2 lac lacinia mp2 mp3 mp4 maxillary palpomeres 2–4 respectively pf palpifer s stipes. Scale bar 0.1 mm.
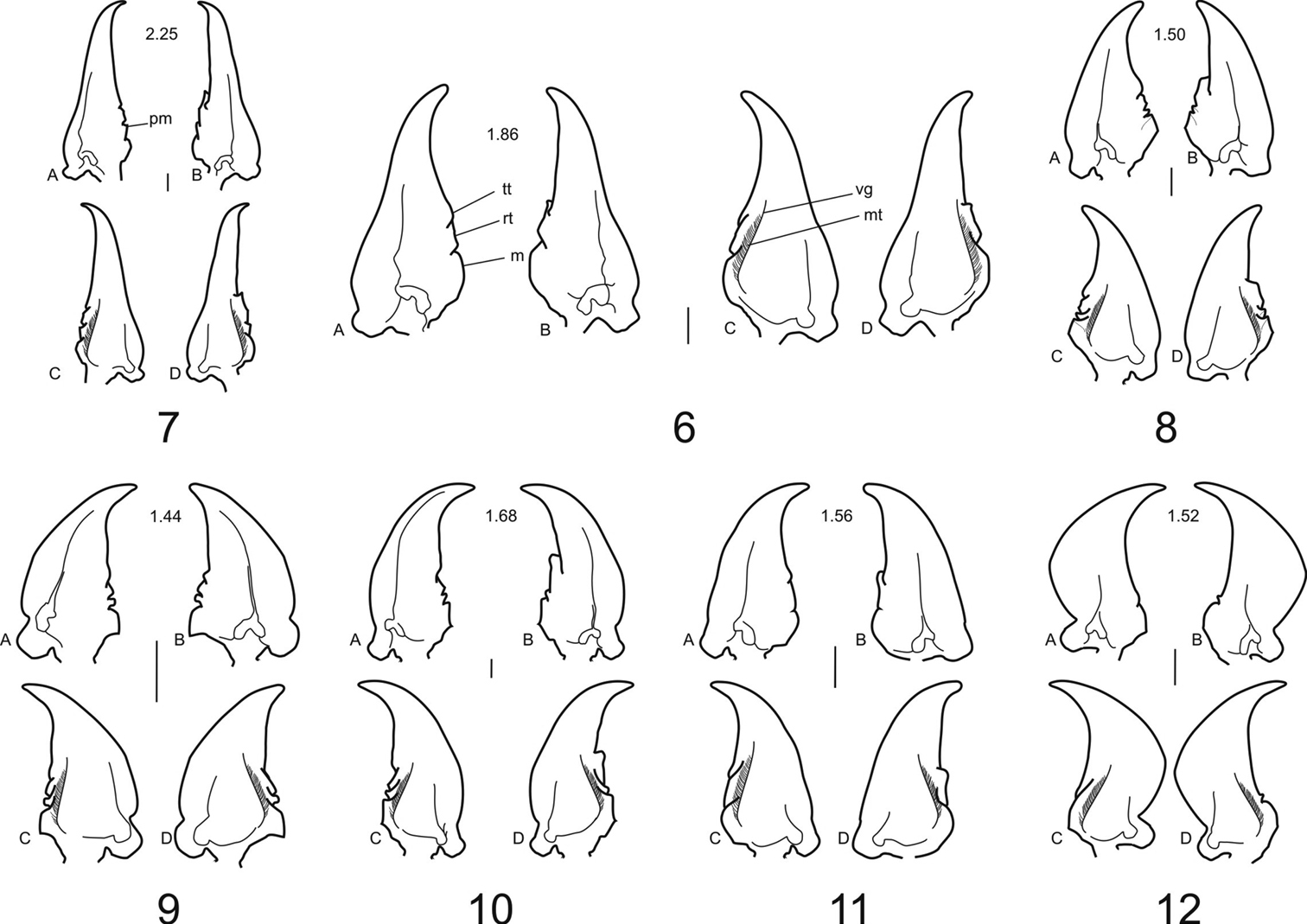
Mandibles ofArdistomina, Clivinina and Dyschiriina with length/width ratios: A left, dorsal aspect B right, dorsal aspect C left, ventral aspect and D right, ventral aspect) 6 Semiardistomis labialis (Chaudoir) 7 Ardistomis fasciolatus Putzeys 8 Aspidoglossa mexicana (Chaudoir) 9 Oxydrepanus rufus (Putzeys) 10 Clivina dentipes Dejean 11 Schizogenius arimao Darlington 12 Dyschiriodes larochellei Bousquet. Legend: Occlusal teeth: m molar pm premolar rt retinacular tt terebral. Ventral surface: vg ventral groove mt microtrichia. Scale bar 0.1 mm.
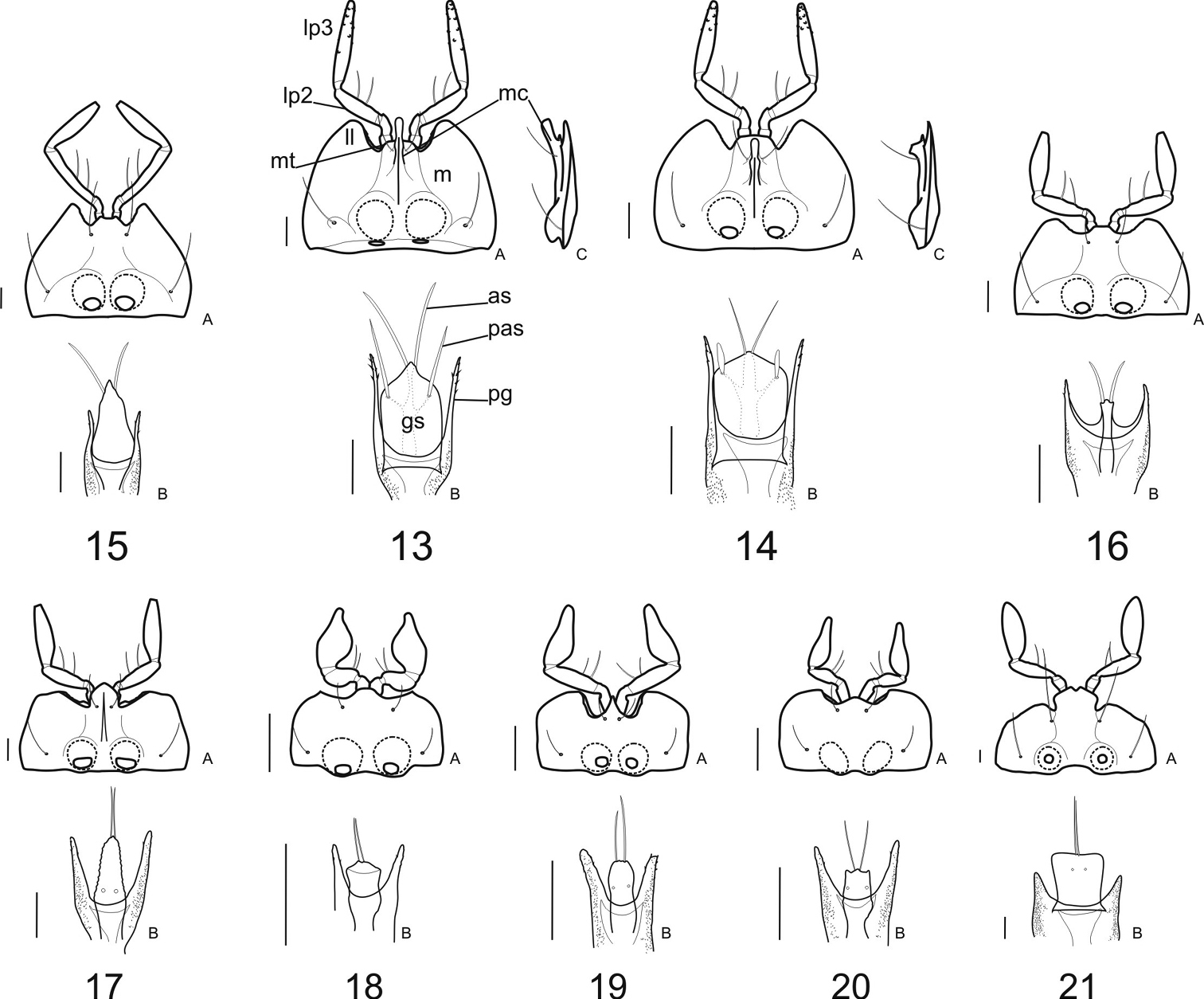
Labium ofClivinini (ventral aspects): A mentum and palpi B ligula C mental carina, lateral aspect. 13–14 Genus Semiardistomis: 13 S. glabratus (Putzeys) 14 Semiardistomis labialis (Chaudoir) 15 Ardistomis fasciolatus Putzeys 16 Aspidoglossa mexicana (Chaudoir) 17 Clivina dentipes Dejean 18 Oxydrepanus rufus (Putzeys) 19 Schizogenius arimao Darlington 20 Ancus excavaticeps (Putzeys) 21 Nyctosyles planicollis (Reiche). Legend: as apical setae of glossal sclerite gs glossal sclerite lp2 labial palpomere 2 lp3 labial palpomere 3 ll lateral lobes mc median carina m mentum mt mental tooth pas preapical seta of glossal sclerite pg paraglossa. Scale bar 0.1 mm.

22 Body extremity (lateral aspect) of Semiardistomis labialis (Chaudoir) 23–24 Protarsus (ventral aspect) of Semiardistomis 23 Semiardistomis labialis (Chaudoir) 24 Semiardistomis glabratus (Putzeys). Legend: app abdominal epipleural projectionof abdominal sternum VII ep elytral plica. Scale bar 1 mm.
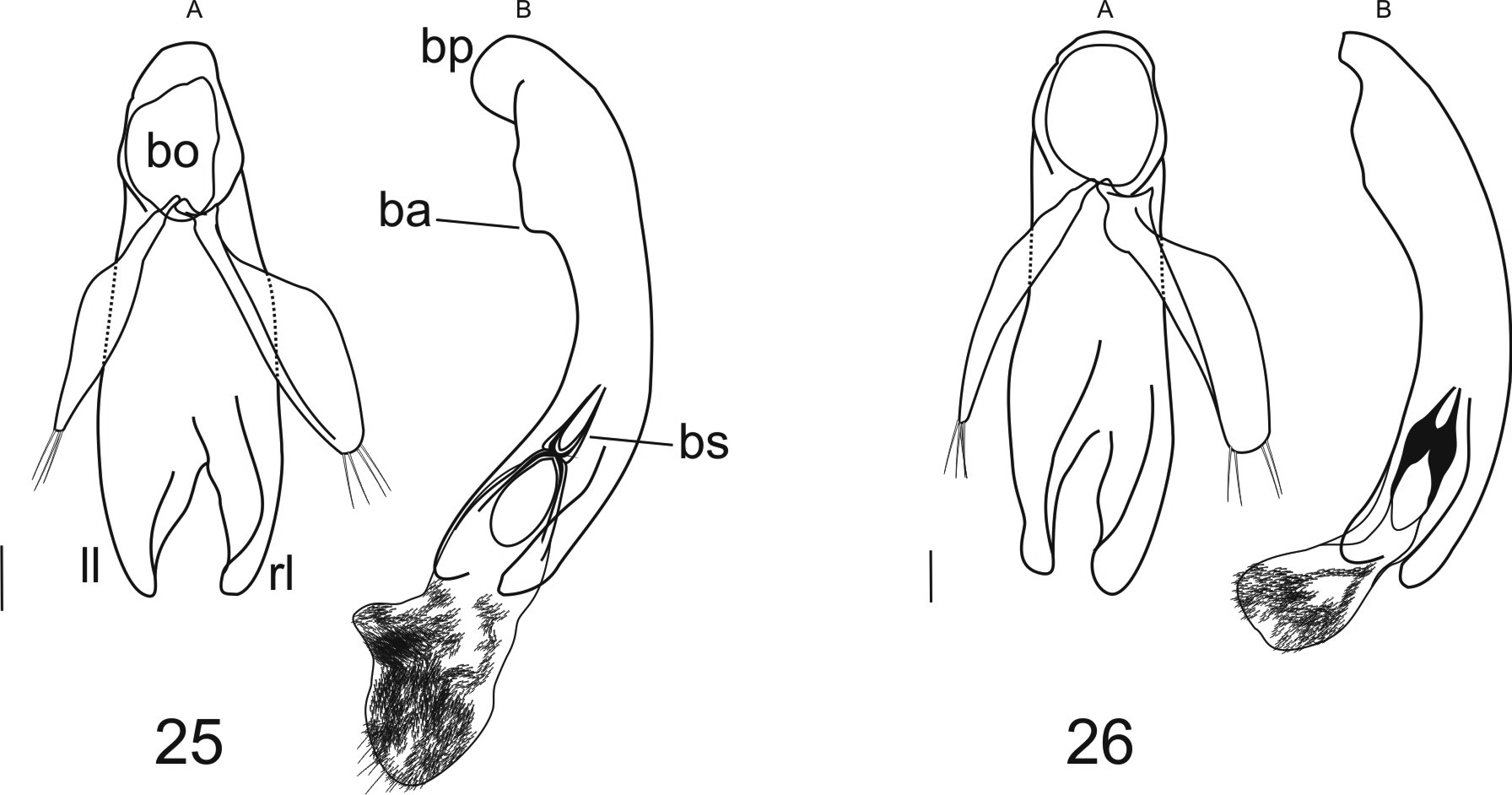
Male genitalia of genus Semiardistomis: A phallus and parameres in dorsal aspect B phallus and endophallus everted in lateral aspect 25 Semiardistomis labialis (Chaudoir) 26 Semiardistomis glabratus (Putzeys). Legend: ba basal apophysis bo basal opening bp basal projection bs basal sclerite ll left lobe rl right lobe. Scale bar 0.1 mm.
