





(C) 2011 Loris Galli. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
The Italian Protura were studied basing on 5103 specimens from 198 sampling areas, along with bibliographic data from 49 collecting sites. 17 out of the 20 Italian regions are covered. As a result, 40 species have been identified (belonging to 8 genera and 4 families), 6 of which are new records for the Italian fauna.
A key to the Italian species is reported, followed by a series of distribution maps and brief remarks for some of them. A preliminary biogeographical overview allowed us to delineate the chorological categories of these species, 10 of which are actually known only in Italy. The comparison with the species richness known for some best studied Central and Eastern European Countries leads us to speculate that widening our research, Italian Protura check-list will be much implemented.
Protura, Italy, distribution, key to species
Protura is a group of Hexapoda which has been discovered recently: the first species described is Acerentomon doderoi, collected from soil samples taken from the grounds of a small villa actually in the center of Genoa (
Knowledge of Protura has rapidly increased all over the world thanks to the careful research of many specialists. Just to mention the main publications, in 1964 Tuxen published his valuable book about the Protura of the World; Nosek’s monograph on European Protura was printed in 1973; a year later Imadaté’s volume about Japanese species was released (1974); while the impressive monograph about Chinese Protura was published more recently (
The European research about this group, although with some exceptions, was concentrated in Central and Eastern Countries due to the work of some Authors such as Nosek, Rusek, Tuxen and, more recently, Szeptycki and Shrubovych.
In Italy, knowledge regarding Protura (see species list for detailed bibliographic references) can be summarized with the identification of 31 species belonging to the Italian fauna by the national check-list (
With this paper we hope to lay the foundations for the advancement and improvement of studies regarding this little known taxon in Italy as well in the Mediterranean Region, one of the biodiversity hotspots on the planet, reaching the highest peaks of diversity of soil-borne organisms (e.g.
Many of the Protura examined in this paper were collected by colleagues and given us in tubes containing 70% ethanol. However, we have obtained some specimens by extraction from soil or litter samples by Berlese-Tullgren funnels (2.5 mm mesh size). Specimens were incubated at 40–50° C for 24 hours in lactic acid to clarify, mounted on slides in Marc André medium and were observed and identified by an interference contrast microscope.
In total 5103 specimens from 198 sampling areas were examined. 3929 specimens were identified to species level (Table 1).
Number of Protura specimens examined from each Italian region.
| Regions | Nr of specimens (bibliographic data excluded) | Nr of specimens identifiable to species level (bibliographic data excluded) |
| Aosta Valley | 134 | 94 |
| Piedmont | 748 | 546 |
| Lombardy | 214 | 189 |
| Trentino-Alto Adige | 40 | 20 |
| Veneto | 193 | 161 |
| Friuli-Venezia Giulia | 68 | 58 |
| Liguria | 2878 | 2158 |
| Emilia-Romagna | 149 | 143 |
| Tuscany | 264 | 228 |
| Marches | 3 | 2 |
| Umbria | 45 | 46 |
| Lazio | 64 | 51 |
| Abruzzo | 22 | 16 |
| Molise | - | - |
| Campania | - | - |
| Apulia | 16 | 7 |
| Basilicata | 67 | 45 |
| Calabria | - | - |
| Sicily | 33 | 26 |
| Sardinia | 165 | 139 |
| Total | 5103 | 3929 |
In our analysis we also considered the data taken from 49 Italian collecting sites known in literature. 17 out of 20 Italian regions are covered, missing specimens from Molise, Campania and Calabria (Southern part of the peninsula).
Key to genera of Italian ProturaThis key and the following ones to species are based, and adapted to the Italian fauna, on
| 1 | Tracheal system present (meso and metanotum with spiracles); all three pairs of abdominal legs two segmented, with terminal vescicle and with 5 setae on each | Eosentomidae – Genus Eosentomon |
| – | Spiracles absent | Acerentomoidea 2 |
| 2 | Only the first pair of abdominal legs with a terminal vescicle and 4 setae; pairs II and III unsegmented with 2–3 setae; abdominal segment VIII with a more or less developed striate band | Acerentomidae 4 |
| – | First two pairs of abdominal legs with terminal vescicle; third pair unsegmented | 3 |
| 3 | Maxillary gland with a long dilatated sausage–like part; pseudoculus pear–like with a long and broad S shaped median opening; 8 setae in the anterior row of abdominal tergites II–VII | Hesperentomidae – Ionescuellum condei (Nosek, 1965) |
| – | At most 4 setae in the anterior row of abdominal tergites II–VII; maxillary gland with heart–shaped or circular dilatation; pseudoculi without median opening | Protentomidae 7 |
| 4 | Abdominal legs II and III with 3 setae (a longer median one and two shorter sub–apical) | Genus Acerentulus |
| – | Abdominal legs II and III with 2 setae | 5 |
| 5 | Abdominal legs II and III with 2 setae of the same length; maxillary gland with racemose appendix; sensillum of labial palp broad | Genus Acerella |
| – | Subapical seta of abdominal legs II and III shorter than the median one | 6 |
| 6 | Abdominal legs II and III with a long median seta and a very short subapical one; pseudoculi small; striate band of tergite VIII complete; maxillary gland with a rather large calyx, heart shaped; tuft of setae on labial palp strongly reduced; anterior row of abdominal sternites I–VII with 3 setae; sternite VIII with a single row of 4 setae | Genus Gracilentulus |
| – | Subapical seta of abdominal legs II and III half the length of the median one or less; head with a rostrum (from very short to long); anterior row of abdominal sternites I–VII with a variable number (≥ 3) of setae | Genus Acerentomon |
| 7 | Pseudoculus with a large triangular proximal prolongation; the “lever” of the same length as the pseudoculus itself and of almost the same width distally; the comb on tergite VIII with distinct teeth | Genus Proturentomon |
| – | Pseudoculus often more elliptical and proximal prolongation usually narrower parallel sided; the comb on tergite VIII with very fine teeth or toothless | Genus Protentomon |
Since this key could lead to a misidentification of similar Palearctic species not already detected in Italy, we suggest a careful examination of the species’ descriptions (and redescriptions) to verify the identification accuracy and also to refer to the keys to species of other European Countries (e.g. those cited at the beginning of the key to genera) as well as to the monographic papers published on certain genera (e.g.
Genus Eosentomon
| 1 | Tergite VII with 6 anterior setae | 2 |
| – | Tergite VII with 4 anterior setae | 3 |
| 2 | Head with only posterior additional seta; seta p2’ on nota shorter than p3’ | Eosentomon transitorium Berlese, 1909 |
| – | Head with only posterior additional seta; seta p2’ roughly the same length of p3’ | Eosentomon germanicum Prell, 1912 |
| 3 | Tergites IV–VI missing seta p4’; chaetotaxy of sternite XII 8/7 | Eosentomon foroiuliense Torti & Nosek, 1984 |
| – | Tergites IV–VI with seta p4’; chaetotaxy of sternite XII 8/4 | 4 |
| 4 | Tergites II–VI missing seta p3’ | Eosentomon romanum Nosek, 1969 |
| – | Tergites II–VI with seta p3’; head with both anterior and posterior additional setae; seta p2’ on nota subequal or longer than p3’ | 5 |
| 5 | On tergite VII seta p1’ situated at the same level and near the base of p2 | Eosentomon delicatum Gisin, 1945 |
| – | On tergite VII seta p1’ placed close to the posterior border and p2’ in a cavity on the hind margin | 6 |
| 6 | Sensillum c’ behind the line α6–δ5; body length 750 μm; pseudoculus fairly big (PR = 7.5) | Eosentomon noseki Tuxen, 1982 |
| – | Sensillum c’ proximally to line α6–δ5; body length 1610 μm; PR = 8.6–11.6 | Eosentomon armatum Stach, 1926 |
Genus Acerentulus
| 1 | Sensillum a long reaching nearly or passing seta γ3; sensillum b subequal or shorter than c | 4 |
| – | Sensillum a of medium length or short, not reaching or barely reaching seta γ3 | 2 |
| 2 | Sensillum b subequal or shorter than c | 3 |
| – | Sensillum b much longer than c, reaching the empodium | Acerentulus traegardhi Ionescu, 1937 |
| 3 | Tergites II–VI without seta p1’ | Acerentulus cunhai Condé, 1950 |
| – | Tergites II–VI with seta p1’ | Acerentulus tuxeni Rusek, 1966 |
| 4 | Sensillum a’ broad, relatively short, not reaching the base of b’ | 5 |
| – | Sensillum a’ broad, long, reaching the base of b’ | 8 |
| 5 | Seta p3’ missing in tergite VII | 6 |
| – | Seta p3’ present in tergite VII | 7 |
| 6 | Seta a1 missing in tergite VII | Acerentulus exiguus Condé, 1944 |
| – | Seta a1 present in VII | Acerentulus apuliacus Rusek & Stumpp, 1988 |
| 7 | Setae a1 and p1’ missing in tergite VII | Acerentulus gisini Condé, 1952 |
| – | Setae a1 and p1’ present in tergite VII | Acerentulus confinis (Berlese, 1908) |
| 8 | Setae a1 and p1’ missing in tergite VII | 9 |
| – | Tergite VII: seta a1 present, p1’ missing | Acerentulus terricola Rusek, 1965 |
| 9 | Seta p1 present in tergite VIII; sternite XI with 4 setae | Acerentulus condei Nosek, 1983 |
| Seta p1 missing in tergite VIII; sternite XI with 6 setae | Acerentulus alpinus Gisin, 1945 |
Genus Acerella
| 1 | Sensillum t2 nearly 3 times the length of t1 | Acerella tiarnea (Berlese, 1908) |
| – | Sensillum t2 one and a half to twice the length of t1 | Acerella muscorum (Ionescu, 1930) |
Genus Gracilentulus
| 1 | Sensillum b not reaching seta γ3 | 2 |
| – | Sensillum b passing seta γ3 | Gracilentulus gracilis (Berlese, 1908) |
| 2 | Chaetotaxy of tergites II–VI 7/16; TR = 2, 7 | Gracilentulus sardinianus Nosek, 1979 |
| – | Chaetotaxy of tergites II–VI 8/14; TR = 3, 3 | Gracilentulus meridianus (Condé, 1945) |
Genus Acerentomon
| 1 | Chaetotaxy of sternite VIII 4/0 | 2 |
| – | Chaetotaxy of sternite VIII 4/2 | 3 |
| 2 | Seta x present; rostrum very long, LR = 3, 3 | Acerentomon noseki Torti, 1981 |
| – | Seta x absent; rostrum short, LR ≥ 6 | 4 |
| 3 | Seta x present; rostrum long, LR = 3, 5–4, 7 | 7 |
| – | Seta x absent; rostrum of medium length, LR = 4, 5–5 | Acerentomon affine Bagnall, 1912 |
| 4 | Rostrum short, LR nearly 6 | 5 |
| Rostrum very short, LR ≥ 9 | 6 | |
| 5 | Sensillum b extremely broad, not spindle–shaped and shorter than c; a long and reaching the base of e; pleural pectines strongly developed | Acerentomon meridionale Nosek, 1960 |
| – | Sensillum b broad, spear shaped, almost reaching seta γ4 and longer than c; a short, barely reaching the base of d; pleural pectines only on segments VI–VII | Acerentomon balcanicum Ionescu, 1933 |
| 6 | Comb VIII with 10–14 teeth; pleural pectin VI with a row of long teeth; ratio of sensilla a:b = 1, 1 | Acerentomon microrhinus Berlese, 1909 |
| – | Comb VIII with 17–20 teeth; pleural pectin VI strongly reduced to a group of 4 distinct teeth; ratio of sensilla a:b = 0, 8 | Acerentomon condei Nosek & Dallai, 1982 |
| 7 | Sensillum b thin and small, distinctly shorter than c | 8 |
| – | Sensillum b distinctly broad, subequal or longer than c | 10 |
| 8 | Head with additional setae | Acerentomon gallicum Ionescu, 1933 |
| – | Head without additional setae | 9 |
| 9 | Sensillum a short, barely reaching d; pleural line VI with a fine serration | Acerentomon italicum Nosek, 1969 |
| – | Sensillum a long, extending beyond the base of d, sometimes even reaching e; pleural line VI with a row of conspicuous teeth | Acerentomon fageticola Rusek, 1966 |
| 10 | Seta p3’ present in tergite VII | Acerentomon doderoi Silvestri, 1907 |
| – | Seta p3’ missing in tergite VII | 11 |
| 11 | Comb VIII with 14–16 long teeth, the median ones smaller; body length about 1600 μm | Acerentomon maius Berlese, 1908 |
| – | Comb VIII with 9–12 pointed teeth; body length 1980–2370 μm | Acerentomon baldense Torti, 1986 |
Genus Proturentomon
| 1 | Chaetotaxy of tergites I–VI 0/12 | Proturentomon noseki (Rusek, 1975) |
| – | Chaetotaxy of tergites II–VI 2/12 | 2 |
| 2 | Chaetotaxy of tegites I and VIII 2/10 and 6/12, respectively; sensillum b distinctly shorter than c | Proturentomon minimum Berlese, 1908 |
| – | Chaetotaxy of tegites I and VIII 2/12 and 6/14, respectively; sensillum b subequal or longer than c | 3 |
| 3 | Body length 690 μm; comb on tergite VIII with 8 teeth | Proturentomon condei Nosek, 1967 |
| – | Body length 500 μm; comb on tergite VIII with 4 long and thin teeth | Proturentomon pilosum (Rusek, 1975) |
Genus Protentomon
| 1 | Tergites II–VI without the anterior row of setae; sternite XI with 4 setae | Protentomon perpusillum (Berlese, 1909) |
| – | Tergites II–VI with 2 setae in the anterior row; sternite XI with 6 setae | Protentomon berlesei Nosek, 1969 |
This section provides summaries on species known to date belonging to Italian fauna. For each one the amount of material examined (PI = pre-imago, MJ = maturus junior, LII = larva II, LI = larva I, undet = undetermined), a short description of the global distribution from
ORDO: ACERENTOMATA Familia: Hesperentomidae Price, 1960
4 ♂♂, 4 ♀♀.
Austria, Italy.

