





(C) 2012 Mykola M. Kovblyuk. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Eight species of Haplodrassus are recorded from Crimea: Haplodrassus bohemicus Miller & Buchar, 1977; Haplodrassus dalmatensis (L. Koch, 1866); Haplodrassus isaevi Ponomarev & Tsvetkov, 2006; Haplodrassus minor (O. P.-Cambridge, 1879); Haplodrassus kulczynskii Lohmander, 1942; Haplodrassus pseudosignifer Marusik, Hippa & Koponen, 1996; Haplodrassus signifer (C.L. Koch, 1839) and Haplodrassus umbratilis (L. Koch, 1866). The occurrence of Haplodrassus cognatus (Westring, 1861) in Crimea has not been confirmed. Haplodrassus bohemicus is a new species record for the Crimean fauna. Haplodrassus pseudosignifer is a new species record for Crimea and Ukraine as a whole, with Crimea as the westernmost point of its distribution range. Haplodrassus invalidus is recorded for the first time for the fauna of Azerbaijan, Caucasus and the former Soviet Union. Azerbaijan is the easternmost point of its known distribution range. All Crimean Haplodrassus species have only one peak of activity of adult specimens during the year. In Crimea we found syntopically two closely related species Haplodrassus dalmatensis and Haplodrassus isaevi in two localities (Sudak Distr., 10 km W Sudak, Mezhdurechie Vill., steppe; and Feodosiya Distr., Karadag Nature Reserve, steppes). These species differ in their phenology. The reproductive period of Haplodrassus dalmatensis isin May-July, and that of Haplodrassus isaevi occurs is in October-December. These phenological differences probably represent an additional mechanism of reproductive isolation between the two species. Diagnostic drawings are provided for all mentioned species as well as for Haplodrassus deserticola Schmidt & Krause, 1996 and Haplodrassus pugnans (Simon, 1880).
Haplodrassus, Crimea, Ukraine, Azerbaijan, Kazakhstan, redescriptions, fauna, phenology
Haplodrassus Chamberlin, 1922 with 65 species is a relatively large gnaphosid genus distributed in the Holarctic and India (
In the former Soviet Union 21 species have been recorded to date (
Microphotographs were made using an SEM Jeol JSM-5200 in the Zoological Museum, University of Turku, Finland. Photographs were taken in dishes of different sizes with paraffin at the bottom. Specimens were photographed using an Olympus Camedia E-520 camera attached to an Olympus SZX16 stereomicroscope at the Zoological Museum, University of Turku. Digital images were montaged using “CombineZM” image stacking software.
Coloration was described from specimens preserved in an ethanol/water solution. Leg segments were measured after detaching them from the cephalothorax. All measurements are in mm: minimum-maximum; a figure in brackets represents the average. Illustrations were made using both reflecting- and transmitted-light microscopes. All scale bars equal 0.1 mm.
The morphological terminology follows
All specimens treated in this study are held in the following collections: CP – personal collection of A.V. Ponomarev (Rostov-on-Don, Russia); EMZ – personal collection of E.M. Zhukovets (Minsk, Belarus); ISEA – Siberian Zoological Museum, Institute for Systematics and Ecology of Animals, Novosibirsk, Russia, G.N. Azarkina; TNU – Zoology Department, V.I. Vernadsky Taurida National University, Simferopol, Ukraine (M.M. Kovblyuk); ZMMU – Zoological Museum of the Moscow State University, Moscow, Russia, K.G. Mikhailov; ZMT – Zoological Museum, University of Turku, Finland (S. Koponen); YMC – Yuri M. Marusik’s temporary collection in the Zoological Museum, University of Turku, Finland.
The following abbreviations are used in the text: AM, AL, PM, PL – anterior median, anterior lateral, posterior median and posterior lateral eyes; RTA – retrolateral tibial apophysis.
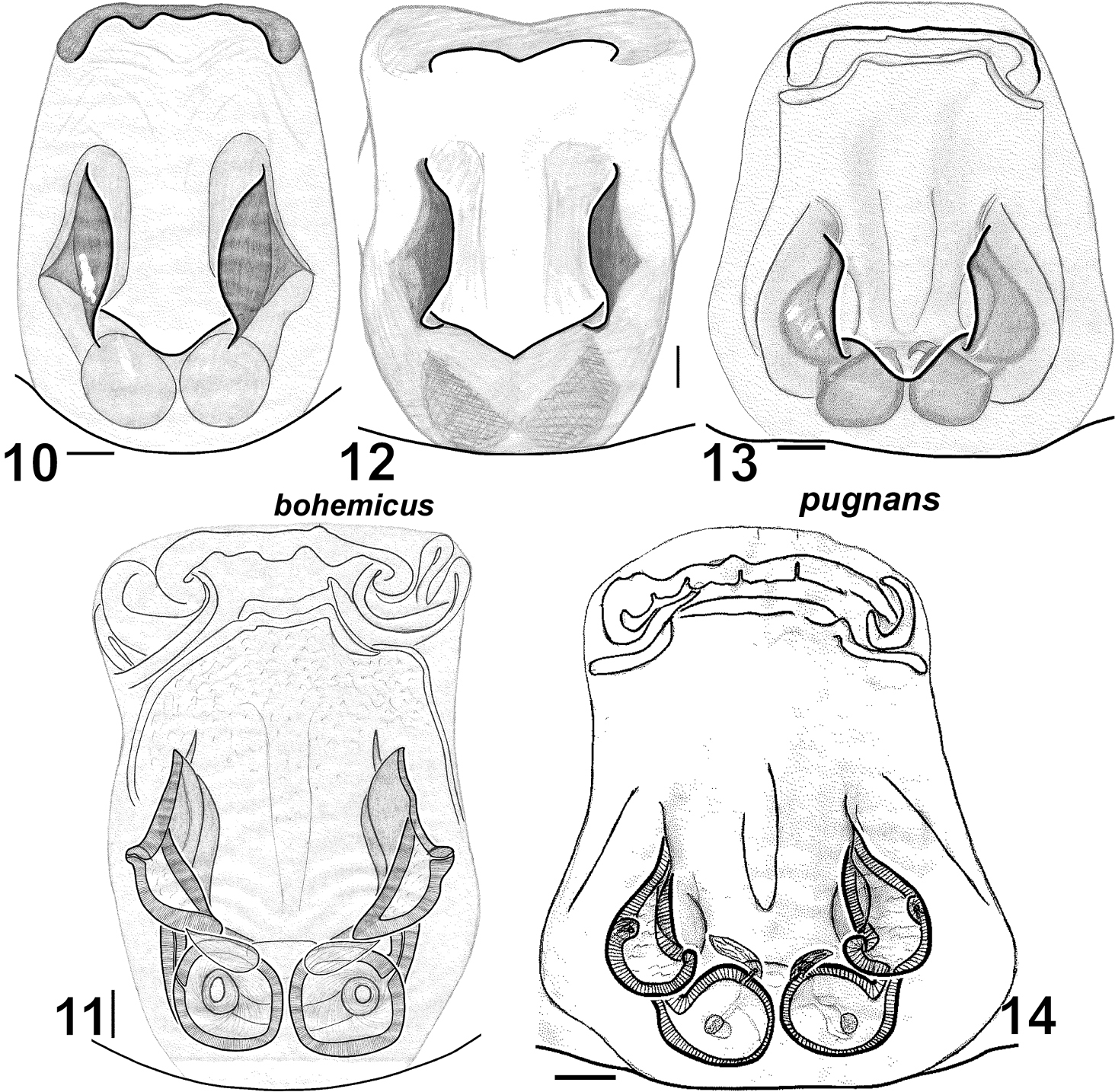
Key to Haplodrassus species found in Crimea Males| 1 | Terminal apophysis toothed (Fig. 37) | Haplodrassus kulczynskii |
| – | Terminal apophysis with 1–2 or without teeth | 2 |
| 2 | Total length ≤ 4 mm, carapace length ≤ 2 mm | Haplodrassus minor |
| – | Total length more than 4 mm, carapace longer than 2 mm | 3 |
| 3 | Terminal apophysis with plate-like bulge (Fig. 81) | Haplodrassus umbratilis |
| – | Terminal apophysis without plate-like bulge | 4 |
| 4 | Terminal apophysis with 1–2 teeth (Figs 23, 26) | 5 |
| – | Terminal apophysis without teeth (Figs 2, 5, 69, 72) | 6 |
| 5 | Terminal apophysis with 1 tooth, embolus without tooth (Fig. 26), metatarsus I with 2 ventral spines | Haplodrassus isaevi |
| – | Terminal apophysis with 2 teeth, embolus with tooth (Fig. 23), metatarsus I without ventral spines | Haplodrassus dalmatensis |
| 6 | RTA with dorsal “step”-like keel (Figs 1, 4), embolus without tooth (Figs 2, 5) | Haplodrassus bohemicus |
| – | RTA without “step”-like keel (Figs 68, 71), embolus with tooth (Figs 69, 72) | 7 |
| 7 | Terminal apophysis short (Fig. 69), length/width ratio ca 2 | Haplodrassus pseudosignifer |
| – | Terminal apophysis long (Fig. 72), length/width ratio ca 3 | Haplodrassus signifer |
| 1 | Epigynal pockets with long protrusion directed anteriorly (Fig. 38) | Haplodrassus kulczynskii |
| – | Epigynal pockets without long protrusion | 2 |
| 2 | Body length ≤ 5 mm, carapace ≤ 2 mm | Haplodrassus minor |
| – | Body longer than 5 mm, carapace longer than 2 mm | 2 |
| 3 | Epigynal fovea constricted anteriorly (Figs 28, 30) | 4 |
| – | Epigynal fovea not constricted anteriorly | 5 |
| 4 | Epigynal fovea with septum, fovea wider than spermathecae (Fig. 28), metatarsus IV with 3 retrolateral spines | Haplodrassus dalmatensis |
| – | Epigynal fovea without septum, fovea narrower than spermathecae (Fig. 30), metatarsus IV with 4–5 retrolateral spines | Haplodrassus isaevi |
| 5 | Spermathecae oval (Figs 82–83) | Haplodrassus umbratilis |
| – | Spermathecae globular | 6 |
| 6 | Fovea without longitudinal groove (Figs 10, 12) | Haplodrassus bohemicus |
| – | Fovea with long longitudinal groove (Figs 75–78) | 7 |
| 7 | Fovea rectangular (longer than wide) (Fig. 75) | Haplodrassus pseudosignifer |
| – | Fovea square-shaped (length subequal to width) (Fig. 77) | Haplodrassus signifer |
Drassus hiemalis Emerton, 1909.
Male palp with large terminal apophysis, thick embolus, hooked median apophysis and RTA flattened, often shifted dorsally. Epigyne with thick sclerotized lateral pockets and with one anterior hood. Posterior median eyes close together, separated by their radius or less (
In terms of habitus and coloration Haplodrassus resembles only two other genera that occur in Crimea and the eastern Mediterranean: Parasyrisca Schenkel, 1963 and, to a lesser extent, Drassodes Westring, 1851. However, Haplodrassus is easily distinguished by having a large terminal apophysis (absent in the other genera), a flat retrolateral tibial apophysis widened dorsally (conical or flat and tapering in Drassodes and Parasyrisca), a broad embolus (cylindrical or hidden in the other genera), and the presence of heavily sclerotized lateral epigynal pockets (absent in Drassodes and Parasyrisca).
Holarctic and India (
http://species-id.net/wiki/Haplodrassus_bohemicus
Figs 1–6, 10–12UKRAINE, CRIMEA: Saky Distr.: 12 ♂♂, 2 ♀♀ (TNU), near Pribrezhnaya railway station, 30.04.-24.06.2000, M.M. Kovblyuk.
UKRAINE. Nikolaev Area: 6 ♂♂ (TNU), Pervomaysky Distr., Kuripchane Vil., 5.05.–8.06.2006, N.Yu. Polchaninova. Kherson Area: 2 ♂♂, 2 ♀♀ (TNU), Henichesk Distr., Arabatskaya strelka, 4 km S Henichesk Town, 23.05.–10.06.2011, N.A. Stasyuk. Donetsk Area: 4 ♂♂ (TNU), Pershotravnevy Distr., Belosaraiskaya Kosa, 11–22.06.2001, E.V. Prokopenko; 2 ♂♂ (TNU), Novoazovsk Distr., Khomutovo Vil., “Khomutovskaya Steppe”, N47°16', E38°10', 15–20.06.2004, N.Yu. Polchaninova; 1 ♂ (TNU), Slavyansky Distr., Svyatogorsk Town, N49°02', E37°39', 8–30.06.2004, N.Yu. Polchaninova. RUSSIA. Rostov Area: 7 ♂♂, 1 ♀ (TNU), Ust’-Donetsk Distr., near Razdorskaya Vil., «Pukhlyakovskye sklony», 10.05.–28.06.2004, A.V. Ponomarev.