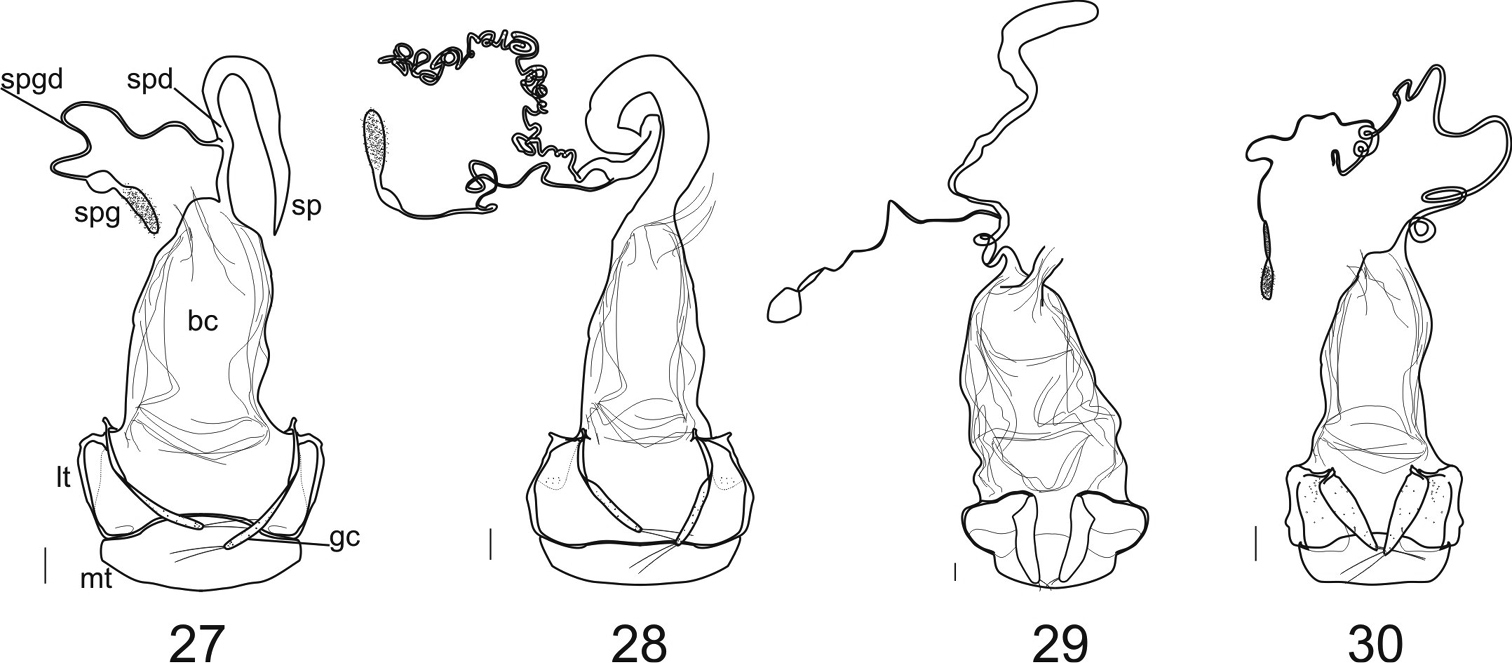
Female genitalia (ventral aspect). of Ardistomina 27 Semiardistomis glabratus (Putzeys) 28 Semiardistomis labialis (Chaudoir) 29 Ardistomis fasciolatus Putzeys 30 Aspidoglossa mexicana (Chaudoir). Legend: bc bursa copulatrix gc gonocoxa (gonocoxite 1 and 2, fused) lt laterotergite mt mediotergite sp spermatheca spd spermathecal duct spg spermathecal gland spgd spermathecal gland duct.Scale bar 0.1 mm.
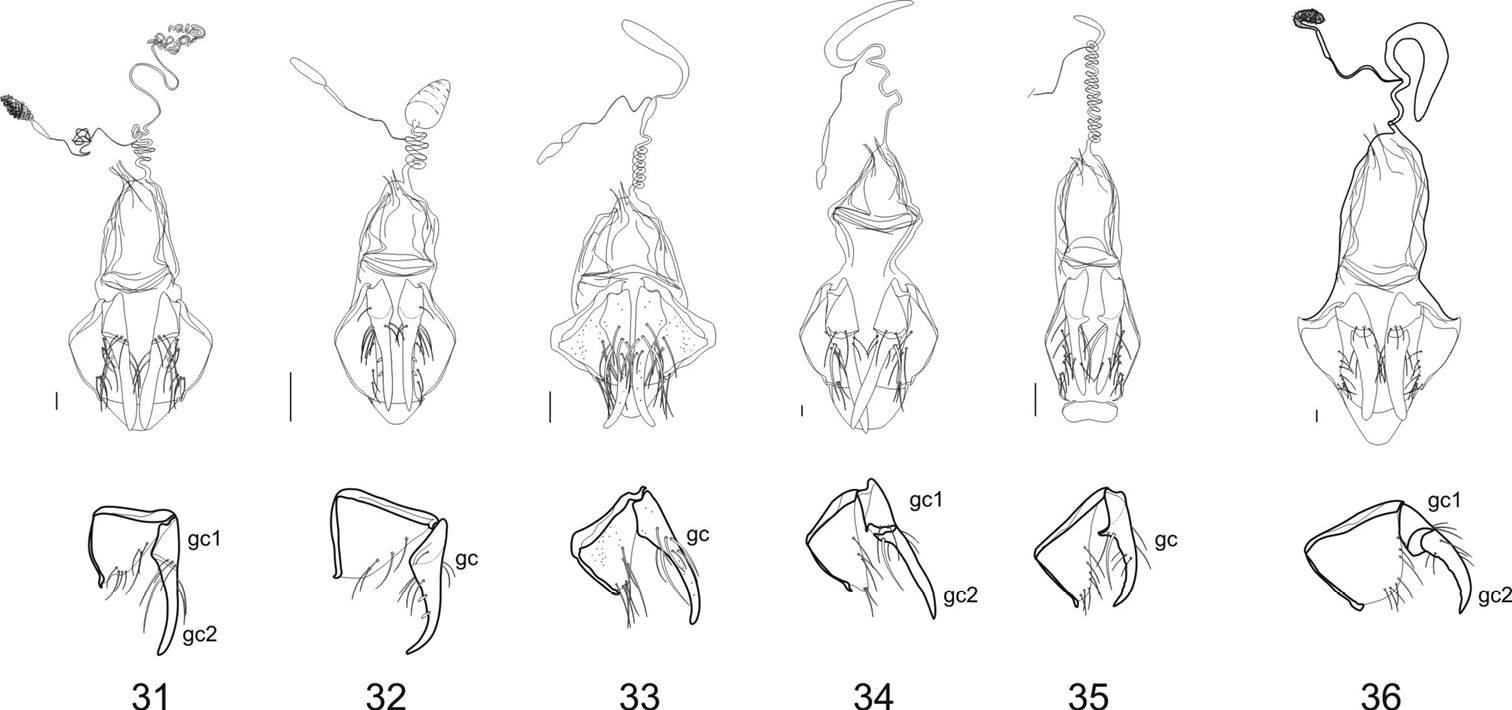
Female genitalia (dorsal aspect) and gonocoxites /laterotergites (lateral aspect) of Clivinina 31 Clivina dentipes Dejean 32 Oxydrepanus rufus (Putzeys) 33 Schizogenius arimao Darlington 34 Nyctosyles planicollis (Reiche) 35 Ancus excavaticeps (Putzeys) 36 Obadius insignis Burmeister. Legend: See legend for Figs 27–30, except gonocoxite 1 (gc1) and. gonocoxite 2 (gc2) Scale bar 0.1 mm.
| 1 | Elytral disc plurisetose | 2 |
| 1’ | Elytral disc with setiferous punctures in interval 3, or sometimes also in interval 5 | 5 |
| 2(1) | Profemur plurisetose | 3 |
| 2’ | Profemur with the usual setae | 4 |
| 3(2) | Pronotal disc plurisetose (along margin and on disc). Habitus as in Fig. 48. USA | Semiardistomis puncticollis(Dejean) |
| 3’ | Pronotal disc with the usual 2 pairs of marginal setae. Habitus as in Fig. 44. South America | Semiardistomis pilosellus (Kult) |
| 4(2’) | Pronotal disc plurisetose along margin. Habitus as in Fig. 55. USA | Semiardistomis viridis (Say) |
| 4’ | Pronotal disc with the usual 2 pairs of marginal setae. Habitus as in Fig. 54. Mexico | Semiardistomis propinquus (Putzeys) |
| 5(1’) | Elytron with two setiferous punctures on interval 3; humerus rounded; brachypterous. Habitus as in Fig. 47 Guadeloupe | Semiardistomis laevistriatus (Fleutiaux & Sallé) |
| 5’ | Elytron with three or more setiferous punctures on interval 3; macropterous | 6 |
| 6(5’) | Elytron with three setiferous punctures on interval 3 | 7 |
| 6’ | Elytron with more than three setiferous punctures on interval 3 | 12 |
| 7(6) | Elytral striae impunctate. Habitus as in Fig. 52. South America | Semiardistomis pallipes (Dejean) |
| 7’ | Elytral striae punctate | 8 |
| 8(7’) | Elytral disc with isodiametric mesh pattern | 9 |
| 8’ | Elytral disc smooth | 10 |
| 9(8) | Elytral striae impressed only in basal fifth, punctures deep along three-quarters of elytra, surface completely covered with isodiametric mesh pattern. South America | Semiardistomis deletus (Putzeys) |
| 9’ | Elytral striae obliterated in apical third, punctures moderately deep, surface with isodiametric mesh pattern, evanescent in central area of elytral disc in some specimens. South America | Semiardistomis flavipes (Dejean) |
| 10(8’) | Elytral striae obliterated in apical fifth, punctures moderately deep. Habitus as in Fig. 51. West Indies | Semiardistomis cyaneolimbatus (Chevrolat) |
| 10’ | Elytral striae impressed only in basal fifth | 11 |
| 11(10’) | Elytral punctures deeply impressed in basal two-thirds of elytra. Habitus as in Figs 1, 2. Central America | Semiardistomis labialis (Chaudoir) |
| 11’ | Elytral punctures moderately impressed in basal third of elytra. Southern South America | Semiardistomis semipunctatus (Dejean) |
| 12(6’) | Elytral disc surface completely covered with isodiametric mesh pattern. Habitus as in Fig. 46. South America | Semiardistomis maindroni (Kult) |
| 12’ | Elytral disc surface smooth | 13 |
| 13(12’) | Elytral striae complete; impunctate | 14 |
| 13’ | Elytral striae incomplete | 17 |
| 14(13) | Front between antennal lobes with transverse rugae. Habitus as in Fig. 50. Southern Brazil | Semiardistomis rugosus |
| 14’ | Front between antennal lobes smooth | 15 |
| 15(14’) | Elytron with five setiferous punctures on interval 3; body size more than 7 mm. Habitus as in Fig. 49. Peru | Semiardistomis major n. sp. |
| 15’ | Elytron with four setiferous punctures on interval 3; body size less than 6 mm | 16 |
| 16(15’) | Pronotum markedly cordate, 1.25 times wider than long; humeri square, sides subparallel. Habitus as in Fig. 37. Colombia | Semiardistomis cordicollis (Putzeys) |
| 16’ | Pronotum subcordate, 1.14 times wider than long; shoulders rounded. Habitus as in Fig. 38. South America. | Semiardistomis exspectatus n. sp. |
| 17(13’) | Elytron with eight setiferous punctures in interval 3 | 18 |
| 17’ | Elytron with four setiferous punctures in interval 3 | 19 |
| 18(17) | Elytron with three setiferous punctures in interval 5. Habitus as in Fig. 39. Brazil | Semiardistomis darlingtoni (Kult) |
| 18’ | Elytron with interval 5 asetose. Habitus as in Fig. 40. South America | Semiardistomis subglabra (van Emden) |
| 19(17’) | Elytral striae 3, 4, 5 and 6 impressed in basal half of disc, stria 2 present. Body length around 4 mm. Habitus as in Fig. 45. South America | Semiardistomis jedlickai (Kult) |
| 19’ | Elytral striae 3, 4, 5 and 6 impressed in basal third of disc, stria 2 absent. Body length around 6 mm. Habitus as in Fig. 41. South America | Semiardistomis glabratus (Putzeys) |
This group is defined by the following: mentum with median carina (Fig. 13C) extended slightly distad anterior margin of lateral lobes (except Semiardistomis puncticollis with this structure (putatively) reduced); mental pit organs opened through oval orifices on the mental–submental suture. Males with wide phallus, undefined basal bulb (Fig. 26B) and a prominent, thickly sclerotized endophallic basal sclerite (Fig. 26A).
Female reproductive tract (Fig. 27; cf. Fig. 28) with spermatheca duct (spd) relatively short; spermatheca (sp) bent in its middle portion, wider from this point and with acute apex.
Included speciesSemiardistomis cordicollis (Putzeys, 1846)
Semiardistomis darlingtoni (Kult, 1950)
Semiardistomis exspectatus sp. n.
Semiardistomis glabratus (Putzeys, 1866)
Semiardistomis jedlickai (Kult, 1950)
Semiardistomis laevistriatus (Fleutiaux & Sallé, 1889)
Semiardistomis maindroni (Kult, 1950)
Semiardistomis major sp. n.
Semiardistomis pilosellus (Kult, 1950)
Semiardistomis puncticollis (Dejean, 1831)
Semiardistomis rugosus (Putzeys, 1866)
Semiardistomis subglabra (van Emden, 1949)
http://species-id.net/wiki/Semiardistomis_cordicollis
Figs 37, 57Holotype at MHNP, pinned, labeled: handwritten “Lectotype Ardistomis cordicollis Putz. By Erwin 1976”/ “Lectotype Clivina labialis Chd. Des. S.W. Nichols 1984”/ printed in red paper “lectotype”/ handwritten in box “cordicollis Putzeys. Nlle Grenade C. Reiche”.
Given by
Elytral surface smooth, striae complete, impunctate, 4 setiferous punctures in interval 3, humeri square, sides subparallel. Pronotum notably cordiform. Size given by Putzeys as 4.5 mm.
Fig. 37.
The unique type seen appears to be a member of the puncticollis group. I have been unable to examine the genitalia, but the general structure is similar to the species of this group (I observed 4 setiferous punctures in interval 3 contrary to Putzeys’ original description which mentions 3 punctures). The species appears to be most similar to Semiardistomis exspectatus sp. n.
(Fig. 57). This species is known only from an unspecified location in Colombia.

