Research Article |
|
Corresponding author: Xiaoping Wu ( xpwu@ncu.edu.cn ) Academic editor: Maria Elina Bichuette
© 2019 Jiajun Qin, Xiongjun Liu, Yang Xu, Xiaoping Wu, Shan Ouyang.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Qin J, Liu X, Xu Y, Wu X, Ouyang S (2019) Beta diversity patterns of fish and conservation implications in the Luoxiao Mountains, China. ZooKeys 817: 73-93. https://doi.org/10.3897/zookeys.817.29337
|
Abstract
The Luoxiao Mountains play an important role in maintaining and supplementing the fish diversity of the Yangtze River Basin, which is also a biodiversity hotspot in China. However, fish biodiversity has declined rapidly in this area as the result of human activities and the consequent environmental changes. Beta diversity was a key concept for understanding the ecosystem function and biodiversity conservation. Beta diversity patterns are evaluated and important information provided for protection and management of fish biodiversity in the Luoxiao Mountains. The results showed that the spatial turnover component was the main contributor to beta diversity of Hemiramphidae, Amblycipitidae, Catostomidae, Clariidae, Balitoridae and Percichthyidae in the Luoxiao Mountains, which indicated that a number of protected areas would be necessary to conserve fish biodiversity and that these families would need conservation measures. Most protected areas are currently limited to some regions; therefore, in order to protect fish diversity, conservation efforts must target an increase in the number of protected areas which should be spread across each of the regions.
Keywords
beta diversity, commercial fishes, Luoxiao Mountains, protected areas
Introduction
Biodiversity patterns and their formation mechanisms have been one of the hot issues, and it is also an important foundation for conservation (
Beta diversity is an important tool for conservation planning (
The Luoxiao Mountains range is located in the southeast of China’s mainland and has a long history and complex environmental factors (
Material and methods
Study area
The Luoxiao Mountains (25°32'–29°28'N, 113°09'–114°26'E) are a large system of mountain ranges, located in the southeast of China’s mainland with an overall north-south trend, stretching across Hubei, Hunan, and Jiangxi provinces. It consists of Mufu Mountain, Jiuling Mountain, Wugong Mountain, Zhuguang Mountain, and others. The total length of the Luoxiao Mountains is 400 km and altitude ranges are 82–2120 m. Lingfeng Peak (2122 m) is one of the highest mountains in the southeastern Eurasia. Its average precipitation range is 1341–1943 mm and forest coverage in the watershed reaches 90% (Table
Hydrology and environmental characteristics of the streams of Luoxiao Mountains. JJ: Jinjiang River; YS: Yuanshui River; HS: Heshui River; SS: Shushui River; SC: Suichuan River; SY: Shangyou River; MS: Mishui River; ML: Miluo River; FS: Fushui River; XH: Xiuhe River; LY: Liuyang River.
| Stream | Latitude | Longitude | Length (km) | Area (km2) | Average gradient (%) | Average precipitation (mm) | Average temperature (°C) | Annual average runoff (×108 m3) | Average Altitude (m) |
| LY | 28°24'–28°46' | 112°99'–114°04' | 222 | 4665 | 0.57 | 1598 | 17.3 | 39.41 | 252 |
| MS | 27°16'–26°25' | 112°88'–113°99' | 296 | 10305 | 1.01 | 1483 | 18.1 | 76.03 | 352 |
| ML | 28°86'–29°02' | 112°93'–114°05' | 253 | 5543 | 0.46 | 1400 | 17.6 | 43.04 | 250 |
| FS | 29°49'–29°86' | 114°40'–115°45' | 196 | 5250 | 0.79 | 1275 | 16.6 | 43.5 | 613 |
| XH | 28°31'–29°12' | 114°14'–116°01' | 419 | 14700 | 0.48 | 1663 | 16.7 | 135.1 | 676 |
| SY | 25°37’-25°49’ | 113°43'–114°49' | 204 | 4647 | 0.70 | 1570 | 18.8 | 33 | 615 |
| SC | 26°11'–26°30' | 113°56'–114°44' | 176 | 2882 | 2.36 | 1640 | 16.9 | 27.1 | 971 |
| SS | 126°29'–26°47' | 14°04'–114°50' | 152 | 1301 | 2.14 | 1630 | 16.7 | 11.3 | 1610 |
| HS | 27°04'–27°24' | 114°01'–114°59' | 256 | 9103 | 0.59 | 1580 | 17.8 | 27.4 | 747 |
| YS | 27°27'–28°04' | 114°10'–115°29' | 279 | 6262 | 0.34 | 1678 | 17.2 | 29.6 | 391 |
| JJ | 27°57'–28°25' | 114°01'–115°49' | 307 | 7886 | 0.26 | 1679 | 17.6 | 70 | 391 |
Sampling methods
Sampling sites were selected by considering habitats, variations, and anthropogenic activities in the Luoxiao Mountains. Fish samples were collected from April 2014 to 2017 in eleven streams of the Luoxiao Mountains. We selected eleven streams (42 sampling sites) (Figure

Data analysis
Beta diversity is represented by the difference in species composition between different communities, which was determined by species turnover (species replacement) and nestedness (richness difference;
BAS frameworks (Sørensen index):
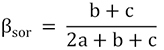
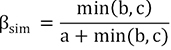
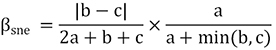
POD frameworks (Jaccard index):
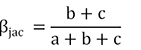
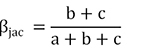
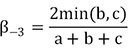
Results
Fish species composition
The fish specimens sampled and identified in the Luoxiao Mountains were categorized into 113 species and 17 families (Figure

Beta diversity patterns
The fish composition similarity in the Luoxiao Mountains had a mean value of 0.50 and 0.67, based on BAS and POD frameworks respectively (SD ± 0.06 and SD ± 0.05, respectively; Table
Fish compositional similarity by BAS and POD frameworks in the streams of Luoxiao Mountains. JJ: Jinjiang River; YS: Yuanshui River; HS: Heshui River; SS: Shushui River; SC: Suichuan River; SY: Shangyou River; MS: Mishui River; ML: Miluo River; FS: Fushui River; XH: Xiuhe River; LY: Liuyang River; ES: Eastern stream of Luoxiao Mountain; WS: Western stream of Luoxiao Mountains.