Haplodrassus pugnans (Simon, 1880): RUSSIA, Magadan Area: 1 ♂, 3 ♀♀ (ISEA, БИ-930), Ten’kynskyi Distr., env. Sibit-Tyellakh, Aborigen Field Station, Betula, 12–22.06.1983, A.V. Avershin.
Haplodrassus bohemicus is most similar to Haplodrassus pugnans (Simon, 1880), Haplodrassus signifer (C.L. Koch, 1839) and Haplodrassus pseudosignifer Marusiket al. 1996, but differs: 1) by the shape of RTA having a “step”-like dorsal margin (RTA dorsal margin without “step” in Haplodrassus pugnans, Haplodrassus signifer and Haplodrassus pseudosignifer); 2) by the apically directed embolus lacking a tooth (retrolaterally directed embolus with a tooth in Haplodrassus pugnans, Haplodrassus signifer and Haplodrassus pseudosignifer); 3) by the outlines of epigynal sclerites and relative proportions of the epigynal fovea (cf. Figs 10, 12 and 13, 75, 78).
Male measurements (n = 5). Total length 5.8–7.5 (6.7); carapace 2.7–3.2 (3.0) long, 2.0–2.6 (2.3) wide. Diameters of eyes and distances between them: AM 0.10–0.15 (0.12), AL 0.10–0.16 (0.13), PM 0.15–0.22 (0.19), PL 0.10–0.14 (0.12), AM-AM 0.09–0.12 (0.10), AM-AL 0.03–0.04 (0.04), PM-PM 0.03–0.04 (0.03), PM-PL 0.12–0.16 (0.14), AM-PM 0.12–0.18 (0.15), AL-PL 0.10–0.16 (0.14). Distances between anterior eyes and margin of clypeus: AM-clypeus 0.18–0.22 (0.20), AL-clypeus 0.10–0.18 (0.15).
Length of leg segments (male):
| Leg | Femur | Patella | Tibia | Metatarsus | Tarsus | Total |
|---|---|---|---|---|---|---|
| I | 1.9–2.2 (2.1) | 1.1–1.4 (1.2) | 1.6–1.7 (1.6) | 1.2–1.4 (1.3) | 0.9–1.0 (1.0) | 6.8–7.6 (7.2) |
| II | 1.6–1.9 (1.8) | 1.0–1.2 (1.1) | 1.2–1.4 (1.3) | 1.1–1.2 (1.2) | 0.9–1.0 (1.0) | 5.8–6.8 (6.3) |
| III | 1.5–1.8 (1.6) | 0.8–1.0 (0.9) | 0.9–1.0 (1.0) | 1.2–1.4 (1.3) | 0.8–1.0 (0.9) | 5.2–6.0 (5.6) |
| IV | 2.1–2.3 (2.2) | 1.0–1.2 (1.1) | 1.6–1.7 (1.6) | 1.8–2.0 (1.9) | 1.0–1.2 (1.1) | 7.5–8.3 (8.0) |
Length of palp segments: femur 1.0–1.2 (1.1), patella 0.4–0.5 (0.5), tibia 0.3–0.5 (0.4), tarsus 1.0–1.2 (1.0). Cheliceral teeth: anterior – 2 (little; proximal tooth connected with keel of cheliceral groove), posterior – 2. Abdomen 2.9–4.0 (3.6) long, 1.8–2.2 (2.0) wide. Scutum is absent. Basal segment of anterior (inferior) spinnerets 0.5–0.6 (0.6) long. Coloration light brown, as in most Haplodrassus species.
Palp as in Figs 1–6. RTA with a “step” (St) on the promargin, terminal apophysis almost straight, without distinct ridge, embolus slightly twisted and lacks a tooth.
Female measurements (n = 2). Total length 7.8–8.0; carapace 2.7–3.5 long, 2.0–2.7 wide. Abdomen 4.3–5.1 long, 2.7–3.1 wide. Coloration light brown, as in most Haplodrassus species.
Epigyne as in Figs 10–12. Lateral pockets long, sub-parallel, spermathecae globular. Fovea of epigyne without narrow longitudinal groove.
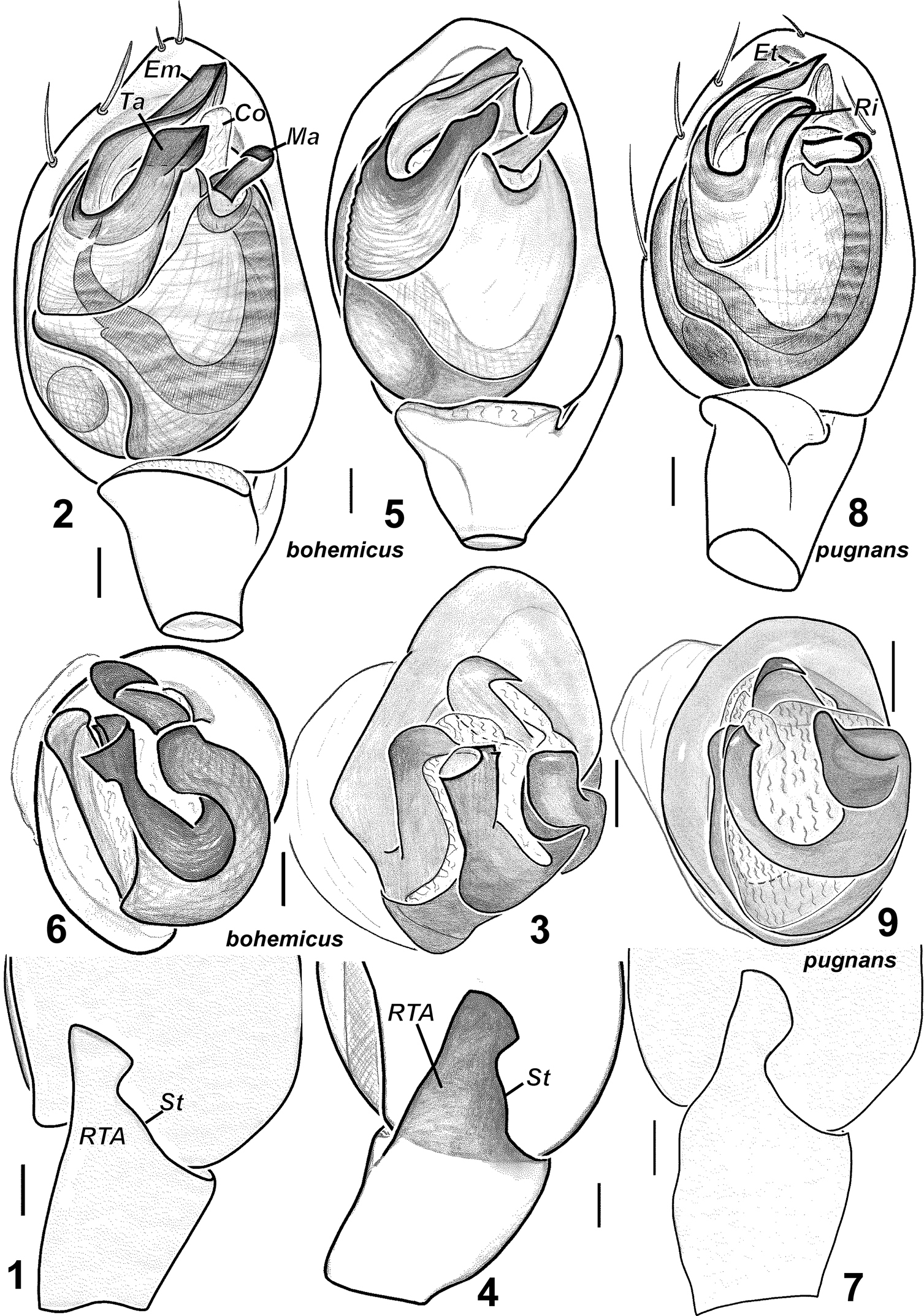
Males of Haplodrassus bohemicus (1–3 from Rostov Area, 4–6 from Crimea) and Haplodrassus pugnans (7–9 from Magadan Area): 1, 4, 7 RTA, retrolateral view 2, 5, 8 palp, ventral view 3, 6, 9 bulbus, apical view. Abbreviations: Coconductor; Emembolus; Ettooth of embolus; Mamedian apophysis; Ri ridgeof terminal apophysis; RTA retrolateral tibial apophysis; St“step”-like keel of RTA; Taterminal apophysis.
Females of Haplodrassus bohemicus (10–11 from Rostov Area, 12 from Crimea) and Haplodrassus pugnans (13–14 from Magadan Area): 10, 12–13 epigyne, ventral view 11, 14 epigyne, dorsal view.
Czech Republic, Macedonia, Greece, Ukraine (Nikolaev, Kherson, Donetsk Areas, Crimea), Russia (Rostov and Stavropol Areas, Kalmykya, Dagestan) (
Haplodrassus bohemicus is a new species record for the Crimean fauna.
Meadows, steppes and sand dunes.
♂♀ – V-VI. In Crimea the peak activity of adults occurs in May.
http://species-id.net/wiki/Haplodrassus_cognatus
Figs 15–16The earlier record of Haplodrassus cognatus from Crimea was based on specimens of unknown sex and number from Karadag Nature Reserve (
UKRAINE. Donetsk Area: 1 ♀ (TNU), Slavyansky Distr., Svyatogorsk Town, N49°02', E37°39', Quercus, 7.06–9.07.2005, N.Yu. Polchaninova.
Haplodrassus silvestris (Blackwall, 1833): UKRAINE. Chernovtsy Area: 1 ♂ (TNU № 2153), Tsetsyno Town, Fagus wood, 23.04–18.05.2009, V.V. Garashchuk & T.O. Auzyak. Kharkiv Area: 1 ♂, 1 ♀ (TNU), Veliko-Burlukskiy Distr., Nesterivka Vill., N49°53', E37°17', 14.06–14.07.2003, N.Yu. Polchaninova. RUSSIA. Belgorod Area: 1 ♀ (TNU), Borisobsky Distr., Borisovka Town, “Les na Vorkle” Reserve, N50°38', E35°58', 5.07.unknown year, N.Yu. Polchaninova.
Haplodrassus cognatus can be distinguished from all other Haplodrassus species by its straight terminal apophysis with a basal tooth in males, and by the shape of the fovea and wide anterior hood (Ah) in females.