37 Semiardistomis cordicollis (Putzeys) 38 Semiardistomis exspectatus sp. n. Dorsal aspect. Scale bar 1 mm.
http://species-id.net/wiki/Semiardistomis_darlingtoni
Figs 39, 57Holotype male (ADVA), glued on pinned card, genitalia attached in microvial, labeled: handwritten “darlingtoni 57 Det. K. Kult”/ “Darlingtoni Kt. det. K.Kult”/ “ Brasilia Jatahy (Goyas)”/ printed on red paper “TYPE”.
Given by
Elytral surface smooth, impunctate, elytral striae impressed in basal third, three setiferous punctures on interval 5 and 8 in interval 3, stria 2 absent, prominent humeral tooth at junction of third stria with marginal channel. Profemur glabrous. Abdominal sternum VII with 5+5 setiferous punctures.
dorsal aspect, as in Fig. 39.
Variation of measurements (mm) and ratios for Semiardistomis darlingtoni (n=1) are: HL = 0, 70; PL = 1, 22; PW = 1, 38; EL = 3, 42; EW = 2, 21; SBL = 5, 4; PW/EW = 0, 62; PW/PL = 1, 13; PL/EL = 0, 36; EW/EL = 0, 64.
(Fig. 57). Known only from the type area.
The holotype is teneral. The species is closely related to Semiardistomis subglabra.

39 Semiardistomis darlingtoni (Kult) (teneral) 40 Semiardistomis subglabra (van Emden). Dorsal aspect. Scale bar 1 mm.
urn:lsid:zoobank.org:act:1C3A9901-2441-4E08-9860-9DBE99ADBA6F
http://species-id.net/wiki/Semiardistomis_exspectatus
Figs 38, 57Male (USNM), glued on pinned point, genitalia attached in microvial, labeled: printed "Peru: Madre de Dios Rio Manu, BIOLAT Biol. Sta. Pakitza, 356 m, 24 June 1993 11°56'47"S, 071°17'00"W T. L. Erwin & F. Pfuno"/ "Treading in leaf litter half buried in quick- silt at edge of Que. Trepanatrunco Tr. Tachigali"/ "135 Lot 510"/ "BIOLAT/ COLE 000019005".
19 exemplars (USNM), 2 exemplars (PVCCu) labeled same as the holotype. 14 exemplars (USNM) labeled: printed “Peru: Madre de Dios Rio Manu, BIOLAT Biol. Sta. Pakitza, 356 m, 11 June 1993 11°56'47"S, 071°17'00"W T. L. Erwin & F. Pfuno"/ "Tr. Castañal/ 12.5 Que. Paujil on sand among small stones near stream edge at night Lot 437".
PERU, Madre de Dios, Rio Manu, 11°56'47"S, 071°17'00"W.
After have a picture of zoogeographic patterns for some characters from the rest of described species of Semiardistomis, appearance of this new form filled a gap in the scenario of the group puncticollis . So the specific epithet derived from Latin adjective, meaning expected.
Elytral surface smooth, striae complete, impunctate, 4 setiferous punctures on interval 3, shoulders rounded. Profemur glabrous. Abdominal sternum VII with 4+4 setiferous punctures.
Fig. 38.
Variation of measurements (mm) and ratios for Semiardistomis exspectatus sp.n. (n=10) are: HL = 0, 64–0, 68–0, 74; PL = 1, 09–1, 18–1, 28; PW = 1, 28–1, 35–1, 44; EL = 3, 20–3, 37–3, 68; EW = 2, 14–2, 32–2, 66; SBL = 4, 93–5, 24–5, 70; PW/EW = 0, 58; PW/PL = 1, 14; PL/EL = 0, 35; EW/EL = 0, 69.
Body piceous with green reflections, mouthparts and antennae testaceous, legs ferrugineous.
Frons, gena, antennal lobes, pronotum, proepisternum and elytra smooth. Prosternum with microsculptural mesh pattern isodiametric. Metasternum and abdominal sterna with microsculpture in form of a shallow transverse mesh pattern.
Anterior marginal setae on pronotal disc equidistant between anterior angles and posterior setae. Elytral disc with 4 setae in interval 3. Abdominal sternum VII with ambulatory setae near base, 2 on each side; inner pair of preapical setae equidistant each other. Profemur glabrous. Ventral surface of protibia with many setae on basal half.
Clypeus with anterior margin concave medially; lateral lobes distinct, projected at the same level of anterior margin. Frontal impressions deep and wide. Supraantennal lobes with median sulci across their length. Eyes prominent. Antennomere 2 shorter than antennomere 3; antennomeres 4–10 about 1.9 times longer than wide.
Mentum with median carina extended distad slightly beyond anterior margin of lateral lobes; pit organs opened through oval orifices to the mental- submental suture.
Pronotum ovate, lateral border reaching base.
Elytra oval, humeri curved, striae complete in their length, impunctate; visible humeral tooth at junction of third stria with marginal channel.
Metathoracic wingsmacropterous.
Genitalia as described for the puncticollis species–group.
This species exhibits many plesiotypic character states for the puncticollis species-group.
(Fig. 57). Widespread in the northern parts of South America, east of the Andes mountain range.
Records from labels suggest that the habitat of this species is typical of most species of the genus, living on loose soil adjacent to fresh water bodies. Furthermore the species seems to prefer sandy areas with leaves or stones. Tenerals were collected in August.
In addition to the type material, I have seen 8 specimens from the following localities.ECUADOR. Pichincha Alturiquin Rio Toachi 9–1 Feb 1999 (PBPC, 2). PERU. Loreto. Boca del Rio Samiria 16 Aug 1991 (USNM, 1); Loreto, Hamburgo, Boca del Inglés 150m 23 Aug 1991 (USNM, 1); Loreto, Pithecia 14 Aug 1989 (USNM, 2); Loreto Cmp. Terry 14 May 1990 (USNM, 1). TRINIDAD AND TOBAGO: Trinidad Chatham 17 Jun 1980 (NMNH, 1)
http://species-id.net/wiki/Semiardistomis_glabratus
Figs 3, 13, 24, 26, 27, 41–43, 57Lectotype, here designated, female (IRSNB), glued on cardboard pinned, genitalia attached in microvial, labeled: green paper handwritten "A. glabrata Pz. C Mvideo. (A. 21ª)"/ printed "Soc. Ent. Belg. Coll. Putzeys"/ printed "Syntype."
Paralectotypes, here designated:onefemale (IRSNB) labeled as Lectotype; one exemplar (MHNP) labeled: handwritten in box “glabrata Putz. Mvideo”.
Holotype of Semiardistomis balthasari male (ADVA), glued on cardboard pinned, genitalia attached in microvial, labeled: handwritten “Ardistomis balthasari Kt. Det. K. Kult 1948”/ “balthasari Kt. det. K.Kult”/ printed in red paper “TYPE”/ printed “San Ignacio Missions”.
Given by
Body piceous with green brassy reflections. Elytral surface smooth, striae reduced to basal third, impunctate, 4 setiferous punctures on interval 3, stria 2 absent. Profemur glabrous. Abdominal sternum VII with 5+5 setiferous punctures.
dorsal aspect, as in Fig. 41.
Labrum, dorsal aspect, as in Fig. 3. Labium, as in Fig. 13A; ligula, enlarged, as in Fig. 13B.
male protarsus as in Fig. 24.
as in Fig. 26.
as in Fig. 27.
Variation of measurements (mm) and ratios for Semiardistomis glabratus (n=7) are: HL = 0, 70–0, 76–0, 80; PL 1, 44–1, 51–1, 60; PW 1, 53–1, 68–1, 83; EL 3, 56–3, 84–4, 13; EW 2, 50–2, 60–2, 73; SBL 5, 76–6, 11–6, 53; PW/EW 0, 65; PW/PL 1, 11; PL/EL 0, 39; EW/EL 0, 68.
Variation of measurements (mm) and ratios for Semiardistomis glabratus identified in collections as Semiardistomis balthasari (n= 11) are: HL = 0, 67–0, 78–0, 86; PL 1, 12–1, 32–1, 44; PW 1, 34–1, 49–1, 63; EL 3, 33–3, 66–3, 87; EW 2, 08–2, 36–2, 56; SBL 5, 15–5, 76–6, 11; PW/EW 0, 63; PW/PL 1, 13; PL/EL 0, 36; EW/EL 0, 65.
Data from labels suggest that the species lives along standing water bodies and swamps.
(Fig. 57). Widespread in South America, east of the Andes mountain range, from northern Brazil, north of the Equator, to Uruguay, south of the Tropic of Capricorn.
ARGENTINA. San Ignacio Misiones (ADVA, 2) Rio Salado (IRSNB, 12) Cordoba Argüello Sept 58 (MHNP, 8). BRAZIL. Oberaba (IRSNB, 9). PERU. Loreto 1 km E Hamburgo, Boca del Ingles Camp, 150 m 23 Aug 1991 (USNM, 8). Loreto, Pithecia 14 Aug 1989 (USNM, 5). Madre de Dios Pakitza 07 Oct 1990 (USNM, 5). URUGUAY. Montevideo (IRSBN, 5) (MHNP, 2). VENEZUELA. Amazonas, Dpto Rio Negro 28 Jan 1985 (USNM, 4).
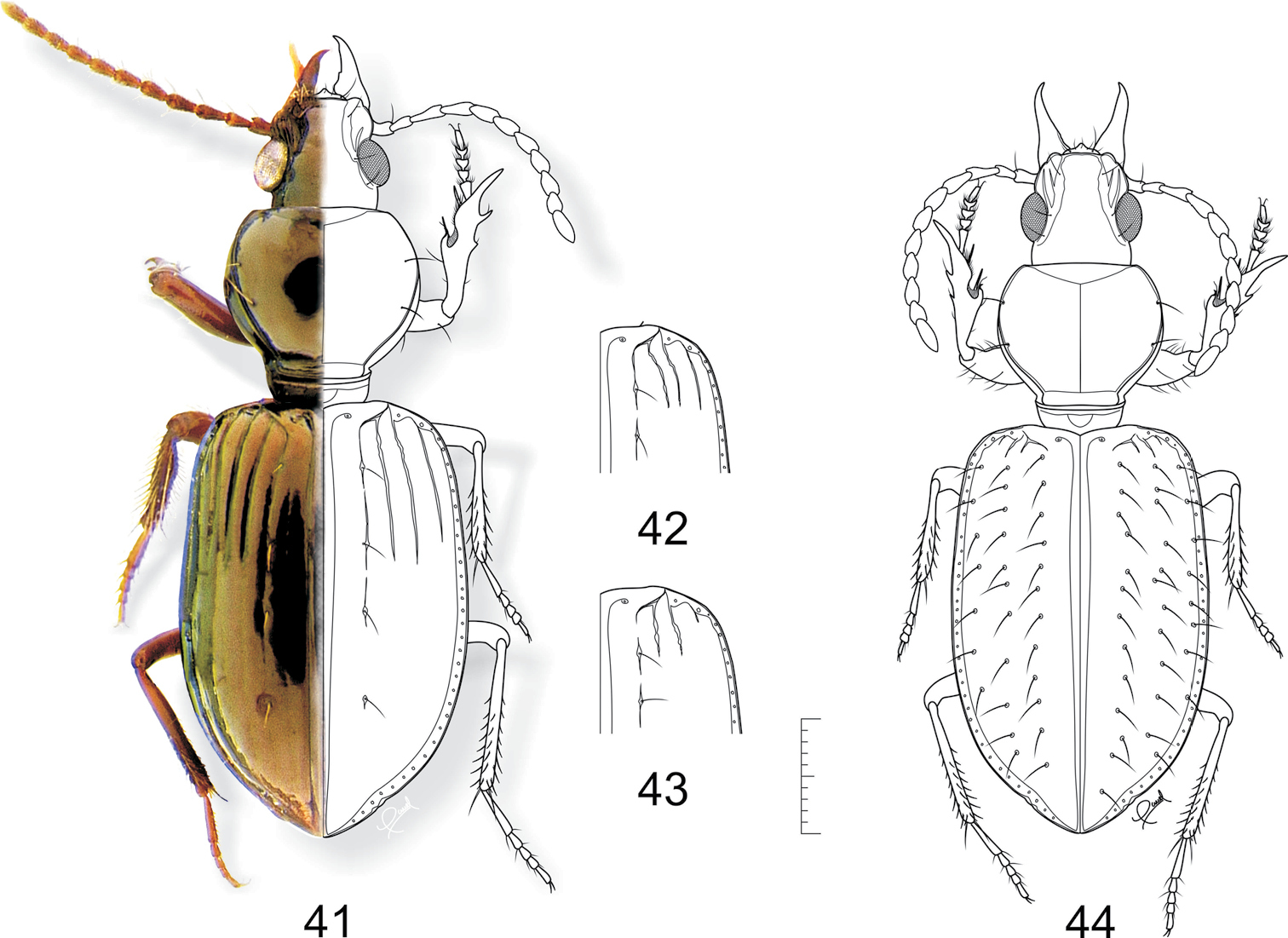
41–43 Semiardistomis glabratus (Putzeys) 41exemplar from Peru, Loreto 42 exemplar from Argentina, Misiones 43 exemplar from Uruguay, Montevideo 44 Semiardistomis pilosellus (Kult). Dorsal aspect. Scale bar 1 mm.
http://species-id.net/wiki/Semiardistomis_jedlickai
Figs 45, 57Holotype at ADVA, glued on pinned card (specimen without genitalia), labeled: handwritten “Ardistomis jedlickai Kt. Det. K. Kult 1948”/ “jedlickai Kt. det. K.Kult”/ printed in red paper “TYPE”/ printed “Corumba Matt. Grosso.”
Given by
Body piceous with green brassy reflections. Elytral surface smooth, striae distinct in basal half only, impunctate, 4 setiferous punctures on interval three, stria 2 distinct. Profemur glabrous. Abdominal sternum VII with 4+4 setiferous punctures. Size small.
dorsal aspect, as in Fig. 45.
Variation of measurements (mm) and ratios for Semiardistomis jedlickai (n=6) are: HL = 0, 50–0, 51–0, 53; PL 0, 96–1, 00–1, 07; PW 0, 99–1, 17–1, 28; EL 2, 30–2, 55–2, 62; EW 1, 70–1, 82–1, 92; SBL 3, 82–4, 07–4, 19; PW/EW 0, 64; PW/PL 1, 16; PL/EL 0, 39; EW/EL 0, 71.