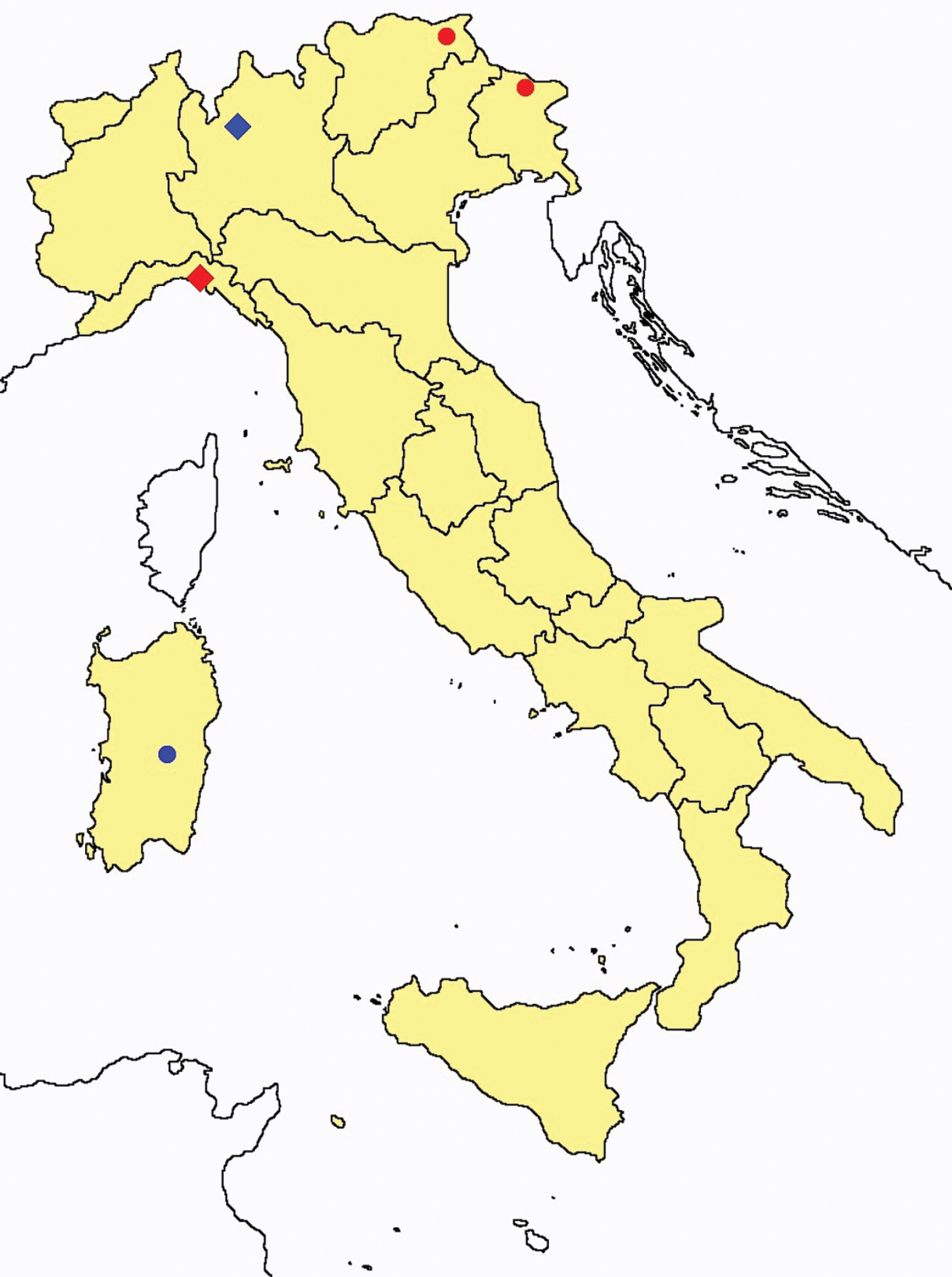
Ionescuellum condei: collecting sites in Italy (red dots: samples personally analyzed by the authors).
First Italian record in
2 ♂♂, 2 ♀♀.
Veneto, Colli Euganei near Padua.
Italy.
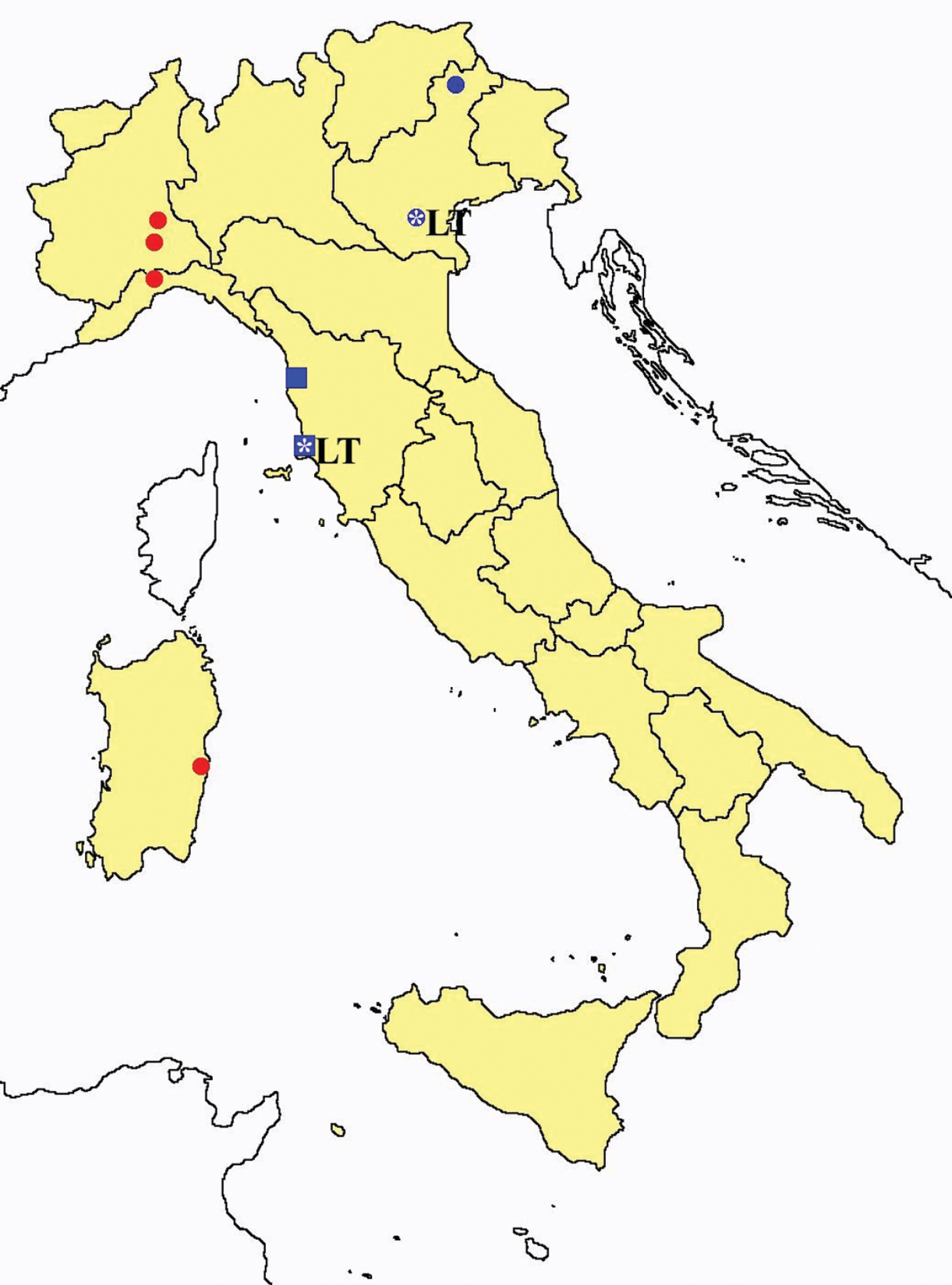
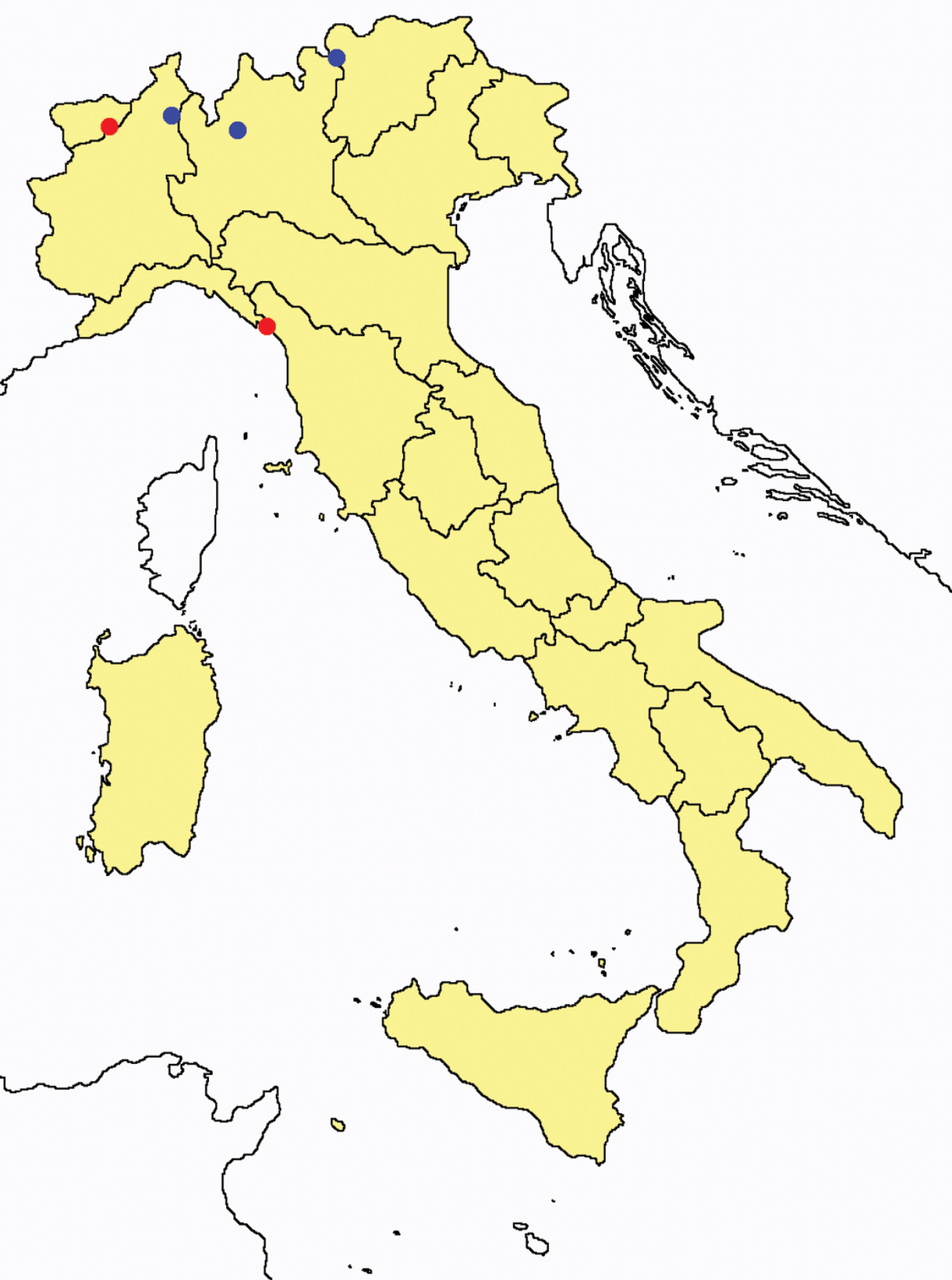
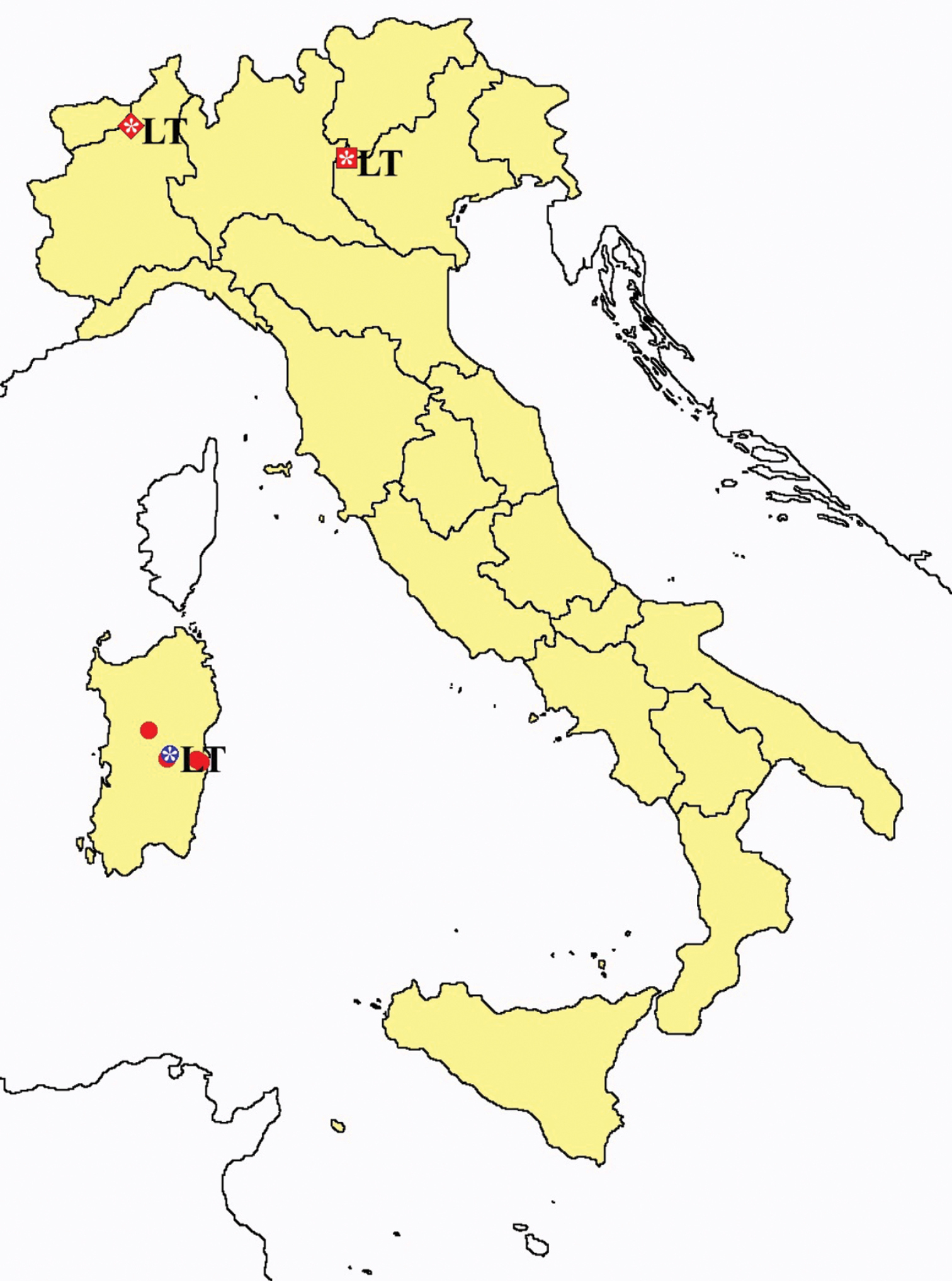
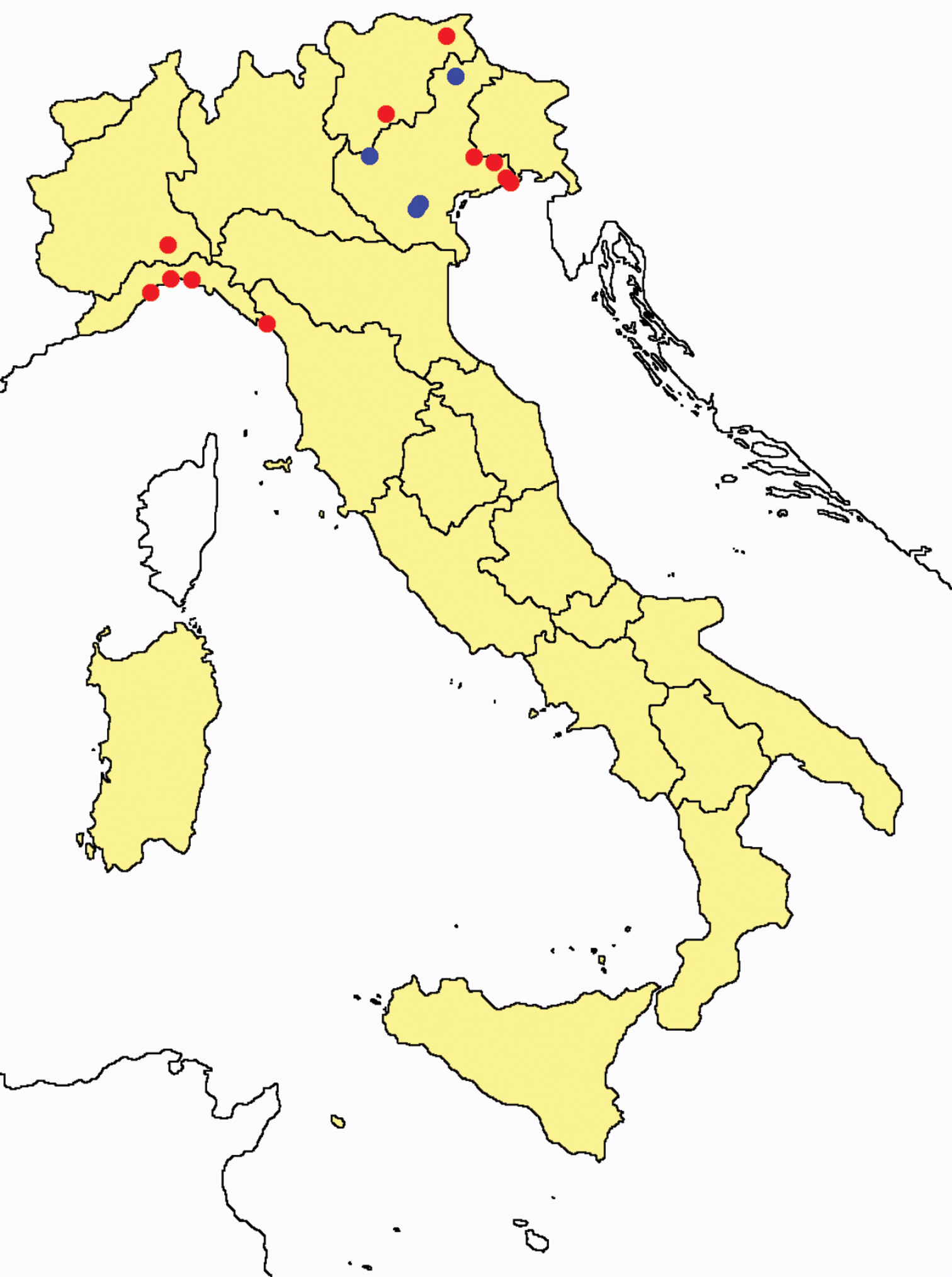
Protentomon spp.: collecting sites in Italy (dots Protentomon berlesei; squares: Protentomon perpusillum; blue: data from literature; red: samples personally analyzed by the authors; LT = type area).