| Stream | β | |||||
| BAS | POD | |||||
| βsor | βsim | βsne | βjac | β-3 | βrich | |
| JJ | 0.52±0.08 | 0.32±0.11 | 0.19±0.13 | 0.68±0.07 | 0.33±0.16 | 0.35±0.19 |
| YS | 0.48±0.08 | 0.27±0.06 | 0.21±0.11 | 0.65±0.07 | 0.27±0.11 | 0.37±0.17 |
| HS | 0.52±0.08 | 0.39±0.09 | 0.13±0.09 | 0.68±0.07 | 0.42±0.14 | 0.26±0.16 |
| SS | 0.49±0.07 | 0.38±0.07 | 0.11±0.07 | 0.66±0.06 | 0.42±0.12 | 0.24±0.14 |
| SC | 0.54±0.11 | 0.31±0.09 | 0.22±0.10 | 0.69±0.09 | 0.28±0.11 | 0.41±0.17 |
| SY | 0.48±0.03 | 0.37±0.10 | 0.11±0.09 | 0.65±0.03 | 0.42±0.15 | 0.22±0.14 |
| MS | 0.47±0.05 | 0.36±0.06 | 0.11±0.08 | 0.64±0.05 | 0.41±0.12 | 0.22±0.14 |
| ML | 0.50±0.04 | 0.38±0.10 | 0.12±0.09 | 0.66±0.04 | 0.42±0.14 | 0.24±0.15 |
| FS | 0.55±0.07 | 0.39±0.06 | 0.16±0.10 | 0.71±0.05 | 0.39±0.15 | 0.32±0.19 |
| XH | 0.48±0.04 | 0.36±0.08 | 0.11±0.08 | 0.65±0.03 | 0.42±0.14 | 0.23±0.15 |
| LY | 0.49±0.03 | 0.39±0.07 | 0.10±0.07 | 0.66±0.03 | 0.45±0.12 | 0.21±0.12 |
| ES | 0.50±0.07 | 0.34±0.09 | 0.16±0.11 | 0.66±0.06 | 0.36±0.14 | 0.30±0.17 |
| WS | 0.50±0.06 | 0.38±0.07 | 0.12±0.09 | 0.67±0.05 | 0.42±0.13 | 0.25±0.15 |
| Total | 0.50±0.06 | 0.36±0.08 | 0.14±0.09 | 0.67±0.05 | 0.39±0.13 | 0.28±0.16 |
At the same time, fish composition similarity (βsør and βjac) for the entire fish fauna had a mean value of 0.66 and 0.76 (SD ± 0.24 and 0.21, Table
BAS and POD frameworks based on all species and 17 families in the streams of Luoxiao Mountain. Values are mean ± standard deviation.
| Family | BAS | POD | ||||
| βsor | βsim | βsne | βjac | β-3 | βrich | |
| Catostomidae | 0.70±0.32 | 0.39±0.05 | 0.31±0.03 | 0.77±0.30 | 0.25±0.03 | 0.51±0.03 |
| Cyprinidae | 0.65±0.24 | 0.41±0.03 | 0.25±0.02 | 0.76±0.20 | 0.32±0.02 | 0.43±0.02 |
| Cobitidae | 0.66±0.26 | 0.37±0.04 | 0.28±0.03 | 0.75±0.23 | 0.28±0.03 | 0.47±0.03 |
| Balitoridae | 0.69±0.25 | 0.49±0.04 | 0.20±0.02 | 0.78±0.21 | 0.39±0.03 | 0.39±0.02 |
| Siluridae | 0.64±0.30 | 0.26±0.03 | 0.38±0.03 | 0.73±0.29 | 0.17±0.02 | 0.56±0.03 |
| Clariidae | 0.70±0.32 | 0.39±0.05 | 0.31±0.03 | 0.77±0.30 | 0.25±0.03 | 0.51±0.03 |
| Bagridae | 0.67±0.24 | 0.41±0.03 | 0.27±0.02 | 0.77±0.21 | 0.32±0.03 | 0.45±0.03 |
| Amblycipitidae | 0.78±0.23 | 0.58±0.04 | 0.21±0.03 | 0.85±0.19 | 0.40±0.03 | 0.45±0.03 |
| Sisoridae | 0.56±0.22 | 0.34±0.03 | 0.22±0.02 | 0.69±0.18 | 0.31±0.03 | 0.38±0.02 |
| Hemiramphidae | 0.93±0.16 | 0.80±0.04 | 0.14±0.03 | 0.96±0.13 | 0.49±0.03 | 0.47±0.03 |
| Syngnathidae | 0.53±0.25 | 0 | 0.53±0.25 | 0.65±0.26 | 0 | 0.65±0.26 |
| Mastacembelidae | 0.64±0.22 | 0.43±0.03 | 0.21±0.02 | 0.75±0.18 | 0.38±0.03 | 0.37±0.02 |
| Percichthyidae | 0.69±0.24 | 0.49±0.04 | 0.20±0.02 | 0.78±0.20 | 0.40±0.03 | 0.39±0.02 |
| Odontobutidae | 0.65±0.25 | 0.49±0.04 | 0.16±0.02 | 0.76±0.20 | 0.33±0.16 | 0.35±0.19 |
| Gobiidae | 0.66±0.25 | 0.42±0.04 | 0.24±0.02 | 0.76±0.22 | 0.32±0.03 | 0.44±0.02 |
| Belontiidae | 0.53±0.23 | 0.31±0.03 | 0.23±0.02 | 0.67±0.20 | 0.28±0.03 | 0.39±0.02 |
| Channidae | 0.58±0.22 | 0.35±0.03 | 0.23±0.02 | 0.71±0.19 | 0.30±0.02 | 0.41±0.02 |
| All species | 0.66±0.24 | 0.41±0.03 | 0.25±0.02 | 0.76±0.21 | 0.32±0.03 | 0.44±0.02 |