(Fig. 57). The known range of this species is confined to central South America.
In addition to holotype: BRAZIL. Matto Grosso Jacare P. N. Xingu at ligth (USNM, 11) PERU. Madre de Dios Pakitza 14 Nov 1990 (USNM, 6). Madre de Dios Rio Manu BIOLAT Pakitza 356m 24 Jun 1993 (USNM, 5).
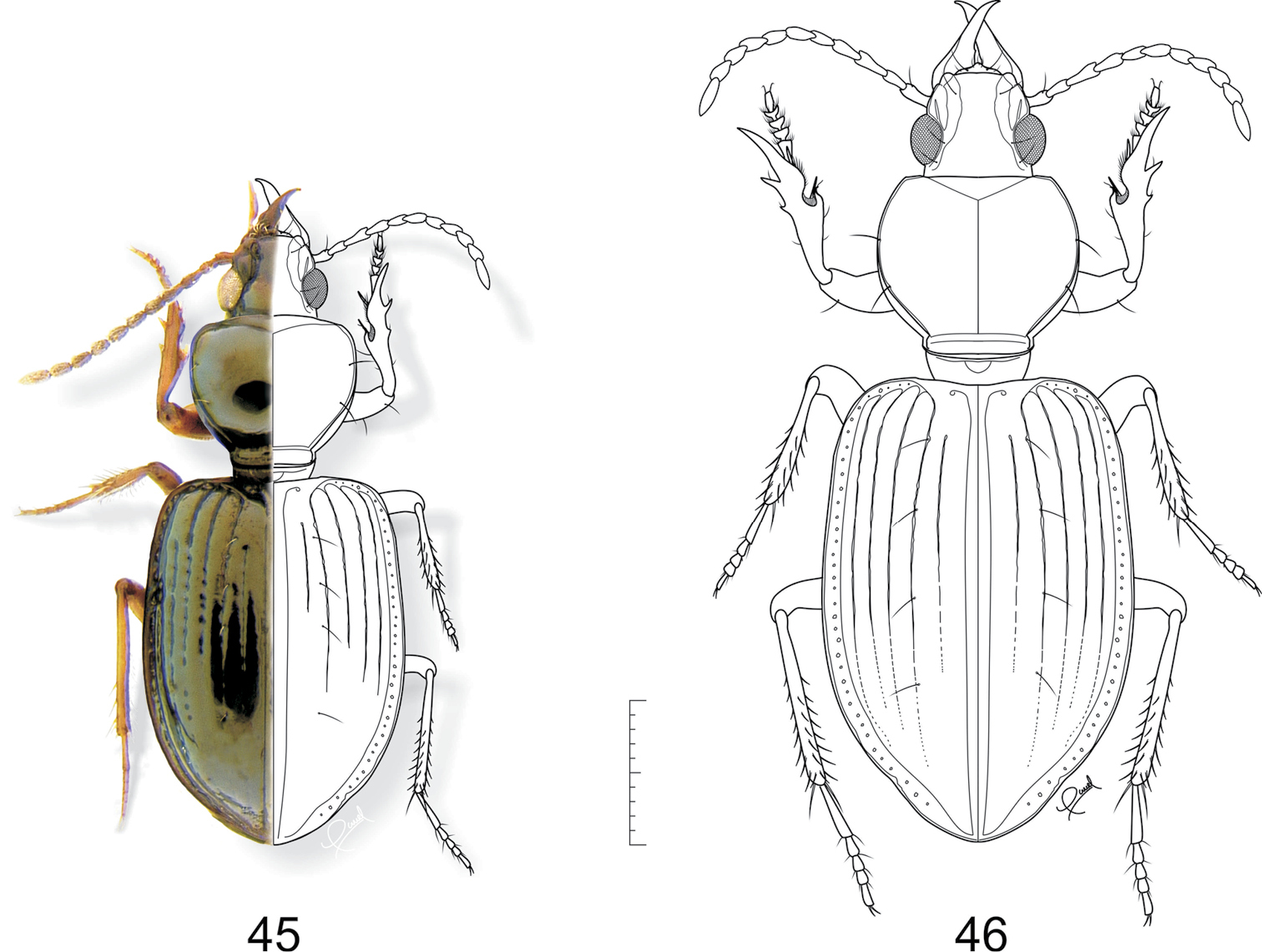
45 Semiardistomis jedlickai (Kult) 46 Semiardistomis maindroni (Kult). Dorsal aspect. Scale bar 1 mm.
http://species-id.net/wiki/Semiardistomis_laevistriatus
Figs 47, 57Lectotype, here designated, male (MHNP), glued on pinned card, labeled: printed “Type”; printed “Guadeloupe Delauney”; handwritten “Ardistomis laevistriata Fleut. & S. type”; “Ardistomis laevistriata Fleut. 47 det. K. Kult type”; “Lectotype Ardistomis laevistriata F. & S. des. S. W. Nichols 1984.”
Paralectotype, here designated, female (MHNP), glued on pinned card, genitalia attached in microvial, labeled: handwritten on paper circle "Guadeloupe Delauney"; printed "ex Musaeo A. Sallé 1897"; "Paralectotype Ardistomis laevistriata F. & S. des. S. W. Nichols 1984”.
Given by
Body ferrugineous dark brown. Elytral surface smooth, striae obliterated in apical fifth, impunctate, 2 setiferous punctures on interval 3, stria 2 distinct, shoulders rounded. Metathoracic wings reduced. Profemur glabrous. Abdominal sternum VII with 5+5 setiferous punctures.
dorsal aspect, as in Fig. 47.
Variation of measurements (mm) and ratios for Semiardistomis laevistriatus (n=4) are: HL = 0, 65–0, 68–0, 70; PL = 1, 13–1, 20–1, 28; PW = 1, 33–1, 39–1, 45; EL = 2, 95–3, 05–3, 15; EW = 2, 00–2, 05–2, 10; SBL = 4, 73–4, 93–5, 13; PW/EW = 0, 68; PW/PL = 1, 16; PL/EL = 0, 39; EW/EL = 0, 67.
(Fig. 57). Restricted to the Islands of Guadeloupe in the Lesser Antilles.
GUADELOUPE. Guadeloupe L. Dufau (ADVA, 1) Bains–Jaunes (USNM, 2). Guadeloupe Marie (MHNP, 1).

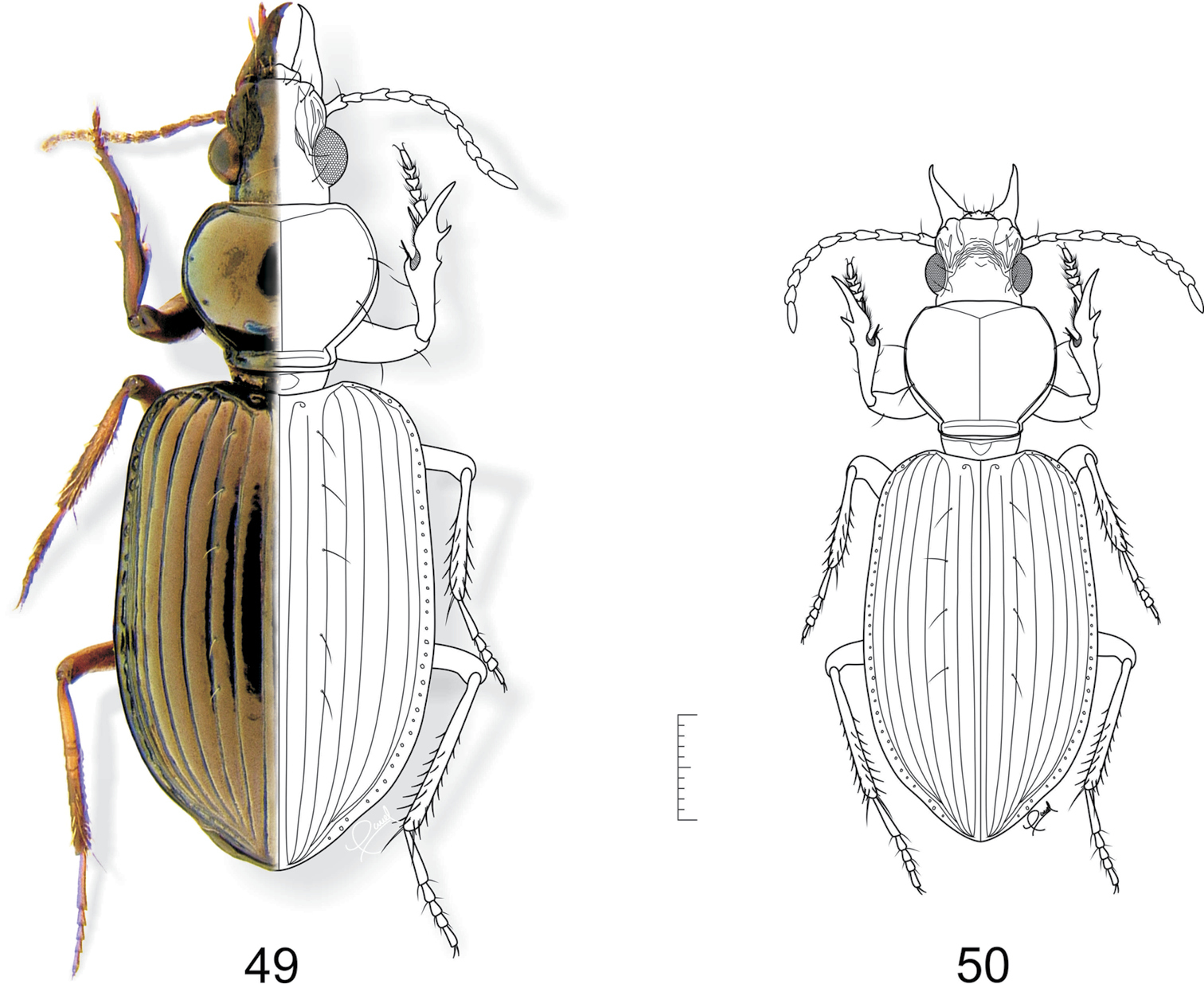
47 Semiardistomis laevistriatus (Fleutiaux & Salle) 48 Semiardistomis puncticollis (Dejean). Dorsal aspect. Scale bar 1 mm.
http://species-id.net/wiki/Semiardistomis_maindroni
Figs 46, 57Holotype at ADVA, glued on cardboard pinned, genitalia attached in microvial, labeled: handwritten “Ardistomis maindroni Kt. Det. K. Kult 1948”/ “maindroni Kt. det. K.Kult”/ printed in red paper “TYPE”/ printed “Cuyaba Matt. Grosso”.
Paratypes. two exemplars labeled as holotype (not checked) designated by Kult in “collection of National Museum of Paris (ex Maindron- Babault)”.
Given by
Body piceous with green reflections. Elytral surface covered with isodiametric mesh pattern, striae obliterated in apical half, impunctate, 4 setiferous punctures on interval 3, stria 2 present. Profemur glabrous. Abdominal sternum VII with 4+4 setiferous punctures.
dorsal aspect, as in Fig. 46.
Variation of measurements (mm) and ratios for Semiardistomis maindroni (n=9) are: HL = 0, 63–0, 66–0, 71; PL = 1, 18–1, 25–1, 32; PW = 1, 31–1, 44–1, 57; EL = 3, 10–3, 38–3, 64; EW = 2, 02–2, 22–2, 40; SBL = 4, 93–5, 30–5, 60; PW/EW = 0, 65; PW/PL = 1, 15; PL/EL = 0, 37; EW/EL = 0, 67.
(Fig. 57). The known range of this species extends from central Brazil southward of the Tropic of Capricorn to central Argentina.
ARGENTINA. Cordoba Arguello Nov 1958 (MHNP, 5); Tucuman Nov 1951 (MHNP, 6); Chaco prov. Capitán Solari env. 3–7 Feb. 2004 (PBPC, 2). BRAZIL. Porta da Bocca am Lagua de Jigua Pernambuco Jul 1937 (BMNH, 4).
urn:lsid:zoobank.org:act:86C18C17-48F7-404A-B43B-76F1707EE250
http://species-id.net/wiki/Semiardistomis_major
Figs 49, 57Male (USNM), glued on pinned point, genitalia attached in microvial, labeled: printed "Peru: Loreto, Cmp. S. Branch, 11 May 90 75°20'W, 05°12'S T. L. Erwin Coll"/ "Running at night on muddy bank of Rio Samiria–Igapó forest soil organic, grasses"/ "ADP 94456".
8 exemplars (USNM), 2 exemplars (PVCCu) labeled as the holotype.
The specific epithet is a Latin adjective alluding to the large size of the adults.
Elytral surface smooth, striae complete, impunctate, 4 setiferous punctures on interval 3, shoulders rounded. Profemur glabrous. Abdominal sternum VII with 5+5 setiferous punctures.
dorsal aspect, as in Fig. 49.
Variation of measurements (mm) and ratios for Semiardistomis major sp. n. (n=11) are: HL = 0, 93–0, 97–0, 99; PL = 1, 50–1, 61–1, 66; PW = 1, 92–1, 98–2, 02; EL = 4, 47–4, 71–4, 80; EW = 3, 13–3, 23–3, 33; SBL = 6, 90–7, 28–7, 46; PW/EW = 0, 61; PW/PL = 1, 23; PL/EL = 0, 34; EW/EL = 0, 69.
Body piceous with green reflections, mouthparts and antennae testaceous, legs ferrugineous.
Frons, gena, antennal lobes, pronotum, prosternum, proepisternum, elytra, metasternum and abdominal segments smooth.
Anterior marginal setae on pronotal disc closer to posterior setae than to anterior angles. Elytral disc with 5 setae in interval 3. Abdominal sternum VII with setae near base, 2 on each side; distance between inner pair of preapical setae 0.6 times that between inner and outer setae. Profemur glabrous. Ventral surface of protibia with many setae on basal half.
Clypeus with anterior margin concave medially; lateral lobes distinct, behind level of anterior margin. Frontal impressions deep and wide. Supraantennal lobes with median sulci impressed in basal half. Eyes prominent. Antennomere 2 shorter than antennomere 3; antennomeres 4–10 about 1.8 times longer than wide.