For nearly 40 years since its description (
Tuscany, S. Vincenzo (Livorno).
Italy, Germany. Data from Denmark and Australia should be confirmed (
Bibliographic data from
6 ♀♀, 1 MJ.
Austria, Slovakia.
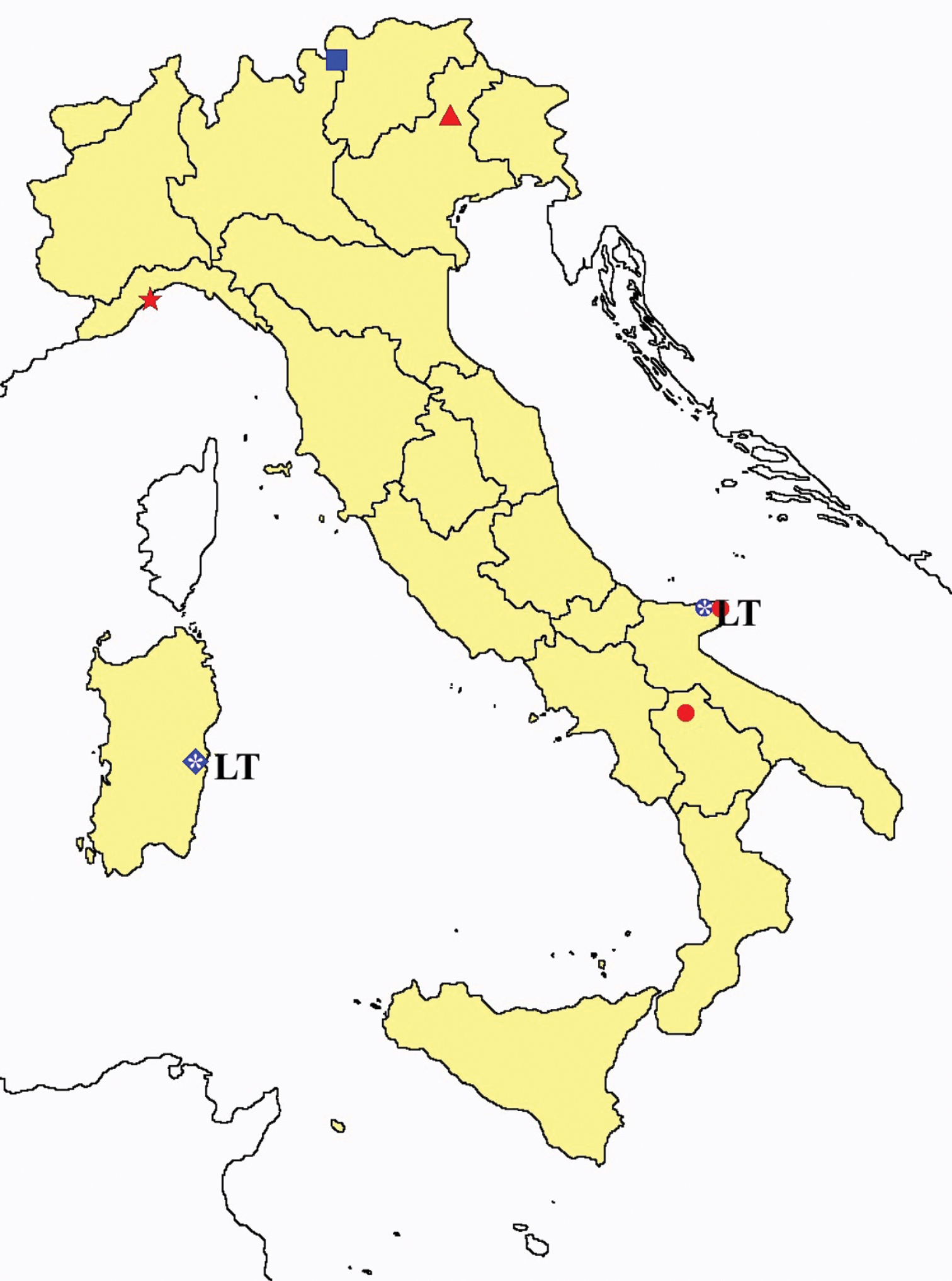
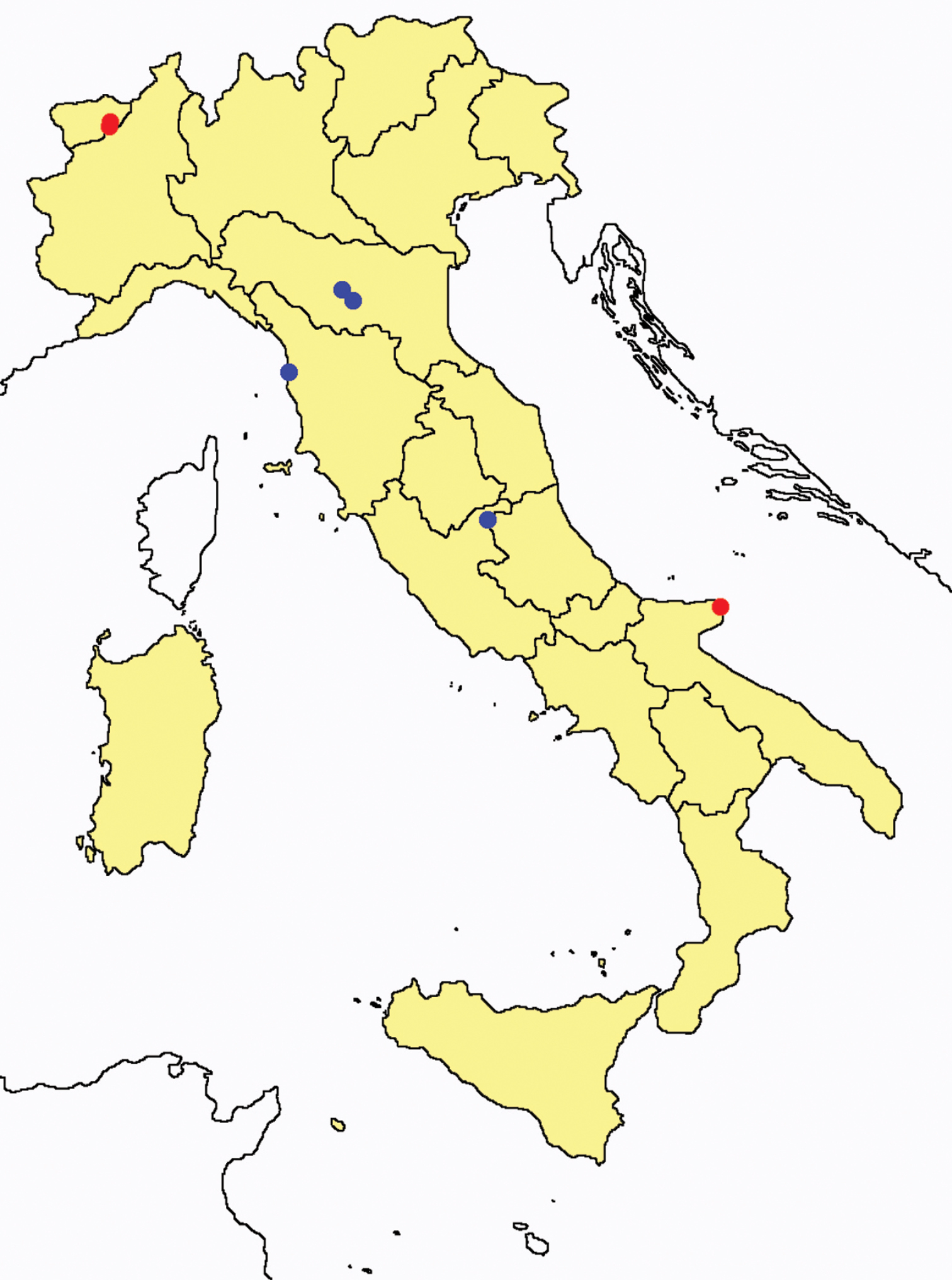
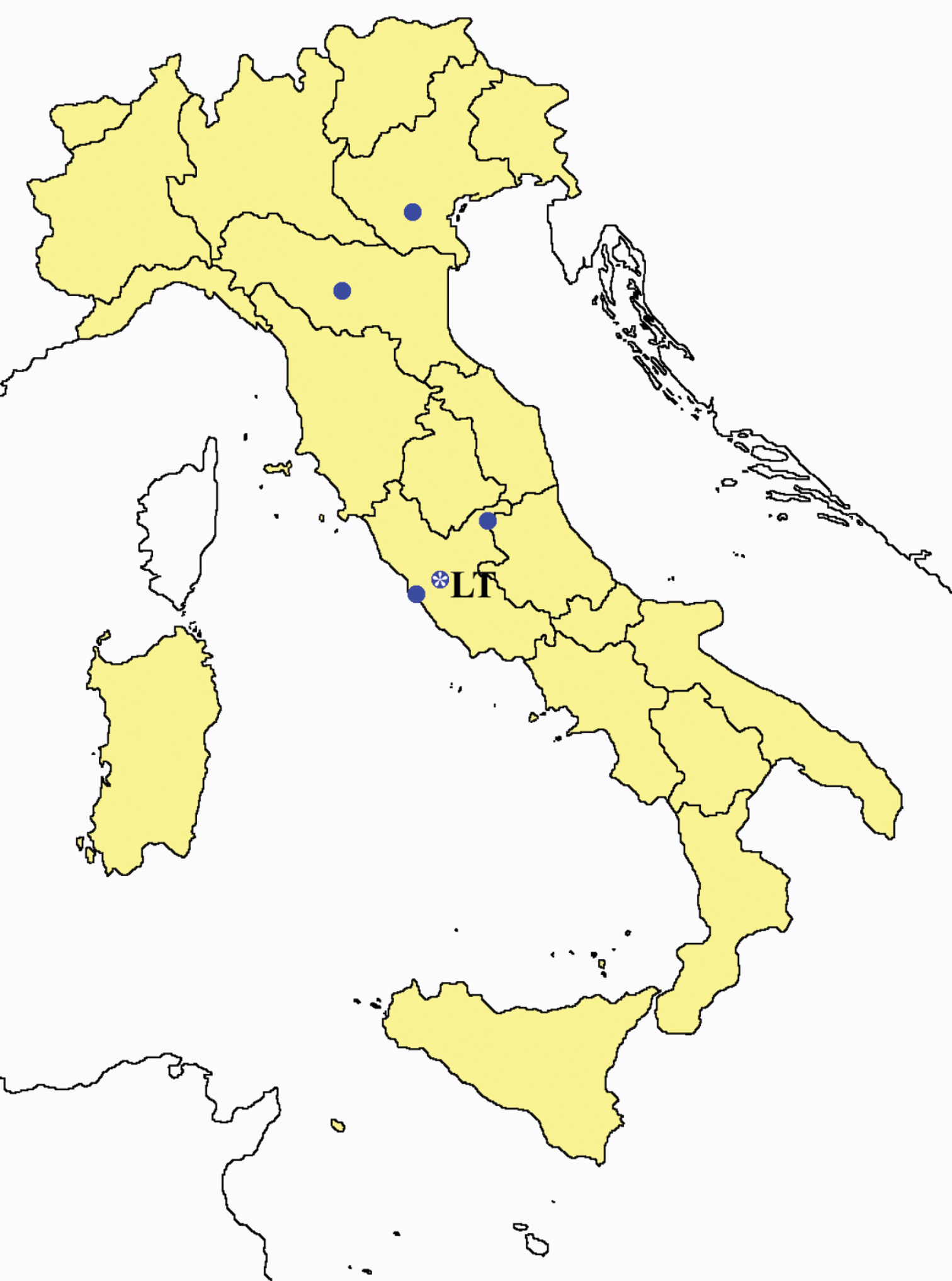
Proturentomon spp.: collecting sites in Italy (squares Proturentomon condei; dots Proturentomon minimum; star: Proturentomon noseki; triangle Proturentomon pilosum; blue: data from literature; red: samples personally analyzed by the authors; LT = type area).