The PCA showed that fish composition similarity of LY, SS, SY, XH, and MS were similar based on BAS and POD frameworks; FS, JJ and SC were similar; HS and ML were similar; and YS was uniquely divided into other areas, respectively (Figure

Results of the principal component analysis (PCA) on the compositional similarity of fish species in the streams of the Luoxiao Mountains. JJ: Jinjiang River; YS: Yuanshui River; HS: Heshui River; SS: Shushui River; SC: Suichuan River; SY: Shangyou River; MS: Mishui River; ML: Miluo River; FS: Fushui River; XH: Xiuhe River; LY: Liuyang River.
We found almost no significant effects of geographical drivers on overall beta diversity for the Luoxiao Mountains (Table
Effects of geographical drivers on pairwise compositional similarity and its partitioned components obtained from BAS and POD frameworks in the streams of Luoxiao Mountain, Jiangxi Province. Significant results (P < 0.05) are in bold.
| Area | Length | Average precipitation | Annual average runoff | Average altitude | Average gradient | Average temperature | |||
| BAS | βsor | r | 0.162 | 0.134 | -0.008 | 0.273 | 0.055 | -0.162 | -0.070 |
| P | 0.190 | 0.203 | 0.454 | 0.069 | 0.333 | 0.850 | 0.647 | ||
| βsim | r | -0.318 | -0.245 | 0.187 | -0.289 | 0.248 | 0.128 | 0.058 | |
| P | 0.931 | 0.871 | 0.127 | 0.888 | 0.032 | 0.272 | 0.415 | ||
| βsne | r | 0.386 | 0.304 | -0.168 | 0.436 | -0.179 | -0.221 | -0.098 | |
| P | 0.022 | 0.062 | 0.805 | 0.011 | 0.861 | 0.943 | 0.658 | ||
| POD | βjac | r | 0.155 | 0.122 | -0.017 | 0.254 | 0.067 | -0.147 | -0.076 |
| P | 0.150 | 0.244 | 0.521 | 0.072 | 0.328 | 0.808 | 0.680 | ||
| β-3 | r | -0.366 | -0.285 | 0.164 | -0.401 | 0.236 | 0.219 | 0.081 | |
| P | 0.961 | 0.913 | 0.160 | 0.970 | 0.028 | 0.051 | 0.370 | ||
| βrich | r | 0.365 | 0.284 | -0.145 | 0.430 | -0.176 | -0.237 | -0.096 | |
| P | 0.019 | 0.076 | 0.758 | 0.015 | 0.851 | 0.981 | 0.630 | ||
Discussion
Fish species composition
Studies on fish composition and diversity in streams is the basis for the conservation and management of stream fishes (
Beta diversity patterns
Abiotic and biotic factors and their ecological processes in different stream sizes varies substantially (
Biogeographical processes
The modern freshwater fish fauna of Eurasia originated in the early Tertiary (
Threats to fish diversity
The headwater stream is a tributary of a larger river, which is often located in a mountainous area with high altitude. Compared with large rivers, it had relatively simple habitat structure, poor nutrition, obvious hydrological change, and low species diversity (
Conservation implications
Freshwater fishes were thought to be the world’s most threatened group of vertebrates after amphibians (
Acknowledgements
This work is supported by grants from the Key Project of Science-Technology Basic Condition Platform from The Ministry of Science and Technology of the People’s Republic of China (Grant No. 2005DKA21402), and the foundation project of the National Ministry of Science and Technology of China (2013FY111500). The authors report no conflict of interest. The authors alone are responsible for the content and writing of this article.