Mentum with median carina extended distad slightly beyond anterior margin of lateral lobes; pit organs opened through oval orifices to the mental- submental suture.
Pronotum ovate, lateral border extended to base.
Elytra oblong, humeri square, striae complete in their length, impunctate.
Metathoracic wingsmacropterous.
Genitalia in both sexes as described for the puncticollis species–group.
(Fig. 57). Known only from the region of Loreto, in Amazonian Peru.
Records from labels suggest that the habitat of Semiardistomis major is typical of most members of the genus, living on loose soil adjacent to fresh water bodies. In this case data indicate a river bank with organic soil. Active at night.
PERU. Loreto. Rio Samiria Cocha Shinguito near lake margin 25 Aug 1991 (USNM, 1).
. 47 Semiardistomis major sp.n. 50 Semiardistomis rugosus (Putzeys). Dorsal aspect. Scale bar 1 mm.
http://species-id.net/wiki/Semiardistomis_pilosellus
Figs 44, 57Holotype female (IRSNB), pinned, genitalia attached in microvial, labeled: green paper handwritten “pilosella. P.”/ green paper handwritten “Mvid Arech.”/ handwritten “Ardistomis pilosella 47 det. K. Kult”/ printed “Soc. Ent. Belg. Coll. Putzeys” red paper printed “TYPE”/ printed “Holotype Ardistomis pilosella Kult des. S. W. Nichols 1984”
Paratypes. Male (ADVA), pinned, genitalia attached in microvial, labeled: handwritten “Montevideo leg. Arechavaleta”/ green paper handwritten “Mvid Arech.”/ handwritten “Ardistomis pilosella 47 det. K. Kult”/ handwritten “pilosellus Kt det. K. Kult”/ printed “Soc. Ent. Belg. Coll. Putzeys”/ red paper printed “Paratype”.
Given by
Body piceous with green brassy reflections. Elytral surface smooth, striae 2–6 erased, surface covered with setiferous punctures. Profemur setose. Abdominal sternum VII with 5+5 setiferous punctures.
dorsal aspect, as in Fig. 44.
Variation of measurements (mm) and ratios for Semiardistomis pilosellus (n=6) are: HL = 0, 70–0.75–0.80; PL = 1, 16–1, 23–1, 31; PW = 1, 30–1, 35–1, 44; EL = 3, 40–3, 50–3, 64; EW = 2, 12–2, 19–2, 27; SBL = 5, 26–5, 48–5, 70; PW/EW = 0, 62; PW/PL = 1, 17; PL/EL = 0, 37; EW/EL = 0, 64.
(Fig. 57). This species is found south of the Rio de la Plata–Uruguay basin, south of the Tropic of Capricorn.
ARGENTINA. Pronunciamiento Prov. Entre Rios (MHNP, 12).
http://species-id.net/wiki/Semiardistomis_puncticollis
Figs 48, 57Lectotype at (MHNP), glued on tip pinned, labeled: printed “Lectotype”; handwritten “36”; “puncticollis Dej. Am. bor. Dej.”; “Lectotype Clivina puncticollis Dej. des. S. W. Nichols 1984.”.
Given by
Body piceous with green reflections. Head with several supraorbital setae. Pronotum with more than two premedial setiferous punctures, pronotal disc with several setiferous punctures. Elytral surface completely smooth, striae absent, punctures deeply impressed, setiferous. Abdominal sternum VII plurisetose. Wing atrophy reported by
dorsal aspect, as in Fig. 48.
(Fig. 57). Known only from southeastern United States and adjacent northeastern Mexico (
USA. Enterprise, Fla 13 5 (USNM, 2) Crescent City, Fla (USNM, 1). L. Harney, Fla. May (USNM, 2) San Jacinto Co. Tex, Jun 68 (USNM, 3) Etats unis Guer (MHNP, 5).
http://species-id.net/wiki/Semiardistomis_rugosus
Figs 50, 57Lectotype, here designated, female (IRSNB), glued on pinned card, head and pronotum missing, genitalia attached in microvial, labeled: green paper handwritten “rugosa. Pz. Sta Cath.(Dhr.)”/ printed “Soc. Ent. Belg. Coll. Putzeys”/ “Syntype”/ “Lectotype Ardistomis rugosa Putz. des. S. W. Nichols 1984”.
Given by
Body piceous. Elytral surface smooth, impunctate, elytral striae complete, 4 setiferous punctures on interval 3, stria 2 absent, prominent humeral tooth at junction of third stria with marginal channel. Profemur glabrous. Abdominal sternum VII with 5+5 setiferous punctures.
dorsal aspect, as in Fig. 50.
Variation of measurements (mm) and ratios for Semiardistomis rugosus (n=2) are: HL = 0, 71; PL = 1, 35; PW = 1, 51; EL = 3, 72–3, 73–3, 75; EW = 2, 34–2, 42–2, 50; SBL = 5, 78; PW/EW = 0, 64; PW/PL = 1, 12; PL/EL = 0, 36; EW/EL = 0, 63.
(Fig. 57). Known only for the Santa Catarina region in Brazil.
BRAZIL. Hansa Humboldt Sta. Catarina 3. 1937 (BMNH, 1).
http://species-id.net/wiki/Semiardistomis_subglabra
Figs 40, 57Holotype female (BMNH), glued on pinned point, without head, genitalia attached in microvial, labeled: green paper handwritten “subglabra Emd.”/ red paper handwritten “Ardistomis subglabra type. Emd.”/ green paper handwritten “amazon”/ printed red circle “Holotype”/ printed “Gesch. 2. 1934 von Prf. Noesske”/ printed “F. van Emden Bequest. B.M. 1960- 129”/ printed “Holotype Ardistomis subglabra Emden des. S.W. Nichols 1984”.
Holotype of Semiardistomis vlastae male (ADVA), glued on pinned card, genitalia attached in microvial, labeled: handwritten “vlastae Kt. det. K.Kult”/ printed in red paper “TYPE”/ printed “Chaco Pilcomayo 5-6 Jan. 1938 Tippmann”.
Given by van
Body piceous with green bluish reflections. Elytral surface smooth, impunctate, striae distinct on basal third only, impunctate, 8 setiferous punctures on interval 3, stria 2 absent. Profemur hirsute. Abdominal sternum VII with 6+6 setiferous punctures.
dorsal aspect, as in Fig. 40.
Variation of measurements (mm) and ratios for Semiardistomis subglabra (n=4) are: HL = 0, 60–0, 70–0, 78; PL = 1, 12–1, 21–1, 28; PW = 1, 23–1, 34–1, 44; EL = 3, 50–3, 64–4, 75; EW = 2, 05–2, 25–2, 40; SBL = 5, 28–5, 52–5, 78; PW/EW = 0, 60; PW/PL = 1, 11; PL/EL = 0, 33; EW/EL = 0, 62.
(Fig. 57). Widespread in South America.
ARGENTINA. Chaco Pilcomayo 5-6 Jan 1938 (ADVA, 1). BRAZIL. Pantanal Jun 1986 (PBPC, 2).
This group is defined by the following: mentum with median carina (Fig. 14C) not extended distad, beyond anterior margin of mental tooth; mental pit organs (Fig. 14A) opened through oval orifices in basal part of mentum; male genitalia with a slender phallus (Fig. 25A) with clearly delineated basal bulb (bp) and basal sclerite (bs) of the endophallus thin and slightly sclerotized (Fig. 25B); female reproductive tract (Fig. 28; cf. Fig. 27) with spermatheca duct (spd) relatively long, wide, bent in its distal portion, spermatheca very narrow, elongate, markedly convoluted in a series of tight twists. The species–group name is based on that of Semiardistomis labialis, the type species of Semiardistomis, being this group nominotypical.
Morphological noteAs in the puncticollis group, the mental carina shows a reduction pattern northward with this prolongation markedly reduced in Semiardistomis viridis.
Included speciesSemiardistomis cyaneolimbatus (Chevrolat, 1863)
Semiardistomis deletus (Putzeys, 1846)
Semiardistomis flavipes (Dejean, 1831)
Semiardistomis labialis (Chaudoir, 1837)
Semiardistomis pallipes (Dejean, 1831)
Semiardistomis propinquus (Putzeys, 1866)
Semiardistomis semipunctatus (Dejean, 1831)
Semiardistomis viridis (Say, 1823)
http://species-id.net/wiki/Semiardistomis_cyaneolimbatus
Figs 51, 53, 59Lectotype, here designated, male (HECO), pinned, labeled: green paper handwritten “Cuba Poey”/ handwritten “ Aspidoglossa cyaneolimbata Chevt. Type Cuba 1461.”/ printed “Chevrolat Carabidae. Fr. V. d. Poll. Pres. 1909, E. B. Poulton”/ printed “type col: 132 Ardistomus cyaneolimbata Chevr. Hope Dept. Oxford”/ printed “Lectotype Ardistomis cyaneolimbatus Chev. des. S.W. Nichols 1984”..
Given by
Body piceous with green brassy reflections, pronotum with bluish reflections. Elytral surface smooth, striae complete but indistinct at apex, punctures moderately impressed, 3 setiferous punctures on interval 3. Abdominal sternum VII with 4+4 setiferous punctures.
dorsal aspect, as in Fig. 51.
Variation of measurements (mm) and ratios for Semiardistomis cyaneolimbatus (n=7) are: HL = 0, 52- 0, 59- 0, 60; PL = 1, 00- 1, 03- 1, 10; PW = 1, 10- 1, 18- 1, 30; EL = 2, 44- 2, 72- 2, 84; EW = 1, 60- 1, 75- 1, 88; SBL = 3, 96- 4, 34- 4, 54; PW/EW = 0, 67; PW/PL = 1, 14; PL/EL = 0, 38; EW/EL = 0, 64.
(Fig. 59): West Indies: Cuba, Cayman Islands and Haiti, in Hispaniola.
I collected this species along margins of the river Itabo, Isla de Pinos, Cuba (Fig. 53); the adults were active during the day on wet, organic soil. Other specimens were collected about 8 meters from the river margins under leaf litter in shady gallery forest.
CUBA. Cuba Rhl (IRSNB, 4); Cuba Chd (MHNP, 1); gundlachii Ptz. Cuba Type (MHNP, 1); Isla de Pinos, márgenes del Rio Itabo 23 Feb. 1999 (PVCCu: 14). HAITI. St. Dominique (MHNP, 1).
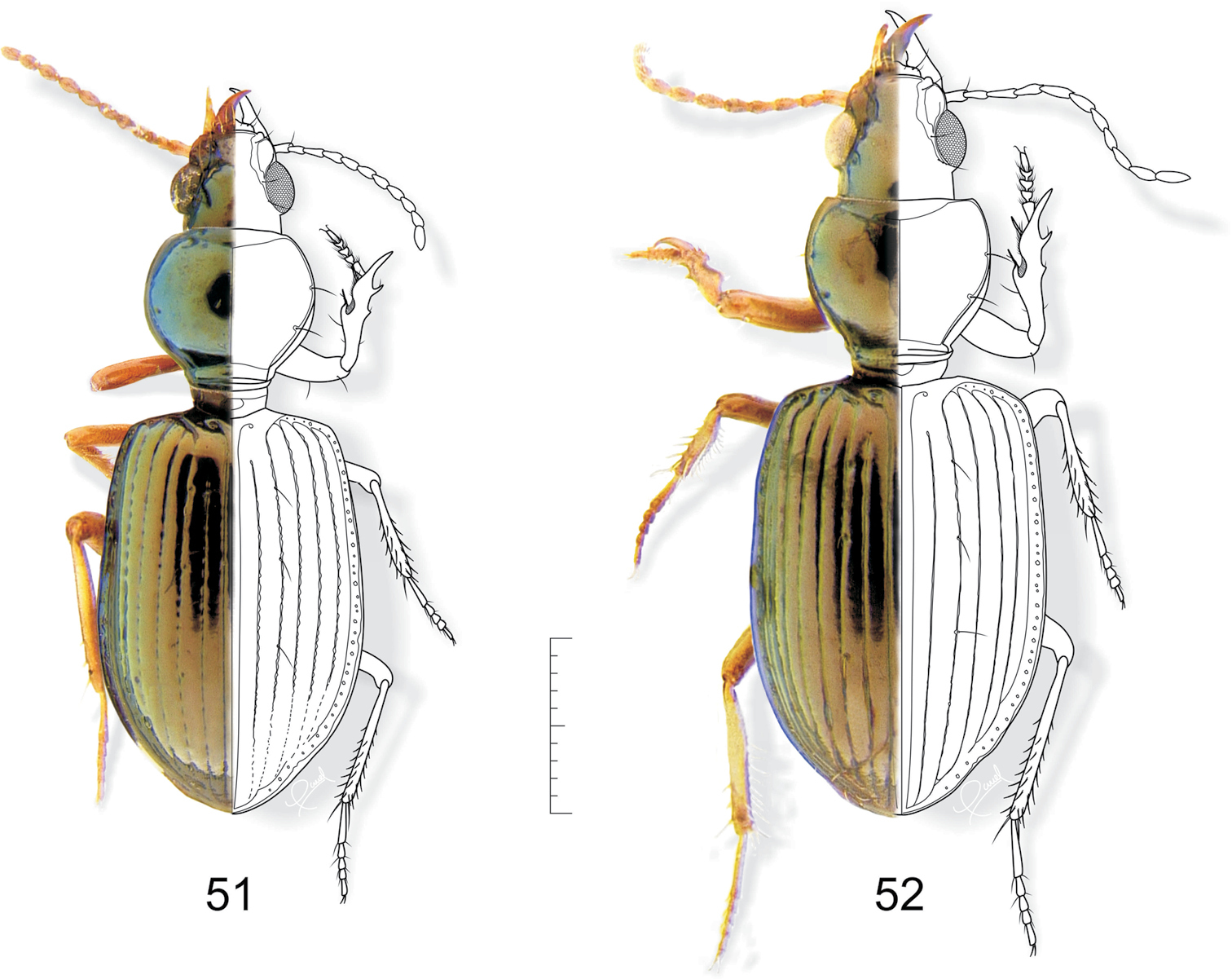
51 Semiardistomis cyaneolimbatus (Chevrolat) 52 Semiardistomis pallipes (Dejean). Dorsal aspect. Scale bar 1 mm.

Habitat of Semiardistomis cyaneolimbatus (Chevrolat) at Rio Itabo, Isla de Pinos, Cuba.
http://species-id.net/wiki/Semiardistomis_deletus
Figure 58Lectotype, here designated, at MHNP, glued on cardboard, labeled: green paper handwritten "Deleta Putz Bresil Reiche"/ handwritten " Lectotype Ardistomis deleta Putz. des. S.W. Nichols 1984"/ handwritten on box "Deleta Reiche Brésil Chevrol"/ red paper printed "Lectotype".