This species is not included in the World Catalogue (
11 ♀♀, 7 MJ, 1 undet.
Tuscany, Giardino di Boboli in Florence.
Recorded from nearly whole Europe (with exception of Scandinavia), but all of the older data should be confirmed (
Bibliographic data from Berlese (1908),
2 ♀♀ (Vignale Monferrato, Alessandria, Piedmont).
Central Europe.
New record for the Italian fauna.
1 ♀ (Concordia, Venice, Veneto).
Central Europe.
New record for the Italian fauna.
South Europe.
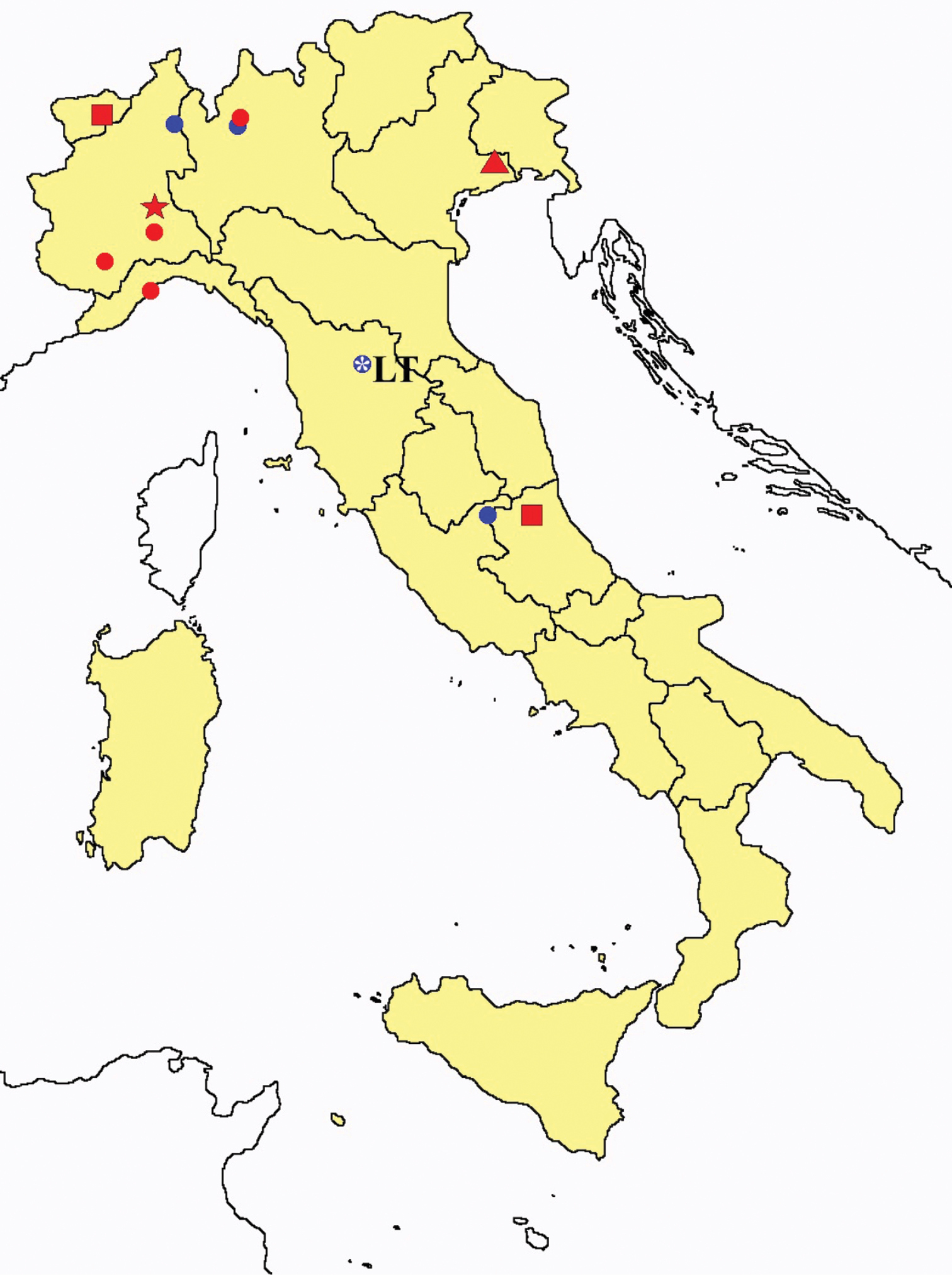
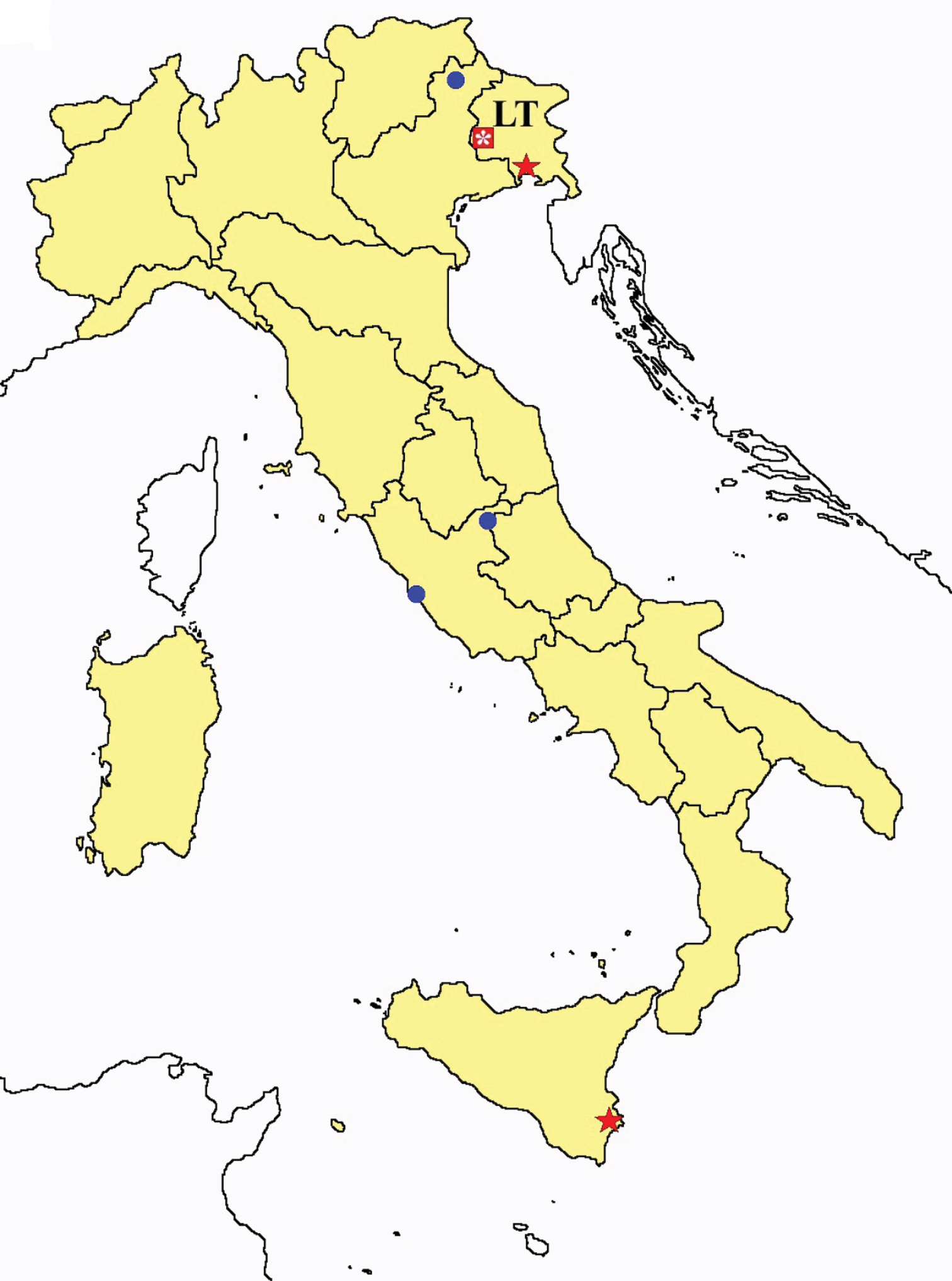
Acerentulus spp.: collecting sites in Italy (square: Acerentulus alpinus; dots: Acerentulus apuliacus; rhombus: Acerentulus condei; star: Acerentulus terricola; triangle: Acerentulus tuxeni; blue: data from literature; red: samples personally analyzed by the authors; LT = type area).
Bibliographic data from
3 ♂♂, 20 ♀♀, 3 PI, 1 MJ.
Apulia, 10 km South of Vico del Gargano, Bosco Sfilzi.
Type area only.
Bibliographic data from
Sardinia, Strada Orientale Sarda km 158.
Mediterranean Europe (Sardinia, Corsica, Slovenia).
Bibliographic data from
104 ♂♂, 187 ♀♀, 5 PI, 24 MJ.
Tuscany, Florence.
Recorded from nearly all Europe (with exception of Scandinavia), North Africa, North America and Australia. Most of the older data are dubious and should be confirmed (
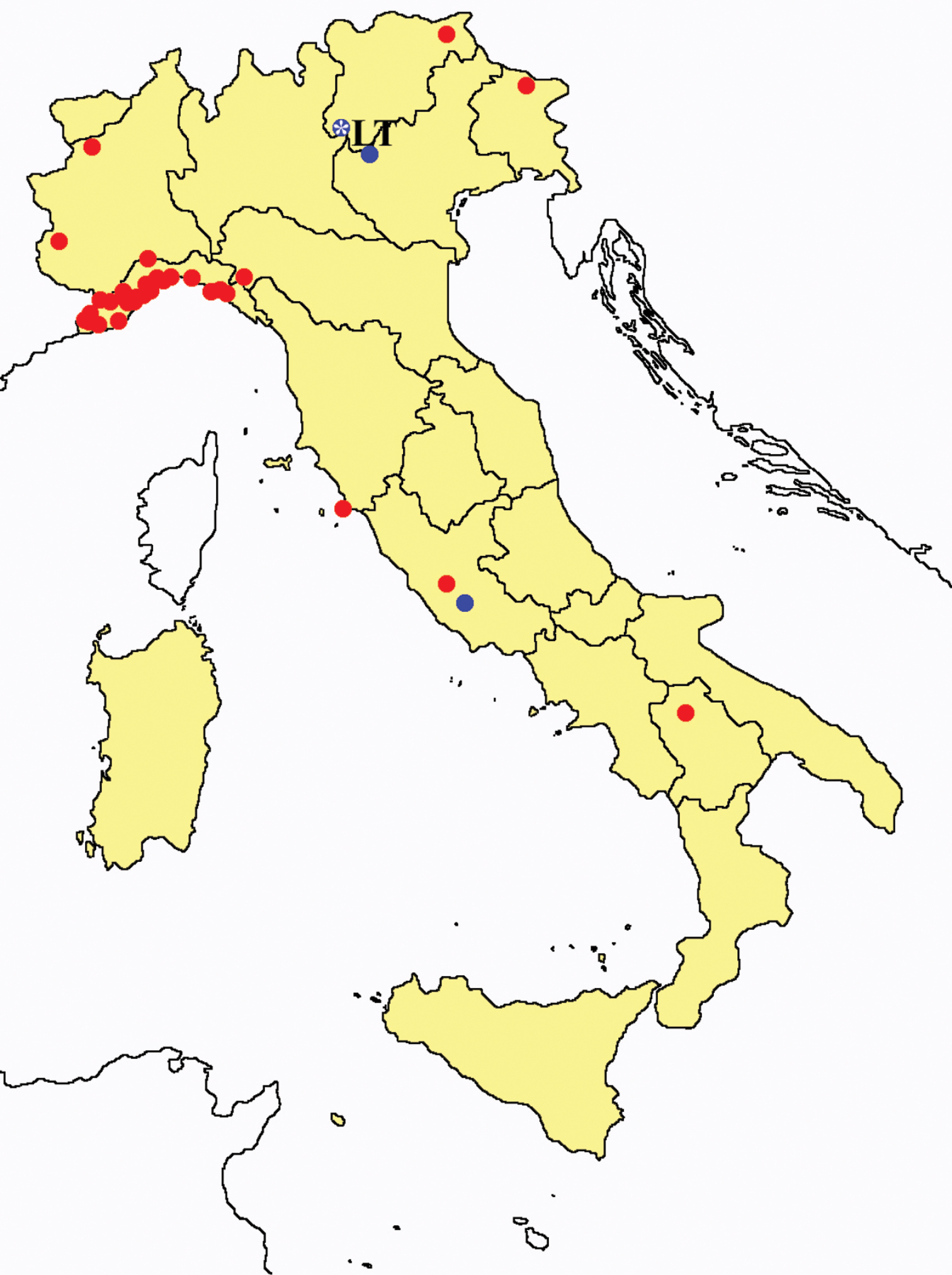
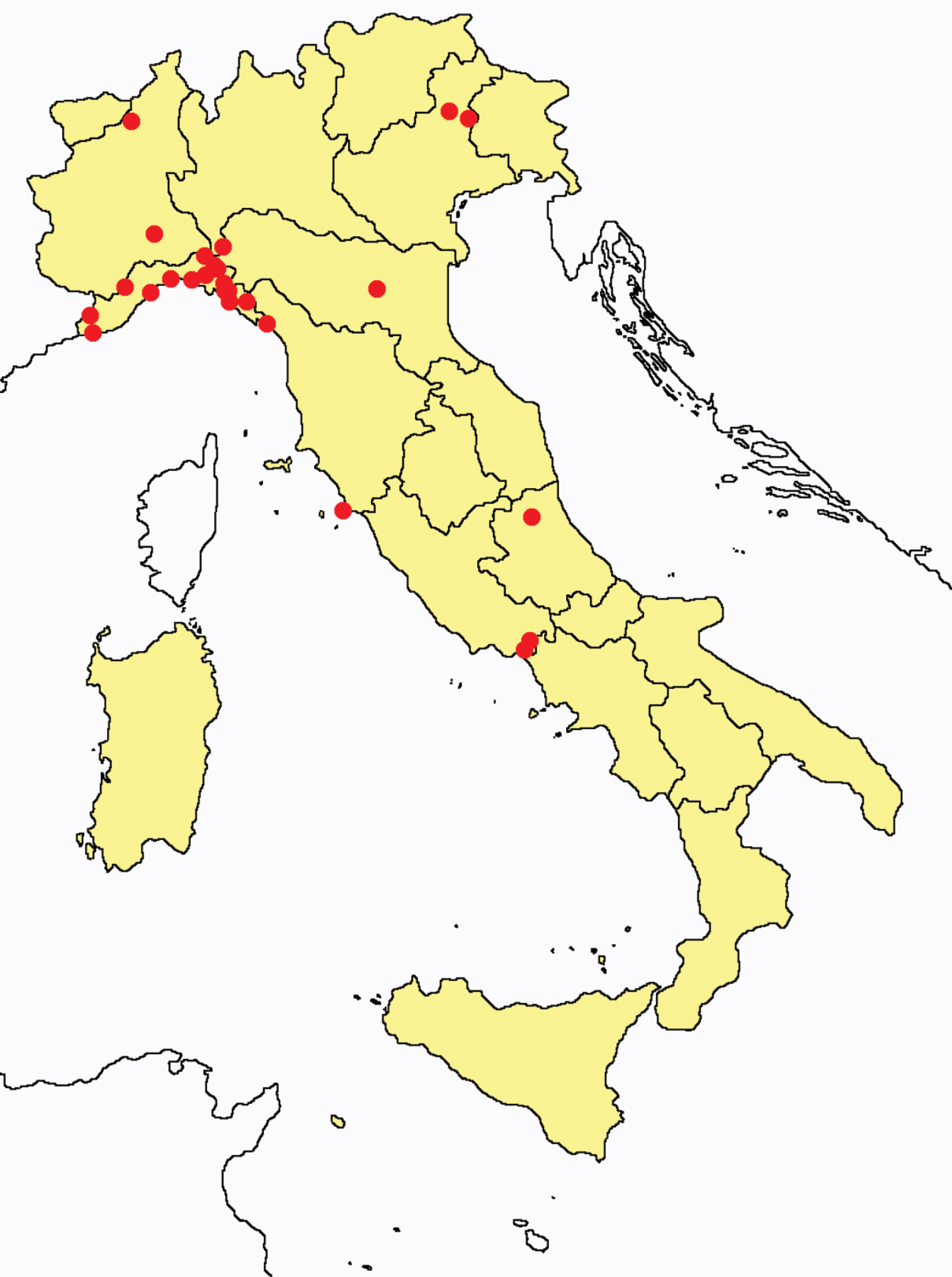
Acerentulus confinis: collecting sites in Italy (blue dots: data from literature; red dots: samples personally analyzed by the authors; LT = type area).
