





(C) 2011 Cristiano Lopes-Andrade. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Three new species of Falsocis Pic are described: Falsocis aquilonius sp. n. from Panamá, Costa Rica and Colombia, Falsocis egregius sp. n. from a single locality in northern Brazil and Falsocis occultus sp. n. from two localities in southeastern and southern Brazil. New records, comparative notes and an identification key for male and female specimens of Falsocis species are also provided.
Minute tree-fungus beetles, Ciinae, Neotropical, Tenebrionoidea
Falsocis Pic, 1916 is a distinctive but relatively uniform genus of Neotropical Ciidae, recently redescribed by
The aims of this work are to provide new records and comparative notes on Falsocis, descriptions of three additional species of the genus and an updated identification key for the species.
Material and methodsSlide preparations of male terminalia, and methods for photographing slide preparations and specimens, followed mostly
Range, mean and standard deviation are given for measurements and ratios. Measurements of antennomeres were taken from the holotypes. The following abbreviations are used for measurements and ratios: CL, length of the antennal club; EL, elytral length (median length from base of scutellum to elytral apex); EW, greatest elytral width; FL, length of the antennal funicle; GD, greatest depth of the body; PL, pronotal length along midline (including the anterior pronotal plate in males); PW, greatest pronotal width; TL, total length (EL+PL; head not included). The ratio GD/EW was taken as an indication of degree of convexity; TL/EW indicates degree of body elongation. Values for TL and TL/EW of the type series of Falsocis brasiliensis and the unique available male Falsocis opacus are also provided here, since in
The distribution map (Fig. 41) was created using latitude and longitude coordinates estimated by tracking localities in the freeware Google Earth 6.0.3 (
The following acronyms are used in this paper:
ANIC Australian National Insect Collection, CSIRO Entomology (Canberra, Australia)
CMN Canadian Museum of Nature (Ottawa, Ontario, Canada)
CMNH Carnegie Museum of Natural History (Pittsburgh, Pennsylvania, USA)
CNCI Canadian National Collection of Insects (Ottawa, Ontario, Canada)
FMNH Field Museum of Natural History (Chicago, Illinois, USA)
LAPC Cristiano Lopes-Andrade Private Collection (Viçosa, MG, Brazil)
MNHN Muséum National d’Histoire Naturelle (Paris, France)
MTD Museum für Tierkunde Dresden (Dresden, Germany)
Taxonomyurn:lsid:zoobank.org:act:01613AEF-7593-43F1-B83C-CB7F8A13AF21
http://species-id.net/wiki/Falsocis_aquilonius
Figs 1–9, 41Cerro Campana, in the province of Panamá, Panama (8°26'N, 81°17'W).
The specific epithet is from the Latin“aquilonius" (adjective), which means “from the North", in reference to its occurrence at the northernmost locality for Falsocis species.
Epipleura enlarged posteriorly, but just slightly explanate (Figs 2–3), not or barely visible from above, with external margin simple (not crenulate). Pronotum with lateral margins visible for their entire lengths from above. Male abdominal sex patch small with a diameter of one-fifth the length of the first abdominal ventrite.
Male holotype (Figs 1–3), measurements in mm: TL 2.21; PL 0.95; PW 1.11; EL 1.26; EW 1.16; GD 0.89. Ratios: PL/PW 0.86; EL/EW 1.09; EL/PL 1.33; GD/EW 0.77; TL/EW 1.91. Body oblong, strongly convex, mostly yellowish brown; coxae and femora pale yellow; mesoventrite, metaventrite and first abdominal ventrite whitish. Head not visible from above; frontoclypeal ridge slightly raised, bearing two very short, barely pronounced tubercles; disc slightly convex, closely and coarsely punctate, glabrous; in between punctures finely granulate. Eyes coarsely facetted; greatest eye width 0.15mm. Each antenna (left antenna measured; FL 0.18mm; CL 0.18mm; CL/FL 1.00) with length of antennomeres (in mm) as follows: 0.08; 0.06; 0.06; 0.05; 0.03; 0.02; 0.02; 0.05; 0.05; 0.08. Pronotum with single and coarse punctation; punctures very close to each other on disc and near the lateral and posterior margins, but somewhat shallower and separated by a distance of about one puncture-width at the anterior projection; in between punctures finely granulate; vestiture grayish, dual (seen in lateral view under a magnification of 100×), consisting of conspicuous stout erect bristles (~0.05mm) and very small decumbent setae (a bit less than 0.02mm); anterior angles rounded, produced forward; anterior edge explanate and produced forward forming a plate that slightly curves upward and tapers toward a rounded apex; lateral margins slightly explanate and visible for their entire lengths from above, irregularly crenulate. Hindwings fully developed. Scutellum subtriangular, densely covered by stout decumbent bristles (better seen in lateral view); basal width 0.15mm. Sides of elytra parallel at basal three-fourths, broadly rounded posteriorly (seen from above) and converging to a blunt apex; punctation single and confused; punctures coarse and separated by a distance of one puncture-width or less; in between punctures dull; vestiture dual, similar to that of pronotum; lateral and apical margins not visible from above; epipleura tapering from base to middle, then enlarging to apex, with external margin simple (not crenulate). Ventral surface of thorax and abdomen finely granulate, with vestiture of decumbent slender setae. Prosternum flat; prosternal process laminate, three-fourths the length of procoxae. First abdominal ventrite twice as long as the second at midline; setose sex patch circular and margined, located near the posterior margin and with a diameter of one-fifth the length of the ventrite at midline.