Paralectotype, here designated, at HECO, pinned, labeled: green paper handwritten “Campos”/ green paper handwritten “deleta (Reiche) Putz 130, 13 Brasilia type”/ red paper handwritten “deleta”/ printed “Chevrolat Carabidae. Fr. V. d. Poll. Pres. 1909, E. B. Poulton”/ printed “17”/ handwritten “Paralectotype Ardistomis deleta Putz. des. S.W. Nichols 1984”/ handwritten “Type col: 805 Ardistomus deleta Putzeys”.
Holotype of Semiardistomis emdeni female (ADVA), glued on pinned card, genitalia attached in microvial, labeled: handwritten “a. emdeni Kt. det. K.Kult, 1948”/ printed on red paper “TYPE”/ printed “Corumba Matt. Grosso”.
Given by
Body piceous with green brassy reflections. Elytral surface with isodiametric mesh pattern, striae impressed in basal eighth, punctures well impressed in basal half, 3 setiferous punctures on interval 3. Abdominal sternum VII with 4+4 setiferous punctures.
Variation of measurements (mm) and ratios for Semiardistomis deletus (n=17) are: HL = 0, 53- 0, 63- 0, 74; PL = 0, 80- 1, 06- 1, 20; PW = 0, 96- 1, 19- 1, 32; EL = 2, 56- 3, 11- 3, 60; EW = 1, 60- 1, 93- 2, 28; SBL = 3, 98- 4, 80- 5, 44; PW/EW = 0, 62; PW/PL = 1, 12; PL/EL = 0, 34; EW/EL = 0, 62.
Variation of measurements (mm) and ratios for type series of Semiardistomis emdeni (n=5) are: HL = 0, 58–0, 61–0, 64; PL = 1, 02–1, 05–1, 09; PW = 1, 12–1, 20–1, 28; EL = 2, 78–2, 95–3, 14; EW = 1, 76–1, 89–2, 02; SBL = 4, 38–4, 61–4, 86; PW/EW = 0, 64; PW/PL = 1, 15; PL/EL = 0, 36; EW/EL = 0, 64.
(Fig. 58). This species is known from southeastern South America, south of the Tropic of Capricorn.
ARGENTINA. Corrientes, Lago Ibera & Santo Tome 26 Sep 1997 (ADVA, 22); Rio Salado (IRSNB, 3); Misiones, Posadas Nov 1962 (MHNP, 6); Formosa Dec 1953 (MHNP, 11). BRAZIL. Nova Teutonia Dec. 1952 (MHNP, 8); Bresil (MHNP, 9); Sta Catarina Nova Teutonia Nov. 1972 (CAS, 17). PARAGUAY. Paraguari Ybycui, La Rosada 13 Apr. 1980 (USNM, 43); Dep. Central. Caacupé road Arroyo Yagua Resa 10 Apr. 1980 (USNM, 2) URUGUAY. Montevideo (IRSBN, 1).
http://species-id.net/wiki/Semiardistomis_flavipes
Fig. 58Holotypeat MHNP, pinned, labeled: handwritten “ Holotype Clivina flavipes Dej. des. S.W. Nichols 1984”/ handwritten on box “flavipes Dej. Brésil C. Dejean"/ red paper printed "Holotype".
Lectotype of Semiardistomis aenea, here designated, at IRSBN, glued on pinned card, labeled: green paper handwritten “R. Jan. Stevens”/ printed “Soc. Ent. Belg. Coll. Putzeys”. Paralectotypes, here designated: 4 at IRSBN labeled as Lectotype. 1 at MHNP labeled as Lectotype.
Holotype of Semiardistomis brittoni at ADVA labeled: handwritten “Ardistomis brittoni Kt. Det. K. Kult 1948”/ “brittoni Kt. det. K.Kult”/ printed in red paper “TYPE”/ handwritten “Argentina Tucuman”.
Holotype of Semiardistomis marani at ADVA labeled: handwritten “Ardistomis brittoni Kt. Det. K. Kult 1946”/ “brittoni Kt. det. K.Kult”/ printed in red paper “TYPE”/ handwritten “Bolivia 450m Sta Cruz Umg Buenavista Steinbach coll”.
Given by
Body piceous with green brassy reflections. Elytral surface with isodiametric mesh pattern, microlines more or less evanescent toward center of elytral disc, striae obliterated in apical fourth, punctures moderately impressed in basal two-thirds, 3 setiferous punctures on interval 3. Abdominal sternum VII with 4+4 setiferous punctures.
Variation of measurements (mm) and ratios for Semiardistomis flavipes (n=34) are: HL = 0, 42–0, 59–0, 68; PL = 0, 96–1, 03–1, 20; PW = 0, 95–1, 15–1, 34; EL = 2, 10–2, 86–3, 36; EW = 1, 44–1, 75–2, 00; SBL = 3, 55–4, 48–5, 16; PW/EW = 0, 66; PW/PL = 1, 11; PL/EL = 0, 36; EW/EL = 0, 61.
Variation of measurements (mm) and ratios for Semiardistomis brittoni (n=3) are: HL = 0, 54–0, 62–0, 67; PL 0, 84–1, 03–1, 09; PW 1, 09–1, 21–1, 28; EL 2, 75–3, 03–3, 20; EW 1, 76–1, 90–2, 02; SBL 4, 26–4, 68–4, 96; PW/EW 0, 63; PW/PL 1, 16; PL/EL 0, 34; EW/EL 0, 63.
Variation of measurements (mm) and ratios for Semiardistomis marani (n=1) are: HL = 0, 58; PL 0, 96; PW 1, 09; EL 2, 62; EW 1, 66; SBL 4, 16; PW/EW 0, 65; PW/PL 1, 13; PL/EL 0, 37; EW/EL 0, 63.
Species Semiardistomis brittoni and Semiardistomis marani measurements are a subset of that of Semiardistomis flavipes specimens studied. Variation in this species is found in the microsculpture on the elytral disc which varies from being distinct on the entire surface to being present only marginally, in the extent of the elytral striae, and the size of the punctures. Such variation is seen even within populations from the same locality.
(Fig. 58). The range of this species extends in central South America from western Brazil southward and westward to Paraguay, south of the Tropic of Capricorn.
Label data indicate that specimens of this species were collected during day at the margins of small water bodies, on sand–slit substrate.
ARGENTINA. Tucuman (ADVA, 3); Salta Sierra Tartagal 14 Nov. 2006 (ADVA, 6). BOLIVIA. Santa Cruz Buena Vista 20 Feb. 1999 (FSCA, 9) Santa Cruz 450m Buenavista (ADVA, 1). BRAZIL. Mato Grosso, Pantanal 1 Sep. 2000 (PBPC, 2); Rondonia 62 km SW Ariquemes 15- 19 May 1996 (FSCA, 9); Mato Grosso Varzea Grande, Cuiaba 5 May 1972 (FSCA, 10); Minas Geraes (HECO, 3); Rio Jan. (IRSBN, 4). PARAGUAY. Rio Confusa 18 Jan. 1937 (IRSBN, 144)
http://species-id.net/wiki/Semiardistomis_labialis
Figs 1, 2, 4- 6, 14, 22, 23, 25, 28, 59Lectotype, here designated, at MHNP, glued on point, labeled: green paper handwritten “Labialis Chaud Mexico Dej.”/ handwritten in box “labialis Chaud Mexique Chevrol”/ handwritten “38”/ “Lectotype Ardistomis labialis Chd. Det George E. Ball 72”/ “Lectotype Clivina labialis Chd. Des. S.W. Nichols 1984”/ printed in red paper “lectotype”/ printed in circle “Lectotype
Paralectotypes, here designated: two exemplars at MHNP labeled: handwritten in box “labialis Chaud Mexique Chevrol”; handwritten “Paralectotype Clivina labialis Chd. Des. S.W. Nichols 1984”.
Lectotype of Ardistomis tuspanensis, here designated, at IRSBN labeled: green paper handwritten “ Tuspanensis Tuspan (Chev.)”/ printed “Soc. Ent. Belg. Coll. Putzeys”/ handwritten “P. Basilewsky Ardistomus tuspanensis Putz”.
Given by
Body piceous with green reflections. Elytral surface completely smooth, striae continuously impressed in basal 1/8, punctures deeply impressed in basal 2/3, 3 setiferous pores on interval 3. Abdominal sternum VII with 4+4 setiferous punctures.
dorsal and ventral aspects, respectively, as in Figs 1, 2.
Labrum, dorsal aspect, as in Fig. 4. Labium, as in Fig. 14A; ligula, enlarged, as in Fig. 14B..
Legs: male protarsus as in Fig. 23.
Male genitalia: as in Fig. 25.
Female genitalia: as in Fig. 28.
Variation of measurements (mm) and ratios for Semiardistomis labialis from Mexico (n=11) are: HL = 0, 53–0, 58–0, 68; PL = 0, 90–1, 11–1, 25; PW = 1, 00–1, 23–1, 40; EL = 2, 65–3, 10–3, 40; EW = 1, 70–1, 94–2, 20; SBL = 4, 10–4, 80–5, 23; PW/EW = 0, 63; PW/PL = 1, 10; PL/EL = 0, 36; EW/EL = 0, 63.
Variation of measurements (mm) and ratios for Semiardistomis labialis from Costa Rica (n=11) are: HL = 0, 54–0, 60–0, 68; PL 1, 00–1, 07–1, 20; PW 1, 10–1, 21–1, 36; EL 2, 60–2, 86–3, 20; EW 1, 68–1, 86–2, 16; SBL 4, 14–4, 54–5, 08; PW/EW 0, 65; PW/PL 1, 13; PL/EL 0, 38; EW/EL 0, 65.
Variation of measurements (mm) and ratios for Semiardistomis labialis identified in collections as Semiardistomis tuspanensis (included syntypes) (n= 5) are: HL = 0, 55–0, 60–0, 68; PL 1, 03–1, 08–1, 23; PW 1, 08–1, 18–1, 30; EL 2, 65–2, 80–3, 00; EW 1, 70–1, 75–1, 85; SBL 4, 33–4, 48–4, 63; PW/EW 0, 67; PW/PL 1, 09; PL/EL 0, 39; EW/EL 0, 63.
(Fig. 59). The known range of this species extends in Middle America from central Costa Rica northward to northwestern Mexico (slightly north of the Tropic of Cancer).
Most label data indicate “attracted to lights”. Tenerals were collected in November.
MEXICO. Sinaloa, Culiacán Apr 1969 (USNM, 9) Veracruz, Cordoba Nov. 1966 (USNM, 7) Oaxaca Tehuantepec Dec. 1964 (USNM, 1) Chiapas Tapilulas May 1974 (USNM, 9) Colima Volcano (USNM, 5) Jalapa (IRSNB, 2) Sa. De Durango (IRSNB, 4) Guerrero Chiapas Jul 2005 (PBPC, 2). BELIZE. Sibun River at Gracy Rock Jun 1974 (USNM, 7). GUATEMALA. Alta Verapaz May 1973 (USNM, 40) Sacapulas 4500ft Dec. 1947 (USNM, 3). HONDURAS. Dept. Conayagua Rancho Chiquito May 1964 (UASM, 8), Belize district Jun 1968 (USNM, 18), 5 mi E. Choluteca Jul 1965 (USNM, 22), 16 mi W. Sabana Grande Jul 1965 (USNM, 7), Pespire Jul 1965 (USNM, 7), San Marcus Colon Jul. 1965 (USNM, 10). EL SALVADOR. Ch. Del Guayabo May 1975 (USNM, 1). NICARAGUA. Rivas Rio Canas Gordas Jun 1964 (UASM, 18). COSTA RICA. Guanacaste Santa Rosa NP Jun 2004 (ADVA, 34), Guanacaste NP Jun 1990 (USNM, 9), Guanacaste Santa Cruz Est. Bosque Diria 150- 250 m Nov 1998 (USNM, 18), Guanacaste Santa Rosa 300 m Mar 1990 (USNM, 5).
http://species-id.net/wiki/Semiardistomis_pallipes
Figs 52, 58Lectotype, here designated, at MHNP, pinned, labeled: handwritten in box “pallipes Dej. Colombie C. Gory”/ handwritten “52”/ “Lectotype Clivina pallipes Dej. Des. S.W. Nichols 1984”/ printed in red paper “lectotype”.
Holotype of Ardistomis striga at IRSNB, glued on cardboard, genitalia attached in microvial, labeled: green paper handwritten “A. striga My. Panama. (Mky)”/ printed “Soc. Ent. Belg. Coll. Putzeys”/ red paper printed “Type”/ handwritten “P. Basilewsky Ardistomus striga Putz”/ Holotype Ardistomis striga Putzeys det. DRWhitehead”.
Given by
Body piceous with green brassy reflections. Elytral surface with isodiametric mesh pattern, striae complete, punctures absent, 3 setiferous punctures on interval 3. Abdominal sternum VII with 4+4 setiferous punctures.
dorsal aspect, as in Fig. 52.
Variation of measurements (mm) and ratios for Semiardistomis pallipes (n=12) are: HL = 0, 44–0, 54–0, 66; PL 0, 86–1, 01–1, 20; PW 0, 92–1, 10–1, 34; EL 2, 30–2, 74–3, 24; EW 1, 48–1, 72–2, 00; SBL 3, 70–4, 30–5, 10; PW/EW 0, 64; PW/PL 1, 09; PL/EL 0, 37; EW/EL 0, 63. More western specimens have complete elytral striae, whereas in those from the Amazon Basin the striae are shorter (Fig. 58).
(Fig. 58). The range of this species extends in South America from northern Brazil and eastern Ecuador north to Colombia, and to Panama in Middle America.
Label data indicate that specimens of Semiardistomis pallipes were collected at margins of water bodies.