References
- Allan JD, Castillo MM (2007) Stream ecology: Structure and function of running waters (2nd edition). Springer, Netherlands. https://doi.org/10.1007/978-1-4020-5583-6
- Anderson MJ, Ellingsen KE, McArdle BH (2006) Multivariate dispersion as a measure of beta diversity. Ecology Letters 9: 683–693. https://doi.org/10.1111/j.1461-0248.2006.00926.x
- Arthington AH, Dulvy NK, Gladstone W, Winfield IJ (2016) Fish conservation in freshwater and marine realms: status, threats and management. Aquatic Conservation Marine & Freshwater Ecosystems 26: 838–857. https://doi.org/10.1002/aqc.2712
- Baselga A (2010) Partitioning the turnover and nestedness components of beta diversity. Global Ecology and Biogeography 19: 134–143. https://doi.org/10.1111/j.1466-8238.2009.00490.x
- Baselga A (2012) The relationship between species replacement, dissimilarity derived from nestedness, and nestedness. Global Ecology and Biogeography 21: 1223–1232. https://doi.org/10.1111/j.1466-8238.2011.00756.x
- Baselga A, Orme CDL (2012) betapart: an R package for the study of beta diversity. Methods in Ecology & Evolution 3: 808–812. https://doi.org/10.1111/j.2041-210X.2012.00224.x
- Bruton MN (1995) Have fishes had their chips? The dilemma of threatened fishes. Environmental Biology of Fish 43: 1–27. https://doi.org/10.1007/BF00001812
- Bush A, Harwood T, Hoskins AJ, Mokany K, Ferrier S, et al. (2016) Current uses of beta-diversity in biodiversity conservation: a response to Socolar. Trends in Ecology and Evolution 31: 337–338. https://doi.org/10.1016/j.tree.2016.02.020
- Cardoso P, Rigal F, Carvalho JC (2015) BAT-Biodiversity Assessment Tools, an R package for the measurement and estimation of alpha and beta taxon, phylogenetic and functional diversity. Methods in Ecology and Evolution 6: 232–236. https://doi.org/10.1111/2041-210X.12310
- Carvalho JC, Cardoso P, Gomes P (2012) Determining the relative roles of species replacement and species richness differences in generating beta-diversity patterns. Global Ecology and Biogeography 21: 760–771. https://doi.org/10.1111/j.1466-8238.2011.00694.x
- Chen YY (1998) Fauna Sinica: OsteichthyesCypriniformes II. Science Press, Beijing, China.
- Chen YY, Cao WX, Zheng CY (1986) Ichthyofauna of the Zhujiang River with a discussion on zoogeographical divisions for freshwater fishes. Acta Hydrobiologica Sinica 10: 228–236.
- Chu XL, Zheng BS, Dai DY (1999) Fauna sinica, Osteichthyes, Siliuriformes. Science Press, Beijing, China.
- Cressey D (2009) Aquaculture: future fish. Nature 458: 398–400. https://doi.org/10.1038/458398a
- De Silva SS (2012) Aquaculture: a newly emergent food production sector and perspectives of its impacts on biodiversity and conservation. Biodiversity and Conservation 21: 3187–3220. https://doi.org/10.1007/s10531-012-0360-9
- Dray S, Dufour A (2007) The ade4 package: implementing the duality diagram for ecologists. Journal of Statistical Software 22: 1–20. https://doi.org/10.18637/jss.v022.i04
- Dulvy NK, Metcalfe JD, Glanville J, Pawson MG, Reynolds JD (2000) Fishery stability, local extinctions and shifts in community structure in skates. Conservation Biology 14: 283–293. https://doi.org/10.1046/j.1523-1739.2000.98540.x
- Dudgeon D, Arthington AH, Gessner MO, Kawabata Z, Knowler D, Leveque C, Naiman RJ, Prieur-Richard AH, Soto D, Stiassny MLJ (2006) Freshwater biodiversity: importance, threats, status, and conservation challenges. Biological Reviews 81: 163–182. https://doi.org/10.1017/S1464793105006950
- Fu C, Wu J, Chen J, Wu Q, Lei G (2003) Freshwater fish biodiversity in the Yangtze River basin of china: patterns, threats and conservation. Biodiversity & Conservation 12: 1649–1685. https://doi.org/10.1023/A:1023697714517
- Gong HL, Zhuang WY, Liao WB (2016) Comprehensive Scientific Survey of Biodiversity in Luoxiao Mountain. China Science and Technology Achievements 17(22): 9–10.
- Grossman GD, Dowd JF, Crawford M (1990) Assemblage stability in stream fishes: A review. Environmental management 14(5): 661–671. https://doi.org/10.1007/BF02394716
- He MC, Wang ZJ, Tang HX (1998) The chemical, toxicological and ecological studies in assessing the heavy metal pollution in Le An River, China. Water Research 2(2): 510–518. https://doi.org/10.1016/S0043-1354(97)00229-7
- Hiddink JG, Mackenzie BR, Rijnsdorp A et al. (2008) Importance of fish biodiversity for the management of fisheries and ecosystems. Fisheries Research 90(1): 6–8. https://doi.org/10.1016/j.fishres.2007.11.025
- Hilborn R, Quinn TP, Schindler DE, Rogers DE (2003) Biocomplexity and fisheries sustainability. Ecological Monographs 75: 3–36.
- Howard PC, Viskanic P, Davenport TRB, Kigenyi FW, Baltzer M, Dickinson CJ, Lwanga JS, Matthews RA, Balmford A (1998) Complementarity and the use of indicator groups for reserve selection in Uganda. Nature 394: 472–475. https://doi.org/10.1038/28843
- Hu ML, Wu ZQ, Liu YL (2009) The fish fauna of mountain streams in the Guanshan National Nature Reserve, Jiangxi, China. Environmental Biology of Fishes 86: 23–27. https://doi.org/10.1007/s10641-009-9496-1
- Huang LL, Wu ZQ, Hu ML, Li Q, Zong DS, Wan ZQ, Zhao WQ (2008) Fish diversity in Lushan Nature Reserve, Jiangxi, China. Journal of Nanchang University 32: 161–164.