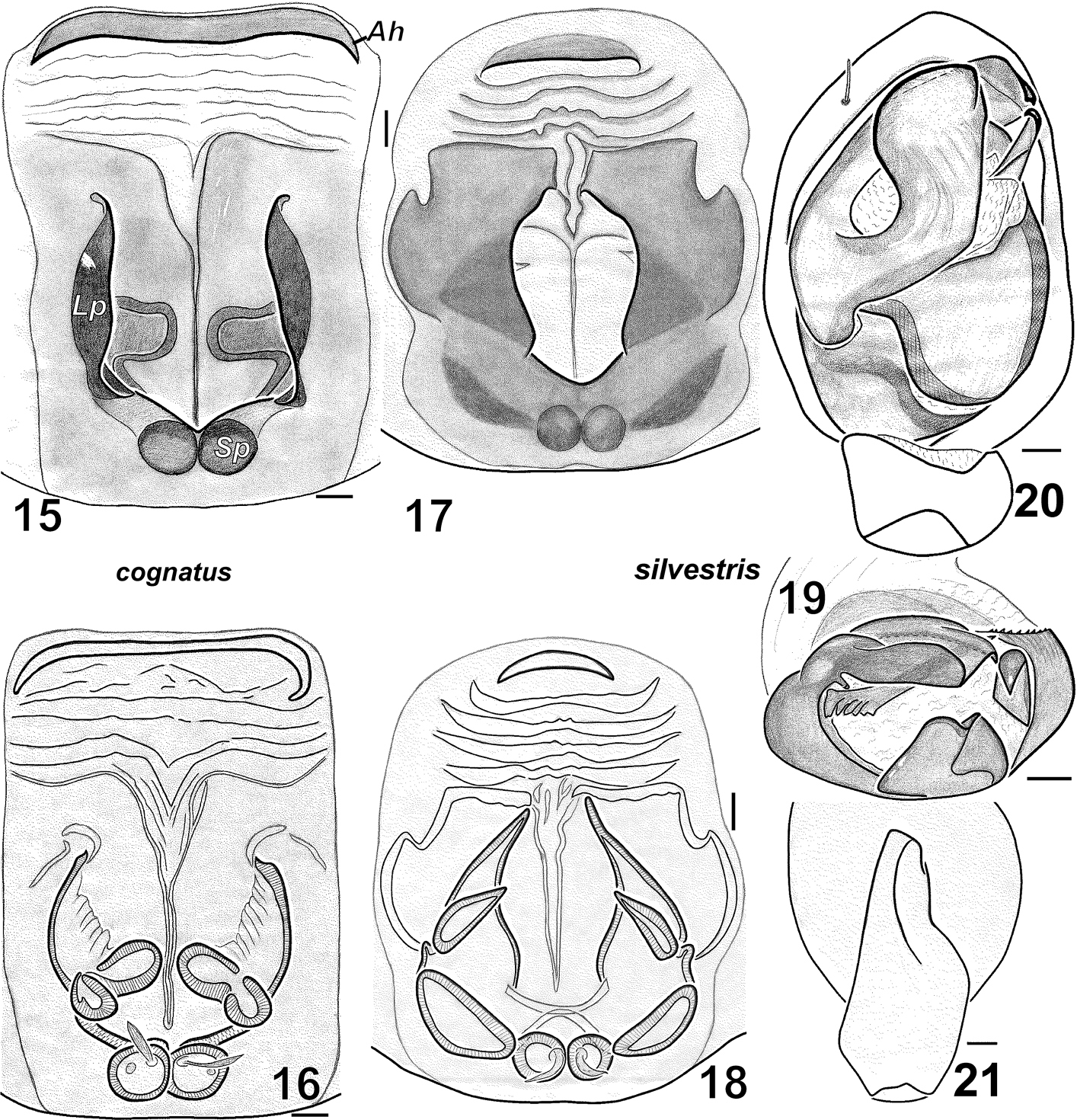
Haplodrassus cognatus (15–16 from Donetsk Area)and Haplodrassus silvestris (17–21 from Kharkiv Area): 15, 17 epigyne, ventral view 16, 18 epigyne, dorsal view 19 bulbus, apical view 20 palp, ventral view 21 RTA, retrolateral view. Abbreviations: Ahanterior hood of epigyne; Lp lateral pocket of epigyne; Sp spermatheca.
It has a trans-Palearctic boreo-nemoral range and occurs from France to Hokkaido, north to north Ural and Tomsk, and south to Greece (
In Central Europe ♂♀ – III-X (
http://species-id.net/wiki/Haplodrassus_dalmatensis
Figs 22–24, 28–29UKRAINE, CRIMEA:Bakhchisaray Distr.: 1 ♂ (TNU), Crimean State Nature Reserve, kordon Asport, 29.06.2001, M.M. Kovblyuk. Feodosiya Distr.: 4 ♂♂, 4 ♀♀ (TNU), Karadag Nature Reserve, 25.05.2003–21.11.2008, M.M. Kovblyuk, O.V. Kukushkin, A.A. Nadolny. Saky Distr.: 5 ♂♂, 7 ♀♀ (TNU), near Pribrezhnaya railway station, 19.05.–3.07.2000, M.M. Kovblyuk. Sevastopol Distr.: 2 ♂ (EMZ), Khersones, 29.05.1996 & 19.06.1998, M.M. Kovblyuk. Simferopol Distr.: 1 ♂, 1 ♀ (EMZ), near Simferopol water reservoir, 30.05.1996, M.M. Kovblyuk; 1 ♀ (TNU), near Fersmanovo Vill., ~ 250 m, 23.06.–16.07.2000, M.M. Kovblyuk; 1 ♀ (TNU), Chatyr-Dagh, Orlinoe canyon, 10–25.06.2000, M.M. Kovblyuk; 3 ♂♂, 1 ♀ (TNU), near Skvortsovo Vill., 19.05.–10.07.2002, M.M. Kovblyuk; 1 ♀ (TNU), Krasnolesye Vill., 10.07.2002, Ya.I. Ibragimova. Sudak Distr.: 2 ♂♂, 2 ♀♀ (TNU), 10 km W Sudak, Mezhdurechie Vill., 23.05–24.06.2010, M.K. Yusufova.
UKRAINE. Kherson Area: 1 ♂ (TNU), Henichesk Distr., Arabatskaya strelka, 4 km S Henichesk Town, 1–10.06.2010, N.A. Stasyuk; 1 ♀ (TNU), Arabatskaya strelka, 7 km S Henichesk Town, 6.07.2010, N.A. Stasyuk.
Haplodrassus dalmatensis can be easily distinguished from all other congeners by the shape of the terminal apophysis with two tooth-like apical processes and by the strong tooth on the embolus in males, and also by the shape of the epigynal fovea with a peculiar medial septum and converging lateral pockets in females.
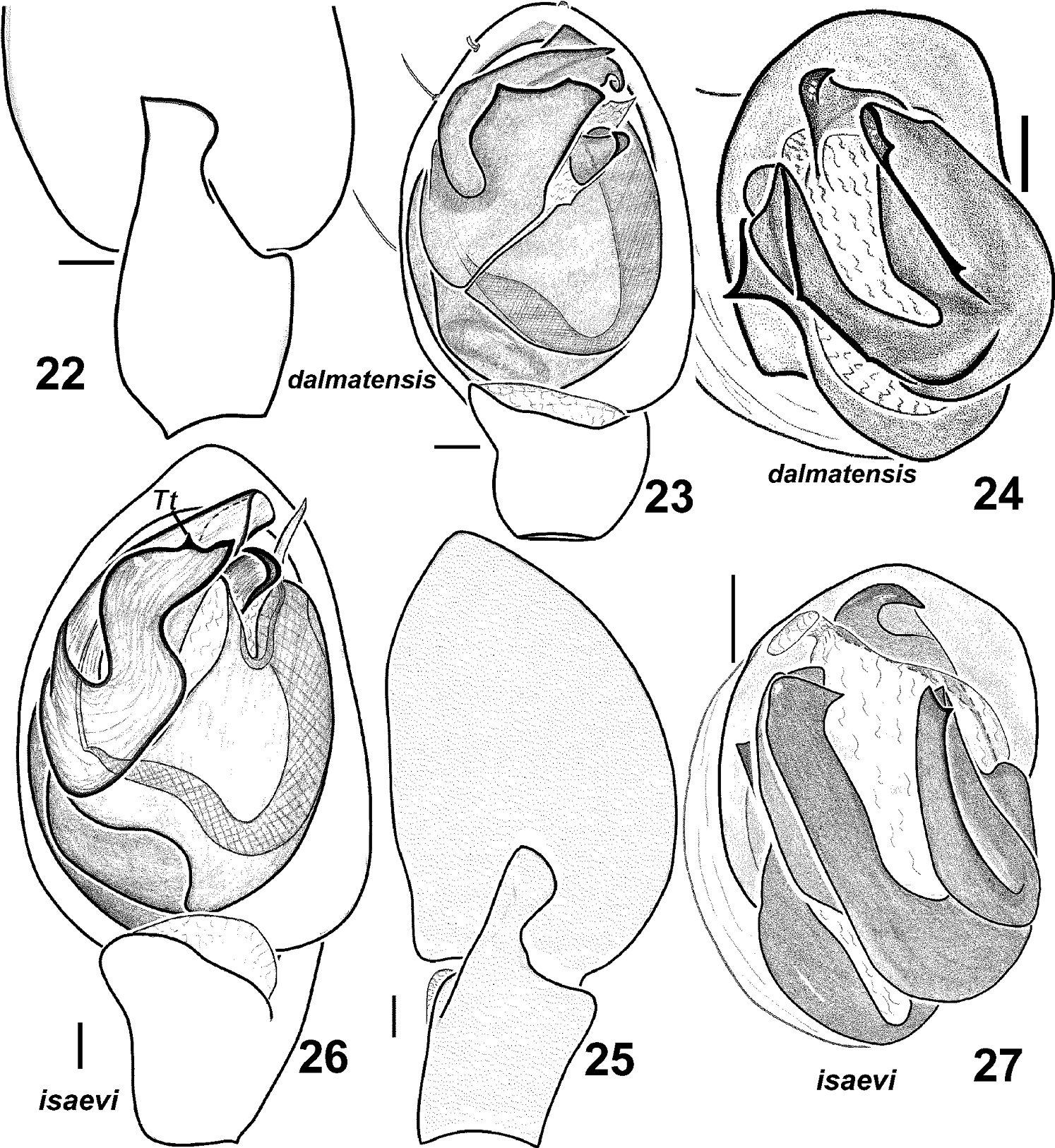
Males of Haplodrassus dalmatensis (22–24 from Crimea) and Haplodrassus isaevi (25–26 paratype from Rostov Area, 27 from Crimea): 22, 25 RTA, retrolateral view 23, 26 palp, ventral view 24, 27 bulbus, apical view. Abbreviations: Tt tooth-like process of terminal apophysis.
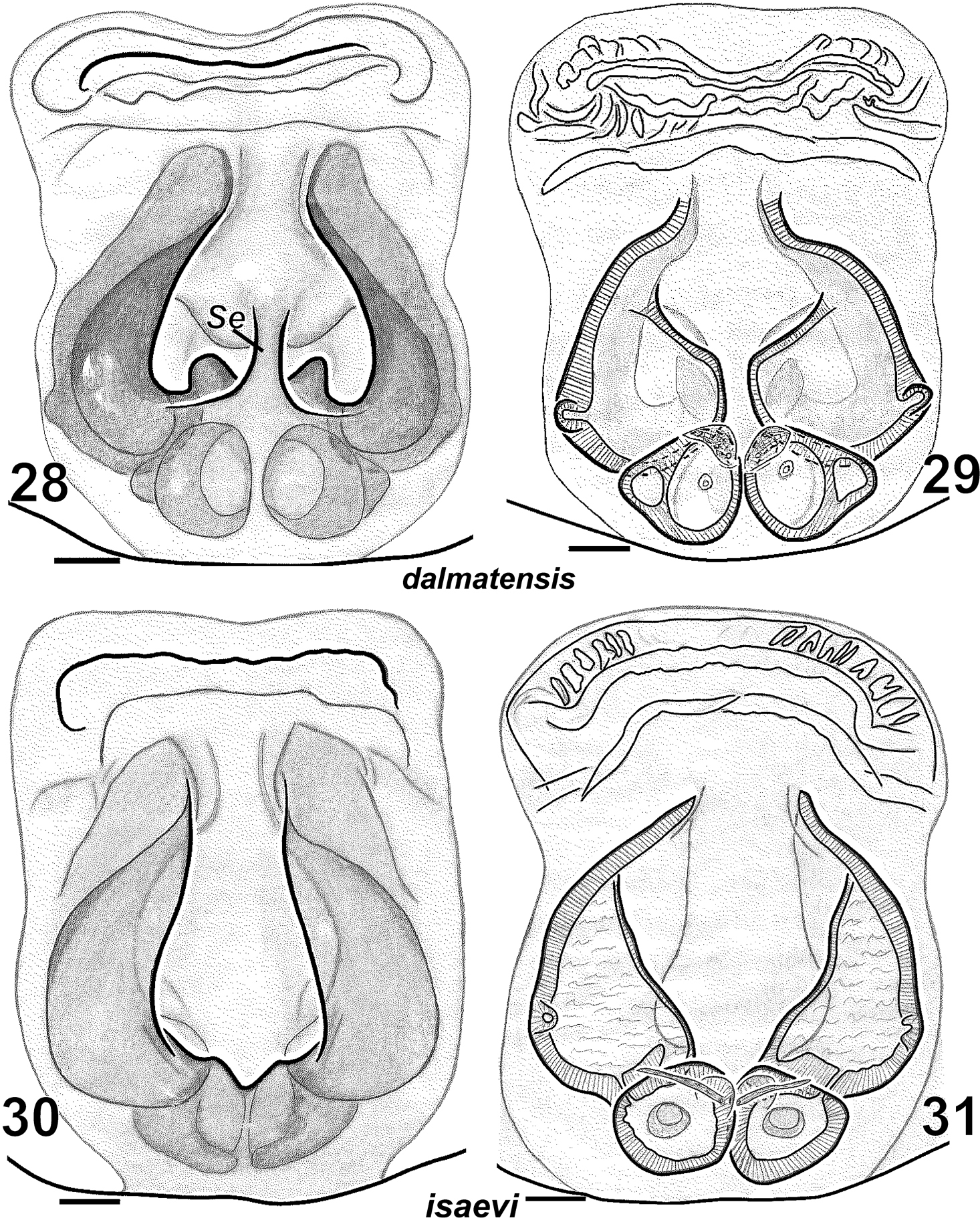
Females of Haplodrassus dalmatensis (28–29 from Crimea) and Haplodrassus isaevi (30–31 from Crimea): 28, 30 epigyne, ventral view 29, 31 epigyne, dorsal view. Abbreviations: Seseptum.