BRAZIL. Pernambuco 4 Jan 1883 (USNM, 30). COLOMBIA. Colombie (MHNP, 9); San Alberto 10 Nov 1968 (MHNP, 8). ECUADOR. Napo Onkone Gare Camp 10 Sep. 1995 (FSCA, 3). PANAMA. Panama Mky (IRSNB, 2). PERU. Madre de Dios Rio Manu 18 Jul 1992 (USNM, 24). VENEZUELA. Venz. (IRSNB, 1)
http://species-id.net/wiki/Semiardistomis_propinquus
Figs 54, 59Lectotype, here designated, male at IRSNB, glued on card, genitalia attached in microvial, labeled: green paper handwritten "A. propinqua Mex. (Sallé) Chd."/ handwritten " Lectotype Ardistomis propinqua Putzeys det. DRWhitehead"/ "Ardistomus propinquus Putz P Basilewsky 1955"/ printed "Soc. Ent. Belg. Coll. Putzeys"/ "Syntype".
Paralectotypes, here designated: two specimens at IRSNB labeled as the lectotype. Another 3 paralectotypes at MHNP labeled: handwritten in box "propinqua Chaud. Mexique Las Peras. Sallé".
Type area. Given by
Diagnosis. Body piceous with green reflections. Elytral surface completely smooth, striae continuously impressed in basal 1/8, punctures deeply impressed in basal 2/3, most punctures with setae, 3 setiferous punctures on interval 3. Abdominal sternum VII with 4+4 setiferous punctures.
Habitus. dorsal aspect, as in Fig. 54.
Variation of measurements (mm) and ratios for Semiardistomis propinquus from Mexico (n=6) are: HL = 0, 60–0, 62–0, 63; PL 1, 03–1, 14–1, 25; PW 1, 03–1, 23–1, 30; EL 3, 31–3, 47–3, 63; EW 1, 88–2, 01–2, 13; SBL 5, 04–5, 23–5, 44; PW/EW 0, 61; PW/PL 1, 08; PL/EL 0, 33; EW/EL 0, 58.
Despite the presence of setiferous punctures (other than the usual three in interval 3) and the proportionally larger elytra, this species is markedly similar to Semiardistomis labialis and is possibly simply a hirsute form of that species. Similar variation has been documented for other Carabidae, including Agonum decorum (Say) (
(Fig. 59). records indicate that this species is restricted to mountains in southern Mexico.
Label data indicate that exemplars of this species have been collected during the night at margins of a small pond.
This species probably constitutes a morph of Semiardistomis labialis (Chaudoir). If this is shown to be so, the name Semiardistomis propinquus will become a junior synonym of the name Semiardistomis labialis.
In addition to types: MEXICO. Guanajuato (IRSNB, 3) Puebla 20 May 1973 (USNM, 42) Oaxaca Jul. 1964 (USNM, 14) Jalisco S. Guadalajara Jul 1964 (USNM, 4) Michoacan (USNM, 1) Chiapas May 1974 (FSCA, 12) Guerrero Picaya 12 Jun 2004 (CAS, 8) Morelos, Tepoztlan 27 Oct 2004 (CAS, 7).
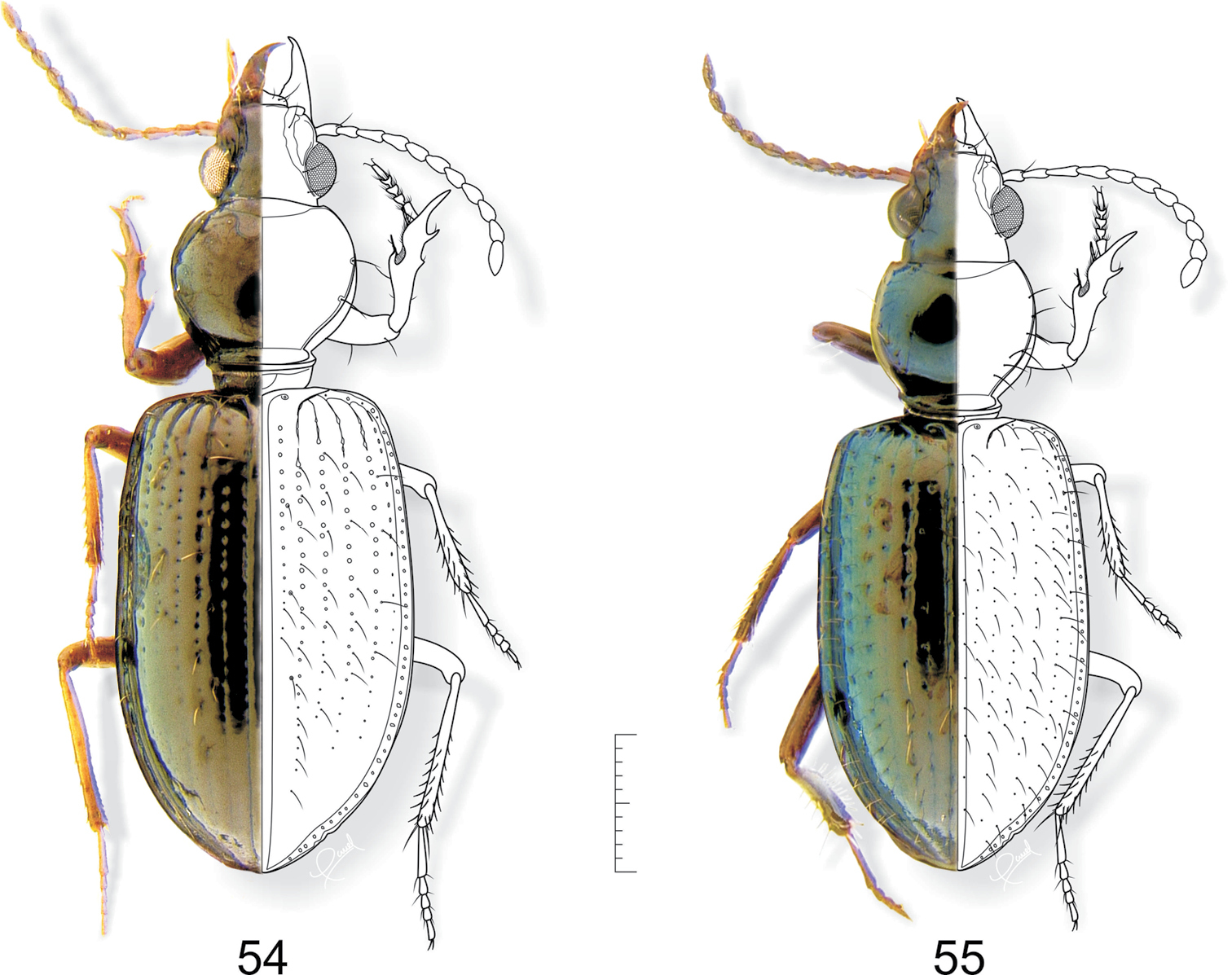
54 Semiardistomis propinquus (Putzeys) 55 Semiardistomis viridis (Say). Dorsal aspect. Scale bar 1 mm.
http://species-id.net/wiki/Semiardistomis_semipunctatus
Fig. 58Lectotype, here designated, at MHNP, pinned, labeled: green paper handwritten “Buenos Ayres Lacordaire”/ handwritten “ 24”/ “Lectotype Clivina pallipes Dej. Des. S.W. Nichols 1984”/ printed on red paper “Lectotype”.
Given by
Body piceous with green bluish reflections. Elytral surface completely smooth, striae continuously impressed in basal 1/8, without or with few punctures, 3 setiferous pores on interval 3. Abdominal sternum VII with 4+4 setiferous punctures.
Variation of measurements (mm) and ratios for Semiardistomis semipunctatus (n=18) are: HL = 0, 46–0, 58–0, 65; PL 0, 93–1, 07–1, 16; PW 0, 95–1, 14–1, 28; EL 2, 25–2, 78–3, 06; EW 1, 38–1, 69–1, 88; SBL 3, 70–4, 43–4, 86; PW/EW 0, 68; PW/PL 1, 07; PL/EL 0, 39; EW/EL 0, 61.
Morphological variation is observed in the elytral punctures. In three different samples from Sta Catarina, Nova Teutonia, Brazil, exemplars with this form are mixed with exemplars of Semiardistomis deletus together with intermediate forms.
(Fig. 58). The known range of this species is confined to a South American area south of the Tropic of Capricorn, extending from southeastern Brazil westward to western Argentina, and south to southern Uruguay.
ARGENTINA. Buenos Aires Delta Parana Jan 1943 (IRSNB, 2). BRAZIL. Campos (HECO, 2); Nova Teutonia (IRSNB, 16); Nova Teutona Sta Catarina Nov 1955 (MHNP, 8); Sta Catarina Nov 1970 (USNM, 21). URUGUAY. Montevideo (IRSNB, 4).
http://species-id.net/wiki/Semiardistomis_viridis
Figs 55 - 57Neotype, designated by
Philadelphia Neck, Pennsylvania, U.S.A.
Body piceous with green reflections. Two pairs of supraorbital setae. More than two premedial setiferous punctures on pronotal disc. Elytral surface completely smooth, striae absent, punctures deeply impressed, setiferous. Abdominal sternum VII with 6+6 setiferous punctures. Body length given by
dorsal aspect, as in Fig. 55.
(Fig. 59). Known from southeastern United States and Bahama Islands in the West Indies (
I collected this species in the “Eagle trail” at the Grassy Waters Preserve in West Palm Beach on April 2, 2011. Adults were active during the day on sandy margins of a fresh water pond (Fig. 56).
USA. Marion Co. V 99 Fla (USNM, 2) Jackson Co, Fla XII 97 (FSCA, 4). Sarasota Co, Fla. May 68 (USNM, 2) San Jacinto Co. Tex, Jun 68 (USNM, 4). West Palm Beach, Fla, March 2011(PVCCu, 12) Etats unis (MHNP, 6).

Habitat of Semiardistomis viridis (Say) at Grassy Waters Preserve, West Palm Beach, Florida, USA
The distribution of the species of the puncticollis group is inadequately known because most of the species have been rarely collected (except for Semiardistomis puncticollis and Semiardistomis laevistriatus). Nonetheless, some patterns can be inferred. The plesiotypic state for some characters seems to be concentrated in the northern portion of South America, with putatively apotypic character states distributed peripherally in a radial fashion. This pattern is similar for the labialis species–group. Since both groups show a similar pattern, I postulate a possible center of origin for the genus and then radiation of descendant lineages from, Northern South America.
The northern radiant of the puncticollis species-group reached temperate southern North America through what is now Middle America (Central America + Mexico). That lineage became extinct, except for its northern descendant, Semiardistomis puncticollis. An eastern lineage colonized the West Indian Lesser Antilles, evidence being the extant Semiardistomis laevistriatus, which became a forest-inhabiting brachypterous humicole, confined now to the Island of Guadeloupe (Basse Terre). Incidentally, Guadeloupe houses other clivinines: Ardistomis atripennis Putzeys (macropterous) and Ardistomis guadeloupensis Kult (brachypterous); and two species of the genus Oxydrepanus.
Southward, the distributional record is too incomplete to infer patterns. Notably are localities with many sympatric species, evidencing high power of dispersal in this southern assemblage, being Loreto and Madre de Dios in Peru the localities with highest number of sympatric species.
The labialis species–group seems to follow a similar zoogeographic structure. Northward from the putative center of radiation (Fig. 59), Semiardistomis viridis is the most derivative form. It is broadly sympatric with Semiardistomis puncticollis, and like that species may be the oldest survivor of an interruption of the ranges of the Central American lineages. But here, contrary to puncticollis species–group, a second event arose in Semiardistomis labialis and related forms in Central America, from which source originated the stock that gave rise to the West Indian Greater Antillean Semiardistomis cyaneolimbatus. That stock may have reached the islands by overseas dispersal, or by means of a now foundered land bridge. More recently, the West Indian Bahamas was invaded by Semiardistomis viridis, no doubt by overseas dispersal.
Southward (Fig. 58) in South America, , the structurally derivative forms of the labialis species– group do not show defined patterns of speciation possibly for several reasons, including the following. First, as far as I can determine, interspecific differentiation is ruled just by some variations in the elytra surface and those variations have shown to be unstable inside localities samples checked. Second, contrary to puncticollis species–group, sympatric “species” share every grade of intermediate forms evidencing a continuous flux between populations. Probably gene flow has not been interrupted southward and we just see the result of the polymorphic condition of only one species derivate from a northern ancestor.
In both species–groups some morphological characteristics, like reduction of the prolongation of the median carina of the mentum and hirsutism, have arisen northward from center of radiation following analog ways, suggesting two different paths of evolution to those states under similar evolutionary pressures.
Checklist of the species names of the genus Semiardistomis KultGenus Semiardistomis Kult, 1950
Species–group puncticollis
Semiardistomis cordicollis (Putzeys, 1846)
Semiardistomis darlingtoni (Kult, 1950)
Semiardistomis exspectatus sp. n.
Semiardistomis glabratus (Putzeys, 1866)
Semiardistomis balthasari (Kult, 1950)
Semiardistomis jedlickai (Kult, 1950)
Semiardistomis laevistriatus (Fleutiaux & Sallé, 1889)
Semiardistomis maindroni (Kult, 1950)
Semiardistomis major sp. n.