- Huang LL, Wu ZQ, Li JH (2013) Fish fauna, biogeography and conservation of freshwater fish in Poyang Lake basin, china. Environmental Biology of Fishes 96: 1229–1243. https://doi.org/10.1007/s10641-011-9806-2
- Huang XP, Gong Y (2007) Fishery resources in Poyang Lake and its conservation. Jiangxi Fishery Science Technology 112(4): 2–6.
- Hughes JM, Schmidt DJ, Finn DS (2009) Genes in streams: using DNA to understand the movement of freshwater fauna and their riverine habitat. Biology Science 59: 573–583. https://doi.org/10.1525/bio.2009.59.7.8
- IUCN (2017) The IUCN Red List of Threatened Species. Version 2016-3. https://www.iucnredlist.org [Accessed on 22 January 2017]
- Jiang ZG, Jiang JP, Wang YZ, Zhang E, Zhang YY et al. (2016) Red List of China’s Vertebrates. Biodiversity Science 24: 500–551. https://doi.org/10.17520/biods.2016076
- Johnson RK, Angeler DG (2014) Effects of agricultural land use on stream assemblages: Taxon-specific responses of alpha and beta diversity. Ecological Indicators 45: 386–393. https://doi.org/10.1016/j.ecolind.2014.04.028
- Kennedy D, Norman C (2005) What don’t we know? Science 309: 75–75. https://doi.org/10.1126/science.309.5731.75
- Kessler M, Abrahamczyk S, Bos M, Buchori D, Putra DD, Gradstein SR et al. (2009) Alpha and beta diversity of plants and animals along a tropical land-use gradient. Ecological Applications 19: 2142–2156. https://doi.org/10.1890/08-1074.1
- Legendre P, Legendre L (2012) Numerical ecology, 3rd edition. Elsevier, Amsterdam.
- Liao WB, Wang YY, Li Z, Peng SL, Chen CQ, Fan Q, Jia FL, Wang L, Liu WQ, Yi GS, Shi XG, Zhang DD (2014) Comprehensive Scientific Survey of Biodiversity in Jinggangshan Area, China. Science Press, Beijing.
- Liu BP, Quan QQ (1996) Course of Historical Geology, 3rd edition edition. Geological Publishing House, Beijing.
- Liu XJ, Hu XY, Ao XF, Wu XP, Ouyang S (2017) Community characteristics of aquatic organisms and management implications after construction of Shihutang Dam in the Gangjiang River, China. Lake and Reservoir Management, 1–16.
- Lytle DA, Poff NL (2004) Adaplation to natural flow regimes. Trends in Ecology and Evolution 19: 94–100. https://doi.org/10.1016/j.tree.2003.10.002
- Margules CR, Pressey RL (2000) Systematic conservation planning. Nature 405: 243–253. https://doi.org/10.1038/35012251
- Moyle PB, Leidy RA (1992) Loss of biodiversity in ecosystems: evidence from fish faunas. In: Fiedler PL, Jain SK (Eds) Conservation biology: the theory and practice of nature conservation, preservation and management.Chapman and Hall, New York, 127–169. https://doi.org/10.1007/978-1-4684-6426-9_6
- Naylor RL, Goldburg RJ, Primavera JH, Kautsky N, Beveridge MCM, Clay J et al. (2000) Effect of aquaculture on world fish supplies. Nature 405: 1017–1024. https://doi.org/10.1038/35016500
- Nogueira C, Buckup PA, Menezes NA, Oyakawa OT, Kasecker TP, Ramos Neto MB et al. (2010) Restricted-range fishes and the conservation of Brazilian freshwaters. Plos One 5: e11390. https://doi.org/10.1371/journal.pone.0011390
- Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin PR, O’Hara R, Simpson GL, Solymos P, Stevens MHH, Wagner H (2015) vegan: community ecology package. R package version 2.3-2. Available at: http://cran. r-project.org
- Osorio D, Terborgh J, Alvarez A, Ortega H, Quispe R, Chipollini V, Davenport LC (2011) Lateral migration of fish between an oxbow lake and an Amazonian headwater river. Ecology of Freshwater Fish 20: 619–627. https://doi.org/10.1111/j.1600-0633.2011.00511.x
- Podani J, Schmera D (2011) A new conceptual and methodological framework for exploring and explaining pattern in presence-absence data. Oikos 120: 1625–1638. https://doi.org/10.1111/j.1600-0706.2011.19451.x
- R Development Core Team (2014) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available at: http://Rproject. org [accessed 23 December 2016]
- Ren P, He H, Song YQ, Cheng F, Xie SG (2016) The spatial pattern of larval fish assemblages in the lower reach of the Yangtze River: potential influences of river-lake connectivity and tidal intrusion. Hydrobiologia 766: 365–379. https://doi.org/10.1007/s10750-015-2471-2
- Roberts JH, Hitt NP (2010) Longitudinal structure in temperate stream fish communities: Evaluating conceptual models with temporal data. American Fisheries Society Symposium 73: 281–299.
- Socolar JB, Gilroy JJ, Kunin WE, Edwards DP (2016) How should beta-diversity inform biodiversity conservation? Trends in Ecology & Evolution 31: 67–80. https://doi.org/10.1016/j.tree.2015.11.005
- Song X, Tang W, Zhang Y (2017) Freshwater fish fauna and zoogeographical divisions in the wuyi-xianxialing mountains of eastern china. Biodiversity Science 25(12): 1331–1338. https://doi.org/10.17520/biods.2017207
- Sutherland WJ, Adams WM, Aronson RB et al. (2009) One hundred questions of importance to the conservation of global biological diversity. Conservation Biology 23: 557–567. https://doi.org/10.1111/j.1523-1739.2009.01212.x
- Tang WQ, Chen YY, Wu HL (2001) Fish species diversity of Wuling Mountains region and its zoogeographic analyses. Journal of Shanghai Ocean University 10: 6–15.