West and Central Palaearctic: North Africa, Europe, South Urals, Caucasus, Anatolia, Near East, Kazakhstan, Western Turkmenistan and mountains of South Siberia (
Juniper forests, forest strips (=shelterbelts), grasslands, steppes, meadows, salt marshes.
In Crimea ♂♀ – V-VI, ♀♀ – VII, XI-XII, the peak of activity in adults occurs in June. In Britain, the peak is in June (
http://species-id.net/wiki/Haplodrassus_invalidus
Figs 32–34AZERBAIJAN. 1 ♂ (TNU), Gobustan, Beyuk-Dash, 17.05.2001, E.F. Huseynov.
Males of Haplodrassus invalidus can be distinguished from all other Haplodrassus species by the peculiar thin embolus with an inner spur-like process, and also the peculiar shape of the RTA, which is not indented and has a claw-like tip (Figs 32–34).
Well described by
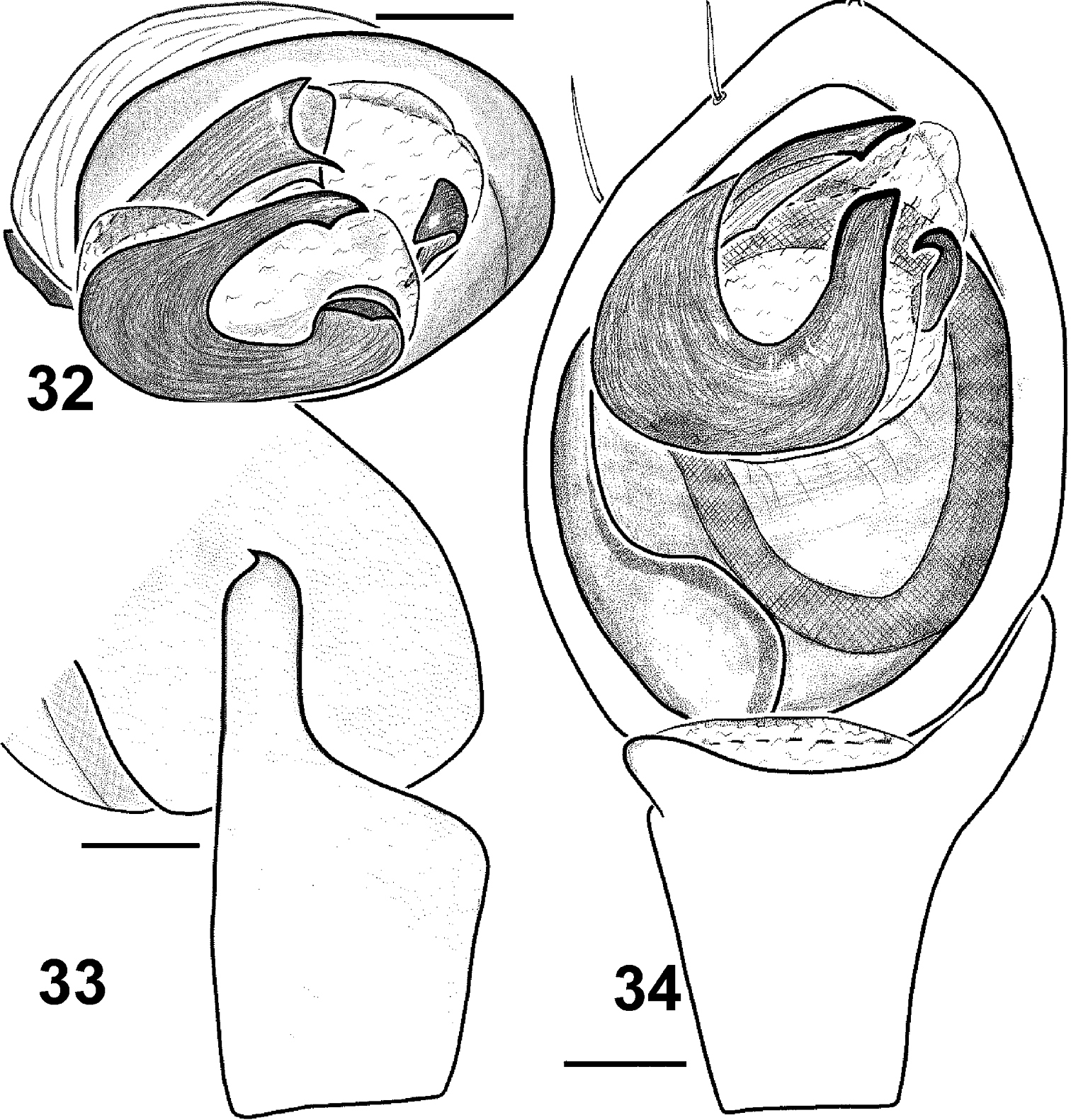
Males of Haplodrassus invalidus from Azerbaijan: 32 bulbus, apical view 33 RTA, retrolateral view 34 palp, ventral view.
Spain, Corsica, Italy (mainland and Sicily), Turkey, Israel and Azerbaijan (
Haplodrassus invalidus is a new species record for the fauna of Azerbaijan, Caucasus and the former Soviet Union. Azerbaijan is the easternmost point of the known distribution range.
http://species-id.net/wiki/Haplodrassus_isaevi
Figs 25–27, 30–31RUSSIA, ROSTOV AREA: 3 ♂♂ paratypes (TNU from CP 18.24.8), Orlovskyi Distr., Rostov Reserve, 6.10.2002, A.V. Ponomarev.
UKRAINE, CRIMEA: Sudak Distr.: 2 ♂♂, 3 ♀♀ (TNU), 10 km W Sudak, Mezhdurechie Vill., 3.10.–7.11.2010, A.K. Yusufova. Feodosiya Distr.: 21 ♂♂, 10 ♀♀ (TNU), Karadag Nature Reserve, 28.05.2003–19.12.2008, M.M. Kovblyuk, O.V. Kukushkin.
Haplodrassus isaevi is most similar to Haplodrassus dalmatensis but differs by the shape of the terminal apophysis, which has only one tooth-like process (two tooth-like apical processes in Haplodrassus dalmatensis), in lacking a tooth on the embolus (embolic tooth present in Haplodrassus dalmatensis), and also by the proportions of the epigyne. Differences also occur in the spination of certain leg segments: male metatarsus I with two ventral spines in Haplodrassus isaevi, but without spines in Haplodrassus dalmatensis; female metatarsus IV with 4–5 retrolateral spines in Haplodrassus isaevi, but with 3 spines in Haplodrassus dalmatensis.
Males (n =5) and females (n = 5). Measurements (♂ / ♀): total length 5.4–7.2 (6.4) / 5.5–7.4 (6.3); carapace 2.3–2.9 (2.7) / 2.4–3.0 (2.7) long, 1.9–2.4 (2.2) / 1.9–2.2 (2.0) wide; abdomen 3.1–4.3 (3.7) / 3.0–4.4 (3.7) long, 1.7–2.3 (2.0) / 1.8–2.8 (2.3) wide.
Length of leg segments:
| Leg | Femur | Patella | Tibia | Metatarsus | Tarsus | Total |
|---|---|---|---|---|---|---|
| I♂ | 1.6–2.0 (1.8) | 1.1–1.3 (1.2) | 1.3–1.6 (1.4) | 1.0–1.2 (1.1) | 0.8–1.0 (1.0) | 5.8–7.1 (6.6) |
| II♂ | 1.4–1.7 (1.6) | 0.9–1.2 (1.1) | 1.0–1.3 (1.2) | 0.8–1.1 (1.0) | 0.8–0.9 (0.9) | 5.0–6.3 (5.8) |
| III♂ | 1.3–1.6 (1.5) | 0.7–0.9 (0.8) | 0.8–1.0 (0.9) | 0.9–1.2 (1.1) | 0.6–0.8 (0.7) | 4.3–5.4 (5.0) |
| IV♂ | 1.7–2.0 (1.9) | 1.0–1.2 (1.3) | 1.3–1.6 (1.5) | 1.4–1.8 (1.6) | 0.8–1.0 (0.9) | 6.3–7.6 (7.0) |
| I♀ | 1.4–1.8 (1.7) | 0.9–1.2 (1.1) | 1.0–1.4 (1.3) | 0.8–1.2 (1.0) | 0.8–0.9 (0.8) | 4.9–6.6 (5.9) |
| II♀ | 1.4–1.7 (1.5) | 0.9–1.0 (1.0) | 1.0–1.2 (1.1) | 0.8–1.1 (0.9) | 0.7–0.9 (0.8) | 5.0–5.9 (5.4) |
| III♀ | 1.2–1.4 (1.3) | 0.7–0.8 (0.8) | 0.7–0.9 (0.8) | 0.9–1.1 (1.0) | 0.6–0.8 (0.7) | 4.2–5.2 (4.8) |
| IV♀ | 1.6–2.0 (1.8) | 0.9–1.1 (1.0) | 1.2–1.6 (1.4) | 1.3–1.8 (1.5) | 0.8–1.0 (0.9) | 5.8–7.4 (6.6) |
Length of palp segments (male / female): femur 1.0–1.2 (1.1) / 0.8–1.1 (1.0), patella 0.4–0.5 (0.4) / 0.4–0.5 (0.5), tibia 0.3–0.4 (0.3) / 0.3–0.4 (0.4), tarsus 1.0–1.2 (1.0) / 0.6–0.7 (0.7).
Chelicerae with 2–3 promarginal and 2 retromarginal teeth in males and females. Number of promarginal teeth varies from 3 (most common) to 2 (seldom). One female studied had one chelicera with 3 and the other chelicera with 2 promarginal teeth. Coloration grey.
Male palp as in Figs 25–27. Terminal apophysis sharply turned, without ridge, but with tooth (Tt) in subterminal part, embolus without tooth.
Epigyne as in Figs 30–31. Fovea long, lateral pockets slightly converging, foveal width less that spermathecal span.
Greece, Ukraine (Crimea), Russia (Rostov Area), Kazakhstan (West-Kazakhstan Area) (
Steppes.
In Crimea ♂♀ – X-XII, ♀♀ – II-III, V, the peak activity of adults occurs in December.
In Crimea we found both closely related species, Haplodrassus dalmatensis and Haplodrassus isaevi, to be syntopical in two localities (Sudak Distr., 10 km W Sudak, Mezhdurechie Vill. and Feodosiya Distr., Karadag Nature Reserve). However, these species have quite different phenologies and adults of the two different species do not co-occur. The reproductive period of Haplodrassus dalmatensis is in May-July with the peak in June, and in Haplodrassus isaevi adults can be found in October-December, with their peak of activity in December.
http://species-id.net/wiki/Haplodrassus_kulczynskii
Figs 35–39UKRAINE, CRIMEA:Simferopol Distr.: 2 ♂♂, 1 ♀ (YMC), near Fersmanovo Vill., ~ 250 m, 18.04.–1.05.2000, M.M. Kovblyuk; 1 ♂, 1 ♀ (YMC), Chatyr-Dag Mt., Orlinoe canyon, 27.04.–1.06.2000, M.M. Kovblyuk. Yalta Distr.:2 ♂♂ (TNU), Nikitskaya Yaila Mt. (=Skrinita), 22.04.–25.05.2001, M.M. Kovblyuk.
UKRAINE. Nikolaev Area: 1 ♂ (TNU), Pervomaysky Distr., Migiya Vill., 5.05.–8.06.2006, N.Yu. Polchaninova. AZERBAIJAN. Lenkoran Distr.: 1 ♀ (TNU), Alexeevka Vill., 27.04.2001, E.F. Huseynov.