Male terminalia in paratypes (Figs 5–6). Eighth sternite (Fig. 5) with posterior margin straight; posterior angles slightly produced forming two small lateral prominences. Basal piece (Fig. 6) nearly one-third the length of tegmen. Tegmen (Fig. 6) with anterior portion subtriangular; lateral margins straight, slightly diverging to apex; posterior portion with a median V-shaped emargination of about one-third the length of tegmen, forming two lateral lobes. Penis (Fig. 6) as long as tegmen, subcylindrical; lateral margins straight for half of their lengths then slightly curved inwards to a narrowly rounded apex.
Females (Fig. 4). Similar to males, but frontoclypeal ridge straight, barely sinuous, with rounded angles. Anterior pronotal margin broadly rounded. Abdomen devoid of sex patch.
Males, measurements in mm (n = 6, including the holotype): TL 2.00–2.84 (2.45 ± 0.32); PL 0.74–1.32 (1.02 ± 0.20); PW 1.00–1.37 (1.16 ± 0.12); EL 1.21–1.47 (1.37 ± 0.11); EW 1.05–1.42 (1.23 ± 0.12); GD 0.84–1.16 (0.97 ± 0.13). Ratios: PL/PW 0.74–0.96 (0.87 ± 0.09); EL/EW 1.04–1.17 (1.12 ± 0.05); EL/PL 1.12–1.64 (1.37 ± 0.18); GD/EW 0.74–0.88 (0.79 ± 0.05); TL/EW 1.90–2.17 (1.99 ± 0.10). The anterior plate (projected anterior margin) of pronotum in males varies from very small (Fig. 7), medium size (Fig. 8, similar to that of holotype) to extremely projected (Fig. 9); but the apex of this plate is rounded or blunt and devoid of tufts. Females, measurements in mm (n = 5): TL 2.21–2.53 (2.38 ± 0.14); PL 0.79–0.89 (0.83 ± 0.04); PW 1.11–1.37 (1.24 ± 0.10); EL 1.37–1.63 (1.47 ± 0.11); EW 1.16–1.47 (1.33 ± 0.13); GD 0.95–1.16 (1.05 ± 0.08). Ratios: PL/PW 0.60–0.81 (0.67 ± 0.08); EL/EW 1.04–1.27 (1.12 ± 0.09); EL/PL 1.63–1.94 (1.78 ± 0.15); GD/EW 0.74–0.95 (0.80 ± 0.09); TL/EW 1.68–2.14 (1.81 ± 0.19).
Holotype. (ANIC) Panama: \R.P.: Panama Cerro Campana Feb. 22, 1975 Lawrence, Erwin \ Phellinus sp. \ J.F. Lawrence Lot [printed] 3799 [handwritten] \ Falsocis aquilonius Lopes-Andrade & Lawrence HOLOTYPUS [printed on red paper]\. Paratypes. Panama: 1 male and 1 female (ANIC), same data as holotype; 3 males (1 ANIC, 2 LAPC), 4 females (2 ANIC, 1 LAPC) \2.5mi S.W.Rincon Puntarenas, Costa Rica, Mar. 1–7, ’67 \ J.F. Lawrence Lot. [printed] 2175 [handwritten] \ R. Andrews Collector\. Costa Rica. 1 male (ANIC) \La Lola, C.R. III-8–1958 M.J. Stelzer [handwritten] \ MS 58.4 [handwritten]\; 1 female (ANIC) \Puerto Viejo COSTA RICA VIII-4–65 \ J.F. Lawrence Lot [printed] 1611 [handwritten]\. Colombia. 1 male (Marseul Collection, MNHN) \Dupont Colomb [18]41 [handwritten in circular green paper] \ MARSEUL [handwritten in rectangular small green paper] \ [circular blue paper, without information] \ FALSOCIS SPP. [handwritten] det. J.F. Lawrence 19 [printed]\. All paratypes distinguished labeled \Falsocis aquilonius Lopes-Andrade & Lawrence PARATYPUS [printed on yellow paper]\.