- Tisseuil C, Leprieur F, Grenouillet G, Vrac M, Lek S (2013) Projected impacts of climate change on spatio-temporal patterns of freshwater fish beta diversity: A deconstructing approach. Global Ecology and Biogeography 21: 1213–1222. https://doi.org/10.1111/j.1466-8238.2012.00773.x
- Vannote RL, Minshall GW, Cummins KW et al. (1980) The river continuum concept. Canadian Journal of Fishery & Aquatic Science 37(2): 130–137. https://doi.org/10.1139/f80-017
- Wei SG, Li L, Xu R, Huang ZL, Cao HL (2015) Spatial Pattern and Interspecific Relationship of Dominant Species in Plant Community in Jinggang Mountain. Journal of Tropical and Subtropical Botany 23(1): 74–80.
- Wiersma YF, Urban DL (2005) Beta diversity and nature reserve system design in the Yukon, Canada. Conservation Biology 19: 1262–1272. https://doi.org/10.1111/j.1523-1739.2005.00099.x
- Wu YF, Tan Q J (1991) Characteristics of the fish-fauna of the characteristics of Qinghai-Xizang plateau and its geological distribution and formation. Acta Zoologica Sinica 37(2): 135–152.
- Xu YZ, Liu JJ, Cheng YX (2016) Characteristics and ecological risk assessment of heavy metals contamination in sediments of the Xiangjiang River. Environmental Chemistry 35(1): 189–198.
- Yan YZ, Xiang XY, Chu L, Zhan YJ, Fu CZ (2011) Influences of local habitat and stream spatial position on fish assemblages in a dammed watershed, the Qingyi stream, China. Ecology of Freshwater Fish 20: 199–208. https://doi.org/10.1111/j.1600-0633.2010.00478.x
- Yang YC, Li BY, Yin ZS, Zhang QS (1982) The formation and evolution of landforms in the Xizang Plateau. Acta Geographica Sinica 37: 76–87.
- Ye ZH, Liu JQ, Yin GS, Yan ZB (2013) Landform Features and Formation Mechanism in Mt. Jinggangshan, Jiangxi, China. Journal of Mountain Science 31(2): 250–256.
- Yue PQ (2000) Fauna sinica, osteichthyes, cypriniformes III. Beijing, China: Science Press.
- Zhang E, Chen YY (1997) Fish fauna in northeastern Jiangxi Province with a discussion on the zoogeographical division of east china. Acta Zoologica Sinica 21(3): 254–261.
- Zhang D, Wan FY, Chu L (2018) Longitudinal patterns in α and β diversity of the taxonomic and functional organizations of stream fish assemblages in the Qingyi River. Biodiversity Science 26(1): 1–13. https://doi.org/10.17520/biods.2017263
- Zhang JM, Wu ZQ, Hu ML (2010) Resource status of four major Chinese carps in the Xiajiang reach of Ganjiang River. Journal of Hydrology 3(1): 34–37.
- Zhang LS (2012) Ancient Geography of China – the Formation of China’s Natural Environment. Science Press, Beijing.
- Zhao YH, Zhang CG (2001) Fish fauna and zoogeographical analysis of ShiWan Da Shan Mountains, Guangxi, China. Biodiversity Science 9(4): 336–344.
Appendix 1
Species occurrence in the streams of Luoxiao Mountains. 1 = presence of the species as native in the stream, 0 = the species is absent from the stream, 2 = the species is present in the stream, but non-native from this stream. JJ: Jinjiang River; YS: Yuanshui River; HS: Heshui River; SS: Shushui River; SC: Suichuan River; SY: Shangyou River; MS: Mishui River; ML: Miluo River; FS: Fushui River; XH: Xiuhe River; LY: Liuyang River.