Bibliographic data from Berlese (1908),
1 ♀.
Central and West Europe, Macaronesia.
Acerentulus spp.: collecting sites in Italy (rhombus: Acerentulus cunhai; dots: Acerentulus exiguus; blue: data from literature; red: samples personally analyzed by the authors).
Bibliographic data from
1 ♂, 1 MJ.
Central and South Europe.
Bibliographic data from
3 ♀♀.
Central Europe, Italy; data from Bulgaria should be confirmed (
Acerentulus gisini: collecting sites in Italy (blue dots: data from literature; red dots: samples personally analyzed by the authors).
Bibliographic data from (
2 ♂♂ (Bergeggi, Savona, Liguria).
Czech Republic (type area: Czech Rep., ‘Tal Suchý _leb im Nordteil des Mährischen Karstes“).
New record for the Italian fauna.
15 ♂♂, 14 ♀♀, 5 PI, 16 MJ, 2 LII.
Recorded from nearly whole Europe, but it was commonly mistaken with Acerentulus insignis. Many data (especially from the West Europe) should be confirmed (
Acerentulus traegardhi: collecting sites in Italy (blue dots: data from literature; red dots: samples personally analyzed by the authors).
Bibliographic data from
3 ♂♂, 3 ♀♀ (Ponte delle Alpi, Belluno, Veneto).
Central Europe.
New record for the Italian fauna.
3 ♂♂, 5 ♀♀.
Tuscany, Toiana (Pisa).
Recorded from many European countries, from North Africa, South Africa, Australia and New Zealand.
Gracilentulus spp.: collecting sites in Italy (dots: Gracilentulus gracilis; square: Gracilentulus sardinianus; rhombus: Gracilentulus gracilis + Gracilentulus meridianus + Gracilentulus sardinianus; blue: data from literature; red: samples personally analyzed by the authors; LT = type area).
Bibliographic data from Berlese (1908),
4 ♂♂, 4 ♀♀ (Elini, Ogliastra, Sardinia).
France, Spain.
New record for the Italian fauna.
1 ♂, 3 ♀♀, 1 MJ.
Sardinia, between Luogosanto and Tempio Pausania.
Type area only.
Bibliographic data from
43 ♂♂, 63 ♀♀, 3 PI, 7 MJ, 1 undet.
West Europe; data from Romania and “Czechoslovakia” should be confirmed (
Acerentomon affine: collecting sites in Italy (blue dots: data from literature; red dots: samples personally analyzed by the authors).
Species confirmed for Italy. Bibliographic data from
17 ♂♂, 14 ♀♀, 1 PI, 1 MJ.
Southeast Europe, Ukraine.
Acerentomon balcanicum: collecting sites in Italy (blue dots: data from literature; red dots: samples personally analyzed by the authors).
Bibliographic data from
5 ♂♂, 7 ♀♀.
Type area: Veneto, Monte Balbo (Venetian PreAlps) surroundings of Prà Alpesina (Verona).
Type area only.
Bibliographic data from
6 ♂♂, 2 ♀♀, 1 MJ.
Sardinia, Desulo (Gennargento).
Type area only.
Bibliographic data from
64 ♂♂, 94 ♀♀, 5 PI, 2 MJ.
Liguria, Genoa.
Known only from Italy and Slovenia. All data from Central and West Europe and from USA are highly doubtful (
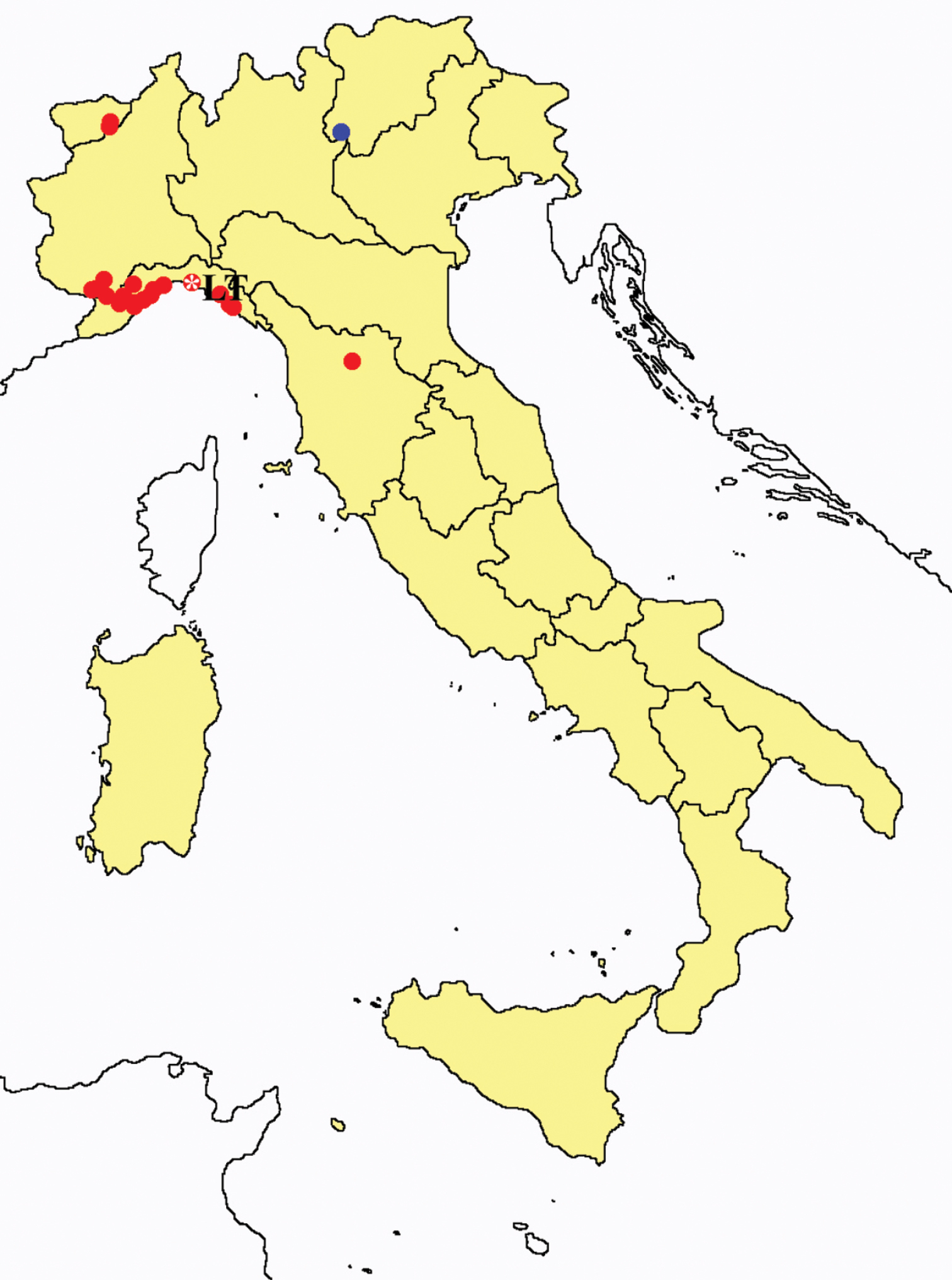
Acerentomon doderoi: collecting sites in Italy (blue dots: data from literature; red dots: samples personally analyzed by the authors; LT = type area).
We have not yet been able to analyse the type material from Villetta Dinegro (Genoa Town). Four specimens originally labelled as “cotypus doderoi” in Genoa Museum collection have been recently identified by the authors as Acerentomon italicum.
Central Europe.
Three specimens from Veneto (Cison, Treviso), and two from Liguria (Lavagna, Genoa) were identified by Prof. Nosek as Acerentomon fageticola and cited in a short note by
We hope that our current redescription of Acerentomon italicum could shed more light on the differences between this species and the related Acerentomon fageticola.
This species has been cited here and in the identification key only for exactness of information.
42 ♂♂, 90 ♀♀, 7 PI, 11 MJ, 6 LII, 1 LI, 1 undet.
West and Central Europe, recorded also from Africa (Uganda – introduced?).
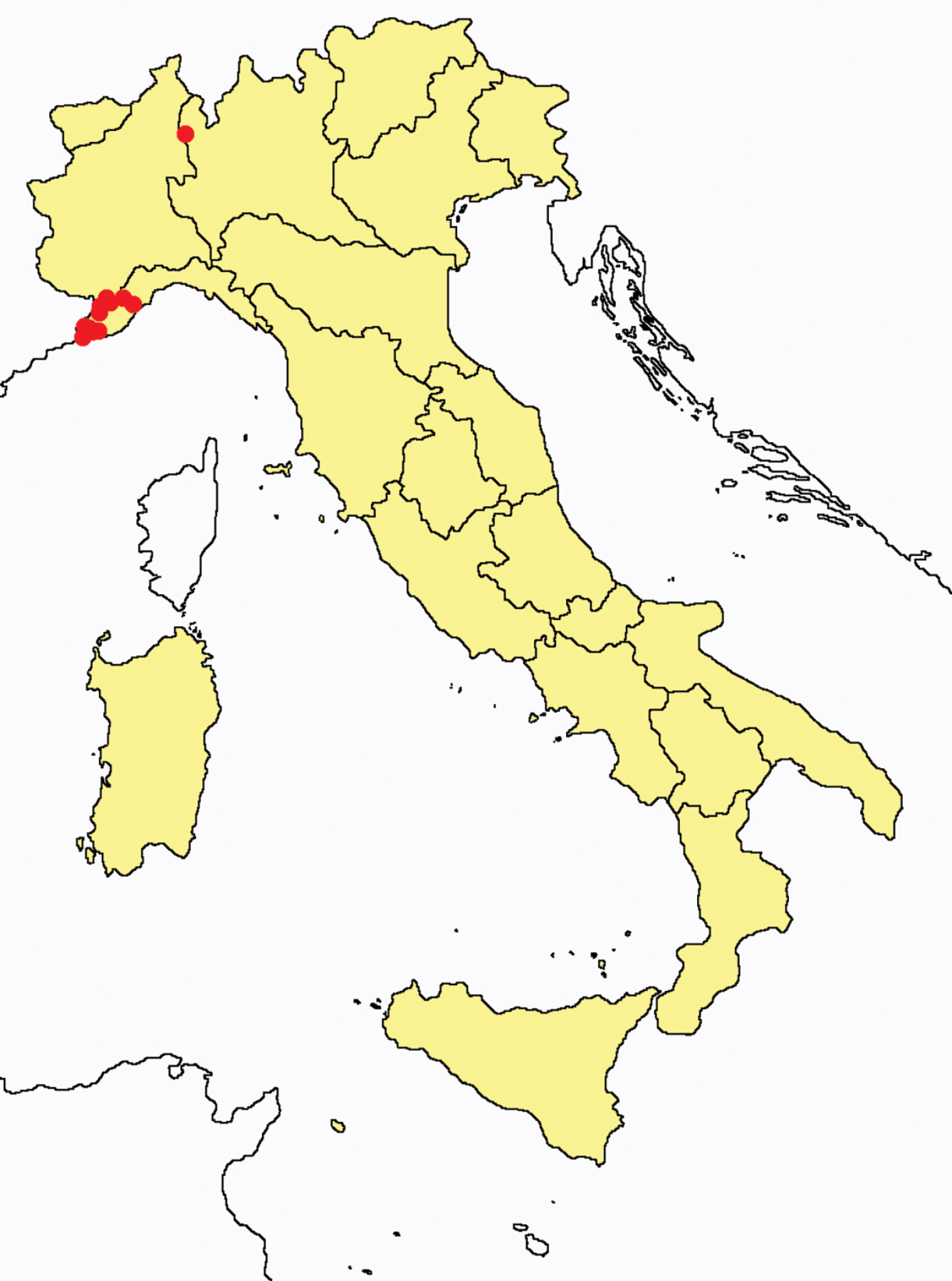
Acerentomon gallicum: collecting sites in Italy (red dots: samples personally analyzed by the authors).
Although in
433 ♂♂, 573 ♀♀, 18 PI, 16 MJ, 14 LII, 6 undet.
Veneto, Colli Euganei near Padua, Italy.
Italy.