Haplodrassus kulczynskii is similar to Haplodrassus rugosus Tuneva, 2005 from Kazakhstan and Haplodrassus taepaikensis Paik, 1992 from Korea and the Russian Far East. Both species have a toothed terminal apophysis. Haplodrassus kulczynskii can be easily distinguished from similar species by having a much wider terminal apophysis, having a step-like subterminal outgrowth on the dorsal margin of the RTA (in Haplodrassus rugosus and Haplodrassus taepaikensis such an outgrowth is absent), and by the longer lateral pockets of the epigyne (in Haplodrassus taepaikensis they are shorter; the female of Haplodrassus rugosus is unknown).
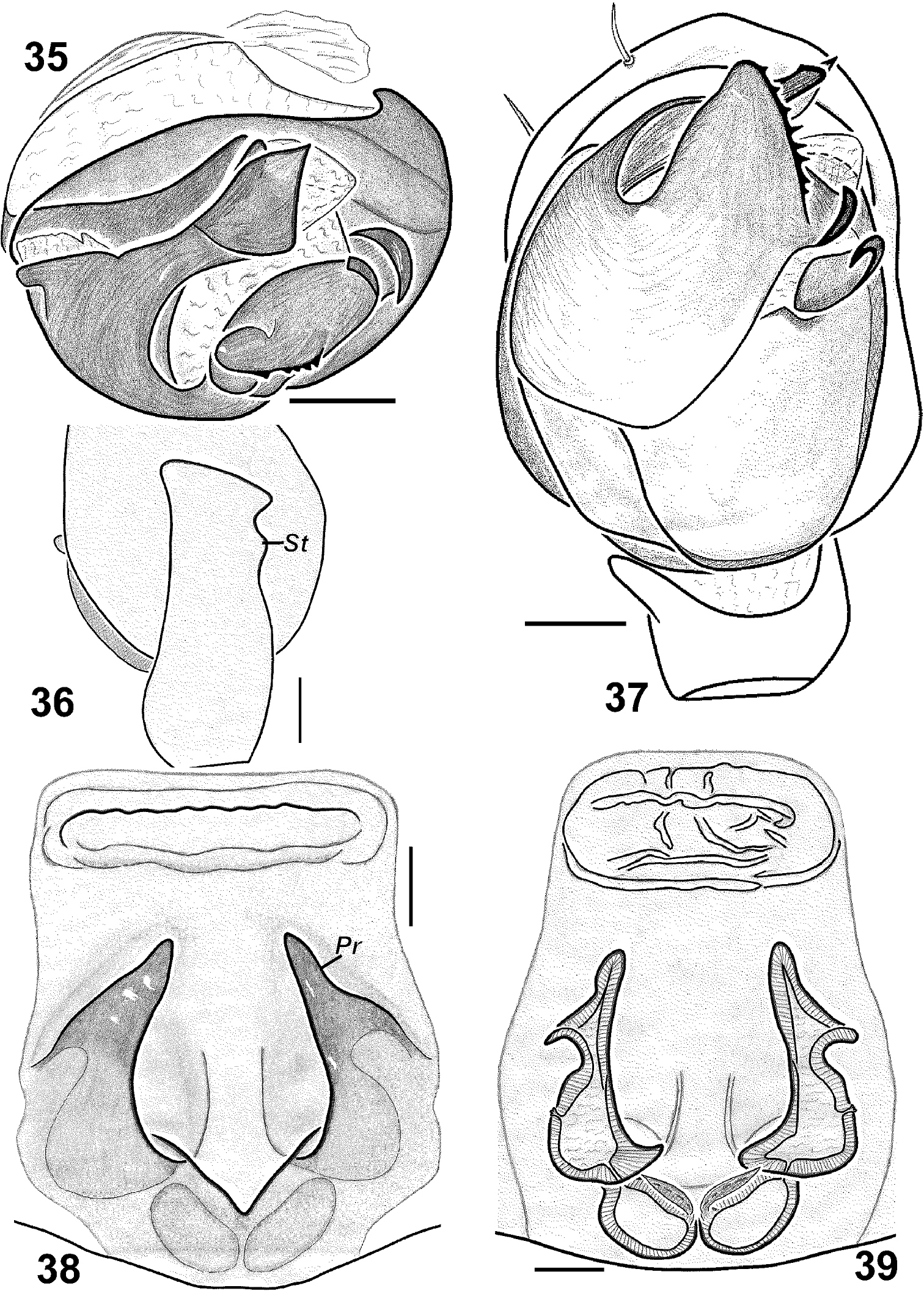
Haplodrassus kulczynskii from Crimea: 35 bulbus, apical view 36 RTA, retrolateral view 37 palp, ventral view 38 epigyne, ventral view 39 epigyne, dorsal view. Abbreviations: Pr protrusion of epigynal pocket; St “step”-like keel of RTA.
West Palaearctic – Far East disjunct nemoral-subtropical range: Central and Southern Europe to Urals, Caucasus, Turkey, Eastern China, Far East Russia and Korea (
Steppe.
In Crimea ♂♀ – IV-V, the peak of activity in adults occurs in April. In Central Europe ♂♀ – IV-VIII (
http://species-id.net/wiki/Haplodrassus_minor
Figs 43–49, 52 –65Ukraine. CRIMEA. Lenino Distr.: 1 ♀ (EMZ), Kerch peninsula, NW coast of Aktash lake, 8.06.1999, M.M. Kovblyuk. Saki Distr.: 5 ♂♂ (TNU), near Pribrezhnaya railway station, 9.05.–3.07.2000, M.M. Kovblyuk. Simferopol Distr.: 1 ♂ (TNU), Kirpichnoe Vill., 31.05.–12.06.1997, M.M. Kovblyuk; 18 ♂♂, 5 ♀♀ (YMC, TNU), Skvortsovo Vill., 27.04.–10.07.2002, M.M. Kovblyuk. Sovietsky Distr.: 1 ♂ (TNU), Uvarovka Vill., 28.04.1999, M.M. Kovblyuk. Yalta Distr.: 1 ♂ (TNU), Yalta, Nikita Vill., 13–30.05.2000, M.M. Kovblyuk.
UKRAINE. Donetsk Area: 1 ♂ (TNU), Slavyansk Distr., Dronovka Vil., 4–8.07.2002, E.V. Prokopenko; 1 ♂, 1 ♀ (TNU), Volodarsky Distr., Nazarovka Vil., “Kamennye Mogily” Nature Reserve, N47°20', E37°06', 20.06.1983, N.Yu. Polchaninova. Nikolaev Area: 1 ♂, 1 ♀ (TNU), Ochakov Distr., Pokrovka Vil., ‘Volyzhin Les’ Department of the Chernomorsky Nature Reserve, 21.05.1987, N.Yu. Polchaninova; 2 ♂♂ (TNU), Pervomaysky Distr., Kuripchane Vil., 6.05.-8.06.2006, N.Yu. Polchaninova. Drawings from these specimens – see in Figs 43–45, 52–53. RUSSIA. Orenburg Area: 5 ♂♂ (TNU), Kuvandyk Distr., Aituar Vill., 22.05.1996, 5.07.2002, N.S. Mazura, T.K. Tuneva; 4 ♂♂, 1 ♀ (TNU), Sol-Iletsk Distr., Chybynda, 5–12.06.2000, S.L. Esyunin. POLAND. 2 ♂♂, 2 ♀♀ (TNU), “Krusne hoty a Polsko, 2001–2002, leg. Prof. E. Kula”.
Haplodrassus deserticola Schmidt & Krause, 1996 from the Canary Islands: 2 ♂♂, 1 ♀ (TNU), leg. et det. J. Wunderlich.
This species is most similar to Haplodrassus deserticola from the Canary Islands (Figs 40–42, 50–51). Haplodrassus deserticola differs from Haplodrassus minor by having a dorsal abdominal pattern (Fig. 66). Haplodrassus minor and Haplodrassus deserticola also differ by the shape of the embolus (thick in Haplodrassus minor, andthin in Haplodrassus deserticola), terminal apophysis (thick in Haplodrassus minor, and thin in Haplodrassus deserticola), median apophysis (short in Haplodrassus minor, and long in Haplodrassus deserticola), epigyne and spermathecae.
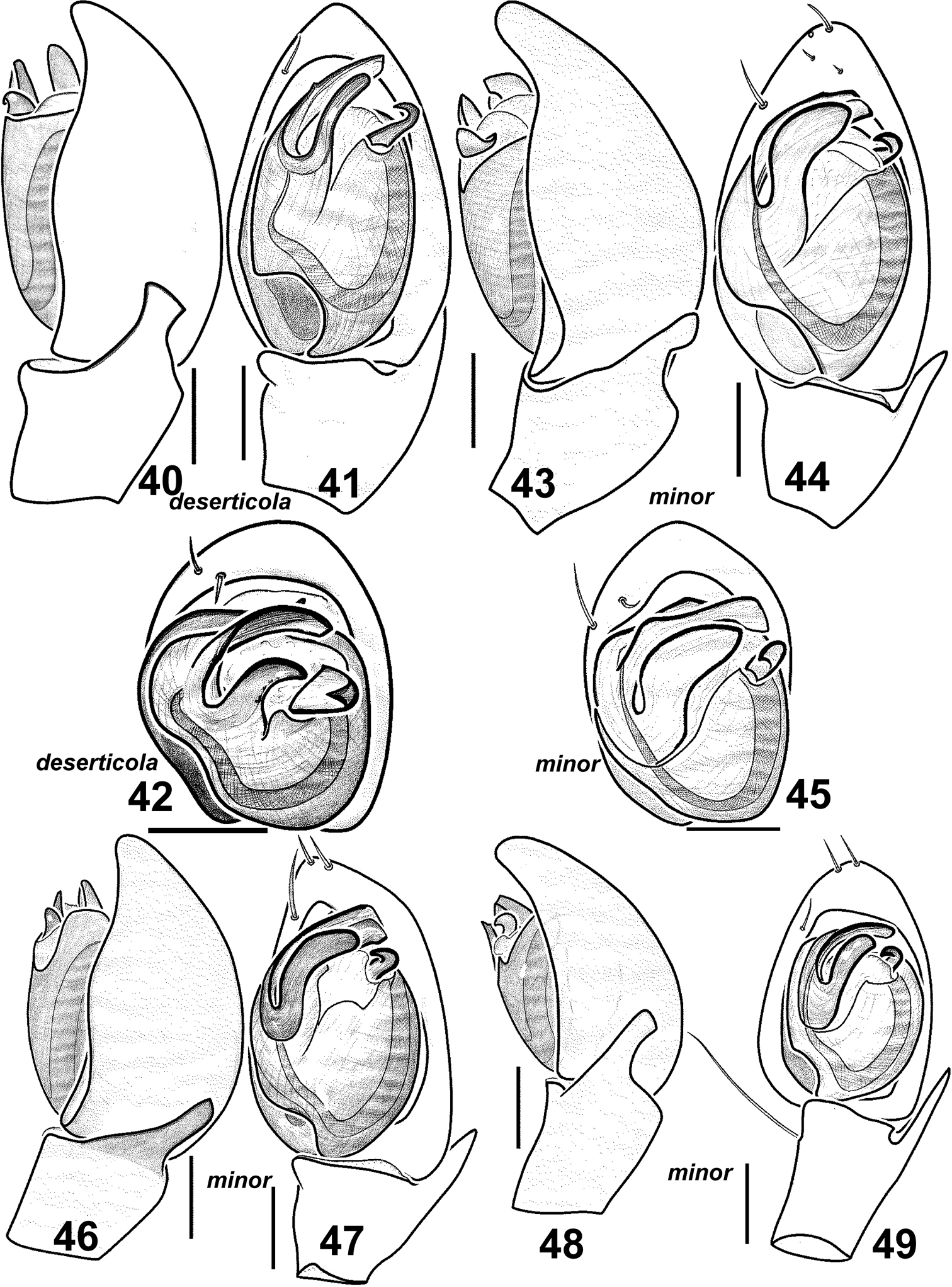
Males of Haplodrassus deserticola (40–42 from Canary Islands) and Haplodrassus minor (43–45 from Poland, 46–47 from Nikolaev Area, 48–49 from Orenburg Area): 40, 43, 46, 48 palp, retrolateral view 41, 44, 47, 49 palp, ventral view 42, 45 bulbus, apical view.