| Species | JJ | YS | HS | SS | SC | SY | MS | ML | FS | XH | LY |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Myxocyprinus asiaticus | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Zacco platypus | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Opsariichthys bidens | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Mylopharyngododon piceus | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 |
| Ctenopharyngodon idella | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 0 |
| Elopichthys bambusa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Squaliobarbus curriculus | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 |
| Hemiculter leucisculus | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 1 |
| Hemiculter bleekeri | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 0 |
| Hemiculterella sauvagei | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Hemiculterella wui | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 1 | 0 |
| Pseudohemiculter dispar | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Pseudolaubuca sinensis | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sinibrama macrops | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 0 |
| Chanodichthys erythropterus | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 |
| Culter alburnus | 0 | 1 | 0 | 1 | 0 | 1 | 1 | 0 | 1 | 0 | 0 |
| Chanodichthys mongolicus | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Chanodichthys dabryi | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 1 |
| Culter oxycephaloides | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| Parabramis pekinensis | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | 0 |
| Megalobrama terminalis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| Megalobrama amblycephala | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 0 |
| Xenocypris macrolepis | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 0 |
| Xenocypris davidi | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| Plagiognathops microlepis | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Distoechodon tumirostris | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 |
| Hypophthalmichthys molitrix | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 1 |
| Hypophthalmichthys nobilis | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 1 |
| Abbottina rivularis | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 1 |
| Pseudorasbora parva | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | 0 | 1 |
| Pseudogobio vaillanti | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Pseudogobio guilinensis | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Hemibarbus labeo | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Hemibarbus maculatus | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 1 |
| Huigobio chenhsienensis | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sarcocheilichthys sinensis | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 1 |
| Sarcocheilichthys kiangsiensis | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Sarcocheilichthys nigripinnis | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 |
| Squalidus argentatus | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 1 |
| Rhinogobio typus | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 |
| Platysmacheilus exiguus | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Saurogobio dabryi | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 1 |
| Saurogobio xiangjiangensis | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 |
| Microphysogobio kiatingensis | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Microphysogobio fukiensis | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 |
| Gobiobotia filifer | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 |
| Gobiobotia longibarba | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Acheilognathus macropterus | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 1 |
| Acheilognathus gracilis | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 1 |
| Acheilognathus chankaensis | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 0 |
| Acheilognathus tonkinensis | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 1 |
| Acheilognathus barbatulus | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Rhodeus ocellatus | 0 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Rhodeus lighti | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 |
| Acrossocheilus fasciatus | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 |
| Acrossocheilus paradoxus | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 |
| Acrossocheilus hemispinus | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 |
| Acrossocheilus parallens | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 |
| Spinibarbus hollandi | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 |
| Onychostoma barbatulum | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Carassius auratus | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Cyprinus carpio | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Garra orientalis | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 |
| Cobitis sinensis | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 0 |
| Misgurnus anguillicaudatus | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Paramisgurnus dabryanus | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Schistura fasciolata | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Schistura incerta | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Leptobotia elongata | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Parabotia banarescui | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Parabotia fasciata | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Parabotia maculosa | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Erromyzon sinensis | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Lepturichthys fimbriata | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 |
| Vanmanenia stenosoma | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 0 |
| Vanmanenia pingchowensis | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Pseudogastromyzon changtingensis | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Silurus asotus | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Silurus meridionalis | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Pterocryptis cochinchinensis | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Clarias fuscus | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Hemibagrus macropterus | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 1 |
| Pseudobagrus crassilabris | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 |
| Pseudobagrus tenuis | 0 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 0 |
| Pseudobagrus ondon | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| Pseudobagrus pratti | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Pseudobagrus albomarginatus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Tachysurus fulvidraco | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Tachysurus nitidus | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 |
| Liobagrus anguillicauda | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Liobagrus marginatus | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Liobagrus nigricauda | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Glyptothorax sinense | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 0 |
| Hyporhamphus intermedius | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| Monopterus albus | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Macrognathus aculeatus | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| Sinobdella sinensis | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | 0 |
| Siniperca chuatsi | 0 | 1 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 0 |
| Siniperca knerii | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Siniperca obscura | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Siniperca roulei | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Siniperca scherzeri | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 0 |
| Siniperca undulata | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 |
| Odontobutis sinensis | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| Rhinogobius cliffordpopei | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 |
| Rhinogobius duospilus | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Rhinogobius giurinus | 0 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 0 |
| Rhinogobius lindbergi | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Rhinogobius leavelli | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| Macropodus opercularis | 0 | 1 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 |
| Channa argus | 1 | 1 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 1 | 0 |
| Channa asiatica | 0 | 1 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 |
| Channa maculata | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 0 |
Appendix 2
| Order | Species (proportion) | Family | Species (proportion) | Subfamily | Species (proportion) |
| Cypriniformes | 77(68.1%) | Catostomidae | 1(0.9%) | Danioninae | 2(3.2%) |
| Cyprinidae | 62(54.9%) | Leuciscinae | 4(6.5%) | ||
| Cobitidae | 9(8.0%) | Culterinae | 15(24.2%) | ||
| Balitoridae | 5(4.4%) | Xenocyprininae | 4(6.5%) | ||
| Siluriformes | 16(14.2%) | Siluridae | 3(2.7%) | Hypophthalmichthyinae | 2(3.2%) |
| Clariidae | 1(0.9%) | Gobioninae | 17(27.4%) | ||
| Bagridae | 8(7.1%) | Gobiobotinae | 2(3.2%) | ||
| Amblycipitidae | 3(2.7%) | Acheilognathinae | 7(11.3%) | ||
| Sisoridae | 1(0.9%) | Barbinae | 6(9.7%) | ||
| Beloniformes | 1(0.9%) | Hemiramphidae | 1(0.9%) | Cyprininae | 2(3.2%) |
| Syngnathiformes | 3(2.7%) | Syngnathidae | 1(0.9%) | Labeoninae | 1(1.6%) |
| Mastacembelidae | 2(1.8%) | ||||
| Perciformes | 16(14.2%) | Percichthyidae | 6(5.3%) | ||
| Odontobutidae | 1(0.9%) | ||||
| Gobiidae | 5(4.4%) | ||||
| Belontiidae | 1(0.9%) | ||||
| Channidae | 3(2.7%) | ||||
| Total | 113(100%) | 113(100%) | 62(100%) |
Appendix 3
Endangered categories of fish species in the Luoxiao Mountains. Key: DD: Data Deficient; LC: Least Concern; NT: Near Threatened; VU: Vulnerable; EN: Endangered; CR: Critically Endangered.