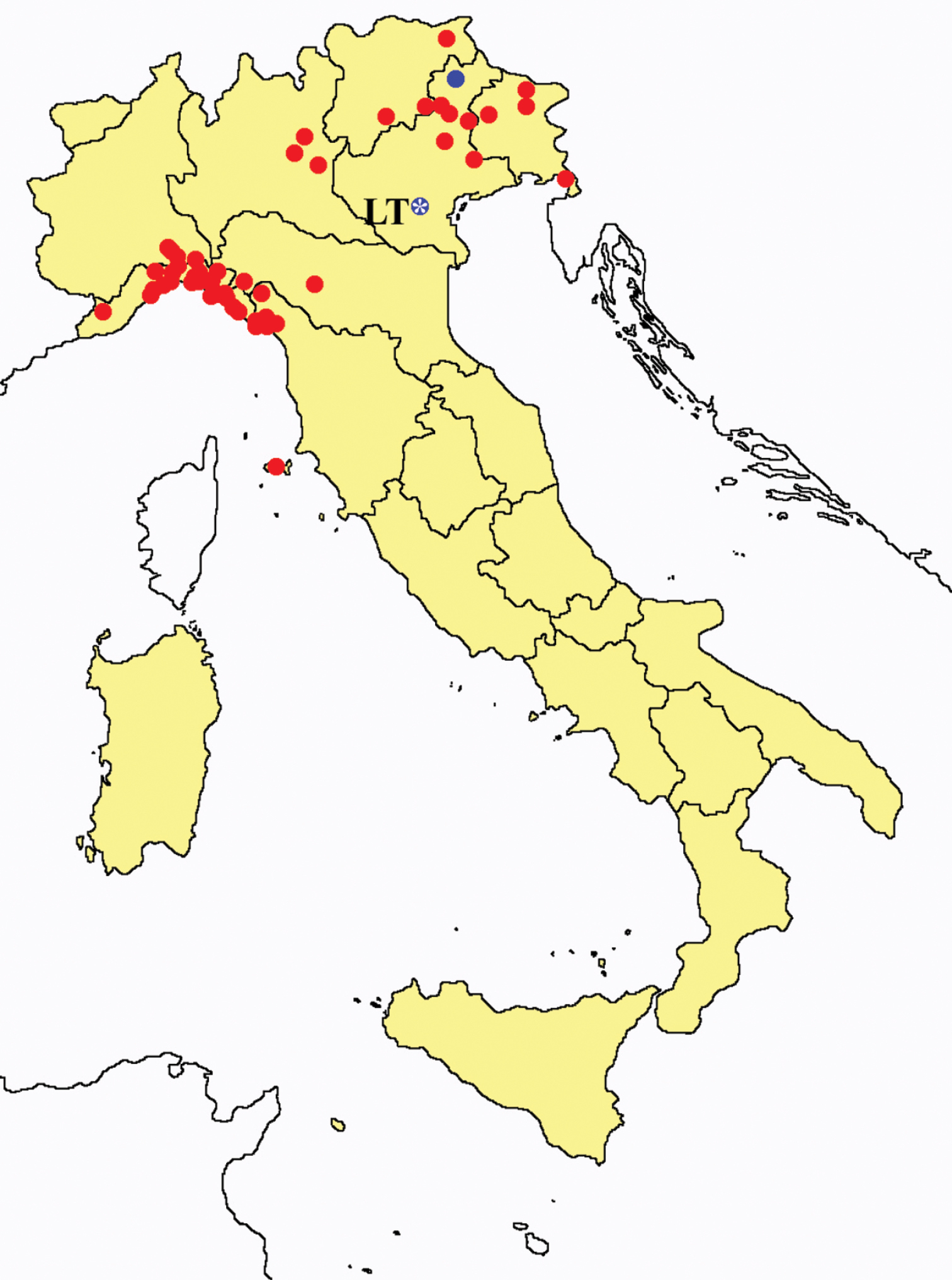
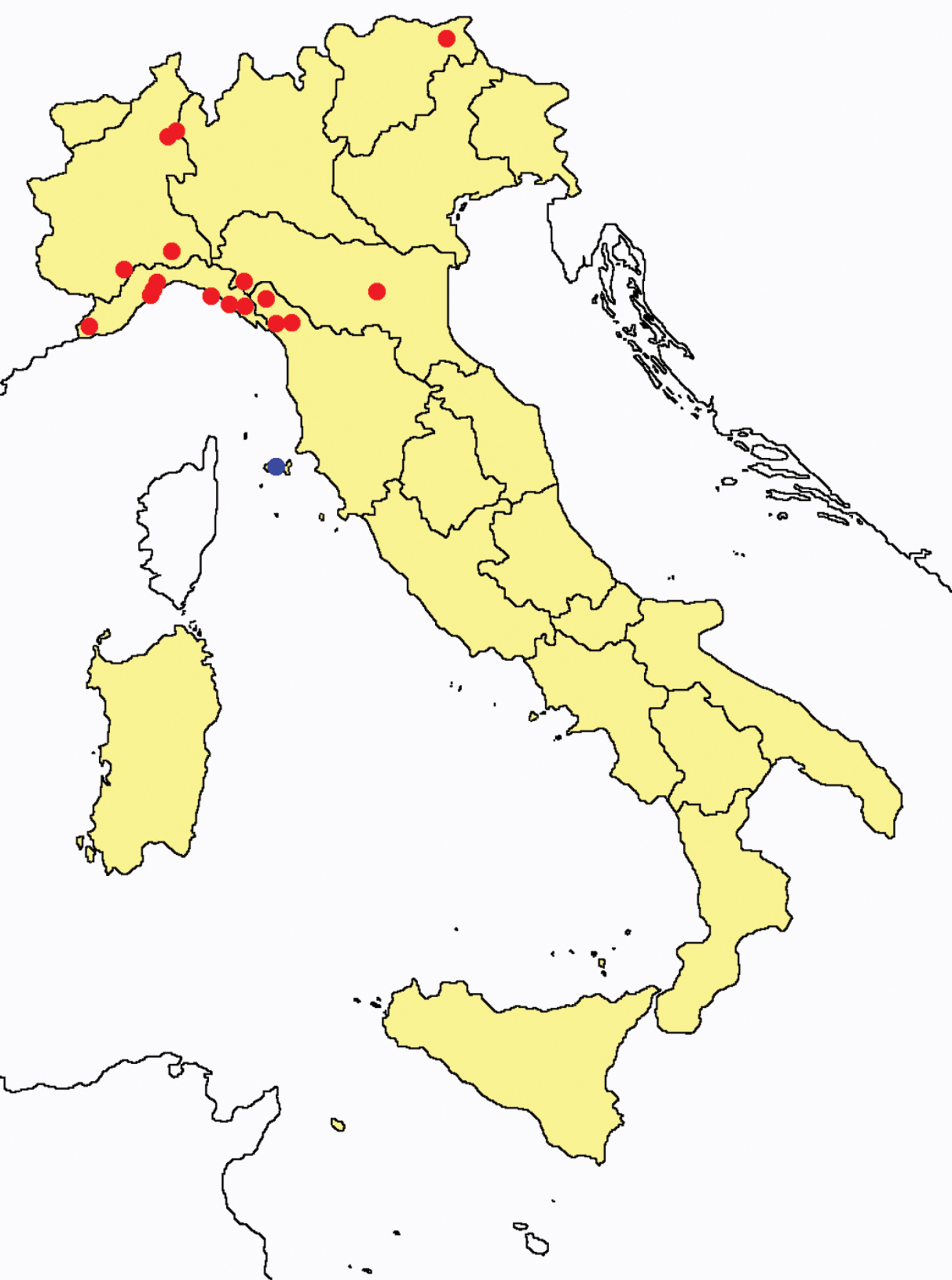
Acerentomon italicum: collecting sites in Italy (blue dots: data from literature; red dots: samples personally analyzed by the authors; LT = type area).
Species currently under redescription by the authors of this paper. Bibliographic data from (
353 ♂♂, 455 ♀♀, 40 PI, 25 MJ, 3 LII, 2 LI, 3 undet.
Trentino Alto Adige, Tiarno.
Italy, Central Europe.
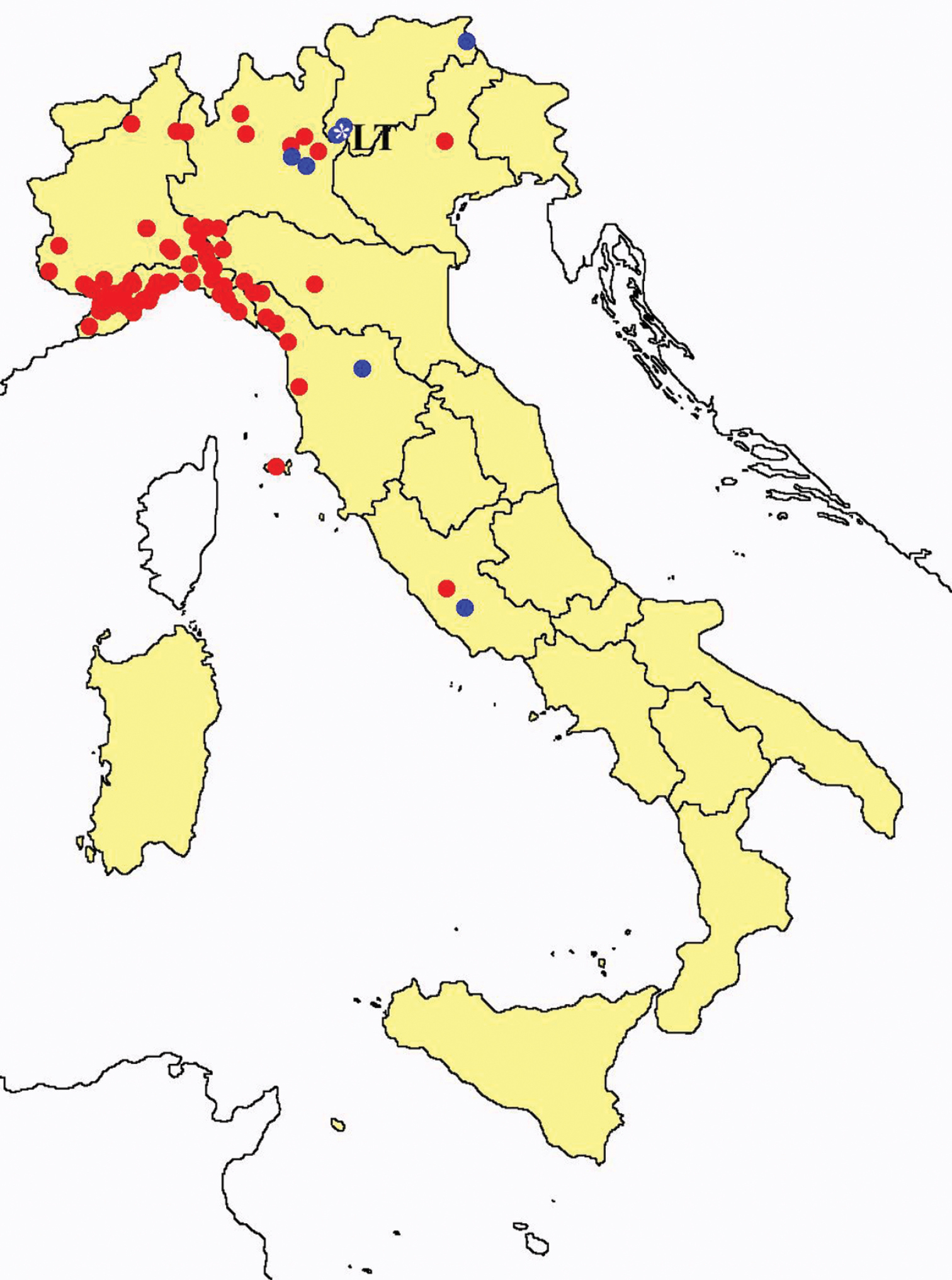
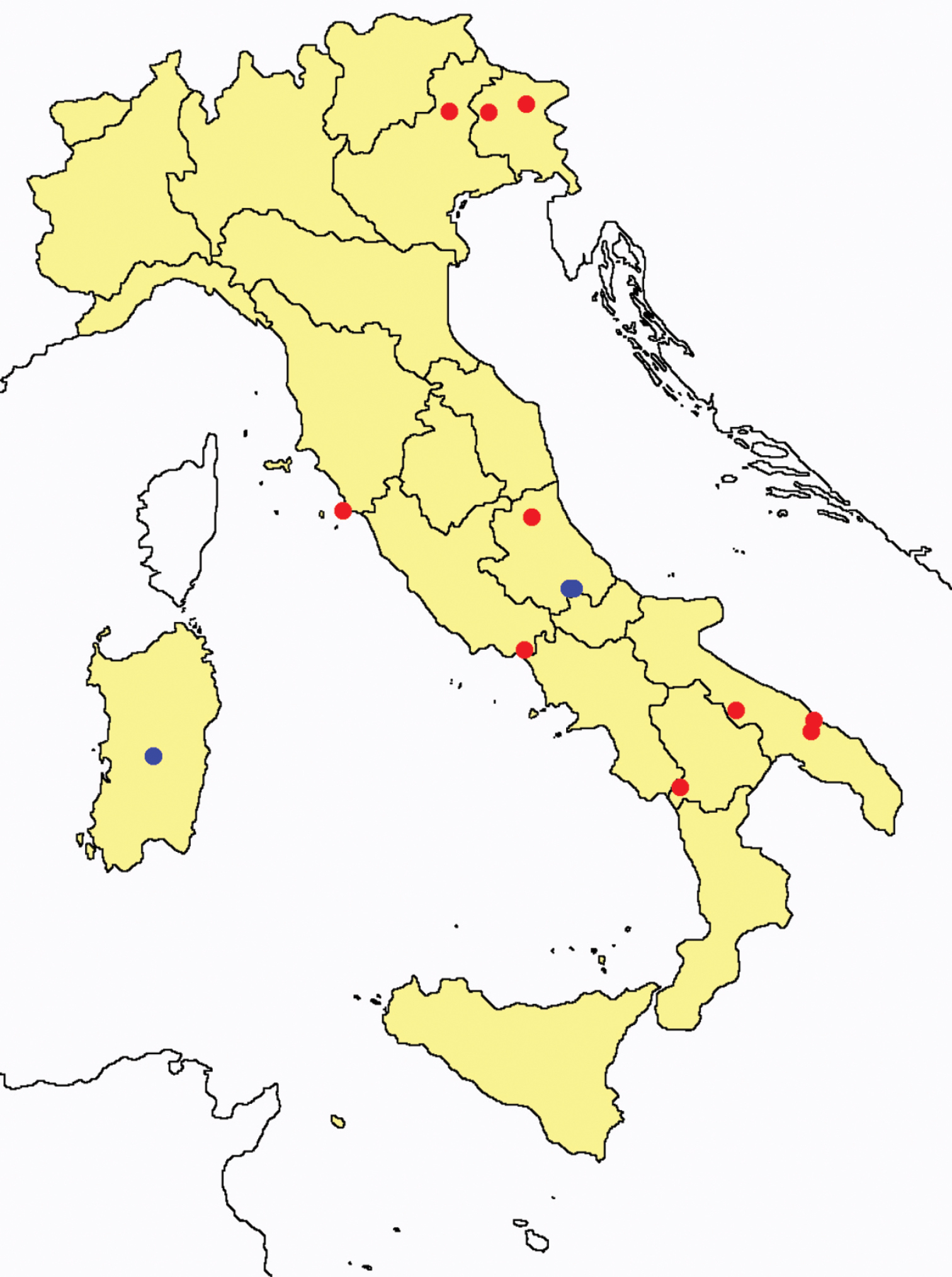
Acerentomon maius: collecting sites in Italy (blue dots: data from literature; red dots: samples personally analyzed by the authors; LT = type area).
Bibliographic data from Berlese (1908),
52 ♂♂, 93 ♀♀, 2 PI, 8 MJ, 1 undet.
South and Central Europe, Near East (Israel).
Acerentomon meridionale: collecting sites in Italy (blue dots: data from literature; red dots: samples personally analyzed by the authors).