Semiardistomis pilosellus (Kult, 1950)
Semiardistomis puncticollis (Dejean, 1831)
Semiardistomis rugosus (Putzeys, 1866)
Semiardistomis subglabra (van Emden, 1949)
Semiardistomis vlastae (Kult, 1950)
Species–group labialis
Semiardistomis cyaneolimbatus (Chevrolat, 1863)
Semiardistomis gundlachi (Putzeys, 1866)
Semiardistomis deletus (Putzeys, 1846)
Semiardistomis emdeni (Kult, 1950)
Semiardistomis flavipes (Dejean, 1831)
Semiardistomis aenea (Putzeys, 1866)
Semiardistomis brittoni (Kult, 1950)
Semiardistomis marani (Kult, 1950)
Semiardistomis labialis (Chaudoir, 1837)
var. picipes (Bates, 1881)
var. nanus (Bates, 1881)
var. dilatatus (Bates, 1881)
Semiardistomis tuspanensis (Putzeys, 1846)
Semiardistomis pallipes (Dejean, 1831)
var. caerulea (Putzeys, 1846)
Semiardistomis striga (Putzeys, 1866)
Semiardistomis propinquus (Putzeys, 1866)
Semiardistomis semipunctatus (Dejean, 1831)
Semiardistomis viridis (Say, 1823)
Semiardistomis vicinus (Putzeys, 1846)
Semiardistomis rostrata (Dejean, 1825)
Concluding remarksThis revision has added an important part in understanding of the structure of subtribe Ardistomina. At this point the road is clear to complete revisions for the rest of Ardistomine genera, which are under construction and will help with new elements to a better comprehension of inter generic relationships. Some questions have to be solved inside the genus originated in part by poor representation in collections of material from many areas mainly from South America. For sure, new samples and DNA studies will contribute to solve unclear specific status. In species–group labialis we have most of problems due to polymorphic condition of most species. In Central America Semiardistomis propinquus must be evaluated using consistent genetic data to determinate whether or not it is an isolated species from Semiardistomis labialis. In South America we just will be able to corroborate denominations of species Semiardistomis pallipes, Semiardistomis flavipes, Semiardistomis deletus and Semiardistomis semipunctatus when gaps in distributional records will filled with new material mainly from Amazonian and Central Brazil and meridional half of Argentina; only them, we will have data enough to picture how different characters with taxonomic value behave in relation with geographic position.
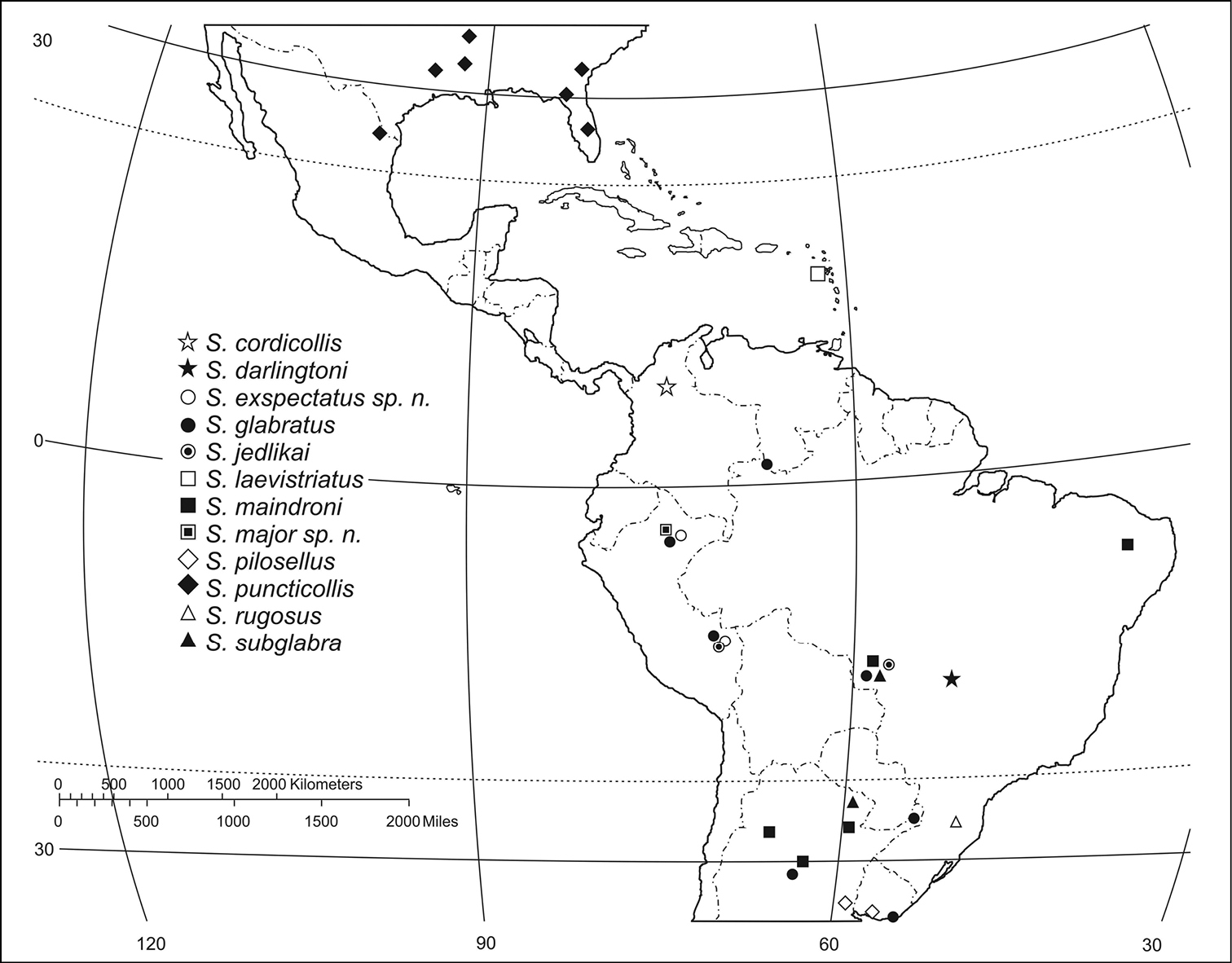
Distribution map of members of the puncticollis species–group.
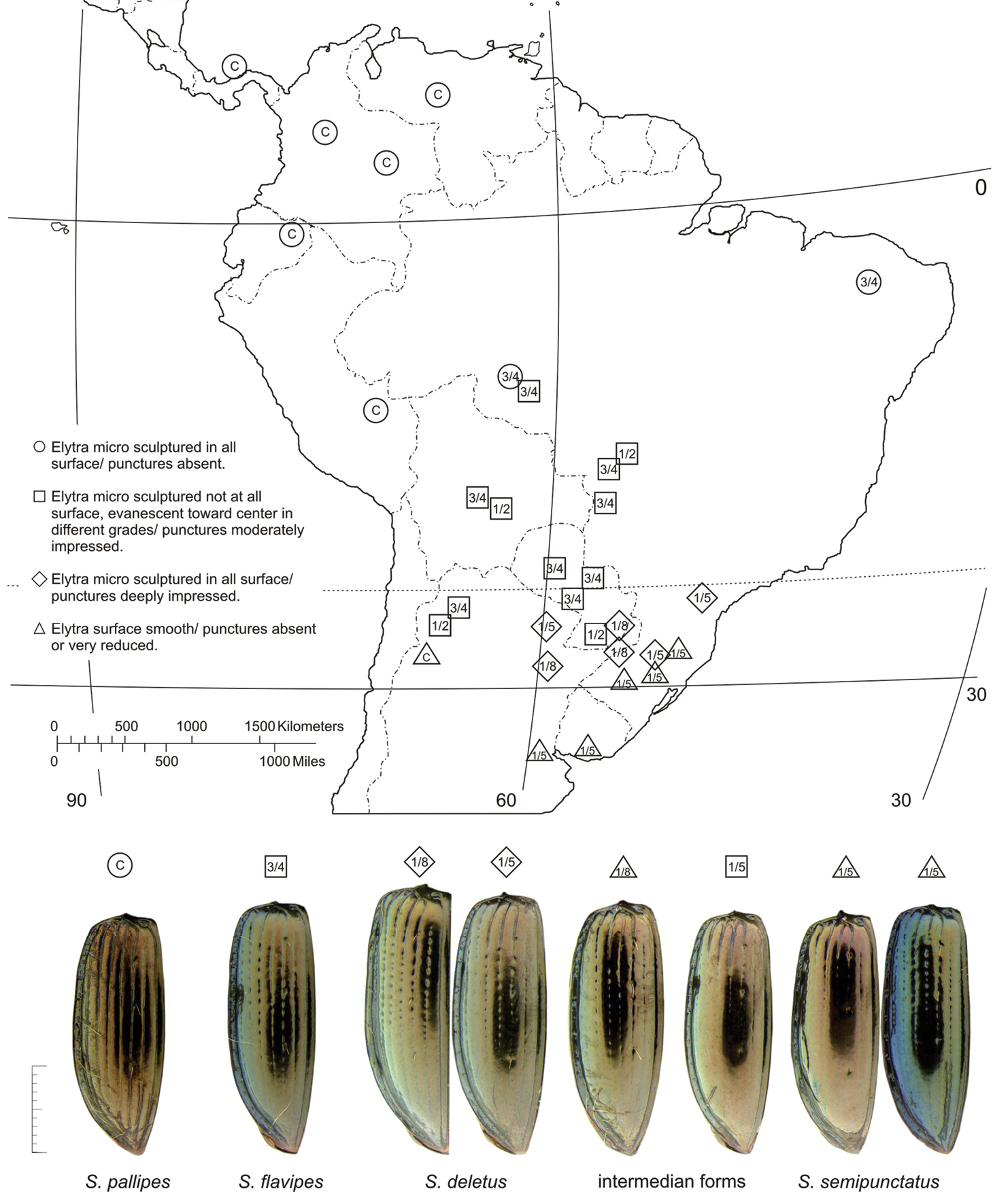
Distribution map, illustrations of exemplar left elytra, and variability of members of the labialis species–group in South America. Value inside symbol indicates portion of basal elytra in which striae are impressed, (C, complete impressed). Scale bar 1 mm.
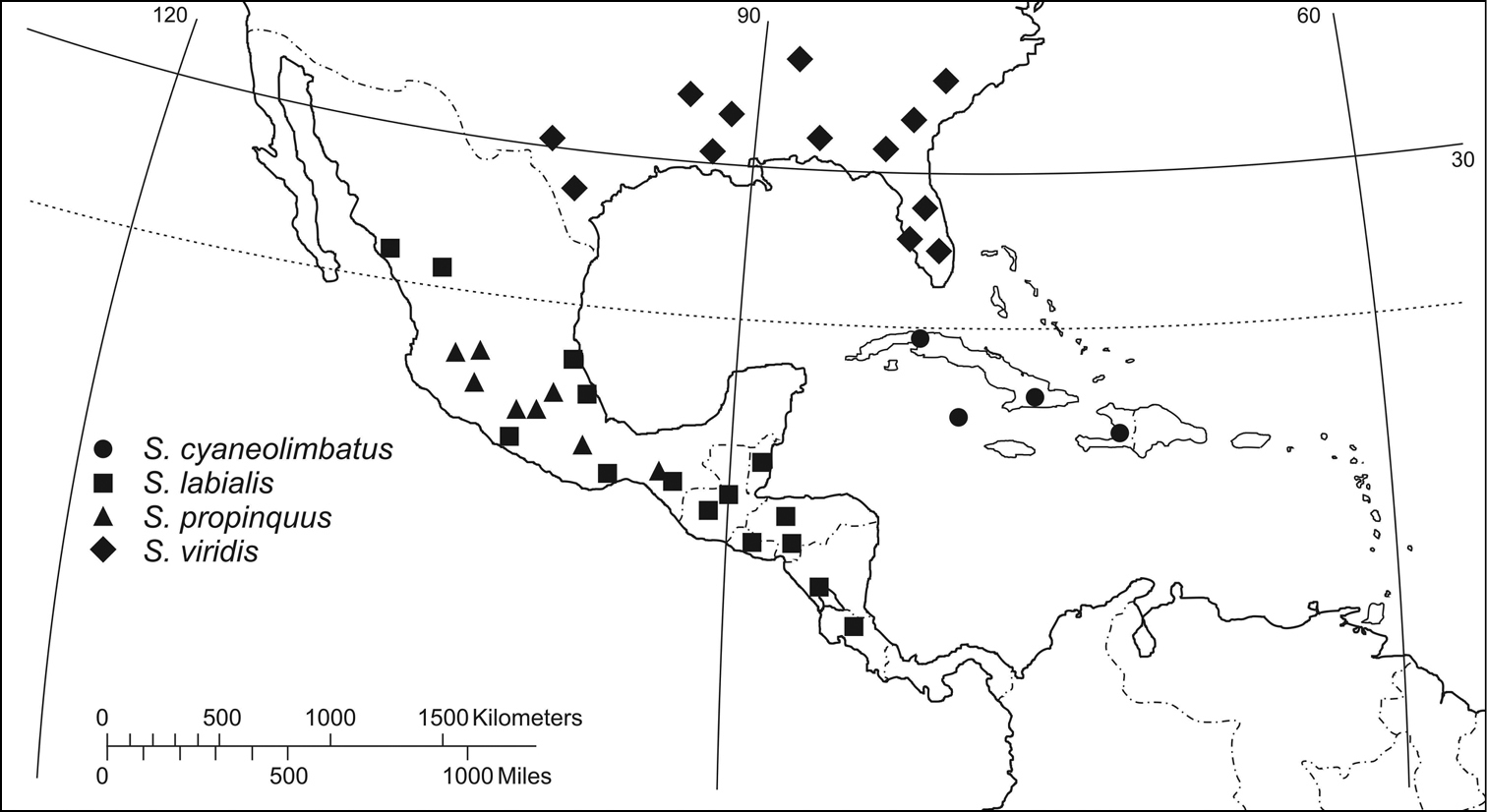
Distribution map of members of the labialis species–group in Central America.
I thank Dr. Terry L. Erwin for his assistance during my visit to the Smithsonian Institution, for his suggestions that clarifies my taxonomic interpretations, for reviewing of the manuscript and for general encouragement all this time. I appreciate revisions of the manuscript made by Dr. Yves Bousquet and Dr. George E. Ball. When in Washington DC I thank assistance provide by Charyn Micheli at the Department of Entomology of the Smithsonian Institution as well as the kind hospitality provide by Héctor Malagón and his family. I also thank Dr. Alain Drumont, Dr. Yves Samyn and Dr. Marie–Lucie Susini for their assistance during my visits to Brussels. I acknowledge loans from curators of public or private collections mentioned above. Field work was possible due to assistance of Dr. Luis M. Diaz in Isla de Pinos, Cuba and Corbary Interian in West Palm Beach, Florida. Financial support for revision of material in USA came from grant offered by the Office of Fellowships of the Smithsonian Institution in 2011 and for revision of material in Europe came from the Belgian Development Cooperation and the Royal Belgian Institute of Natural Sciences through the Belgian Focal Point of the Global Taxonomy Initiative held during 2007, 2008 and 2010. Open access to this paper was supported by the Encyclopedia of Life (EOL) Open Access Support Project (EOASP).