One specimen collected in an unidentified Phellinus. This is the only Falsocis species known from Central America, but it also occurs at the northernmost South America (Fig. 41). Recent field collections in Colombia were unsuccessful in recapturing the species, so the country record is based only on a single old male without precise locality data. In Falsocis opacus, the epipleura is much more enlarged, easily visible from above, with external margin crenulate; the male eighth sternite is similar, but basal piece is strongly developed and is almost half the length of aedeagus; tegmen is elongate, four times as long as wide. In the remaining Falsocis, the epipleura is narrower near the apex.

Falsocis aquilonius Lopes-Andrade & Lawrence, sp. n., male holotype 1–3, shown in the same scale, female paratype from Puntarenas, Costa Rica 4 and slide preparation of male terminalia of a paratype 5–6. 1 Dorsal view 2 Lateral view 3 Ventral view 4 Dorsal view 5 Eighth sternite 6 Aedeagus (left) and gross outline of aedeagus (right) showing tegmen (continuous line), penis (dashed line) and basal piece (dotted line).

Falsocis aquilonius Lopes-Andrade & Lawrence, sp. n., male paratypes, dorsal view, shown in the same scale. 7 Low male from Cerro Campana (Panama) 8 Medium male from La Lola (Costa Rica) 9 High male from Puntarenas (Costa Rica).
urn:lsid:zoobank.org:act:E6BFE936-A896-449F-8D6A-ADE8528F8728
http://species-id.net/wiki/Falsocis_egregius
Figs 10–16, 41Santarém, in the state of Pará, northern Brazil (2°26'S, 54°42'W).
The specific epithet is from the Latin “egregius” (adjective), which means “singular”, “extraordinary”, in a reference to the outstanding head morphology of males.
Head of male with sides of vertex produced laterally to form a pair of triangular plates, each with acute apex (Fig. 13, arrows); frontoclypeal horns wide at base and tapering to apex, slightly arcuate. Pronotum with anterior angles broadly rounded, barely produced forward; males with a row of long setae along the apex of the anterior projection but not forming distinct tufts (Figs 10, 12, arrows).
Male holotype (Figs 10-13), measurements in mm: TL 2.85; PL 1.25; PW 1.50; EL 1.55; EW 1.55; GD 1.35. Ratios: PL/PW 0.83; EL/EW 1.00; EL/PL 1.24; GD/EW 0.87; TL/EW 1.84. Body oblong, strongly convex, mostly light yellowish brown; mouthparts and apices of femora reddish brown; ventral vestiture consisting of very fine decumbent setae. Head strongly developed, the anterior and lateral margins easily visible from above; dorsum concave, slightly tumid on disc; punctation relatively fine, shallow, sparse; in between punctures finely granulate; frontoclypeal ridge explanate, each side produced upward forming a conspicuous horn that is slightly arcuate, bearing two small tubercles between them; each side of vertex explanate and produced laterally forming a conspicuous triangular plate with an acute apex (Fig. 13, arrows). Eyes coarsely facetted; greatest width 0.18mm. Each antenna (left antenna measured; FL 0.22mm; CL 0.23mm; CL/FL 1.05) with length of antennomeres (in mm) as follows: 0.12; 0.07; 0.08; 0.04; 0.04; 0.03; 0.03; 0.06; 0.06; 0.11. Pronotum with shallow, coarse, single punctation; punctures separated by a distance about a puncture-width; in between punctures smooth at disc and finely granulate at the anterior projection; vestiture indistinctly dual, consisting of conspicuous yellowish stout erect bristles (~0.05mm) and very fine decumbent setae (<0.02mm); anterior angles broadly rounded, just slightly produced forward; anterior margin (beyond anterior angles) explanate, produced forward forming a plate that slightly curves downward and narrows toward a slightly arcuate apex with a row of very long slender setae (>0.35mm; Figs 10, 12, arrows). Scutellum subtriangular; punctation finer than those of pronotum and elytra; vestiture consisting of stout decumbent bristles (better seen in lateral view); basal width 0.21mm. Hindwings fully developed. Elytra subparallel at basal two-thirds, with posterolateral angles broadly rounded (as seen from above) and then converging to a blunt apex; punctation single and confused, a bit sparser than that of pronotum; in between punctures smooth; vestiture indistinctly dual, consisting of conspicuous stout erect bristles (~0.07mm) and very minute decumbent setae (<0.02mm); lateral and apical margins not visible from above; epipleura tapering from base to apex. Surface of the ventral thoracic and abdominal sclerites granulate, somewhat rugose. Prosternum flat; prosternal process laminate, almost half the length of procoxae. First abdominal ventrite more than twice as long as the second at midline; setose sex patch circular and margined, located at the middle of the ventrite and with a diameter of one-third the length of the ventrite at midline.