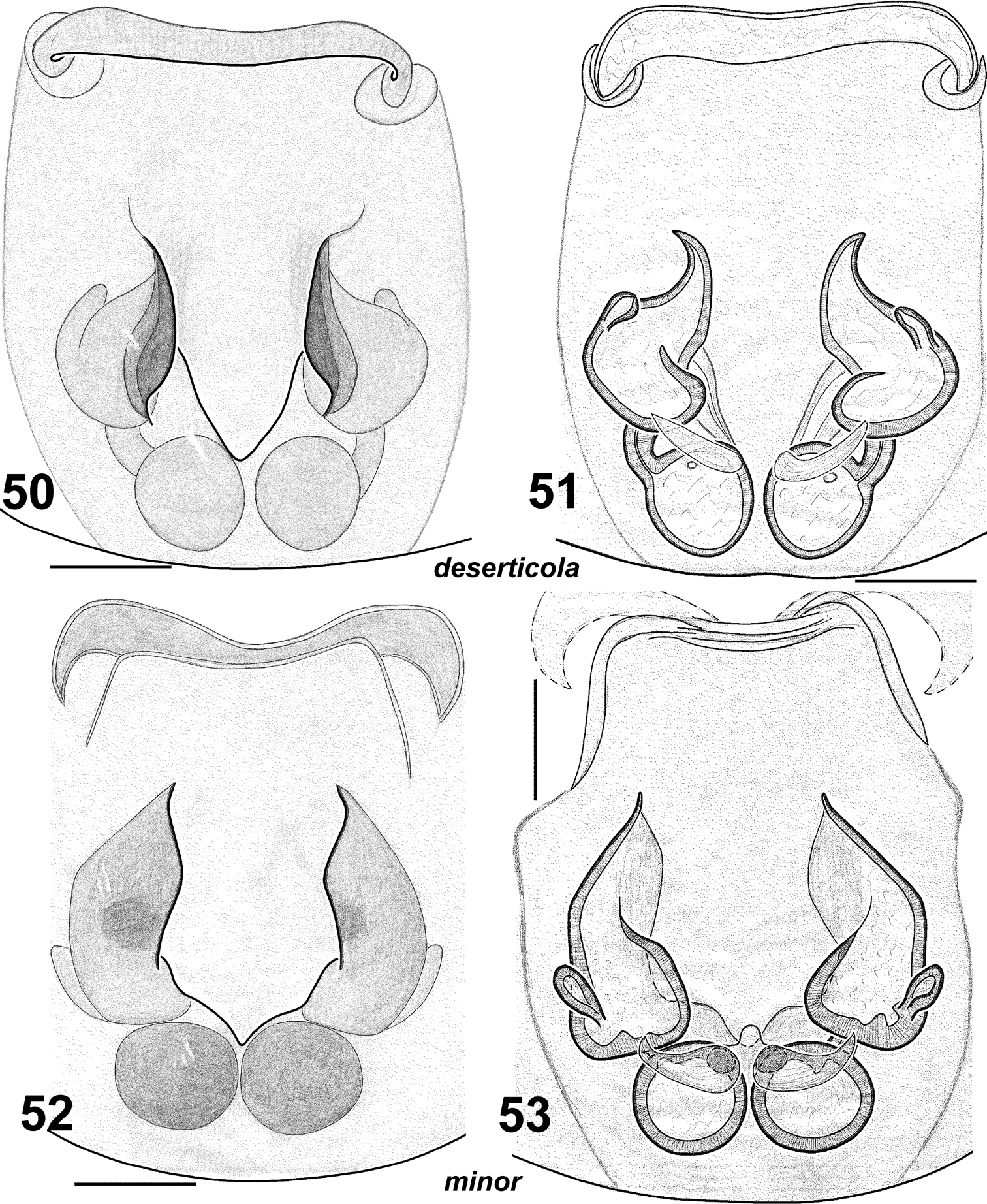
Females of Haplodrassus deserticola (50–51 from the Canary Islands) and Haplodrassus minor (52–53 from Nikolaev Area): 50, 52 epigyne, ventral view 51, 53 epigyne, dorsal view.
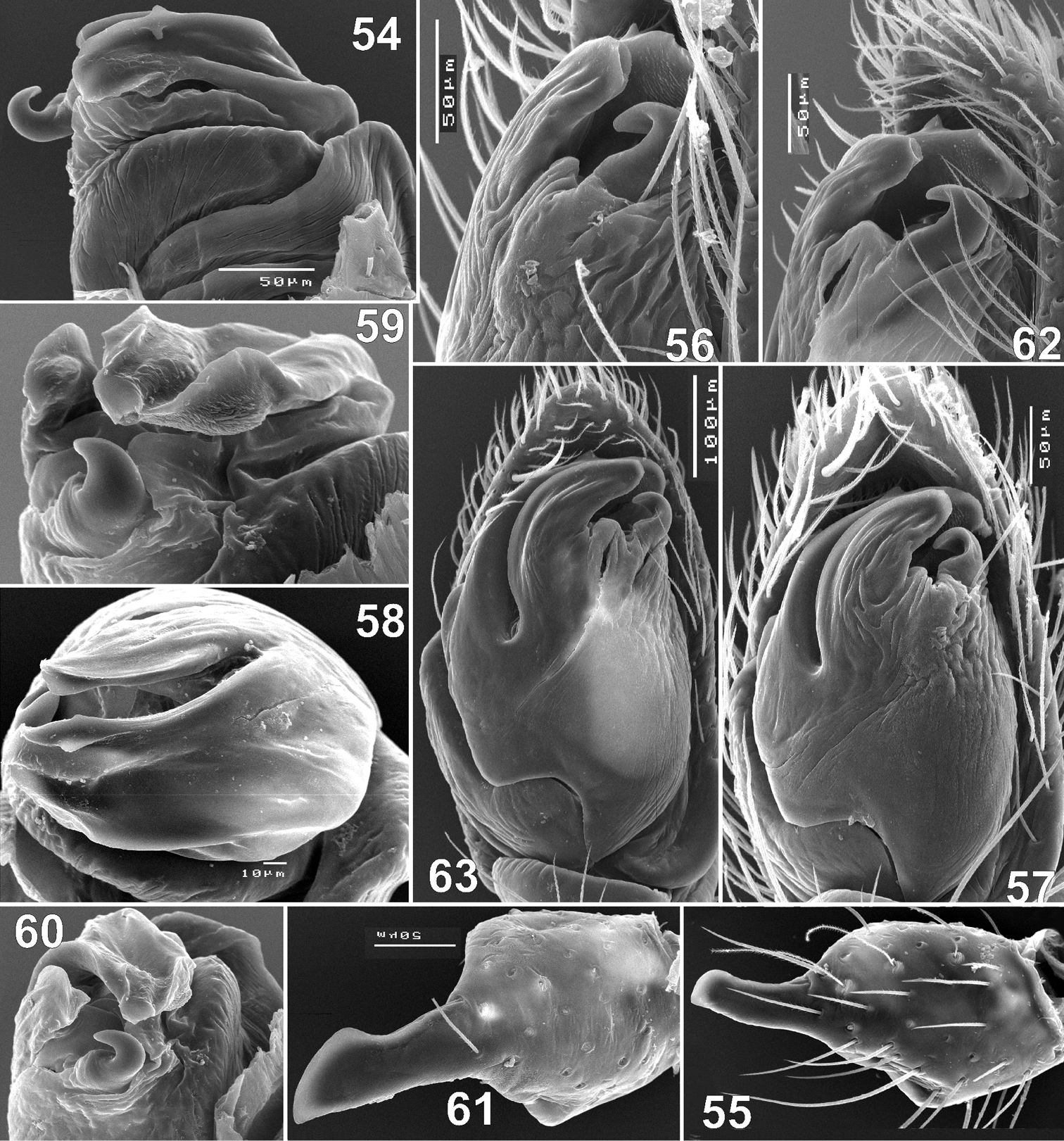
Males of Haplodrassus minor (54–57 from Crimea, 58–63 from Orenburg Area): 54 apical part of bulbus, dorsal view 55, 61 tibia of palp, retrolateral view 56, 62 bulbus, retrolateral view 57, 63 bulbus, ventral view 58 bulbus, apical view 59 bulbus, retrolateral-dorsal view 60 apical part of bulbus, retrolateral view.
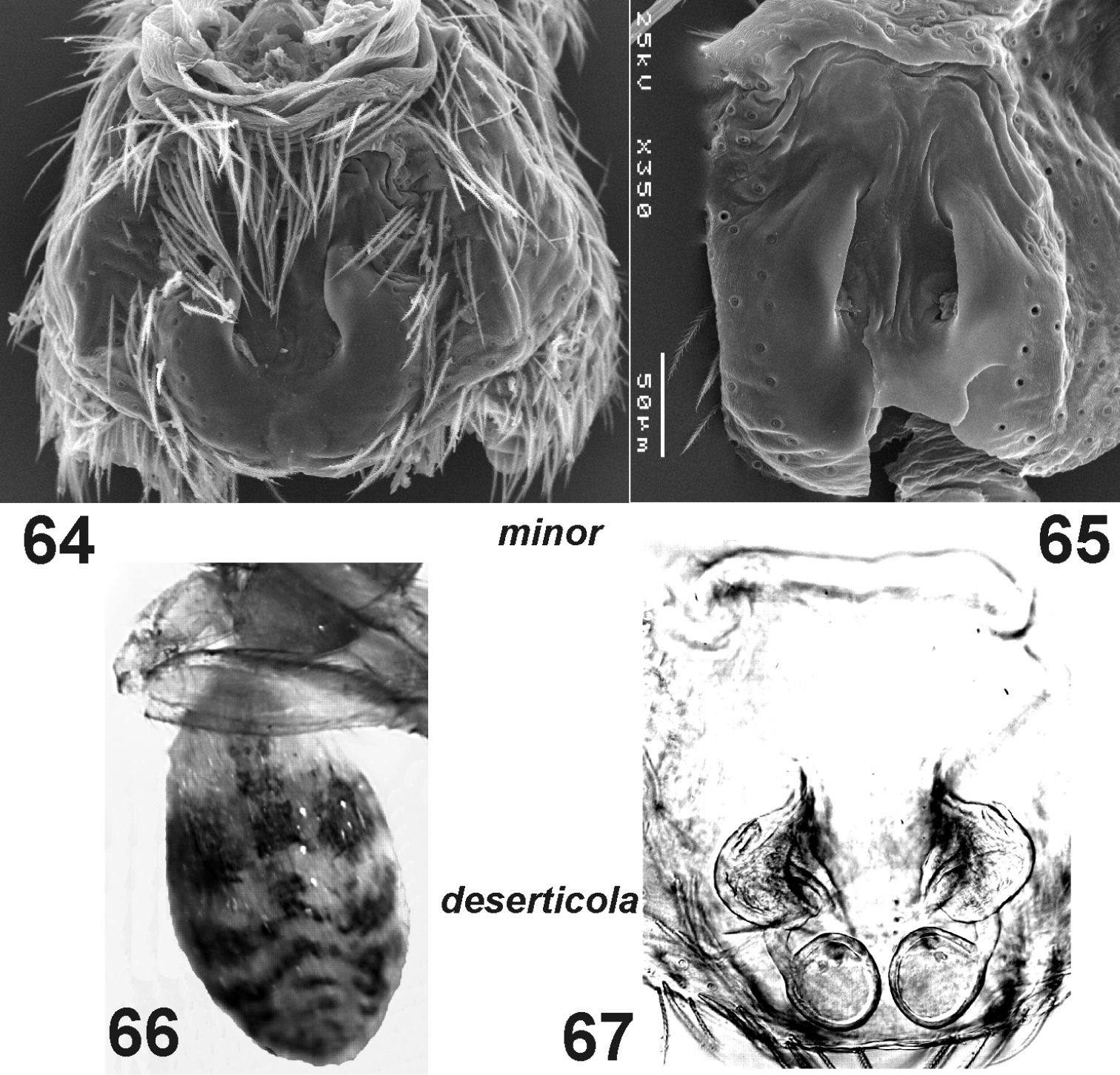
Haplodrassus minor (64 from Crimea, 65 from Orenburg Area) and Haplodrassus deserticola (66–67 from the Canary Islands): 64–65 epigyne, ventral view 66 male abdomen, dorsal view 67 epigyne, dorsal view.
Haplodrassus minor ishighly variable in body size and also in the shape of the bulbal apophyses, RTA (slightly indented or not indented) and the epigyne (Figs 43–65). It is interesting to note that the width of the terminal apophysis decreases with increasing longitude (i.e. from west to east) (cf. Figs 44, 47, 49, 57, 63).
The species has a European range and is known from Portugal to Ural, north to Wales and south to Crete (Tuneva, Esyunin 2003;
Steppes, meadows, forest strips (=shelterbelts) within steppes.