Bibliographic data from
95 ♂♂, 161 ♀♀, 21 PI, 15 MJ, 1 LII, 2 undet.
Piedmont, Casale Monferrato.
South and Central Europe.
Acerentomon microrhinus: collecting sites in Italy (blue dots: data from literature; red dots: samples personally analyzed by the authors; LT = type area).
Bibliographic data from
2 ♀♀.
Piedmont, surroundings of Santuario di Oropa near Biella.
Type area only.
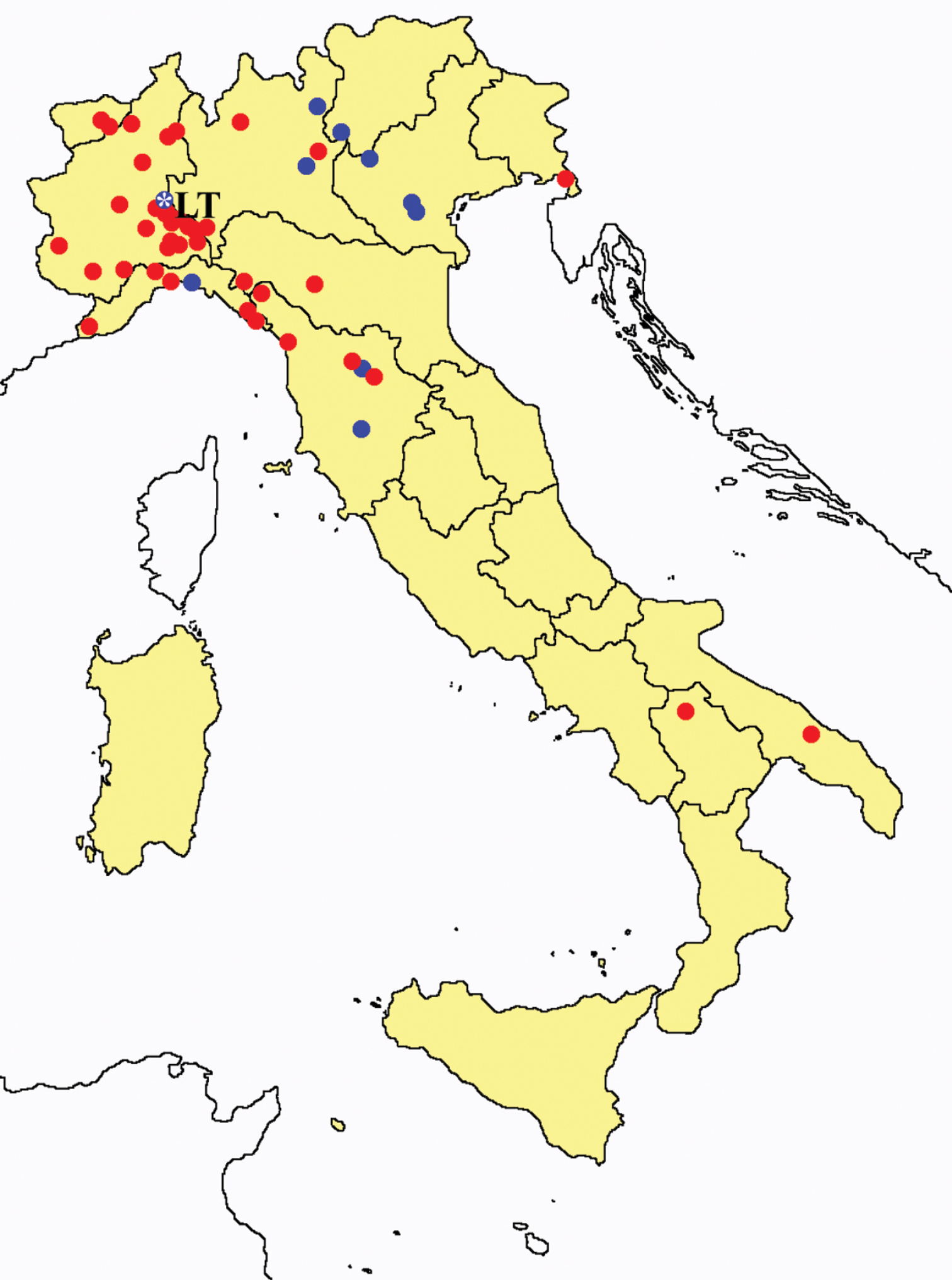
Acerentomon spp.: collecting sites in Italy (square: Acerentomon baldense; dots: Acerentomon condei; rhombus: Acerentomon noseki; blue: data from literature; red: samples personally analyzed by the authors; LT = type area).
Bibliographic data from
3 ♂♂, 6 ♀♀.
Central and West Europe, Near East.
Acerella muscorum: collecting sites in Italy (blue dots: data from literature; red dots: samples personally analyzed by the authors).
Bibliographic data from
30 ♂♂, 95 ♀♀, 1 PI, 1 MJ, 1 LI.
Trentino Alto Adige, Tiarno.
Mediterranean Europe; all data from the Central and North Europe should be checked (
Acerella tiarnea: collecting sites in Italy (blue dots: data from literature; red dots: samples personally analyzed by the authors; LT = type area).
Bibliographic data from Berlese (1908),
ORDO: EOSENTOMATA Familia: Eosentomidae Berlese, 1909
1 ♂ (Carlino, Udine, Friuli-Venezia Giulia) – 1 ♂, 6 ♀♀, 4 MJ (Floridia, Siracusa, Sicily).
Probably widely distributed in Europe, but all data before 1986 should be checked – they most likely concern not only Eosentomon armatum, but also some other similar species (
Eosentomon spp.: collecting sites in Italy (stars: Eosentomon armatum; square: Eosentomon foroiuliense; dots: Eosentomon germanicum; blue: data from literature; red: samples personally analyzed by the authors; LT = type area).
New record for the Italian fauna.
8 ♂♂, 9 ♀♀, 2 MJ, 1 LII, 1 undet.
Europe, North Africa.
Eosentomon delicatum: collecting sites in Italy (blue dots: data from literature; red dots: samples personally analyzed by the authors).
Bibliographic data from
http://species-id.net/wiki/Eosentomon_foroiuliense
Fig. 211 ♀.
Friuli-Venezia Giulia, Aviano.
Type area only.
Bibliographic data from
Central Europe, Scandinavia. The data from West Europe, Italy and Madeira (under Eosentomon germanicum and Eosentomon forsslundi) should be checked – Eosentomon germanicum was commonly mistaken with similar species (
Bibliographic data from
43 ♂♂, 43 ♀♀, 1 PI, 18 MJ, 2 LII.
Macaronesia, Spain.
Eosentomon noseki: collecting sites in Italy (red dots: samples personally analyzed by the authors).
This species is not included in the World Catalogue (
Lazio, Rome.
Italy.
Eosentomon romanum: collecting sites in Italy (blue dots: data from literature; LT = type area).
Bibliographic data from
112 ♂♂, 107 ♀♀, 38 MJ, 11 LII, 6 LI, 13 undet.
Tuscany, Florence.
Probably whole Europe and North Africa, but most of the data should be confirmed (

Eosentomon transitorium: collecting sites in Italy (blue dots: data from literature; red dots: samples personally analyzed by the authors; LT = type area).
Bibliographic data from Berlese (1908),
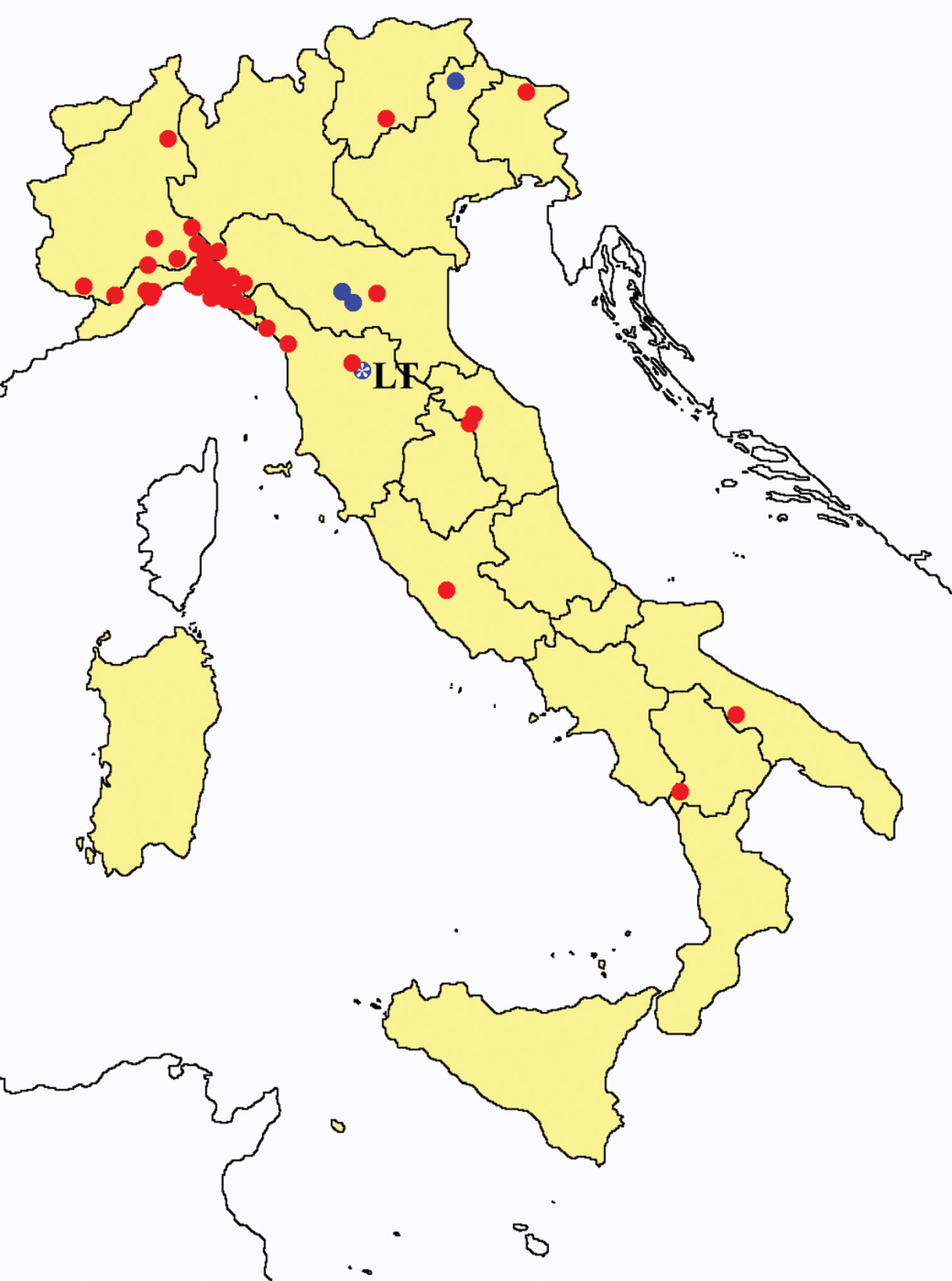
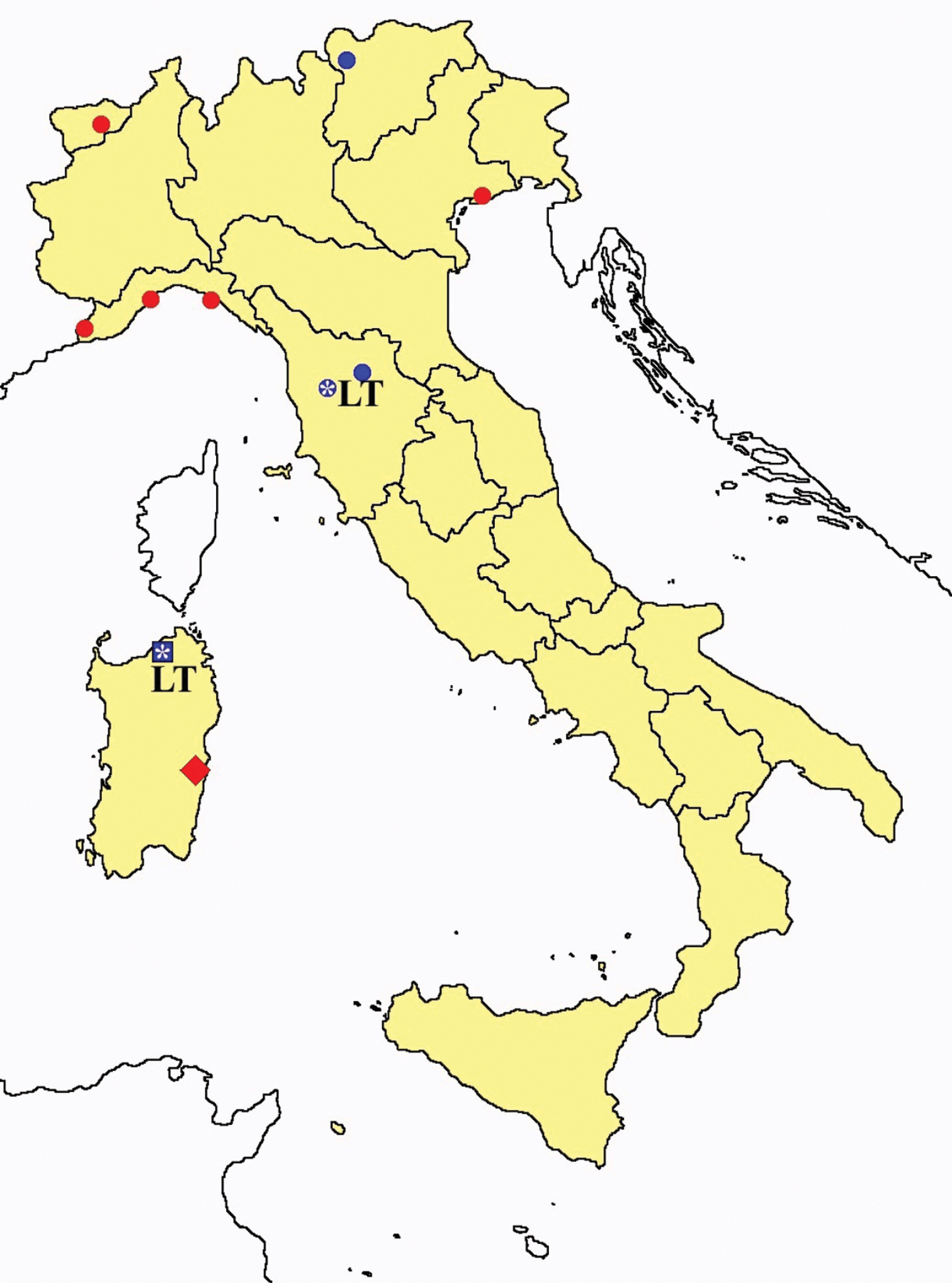
In Figures 26 and 27 the distribution of the sampling sites in Italy and the species richness in the Italian regions are shown, respectively. Unfortunately we regret for the lack of samples from Molise, Campania and Calabria (Southern part of the peninsula); but, apart from that, comparing maps on these Figures, it is clear that the species richness reflects the sampling effort in the different regions, with higher numbers of species known from regions such as Piedmont, Veneto and Liguria, where many more samples have been collected (for a detailed analysis of Protura of Liguria see