Male terminalia in paratypes (Figs 15–16). Eighth sternite (Fig. 15) with posterior margin curved inward; angles not produced. Basal piece (Fig. 16) almost half the length of tegmen. Tegmen (Fig. 16) with anterior portion broadly rounded; lateral margins almost straight; posterior portion bearing a deep V-shaped emargination reaching the middle of the structure and forming two lateral lobes. Penis (Fig. 16) subcylindrical, lateral margins straight for most of their lengths and a bit expanded to a rounded apex; the penis is turned and lays laterally in the slide preparation.
Female (Fig. 14).Similar to males, but frontoclypeal ridge straight, barely sinuous, with rounded angles. Anterior pronotal margin broadly rounded. Abdomen devoid of sex patch.
Males, measurements in mm (n = 5, including the holotype): TL 2.75–2.95 (2.87 ± 0.08); PL 1.20–1.35 (1.26 ± 0.05); PW 1.50–1.55 (1.52 ± 0.03); EL 1.50–1.65 (1.56 ± 0.05); EW 1.55–1.65 (1.57 ± 0.04); GD 1.30–1.40 (1.35 ± 0.04). Ratios: PL/PW 0.80–0.87 (0.83 ± 0.03); EL/EW 0.97–1.00 (0.99 ± 0.01); EL/PL 1.15–1.32 (1.24 ± 0.06); GD/EW 0.84–0.87 (0.86 ± 0.02); TL/EW 1.77–1.87 (1.83 ± 0.05).
Female, measurements in mm (n = 1): TL 2.50; PL 0.95; PW 1.45; EL 1.55; EW 1.50; GD 1.05. Ratios: PL/PW 0.66; EL/EW 1.03; EL/PL 1.63; GD/EW 0.70; TL/EW 1.67.
Holotype. (CMNH) Brazil: \Santarem Brazil Acc. No.2966 \ CM \ Falsocis egregius Lopes-Andrade & Lawrence HOLOTYPUS [printed on red paper]\. Paratypes. Brazil: 4 males (1 ANIC; 2 CMNH; 1 LAPC, dissected) and 1 female (CMNH) \Santarem Brazil Acc. No.2966 \ CM\, same data as holotype. All paratypes distinguished labeled \Falsocis egregius Lopes-Andrade & Lawrence PARATYPUS [printed on yellow paper]\.
The species is currently known from a single collection in Santarém, northern Brazil (Fig. 41). Only one female was available for examination. The series is very small, but species sufficiently distinct to allow description. Differs from Falsocis aquilonius sp. n. and Falsocis opacus in having the epipleura narrow posteriorly. Falsocis occultus sp. n. has deeper and closer pronotal punctation. Very similar to Falsocis brasiliensis, but differing mainly in features mentioned in the diagnosis and in the conspicuous basal piece of male terminalia. In Falsocis brasiliensis, the eighth sternite has the anterior margin only slightly curved inward; the basal piece was not observed in the available slide preparations and is possibly membranous (

Falsocis egregius Lopes-Andrade & Lawrence, sp. n., male holotype 10–13 and female paratype 14 shown in the same scale, and slide preparation of male terminalia of a paratype 15–16. 10 Dorsal view (pronotal tuft of long setae, arrow) 11 Lateral view 12 Ventral view (pronotal tuft of long setae, arrow) 13 Head and pronotum view from above (triangular plate behind eyes, arrows) 14 Female 15 Eighth sternite 16 Aedeagus (left) and gross outline of aedeagus (right) showing tegmen (continuous line), penis (dashed line) and basal piece (dotted line).
urn:lsid:zoobank.org:act:EF30CDE6-0A8A-444A-9A89-0ECB68E9CB17
http://species-id.net/wiki/Falsocis_occultus
Figs 17–25, 41Linhares, in the state of Espírito Santo, southeastern Brazil (19°23'S, 40°04'W).
The specific epithet is from the Latin “occultus” (adjective), which means “hidden”, in reference to the fact that the population from the type locality stayed hidden among fungi forgotten in a field-base for near four years.
Pronotum with anterolateral angles not produced forward; lateral margins not visible from above; male with anterior projection ending in an acute apex (Figs 19, 23-25, arrows) bordered by medium-size bristles.