| Species | Endangered categories | |
|---|---|---|
|
|
|
|
| Myxocyprinus asiaticus | CR | DD |
| Zacco platypus | LC | DD |
| Opsariichthys bidens | LC | LC |
| Mylopharyngododon piceus | LC | DD |
| Ctenopharyngodon idella | LC | DD |
| Elopichthys bambusa | LC | DD |
| Squaliobarbus curriculus | LC | DD |
| Hemiculter leucisculus | LC | LC |
| Hemiculter bleekeri | LC | DD |
| Hemiculterella sauvagei | LC | LC |
| Hemiculterella wui | LC | DD |
| Pseudohemiculter dispar | LC | VU |
| Pseudolaubuca sinensis | LC | LC |
| Sinibrama macrops | LC | LC |
| Chanodichthys erythropterus | LC | LC |
| Culter alburnus | LC | DD |
| Chanodichthys mongolicus | LC | LC |
| Chanodichthys dabryi | LC | LC |
| Culter oxycephaloides | LC | DD |
| Parabramis pekinensis | LC | DD |
| Megalobrama terminalis | LC | DD |
| Megalobrama amblycephala | LC | LC |
| Xenocypris macrolepis | LC | LC |
| Xenocypris davidi | LC | DD |
| Plagiognathops microlepis | LC | LC |
| Distoechodon tumirostris | LC | LC |
| Hypophthalmichthys molitrix | LC | NT |
| Hypophthalmichthys nobilis | LC | DD |
| Abbottina rivularis | LC | DD |
| Pseudorasbora parva | LC | LC |
| Pseudogobio vaillanti | LC | LC |
| Pseudogobio guilinensis | LC | DD |
| Hemibarbus labeo | LC | DD |
| Hemibarbus maculatus | LC | DD |
| Huigobio chenhsienensis | LC | LC |
| Sarcocheilichthys sinensis | LC | LC |
| Sarcocheilichthys kiangsiensis | LC | DD |
| Sarcocheilichthys nigripinnis | LC | DD |
| Squalidus argentatus | LC | DD |
| Rhinogobio typus | LC | DD |
| Platysmacheilus exiguus | LC | LC |
| Saurogobio dabryi | LC | DD |
| Saurogobio xiangjiangensis | LC | DD |
| Microphysogobio kiatingensis | DD | LC |
| Microphysogobio fukiensis | DD | LC |
| Gobiobotia filifer | LC | DD |
| Gobiobotia longibarba | DD | DD |
| Acheilognathus macropterus | LC | DD |
| Acheilognathus gracilis | LC | DD |
| Acheilognathus chankaensis | LC | DD |
| Acheilognathus tonkinensis | LC | DD |
| Acheilognathus barbatulus | LC | LC |
| Rhodeus ocellatus | LC | DD |
| Rhodeus lighti | LC | LC |
| Acrossocheilus fasciatus | LC | DD |
| Acrossocheilus paradoxus | LC | DD |
| Acrossocheilus hemispinus | LC | LC |
| Acrossocheilus parallens | LC | LC |
| Spinibarbus hollandi | LC | DD |
| Onychostoma barbatulum | NT | DD |
| Carassius auratus | LC | LC |
| Cyprinus carpio | LC | VU |
| Garra orientalis | LC | LC |
| Cobitis sinensis | LC | LC |
| Misgurnus anguillicaudatus | LC | LC |
| Paramisgurnus dabryanus | LC | DD |
| Schistura fasciolata | DD | DD |
| Schistura incerta | DD | DD |
| Leptobotia elongata | VU | VU |
| Parabotia banarescui | LC | DD |
| Parabotia fasciata | LC | LC |
| Parabotia maculosa | LC | LC |
| Erromyzon sinensis | DD | DD |
| Lepturichthys fimbriata | DD | LC |
| Vanmanenia stenosoma | DD | DD |
| Vanmanenia pingchowensis | DD | DD |
| Pseudogastromyzon changtingensis | DD | DD |
| Silurus asotus | LC | LC |
| Silurus meridionalis | LC | LC |
| Pterocryptis cochinchinensis | LC | LC |
| Clarias fuscus | LC | LC |
| Hemibagrus macropterus | LC | LC |
| Pseudobagrus crassilabris | LC | DD |
| Pseudobagrus tenuis | DD | DD |
| Pseudobagrus ondon | DD | LC |
| Pseudobagrus pratti | VU | DD |
| Pseudobagrus albomarginatus | LC | DD |
| Tachysurus fulvidraco | LC | LC |
| Tachysurus nitidus | LC | DD |
| Liobagrus anguillicauda | DD | DD |
| Liobagrus marginatus | VU | DD |
| Liobagrus nigricauda | DD | EN |
| Glyptothorax sinense | LC | DD |
| Hyporhamphus intermedius | LC | DD |
| Monopterus albus | LC | LC |
| Macrognathus aculeatus | LC | DD |
| Sinobdella sinensis | DD | LC |
| Siniperca chuatsi | LC | DD |
| Siniperca knerii | LC | DD |
| Siniperca obscura | NT | LC |
| Siniperca roulei | VU | DD |
| Siniperca scherzeri | LC | DD |
| Siniperca undulata | NT | NT |
| Odontobutis sinensis | LC | DD |
| Rhinogobius cliffordpopei | LC | DD |
| Rhinogobius duospilus | DD | DD |
| Rhinogobius giurinus | LC | LC |
| Rhinogobius lindbergi | DD | DD |
| Rhinogobius leavelli | LC | LC |
| Macropodus opercularis | LC | LC |
| Channa argus | LC | DD |
| Channa asiatica | LC | LC |
| Channa maculata | LC | LC |