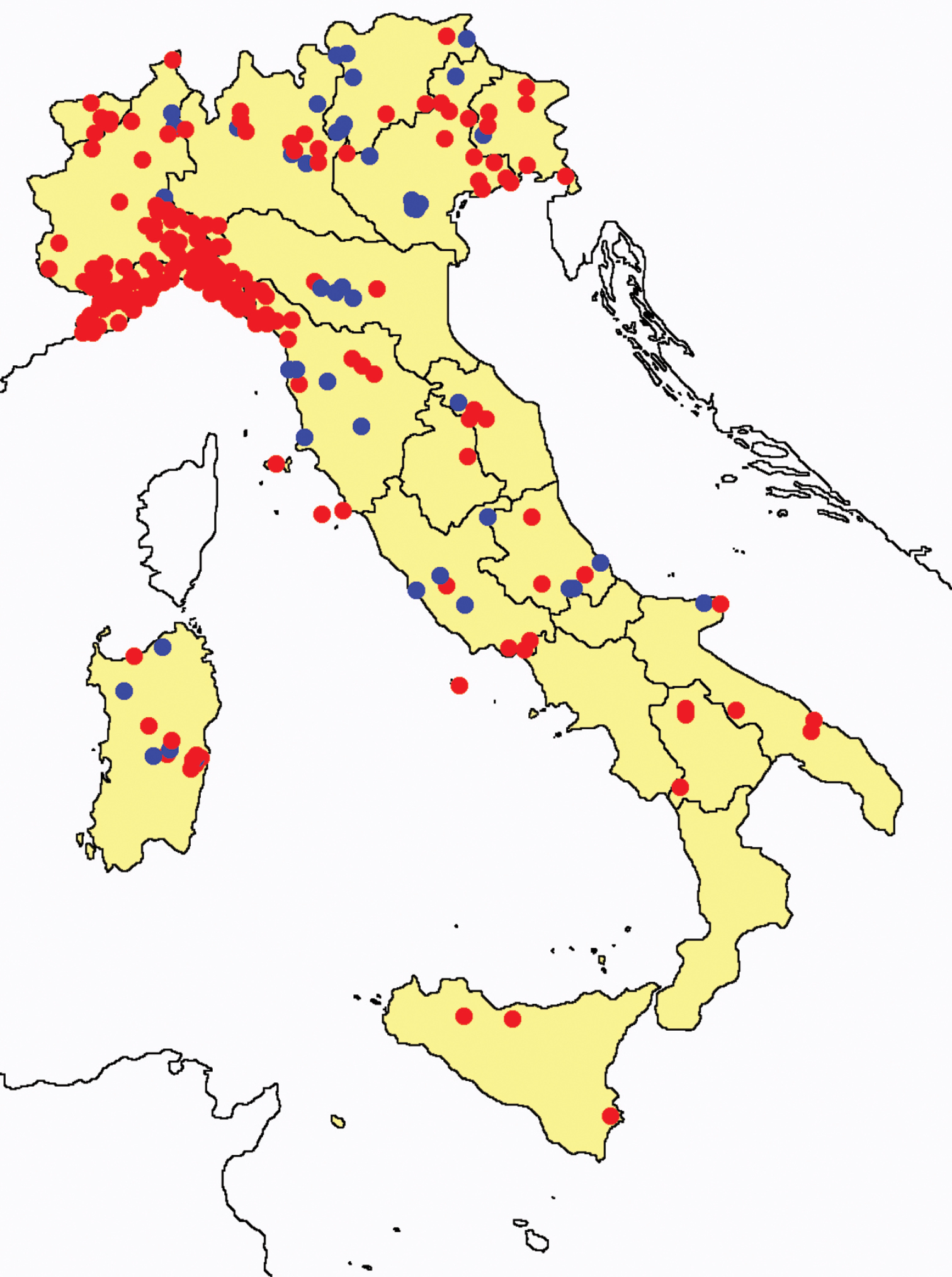
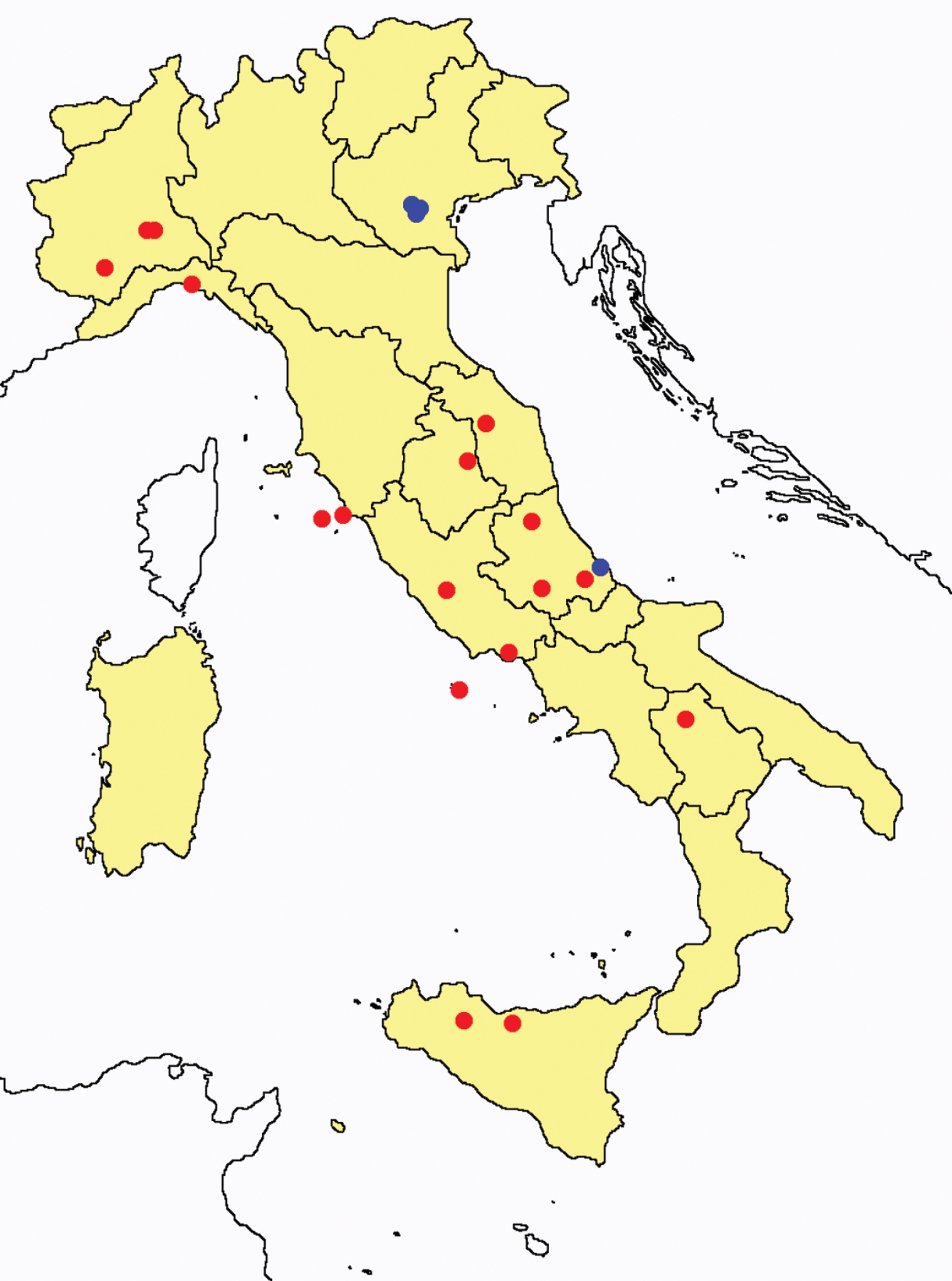
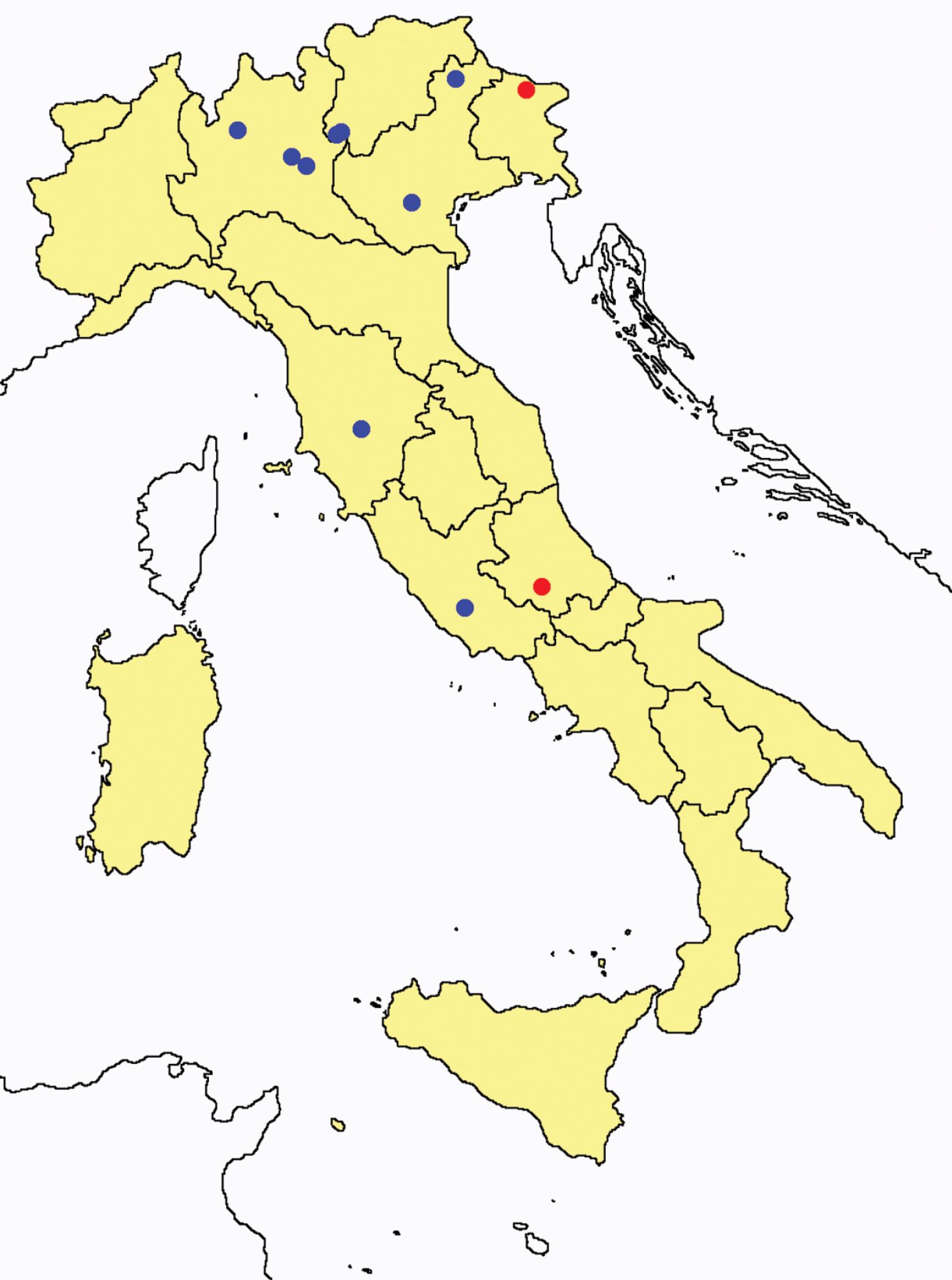
Distribution of the Protura sampling sites in Italy (blue dots: data known only from literature; red dots: data about specimens examined by the authors of this paper).

Species richness in the 20 Italian regions.
According to the analysis made in this paper, we have been able to identify 40 Protura species in Italy, belonging to the families Hesperentomidae (1), Protentomidae (6), Acerentomidae (26) and Eosentomidae (7). At the species level, according to
Chorotypes of the Italian Protura.
| Chorotypes | Species nr |
| Sub-Cosmopolitan | 2 |
| W-Palearctic | 2 |
| Turanic-European-Mediterranean | 4 |
| European-Mediterranean | 5 |
| European | 3 |
| Central-European | 8 |
| S-European | 2 |
| Mediterranean | 4 |
| Known only in Italy | 10 |
Based on the findings, the Italian fauna is mainly composed of species having a European or Mediterranean distribution. With regard to the 10 species known only in Italy, it cannot be said to be endemic due to the poor level of knowledge of this taxon. For the same reason, that given in Table 2 should be considered only a preliminary attempt at classification, which, most likely, is susceptible to changes in the future.
The number of species and genera known in the European Countries (according to
Number of Protura species and genera in the European Countries.
| Country | Species nr | Genera nr |
| Austria | 58 | 10 |
| Balearic islands | 7 | 5 |
| Belgium | 4 | 3 |
| Bosnia and Herzegovina | 16 | 7 |
| Bulgaria | 4 | 2 |
| Corsica | 15 | 7 |
| Croatia | 4 | 2 |
| Czech Republic | 33 | 7 |
| Denmark | 7 | 5 |
| Finland | 3 | 2 |
| France | 38 | 10 |
| Germany | 44 | 10 |
| Greece | 13 | 9 |
| Hungary | 10 | 5 |
| Iceland | 2 | 1 |
| Ireland | 5 | 3 |
| Italy | 40 | 8 |
| Lithuania | 2 | 1 |
| Luxemburg | 30 | 10 |
| Macedonia | 2 | 2 |
| The Netherlands | 1 | 1 |
| Norway | 4 | 1 |
| Poland | 68 | 11 |
| Portugal | 15 | 5 |
| Romania | 10 | 5 |
| Russia | 7 | 4 |
| Serbia | 3 | 2 |
| Slovakia | 38 | 8 |
| Slovenia | 7 | 3 |
| Spain | 23 | 7 |
| Sweden | 12 | 5 |
| Switzerland | 11 | 6 |
| Ukraine | 58 | 12 |
| United Kingdom | 14 | 6 |
It seems rather unlikely that generally poorer (in terms of biodiversity) Countries such as Poland, Ukraine, Austria and Germany have more Protura species than Italy. It’s more likely that this gap is due to a lack of knowledge of the Italian fauna. In support of this hypothesis, a year spent on sampling project in a small cork oak wood in Liguria (NW Italy) led us to identify (
We therefore hope that in the future we will be able to deepen and broaden our research to obtain a more accurate picture of Protura’s ecology and distribution.
The authors wish to thank Dr. Peter J. Schwendinger and Dr. Lionel Monod (Geneva Natural History Museum), Dr. Roberto Poggi (Genoa Natural History Museum) and Dr. Leonardo Latella (Verona Natural History Museum) for giving us the opportunity to study the museum collections. Many thanks to all people that in the last years sent the specimens to our Lab, and particularly to Dr. Giulio Gardini & Stefano Zoia. A special thank to Mrs Marina Zacco, Director of Italo Britannica Society, for kindly revising the manuscript.