Male holotype (Figs 17–19), measurements in mm: TL 3.55; PL 1.70; PW 1.75; EL 1.80; EW 1.75; GD 1.40. Ratios: PL/PW 0.97; EL/EW 1.03; EL/PL 1.06; GD/EW 0.80; TL/EW 2.03. Body oblong, strongly convex, mostly dark brown; mouthparts, antennae and tarsi dark yellowish brown; femora and tibiae dark reddish brown. Head concealed by the anterior pronotal projection (seen from above) except for its anterolateral angles; dorsum concave with disc slightly tumid; punctation coarse, shallow; in between punctures finely granulate; frontoclypeal ridge explanate and produced forming a broad acute triangular plate at each anterior angle, with two small tubercles between them. Eyes coarsely facetted; greatest eye width 0.21mm. Each antenna (left antenna measured; FL 0.30mm; CL 0.28mm; CL/FL 0.93) with length of antennomeres (in mm) as follows: 0.15; 0.09; 0.08; 0.08; 0.06; 0.05; 0.03; 0.08; 0.08; 0.12. Pronotum with single, coarse, relatively deep punctation; punctures very close to each other, separated by a distance of one puncture-width or less; in between punctures smooth but not shining; vestiture yellowish, indistinctly dual (seen under a magnification of 100×), consisting of stout erect bristles (~0.05mm) and small decumbent setae (~0.03mm), both better seen in lateral view; anterior angles not produced forward; anterior margin (beyond anterior angles) explanate, strongly produced forward forming a plate that slightly curves downward and narrows toward an acute apex (Fig. 19, arrow) ornamented by a row of increasingly stout bristles in either side (Figs 17-19); lateral margins slightly crenulate, not visible from above, bearing a row of stout bristles. Scutellum subtriangular, its margins indistinct so that it seems to be contiguous with elytra; punctation conspicuous but slightly finer than those of pronotum and elytra; vestiture consisting of stout decumbent bristles (better seen in lateral view); basal width 0.20mm. Hindwings fully developed. Elytra parallel at basal three-fourths, posteriorly broadly rounded (as seen from above) and converging to a blunt apex; punctation single and confused, slightly finer than that of pronotum, consisting of relatively deep punctures separated by a distance near a puncture-width; in between punctures smooth, dull; vestiture distinctly dual, the erect bristles about 0.1mm long and the decumbent setae about 0.03mm long; lateral and apical margins not visible from above; epipleura tapering from base to the basal one-sixth, then continuing as a narrow line to the apex. Ventral surfaces of thorax and abdomen finely granulate; vestiture consisting of slender decumbent setae. Prosternum flat; prosternal process laminate, two-thirds the length of the procoxae. First abdominal ventrite more than twice as long as the second at midline; setose sex patch suboval and margined, located at the middle of the ventrite and with a diameter of one-third the length of the ventrite at midline.
Male terminalia in paratypes (Figs 21–22). Eighth sternite (Fig. 21) with posterior margin almost straight; angles not produced. Basal piece (Fig. 22) nearly one-third the length of tegmen. Tegmen (Fig. 22) with anterior portion mostly rounded but apex acute; lateral margins slightly sinuous and diverging; posterior portion bearing a deep V-shaped emargination reaching the middle of the structure, forming two lateral lobes. Penis (Fig. 22) subcylindrical; lateral margins subparallel for most of their lengths; apical third subtriangular, membranous.
Females (Fig. 20). Similar to males, but frontoclypeal ridge straight, barely sinuous, with rounded angles. Anterior pronotal margin broadly rounded. Abdomen devoid of sex patch.
Males, measurements in mm (n = 15, including the holotype): TL 2.11–3.55 (2.84 ± 0.42); PL 0.84–1.70 (1.28 ± 0.25); PW 1.16–1.75 (1.48 ± 0.17); EL 1.25–1.80 (1.53 ± 0.19); EW 1.21–1.84 (1.52 ± 0.17); GD 0.95–1.47 (1.22 ± 0.16). Ratios: PL/PW 0.73–0.97 (0.86 ± 0.07); EL/EW 0.86–1.07 (1.01 ± 0.05); EL/PL 1.00–1.50 (1.22 ± 0.15); GD/EW 0.72–0.93 (0.80 ± 0.05); TL/EW 1.73–2.03 (1.86 ± 0.09). In the unique specimen from Nova Teutonia, a teneral male, pronotal and elytral bristles are larger (0.08mm and 0.15mm, respectively) than in specimens from Linhares. In small males, the anterior pronotal plate is barely projected (Fig. 23). However, all males have an acute pronotal apex (Figs 19, 23–25, arrows). Females, measurements in mm (n = 15): TL 2.30–3.15 (2.63 ± 0.27); PL 0.80–1.16 (0.97 ± 0.11); PW 1.05–1.75 (1.42 ± 0.19); EL 1.40–2.00 (1.64 ± 0.18); EW 1.10–1.85 (1.50 ± 0.19); GD 1.05–1.50 (1.23 ± 0.15). Ratios: PL/PW 0.62–0.81 (0.69 ± 0.05); EL/EW 1.00–1.41 (1.11 ± 0.10); EL/PL 1.50–2.00 (1.71 ± 0.15); GD/EW 0.79–0.95 (0.83 ± 0.04); TL/EW 1.64–2.23 (1.77 ± 0.14).