In Crimea ♂♀ – V-VI, ♂♂ – IV, ♀♀ – VII, the peak of activity in adults occurs in May. In Britain ♂♀ – IV-VI, with the peak in June (
http://species-id.net/wiki/Haplodrassus_pseudosignifer
Figs 68–70, 75–76RUSSIA. 2 ♂♂ paratypes (ZMT), SW Altai, 7 km W Katanda, Katun River valley, forest steppe, pitfall traps, 22.06.–26.07.1983, H. Hippa.
UKRAINE, CRIMEA: Feodosiya Distr.:19 ♂♂, 6 ♀♀ (TNU), Karadag Nature Reserve, 15.10.2006–05.2011, M.M. Kovblyuk, O.V. Kukushkin, A.A. Nadolny. Simferopol Distr.: 2 ♂♂, 2 ♀♀ (TNU), Bayrakly Mt. (519 m), ~ 400 m, 14.05.–23.06.2000, M.M. Kovblyuk; 1 ♂ (TNU), Chatyr-Dag Mt., east slope, 10–28.06.2000, M.M. Kovblyuk; 1 ♀ (TNU), near Skvortsovo Vill., 9–30.06.2002, M.M. Kovblyuk. Sudak Distr.: 28 ♂♂, 7 ♀♀ (TNU), 10 km west from Sudak Town, Mezhdurechie Vill., 3.05.–3.06.2010, M.K. Yusufova. Yalta Distr.: 1 ♂ (TNU), Ay-Petri Yaila Mt., west part, 12–13.06.1999, O.V. Kukushkin; 15 ♂♂, 1 ♀ (TNU), Nikitskaya Yaila Mt. (=Skrinita), ~ 1200 m, 2.06.–24.07.2001, M.M. Kovblyuk.
UKRAINE. Nikolaev Area: 1 ♀ (TNU), Pervomaysky Distr., Kuripchane Vil., 26.05–8.06.2006, N.Yu. Polchaninova.
Identification of this species was based on comparison of our specimens with the male paratypes from ZMT. Specimens from Crimea and Altai differ only slightly in the shape of the tooth on embolus. In our opinion the specimens from Crimea, Nikolaev Area and Altai are conspecific.
Haplodrassus pseudosignifer is very similar to Haplodrassus signifer. The two species have no distinct differences in coloration, size or leg spination, but Haplodrassus pseudosignifer can be differentiated from Haplodrassus signifer by having an almost straight and shorter terminal apophysis and thinner embolus, and by the shape of the lateral pockets and fovea of the epigyne.
Males (n = 5) and females (n = 5). Measurements (♂ / ♀): total length 5.7–8.0 (6.79) / 6.6–10.5 (7.8); carapace 2.8–3.5 (3.1) / 2.6–3.6 (3.3) long, 2.2–2.6 (2.4) / 2.1–3.0 (2.6) wide; abdomen 3.1–4.8 (3.8) / 3.4–6.7 (4.7) long, 1.7–2.4 (2.0) / 2.1–4.2 (2.8) wide.
Length of leg segments:
| Leg | Femur | Patella | Tibia | Metatarsus | Tarsus | Total |
|---|---|---|---|---|---|---|
| I♂ | 1.9–2.6 (2.3) | 1.2–1.6 (1.4) | 1.5–2.1 (1.8) | 1.2–1.7 (1.5) | 1.0–1.2 (1.1) | 6.7–9.2 (8.4) |
| II♂ | 1.6–2.2 (2.0) | 1.0–1.4 (1.2) | 1.2–1.8 (1.5) | 1.1–1.6 (1.3) | 0.8–1.1 (1.0) | 5.8–8.1 (7.0) |
| III♂ | 1.4–1.9 (1.7) | 0.7–1.0 (0.9) | 0.9–1.2 (1.0) | 1.2–1.5 (1.4) | 0.8–1.1 (0.9) | 5.2–6.7 (5.9) |
| IV♂ | 2.0–2.6 (2.3) | 1.0–1.4 (1.2) | 1.5–2.02 (1.8) | 1.7–2.4 (2.0) | 1.0–1.3 (1.1) | 7.2–9.8 (8.5) |
| I♀ | 1.8–2.4 (2.2) | 1.0–1.5 (1.4) | 1.4–1.9 (1.7) | 1.1–1.5 (1.3) | 0.8–1.1 (1.0) | 6.2–8.4 (7.5) |
| II♀ | 1.6–2.2 (1.9) | 1.0–1.4 (1.2) | 1.1–1.6 (1.4) | 1.0–1.4 (1.2) | 0.8–1.0 (0.9) | 5.6–7.5 (6.7) |
| III♀ | 1.4–1.9 (1.7) | 0.8–1.0 (0.9) | 0.8–1.2 (1.0) | 1.1–1.5 (1.3) | 0.7–1.0 (0.9) | 4.9–6.6 (5.8) |
| IV♀ | 2.0–2.6 (2.4) | 1.1–1.4 (1.3) | 1.4–2.0 (1.8) | 1.6–2.2 (2.0) | 0.9–1.2 (1.1) | 7.0–9.5 (8.5) |
Length of palp segments (male / female): femur 1.0–1.3 (1.2) / 0.8–1.2 (1.1), patella 0.4–0.7 (0.5) / 0.5–0.6 (0.5), tibia 0.3–0.4 (0.38) / 0.4–0.6 (0.5), tarsus 1.0–1.2 (1.1) / 0.7–0.9 (0.8).
Chelicerae with 2–3 promarginal and 2 retromarginal teeth in males and females. Number of promarginal teeth varies from 2 (frequently) to 3 (rarely).
Coloration grey.
Male palp as in Figs 68–70. Terminal apophysis short (length/width ratio ~ 2) and straight, ridge poorly developed; embolus almost straight and with a tooth.
Epigyne as in Figs 75–76. Fovea elongated, rectangular (longer than wide) with narrow longitudinal groove.
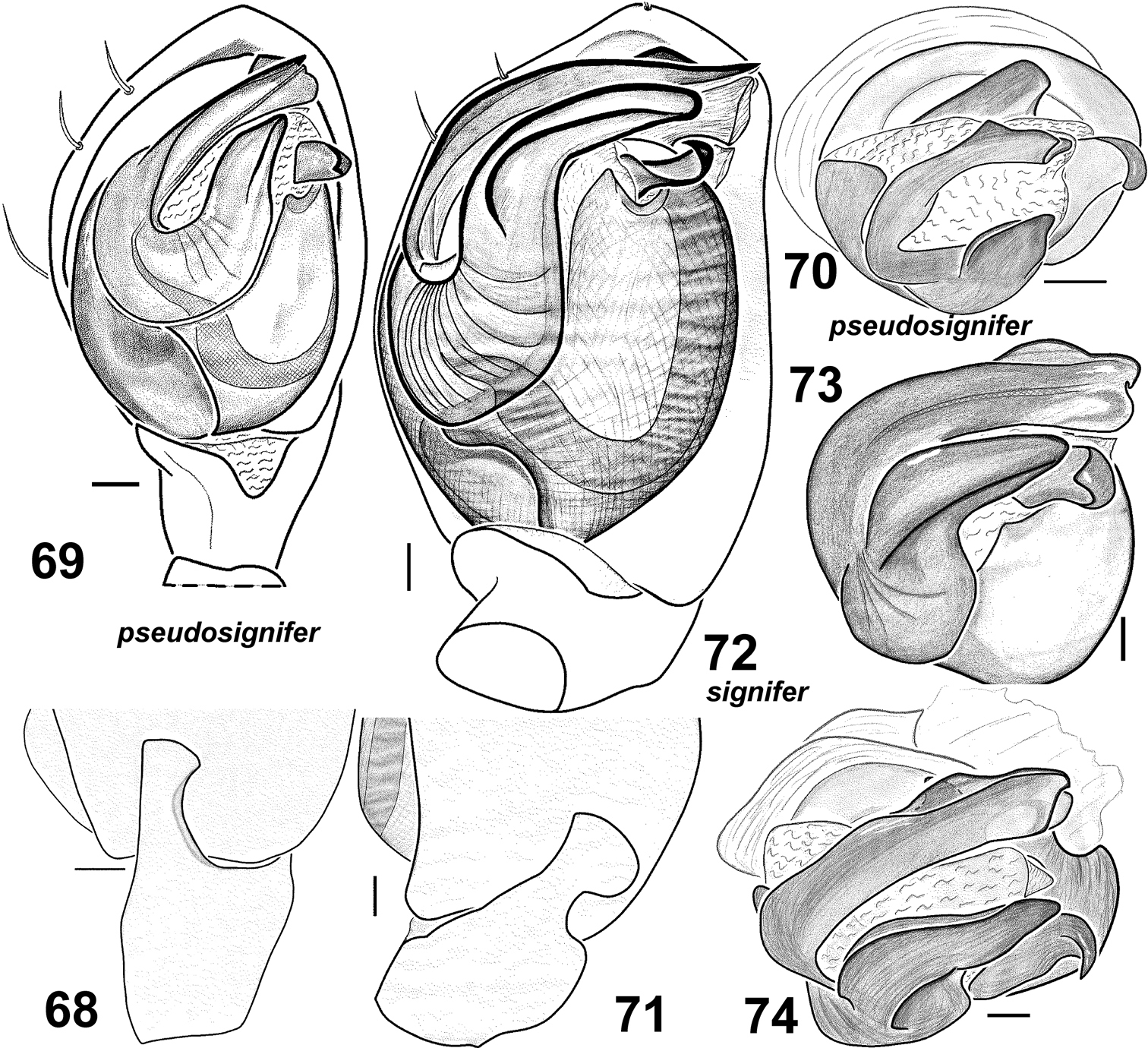
Males of Haplodrassus pseudosignifer (68–70 from Crimea) and Haplodrassus signifer (71–74 from Crimea): 68, 71 RTA, retrolateral view 69, 72 palp, ventral view 70, 73–74 bulbus, apical view.
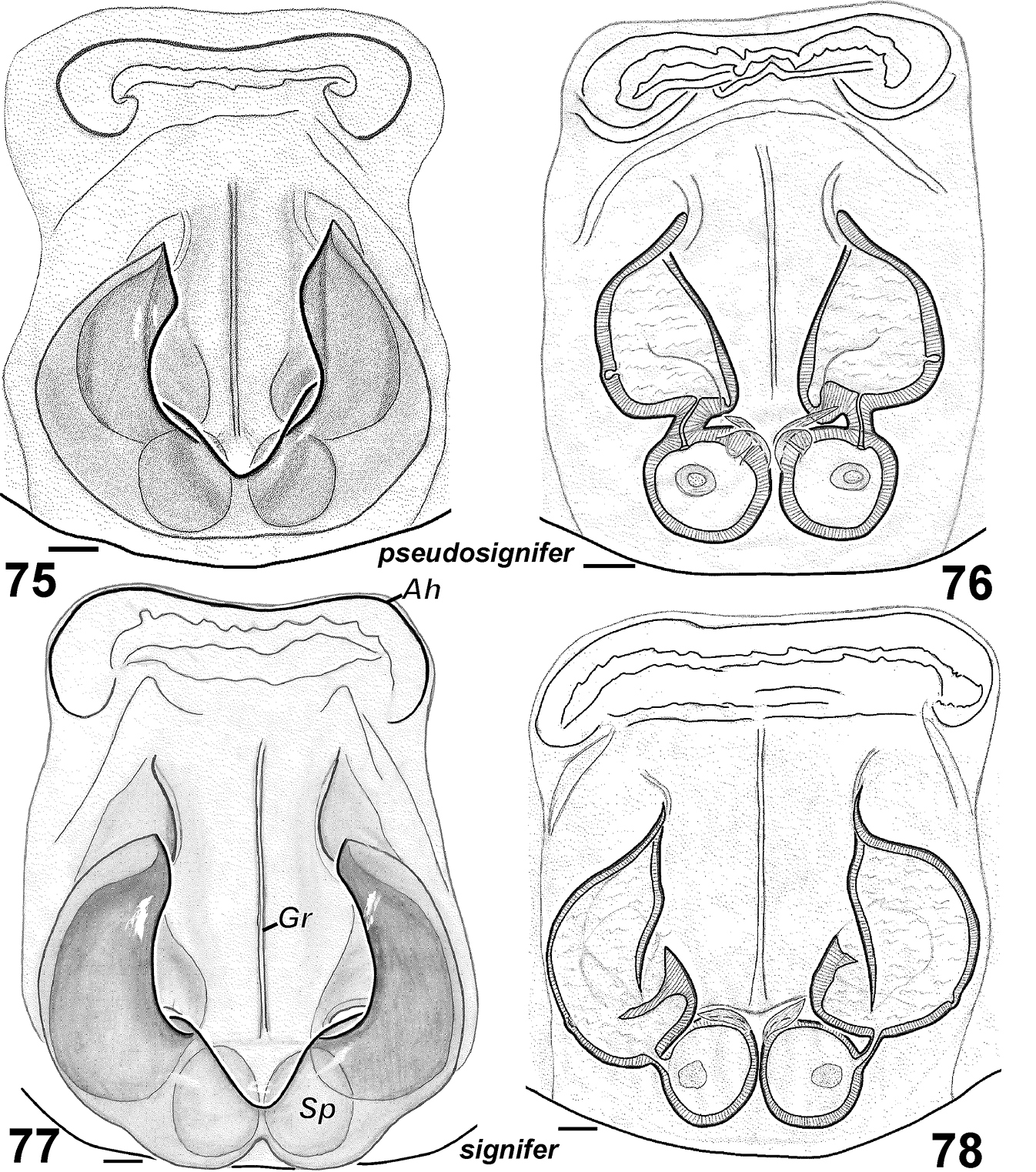
Females of Haplodrassus pseudosignifer (75–76 from Crimea) and Haplodrassus signifer (77–78 from Crimea): 75, 77 epigyne, ventral view 76, 78 epigyne, dorsal view. Abbreviations: Ah anterior hood; Gr groove of epigyne; Spspermatheca.