Holotype. (LAPC) Brazil: \BRASIL: ES Linhares 11–21.x.2004 P.C. Grossi leg. \ Falsocis occultus Lopes-Andrade & Lawrence HOLOTYPUS [printed on red paper]\. Paratypes. Brazil: 56 (27 males, 1 dissected, and 19 females, LAPC; 5 males and 5 females, ANIC), same data as holotype; 1 male (FMNH) \Nova Teutonia, Sta. Catharina, BRAZ. 300–500m alt. Fritz Plaumann leg [printed] XI:1940 [handwritten] \ [circular red paper, without information] \ Falsocis sp. 115 \. All paratypes distinguished labeled \Falsocis occultus Lopes-Andrade & Lawrence PARATYPUS [printed on yellow paper]\.
26 specimens, gender not determined (2 CNCI for molecular analysis, 4 LAPC in absolute alcohol, 20 MTD), same data as holotype.
Known from two localities, in southeastern and southern Brazil (Fig. 41). The specimens from Linhares (in the state of Espírito Santo, southeastern Brazil) were collected in Hexagonia papyracea Berk.(Polyporaceae) and bred in the laboratory until December 2009 in the original basidiomes, without addition of either water or nutrients. In male Falsocis brasiliensis and Falsocis egregius sp. n., the apex of the pronotal projection bears a row of setae that are comparatively longer (Figs 10–12, 26–29, 33–34). Differs from Falsocis aquilonius sp. n. and Falsocis opacus in having the epipleura narrow posteriorly.

Falsocis occultus Lopes-Andrade & Lawrence, sp. n., male holotype 17–19 and female paratype 20 shown in the same scale, and slide preparation of male terminalia of a paratype 21–22. 17 Dorsal view 18 Lateral view 19 Ventral view (acute pronotal apex, arrow) 20 Dorsal view 21 Eighth sternite 22 Aedeagus (left) and gross outline of aedeagus (right) showing tegmen (continuous line), penis (dashed line) and basal piece (dotted line).

Falsocis occultus Lopes-Andrade & Lawrence, sp. n., male paratypes 23–24 shown in the same scale, dorsal view (acute pronotal apex, arrows). 23 Low male from Linhares (in the state of Espírito Santo, southeastern Brazil) 24 Medium male from Linhares 25 The unique specimen known from Nova Teutônia (in the state of Santa Catarina, southern Brazil).
http://species-id.net/wiki/Falsocis_brasiliensis
Figs 26–34, 41Brazil: 1 female (LAPC) \BRASIL: ES Santa Maria de Jetibá 08.xii.2003 leg. Furieri & Nunes\; 3 males (1 LAPC, in absolute alcohol; 2 CNCI, for molecular analysis) and 1 female (LAPC, in absolute alcohol) \BRASIL: MG Viçosa “Belvedere” 03.ii.2011 leg. L.S. Araújo, C.A. Carvalho\; 2 males and 1 female (LAPC) \BRASIL: MG Viçosa Mata da Biologia, ponto 37 16.iv.2010; leg. T. Mariani & C. Lopes-Andrade; ex Hymenochaete luteobadia\; 2 males (LAPC) \BRASIL: MG Viçosa Mata da Biologia, ponto 048 20.iv.2011; C. A. Carvalho & C. Lopes-Andrade; ex Hymenochaete luteobadia\.
The specimens from Viçosa (in the state of Minas Gerais, southeastern Brazil; Figs 26–31) were all collected in Hymenochaete luteobadia (Fr.) Höhn. & Litsch. (Hymenochaetacea). This fungus was previously misidentified as Phellinus sp. (

Falsocis brasiliensis Lopes-Andrade, 2007, specimens from Viçosa (in the state of Minas Gerais, southeastern Brazil), all shown in the same scale. 26 Dorsal view (produced angles, large arrows; two tufts of long setae, small arrows) 27 Lateral view 28 Ventral view (produced angles, large arrows; two tufts of long setae, small arrows) 29 Head and pronotum view from above 30 Low male 31 Female.

Falsocis brasiliensis Lopes-Andrade, 2007, dorsal view, shown in the same scale. 32 The unique female specimen from Santa Maria de Jetibá, in the state of Espírito Santo, southeastern Brazil 33 Male paratype from Venda Nova do Imigrante, Espírito Santo (two tufts of long setae, arrows) 34 Male paratype from Jussari (in the state of Bahia).
The subspecies Falsocis opacus flavus Pic, 1922 was based on a single female collected in Bas-Maroni (French Guyana, near Cayenne), and it is merely a teneral specimen of Falsocis opacus (J. F. Lawrence pers. obs.).The type of Falsocis opacus (MNHN) is 2.10mm long, rather than 3mm as mentioned in the description (
1 female (MNHN) Peru: \Mission de Sarayacu (Riv. Oucayale) [Ucayali] [handwritten in green paper] \ CO 47 [handwritten in a circular yellowish paper] \ MUSEUM PARIS PÉROU PAMPAS [sic] DEL SACRAMENTO DE CASTELNAU 1847 [written in light brown paper] \ [circular blue paper, without information]\.
The specimen was collected at Sarayacu Mission (6°47'S, 75°07'W, Fig. 41) during the French Scientific Expedition to the Pampa del Sacramento, Peru, in 1847. It resembles females of Falsocis brasiliensis, Falsocis egregius sp. n. and Falsocis occultus sp. n. in having the epipleura narrow posteriorly. However, the shallow punctation of pronotum resembles mostly that of the former two species. Female Falsocis brasiliensis from Jussari (Brazil) are similar in color, but body is more elongate. It is very large, with a length of near 3mm. Only females of Falsocis occultus sp. n. reach such a length. A subquadrate body is also observed in a few Falsocis brasiliensis, but the largest females of this species are only 2.4mm long. The very transverse pronotum, twice as long as wide, does not occur in any other known Falsocis species. This female from Peru possibly belongs to another new species, but description is not possible without examining a male. Measurements (in mm) and ratios are as follows: TL 3.05; PL 1.00; PW 2.00; EL 2.00; EW 2.11; GD 1.68; PL/PW 0.50; EL/EW 0.95; EL/PL 2.00; GD/EW 0.80; TL/EW 1.45.

Falsocis opacus Pic, 1916, from Altamira, in the state of Pará, northern Brazil 35–37; arrows pointing out the explanate outer apical margins of epipleura and Falsocis sp., female from Peru 38–40, all shown in the same scale. 35, 38 Dorsal view 36, 39 Lateral view 37, 40 Ventral view.
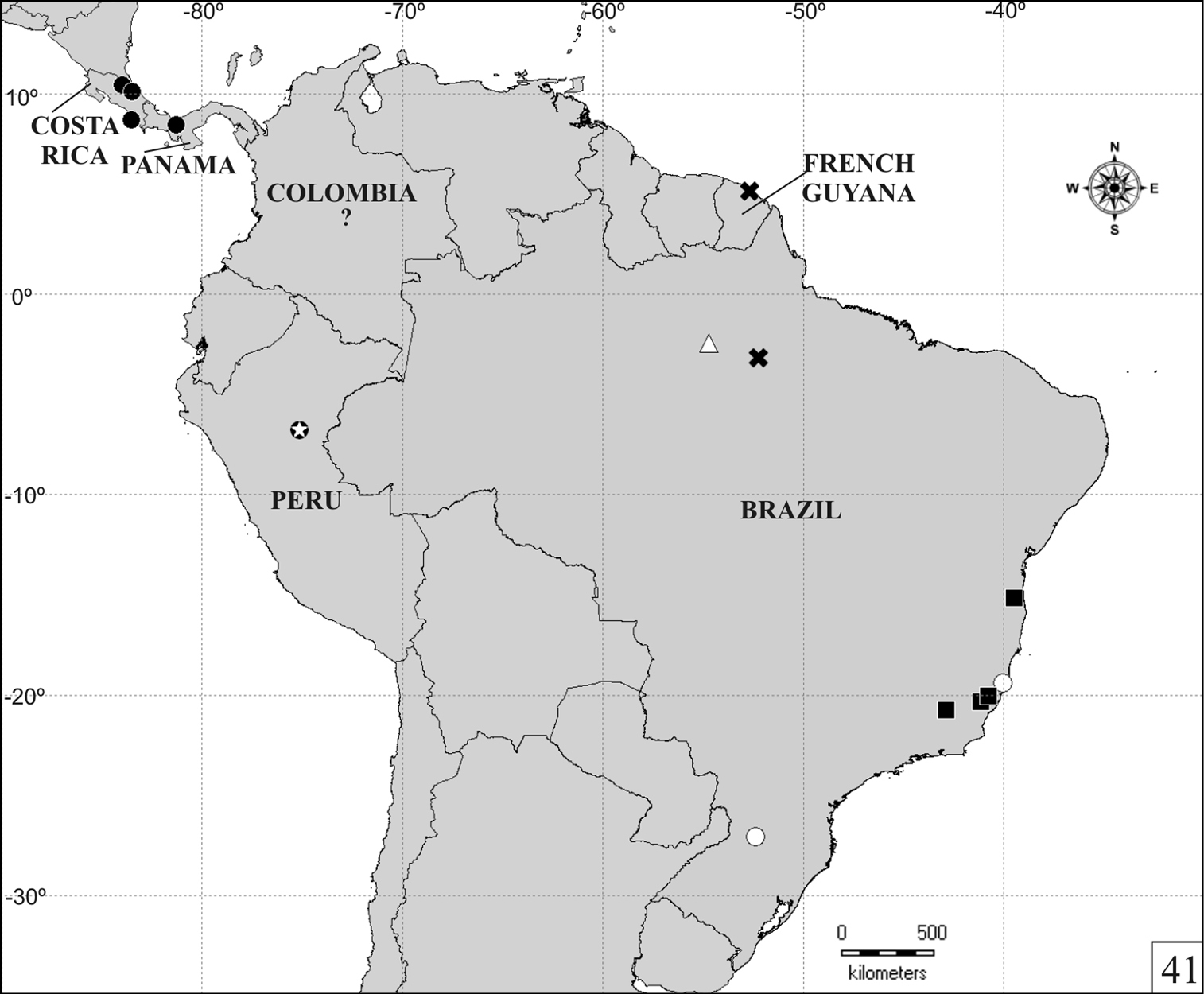
Distribution map for Falsocis Pic species. Falsocis aquilonius sp. n. (full circle), Falsocis brasiliensis Lopes-Andrade (full squares), Falsocis egregius sp. n. (open triangle), Falsocis occultus sp. n. (open circles), Falsocis opacus Pic (x) and Falsocis sp. (open star inside full circle). The question mark indicates an imprecise location of Falsocis aquilonius sp. n. in Colombia.