Steppe zone of Eurasia: from Crimea and Nikolaev Area of Ukraine east to Altai (
Haplodrassus pseudosignifer is a new species record for Crimea and Ukraine. Crimea is the westernmost point of the known distribution range.
Forests with Pistaca mutica or Quercus pubescens, forest-steppes, forest stripes (=shelter belts), rocky steppes, grasslands.
♂♀ – V-VII, ♂♂ – IV, X. In Crimea the peak of activity in adults occurs in May.
http://species-id.net/wiki/Haplodrassus_signifer
Figs 71–74, 77–78UKRAINE, CRIMEA:Feodosiya Distr.:5 ♂♂, 2 ♀♀ (TNU), Karadag Nature Reserve, 27.04.2004.–23.05.2008, M.M. Kovblyuk, O.V. Kukushkin. Saky Distr.: 121 ♂♂, 27 ♀♀ (TNU), near Pribrezhnaya railway station, 16.04.–24.06.2000, M.M. Kovblyuk. Simferopol Distr.: 2 ♂♂ (EMZ), near Simferopol water reservoir, 2.05.1997, M.M. Kovblyuk & G.V. Reutov; 1 ♀ (EMZ), near Strogonovka Vill., 16.05.1997, M.M. Kovblyuk & S. Dyadyushkin; 46 ♂♂, 1 ♀ (EMZ), near Kirpichnoe Vill., 14.05.–12.06.1997, M.M. Kovblyuk; 1 ♀ (TNU), near Fersmanovo Vill., Kesslers’ forest, 350–400 m, 6–23.06.2000, M.M. Kovblyuk; 1 ♂ (TNU), Bayrakly Mt. (519 m), ~ 400 m, 26.05.–6.06.2000, M.M. Kovblyuk; 3 ♂♂, 2 ♀♀ (TNU), near Lozovoe Vill., ~ 250 m, 1.05.–6.06.2000, M.M. Kovblyuk; 8 ♂♂ (TNU), near Simferopol water reservoir, 1.05.–23.06.2000, M.M. Kovblyuk; 2 ♂♂, 1 ♀ (TNU), Simferopol, Bitak Mt., 3.05.–6.06.2000, M.M. Kovblyuk; 4 ♂♂ (TNU), Chatyr-Dag Mt., 20–17.07.2000, M.M. Kovblyuk; 267 ♂♂, 51 ♀♀ (TNU), near Skvortsovo Vill., 12.03.–30.06.2002, M.M. Kovblyuk; 1 ♀ (TNU), Lozovoe Vill., 18.05.2006, M.M. Kovblyuk. Sudak Distr.: 9 ♂♂ (TNU), 10 km W from Sudak Town, Mezhdurechie Vill., 6.05.–22.05.2010, M.K. Yusufova. Yalta Distr.: 5 ♂♂, 3 ♀♀ (TNU), 1 km N Nikita Vill., 22.04.–30.05.2000, M.M. Kovblyuk; 1 ♂, 2 ♀♀ (YMC), Nikitskaya Yaila Mt. range, (=Skrinita), ~ 1200 m, 2.06.–3.07.2001, M.M. Kovblyuk; 2 ♂♂ (TNU), Yalta Mountain-Forest Natural Reserve, near Upper Nikita Lake, 28.04–12.05.2002, A.A. Khaustov; 2 ♂♂, 2 ♀♀ (TNU), near Nikita Vill., 13–26.05.2002, A.A. Khaustov.
SLOVAKIA. 2 ♂♂, 2 ♀♀ (TNU), Nova’ky, 11.05.1990, S. Pekar. UKRAINE. Chernovtsy Area:1 ♀ (TNU), Tsetsino Town, 8–19.05.2009, V.V. Garashchuk, T.O. Auzyak. Donetsk Area: 1 ♀ (TNU), Slavyansky Distr., Svyatogorsk Town, N49°02', E37°39', 07.2004, N.Yu. Polchaninova. Kherson Area: 2 ♂♂, 3 ♀♀ (TNU), Henichesk Distr., Arabatskaya strelka, 4 km S Henichesk Town, 23–30.05.2011, N.A. Stasyuk. RUSSIA. Leningrad Area: 3 ♂♂, 2 ♀♀ (TNU), Nizhnesvirskyi Reserve, 21.07.1994, T.I. Oliger. Kursk Area: 2 ♂♂, 1 ♀ (TNU), Medvensky Distr., “Kazatskaya Steppe” Department of the Tsentral’no-Chernozemny Nature Reserve, 51°30'N, 36°17'E, 20.05.–30.06.2004, N.Yu. Polchaninova. Rostov Area: 8 ♂♂, 1 ♀ (TNU from CP), Orlovsky Distr., Rostovsky Narure Reserve, Starikovskyi region, 15.06.2003 & 7.05.2004, Z.G. Prishutova. ABKHAZIA. Gagra Distr.: 3 ♀♀ (TNU), Gagra Distr., Gagra Range, Mamdzyshkha Mt. (1866 m), from border of forest (43°18'N, 40°19'E, 1705 m) to the peak, wood (Abies, Fagus, Acer) and alpine meadows, 7–15.07.2009, M.M. Kovblyuk.
The species can be easily recognized by the shape of the terminal apophysis, which has a peculiar long ridge, and also by the shape of the epigyne. Haplodrassus signifer is very similar to Haplodrassus pseudosignifer (see the diagnosisfor Haplodrassus pseudosignifer).
Circum-Holarctic polyzonal range (
Steppes, meadows, shrubby communities, forests.
♂♀ – IV-VII, ♂♂ – III. In Crimea the peak activity in adults’ occurs in May. In Central Europe ♂♀ – IV-XII (
Haplodrassus signifer is the largest and most abundant Haplodrassus species in Crimea.
http://species-id.net/wiki/Haplodrassus_umbratilis
igs 79–83Ukraine. CRIMEA. Feodosiya Distr.: 2 ♂♂, 1 ♀ (TNU), Karadag Nature Reserve, thalweg Karadag beams, 44°55'11.4"N, 35°12'25.5"E, 43 m, 9.05.–6.06.2008, M.M. Kovblyuk. Simferopol Distr.: 1 ♀ (TNU), 2 km N Pionerskoe Vill., 10.06.1998, M.M. Kovblyuk; 4 ♂♂ (TNU), 1, 5 km NE Fersmanovo Vill., Kesslers’ forest, 14.05.–6.06.2000, M.M. Kovblyuk; 64 ♂♂, 15 ♀♀ (TNU), Chatyr-Dag Mt., 23.04.–2.09.2000, M.M. Kovblyuk. Sudak Distr.: 1 ♂ (TNU), 10 km W Sudak Town, Mezhdurechie Vill., 6–8.05.2010, M.K. Yusufova.Yalta Distr.: 68 ♂♂, 27 ♀♀ (TNU), Nikitskaya Yaila Mt. (=Skrinita), ~ 1200 m, 4.05.–10.11.2001, M.M. Kovblyuk.
UKRAINE. Donetsk Area: 5 ♀♀ (TNU), Slavyansky Distr., Svyatogorsk Town, N49°02', E37°39', 8–30.06.2004, N.Yu. Polchaninova. Kherson Area:6 ♂♂, 4 ♀♀ (TNU), Golopristynskiy Distr., Chernomorskiy Reserve, Rybalchie Vill., 04–08.1989, Zelinskaya.
Haplodrassus soerenseni (Strand, 1900): UKRAINE. Sumy Area: 1 ♀ (TNU), Vakolovshchina Vill., 1.06.1990, V.A. Gnelitsa.
Haplodrassus umbratilis can be easily differentiated by from all other Haplodrassus species found in Crimea by its terminal apophysis, which has a broad process (Bp). From the similar Haplodrassus soerenseni males differ in the shape of the terminal apophysis and embolus, and females by having longer lateral epigynal pockets.

Haplodrassus umbratilis from Crimea: 79 bulbus, apical view 80 RTA, retrolateral view 81 palp, ventral view 82 epigyne, ventral view 83 epigyne, dorsal view. Abbreviations: Bp broad process of terminal apophysis.
West and Central Palaearctic: all Europe, Turkey, Caucasus, mountains of Central Asia and South Siberia (
Dry forests, forest edges, meadows and steppes.
♂♀ – V-VII, ♂♂ – IV, ♀♀ – VIII, X, the peak of activity in adults occurs in June. In Central Europe ♂♀ –VI-VIII (
The number of Haplodrassus species found in Crimea is rather high. Eight species, as in Crimea are known in the larger area of Bulgaria and Hungary (
Many specimens were collected using pitfall traps, which were regularly checked once in two week during one or two years. Thus, it was possible to analyze the seasonal dynamics of adult activity. All Crimean Haplodrassus species have one peak of activity of adults during the year. The maximum number of individuals and peak of activity for the adults of Haplodrassus kulczynskii occurred in April; for Haplodrassus bohemicus, Haplodrassus minor, Haplodrassus pseudosignifer and Haplodrassus signifer in May; for Haplodrassus dalmatensis and Haplodrassus umbratilis in June; for Haplodrassus isaevi in December. Probably all of the species studied have only one generation per year.
We are very grateful to G.N. Azarkina (Novosibirsk, Russia), S.L. Esyunin (Perm, Russia), M.M. Fedorjak (Chernivtsy, Ukraine), E.F. Huseynov (Baku, Azerbaijan), S. Koponen (Turku, Finland), Yu.M. Marusik (Magadan, Russia), K.G. Mikhailov (Moscow, Russia), T.I. Oliger (Lodeynoe Pole, Russia), S. Pekar (Brno, Czech Republic), N.Yu. Polchaninova (Kharkiv, Ukraine), A.V. Ponomarev (Rostov-on-Don, Russia), E.V. Prokopenko (Donetsk, Ukraine), M. Řezáč (Prague, Czech Republic), T.K. Tuneva (Yekaterinburg, Russia) and J. Wunderlich (Hirschberg, Germany) who provided some important material used in this study. We are deeply obliged to T.O. Auzyak, E.A. Belosludsev, V.V. Garashchuk, V.A. Gnelitsa, A.A. Khaustov, O.V. Kukushkin, N.S. Mazura, A.A. Nadolny, Ya.I. Ibragimova, E. Kula, Z.G. Prishutova, G.V. Reutov, N.A. Stasyuk, M.K. Yusufova for providing some material. We sincerely thank S.L. Esyunin for the photographs of paratypes of Haplodrassus pseudosignifer, T.K. Tuneva for making measurements of Urals specimens of Haplodrassus minor during our work on a joint (with M.K.) unpublished manuscript, Yuri M. Marusik (Magadan, Russia) for making the SEM photos of Haplodrassus minor and for valuable comments to the manuscript. We thank V.I. Ovtsharenko and C. Deltshev for their reviews of the manuscript. We acknowledge the help of P.E. Gol’din who checked the English of the earlier draft. English of the final draft was kindly checked by David Penney (Manchester). This work of M.K. was supported in part by the Karadag Nature Reserve. M.O. was supported in part by the Russian Foundation for Basic Research (grant № 11–0401716).