Identification key to the species of Falsocis Pic
| 1 | Epipleura enlarged posteriorly, forming a slightly or strongly explanate posterolateral margin in elytra, conspicuously or barely visible from above | 2 |
| 1’ | Epipleura narrowing to apex, forming a narrow, not explanate, posterolateral margin in elytra, not visible from above | 3 |
| 2(1) | Pronotum with lateral margins visible from above. Posterior elytral margin (outer margin of epipleura) simple (not crenulate), barely visible from above. Known from Panamá, Costa Rica and Colombia | Falsocis aquilonius sp. n. |
| 2’ | Pronotum with lateral margins not visible from above. Posterior elytral margin (outer margin of epipleura) crenulate, easily visible from below and above (Figs 35–37, arrows). Known from Brazil (Altamira, in the state of Pará) and French Guyana | Falsocis opacus Pic, 1916 |
| 3(1’) | Pronotal punctation deep and coarse, with punctures usually separated by half a puncture-width or less at disc. Anterior plate of pronotum in males devoid of tufts of very long setae and acute at apex (Figs 19, 23–25) | Falsocis occultus sp. n. |
| 3’ | Pronotal punctation shallow and fine, with punctures usually separated by about a puncture-width at disc. Anterior plate of pronotum in males with slender setae usually as long as to twice as long (or more) than an eye-width, with apex straight, barely emarginated or slightly rounded | 4 |
| 4(3’) | Anterior pronotal angles distinctly produced forward and somewhat acute (Figs 26, 28, large arrows). Males with head devoid of conspicuous projection behind each eye (Fig. 29); long setae of anterior pronotal plate organized in two tufts (Figs 26, 28, 33, small arrows) | Falsocis brasiliensis Lopes-Andrade, 2007 |
| 4’ | Anterior pronotal angles barely produced forward and rounded (Figs 10-12). Males with head bearing a conspicuous triangular plate, projected outward, behind each eye (Fig. 13, arrows); long setae of anterior pronotal plate organized as a row, not forming tufts (Figs 10, 12, arrows) | Falsocis egregius sp. n. |
The phylogenetic position of Falsocis is uncertain. In the available phylogenetic analysis of the family, based on molecular data, Falsocis brasiliensis was not part of any defined clade (
Species of Falsocis are not frequently collected and all are allopatric. They are known only from forests or forest remnants and usually one or few specimens are found together in the field. The greatest series is that for Falsocis occultus sp. n., which was reared in laboratory. Accurate data on host fungi are scarce. Available information on Falsocis brasiliensis suggests the species is monophagous (
CLA thanks Paschoal C. Grossi, Caio Antunes de Carvalho, Lucimar Soares de Araújo and Karina S. Furieri for collecting Falsocis species, and Tatiana B. Gibertoni for identifying the host fungi of F. occultus sp. n. and F. brasiliensis. Field collections in Brazil were registered and licensed by Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio, Ministério do Meio Ambiente). Images of male terminalia were made at Laboratório de Acarologia (Departamento de Entomologia, Universidade Federal de Viçosa/UFV). Figures 4, 7 and 9 were provided by Caio Antunes de Carvalho, and figures 15, 16 and 41 by Vivian E. Sandoval Gómez. CLA was supported by Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG: Edital PPP 21/2008 2008, APQ-00049-09; Edital PPM 03/2010, PPM-00017-10), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq: PROTAX 52/2010 nº 562229/2010-8) and the Graduate Program in Entomology of UFV. We also thank current and past curators of ANIC, CMN, CMNH, FMNH and MNHN for the loan of ciids.