





(C) 2010 António Múrias dos Santos. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Recent collections of isopods in Alboran Island and Algeria included several specimens of the species Stenosoma stephenseni sp. n. This is the fourteenth species described in the genus Stenosoma Leach, 1814. Examination of two specimens collected during the Danish oceanographic cruises of the Thor (1908–10) close to the Galite Islands, and identified as Stenosoma acuminatum Leach, 1814, revealed that both belong to Stenosoma stephenseni sp. n. In light of these findings, the Mediterranean records of Stenosoma acuminatum are revised, and it is proposed that Stenosoma acuminatum is a strictly Atlantic species. An updated diagnosis for the genus Stenosoma is given, together with a key for the identification of its species. The nomenclatural status of the name Synisoma Collinge, 1917 is addressed, and although it is in prevailing usage, it is shown that Stenosoma Leach, 1814 is the valid name of the genus.
new species, North Africa, Mediterranean, Alboran Island, Idoteidae
In his work on the isopods collected during the Danish oceanographic cruises of the Thor (1908–10) in the Mediterranean and the Black Sea,
In his revision of the British Idoteids,
While studying the phylogeography of Stenosoma nadejda (Rezig, 1989) we received several specimens from the Alboran Island (provided by JM Guerra García). Analysis of the mitochondrial gene cytochrome c oxidase subunit I revealed that the Alboran specimens belonged to a very divergent lineage from “Stenosoma nadejda” and were potentially a new species (
In this work, we describe a new species of Stenosoma, in which we include the specimens of
Specimens were collected on intertidal algae during low tides, in the winter of 2005 (Alboran Island) and the summer of 2009 (Algeria). All specimens were preserved in 96% ethanol. Description is based on the male holotype unless otherwise stated. Body length measured dorsally from midpoint of cephalon’s anterior margin to posterior of pleotelson. The holotype is deposited in the Zoological Museum, University of Copenhagen, Denmark. All taxonomic work is attributed to A. M. dos Santos and R. Xavier.
AbbreviationsBMNH The Natural History Museum, London, UK
MNHNP Muséum National d’Histoire Naturelle, Paris, France
ZMUC Zoological Museum, University of Copenhagen, Denmark
CIBIO-UP Centro de Investigação em Biodiversidade e Recursos Genéticos, Universidade do Porto, Portugal
ICZN International Code of Zoological Nomenclature (ICZN, 1999).
TaxonomyOrder Isopoda Latreille, 1817
Familiy Idoteidae Samouelle, 1819
http://species-id.net/wiki/Stenosoma
Stenosoma acuminatum Leach, 1814, by subsequent designation of Kussakin, 1982 (under Article 67.8).
The genus Stenosoma was described by
In page 404 of the section “Crustaceology”, Stenosoma was clearly described as a tentative subdivision of the genus Idotea. After the general description of Idotea (numbered as Genus LXIV),
In the Appendix (p. 433), however, Stenosoma is re-described as a genus on its own, this time numbered “XI”, immediately after Idotea (which is Genus X). There, Leach reformulated the diagnosis of Stenosoma (“external antennae longer than the body, the third longer than the fourth joint; body linear”), and included two nominal species, Idotea hectica Pallas, 1772 and Stenosoma acuminatum Leach, 1814. So, under Articles 12.1 and 12.2.5, the name Stenosoma Leach, 1814 is available from page 404, where it is treated as a division of Idotea Fabricius, 1798, and from page 433 of the same publication where it is ranked as a genus. Under Article 24.1, precedence must be accorded to the name proposed at higher rank, i.e., Stenosoma as a genus, in page 433 of the Appendix. The important point here is that on page 433 Leach included in his genus the nominal species Idotea hectica Pallas, 1772 and Stenosoma acuminatum Leach, 1814. Therefore, Stenosoma acuminatum Leach, 1814 is eligible as type species of Stenosoma Leach, 1814 (Articles 67.2 and 67.2.1).
The genus Stenosoma was quickly adopted by some leading French zoologists (e.g.,
In his comprehensive monograph of the Idoteidae,
In his revision of the British idoteids,
Synisoma Collinge, 1917 is currently in prevailing usage, as it has been used virtually in all works published after 1917. To promote nomenclatural stability, the ICZN allows for a reversal of precedence (Article 23.9) whenever a junior synonym is in prevailing usage provided that the two conditions defined in Articles 23.9.1.1 and 23.9.1.2 are both met. In this case, however, the first condition, that the senior synonym has not been used as a valid name after 1899, is not met. In fact, Stenosoma Leach, 1814 was used as a valid name in at least six works posterior to 1899:
Given the complex taxonomic history of the genus Stenosoma and its synonyms, their diagnoses have been modified on an ad-hoc basis to accommodate each new species described. For example, both
Body elongate, lateral margins parallel or sub-parallel, sometimes widening slightly towards pereonites III–IV. Cephalon with pronounced anterolateral lobes, smooth or with a pronounced dorsal tubercle; eyes lateral, small. Antennulae with first article expanded, flagellum composed of a single article. Antenna articles 3–4 longer, flagellum multiarticulated or composed of a single clavate article. Maxillipedal palp with 4 articles. Pereonites smooth, frequently with a shallow dorsal keel, seldom developing into a dorsal triangular tooth; pereonites I–III often with a pair of lateral tubercles. Coxal plates small, round, rarely medium sized and triangular, invisible in dorsal view, or visible dorsally on perionites II–VII or V–VII. Pereopods ambulatory, slender and sub-equal, terminating in a biungulate dactyl with simple setae. Pleon without articulating pleonites (pleotelson), pleonites I–III frequently indicated by incomplete sutures visible ventrolaterally or dorsally (pleotelsonic formula 0+3); pleotelson long, not less than one third of body length, terminally pointed; dorsal surface smooth or with a shallow keel. Penes attached to posterior ventral margin of pleonite 1, fused basally as a penial plate divided over most of its length. Uropod uniramous, endopodite more or less triangular in shape.
Stenosoma acuminatum Leach, 1814; Stenosoma appendiculatum (Risso, 1826); Stenosoma capito (Rathke, 1837); Stenosoma carinatum (Lucas, 1849); Stenosoma lancifer (Miers, 1881); Stenosoma spinosum (Amar, 1957); Stenosoma bellonae (Daguerre de Hureaux, 1968); Stenosoma pacificum (Nunomura, 1974); Stenosoma nadejda (Rezig, 1989); Stenosoma mediterraneum (Rezig, 1989); Stenosoma wetzerae (Ormsby, 1991); Stenosoma raquelae (Hedo & Junoy, 1999); Stenosoma albertoi (Castellanos & Junoy, 2005); Stenosoma stephenseni sp. n.
urn:lsid:zoobank.org:act:EDEC2356-58AE-4DBA-B99D-41042CCB0607
Holotype: ♂ (13.0 mm, partially dissected, preserved in ethanol 96%), Dellys, Boumerdès, Algeria, 36°55'27.14"N, 3°53'42.30"E, 6 Aug 2009, intertidal seaweeds (ZMUC-CRU-20458).
Paratypes: ♂ (12.5 mm), ♀ (11.0 mm), Galite Islands, Bizerte, Tunisia, (approx. 37°31'27.21"N, 8°56'23.54"E), 5 Feb 1909, ‘on the shore' (Stephensen, 1915) (ZMUC-CRU-20228). 2♂ (10.5, 8.9 mm), Dellys, Boumerdès, Algeria, 36°55'27.14" N, 3°53'42.30"E, 6 Aug 2009, intertidal seaweeds, (CIBIO-UP, SstDel5 and SstDel9). 3♂ (10.1, 9.9, 10.5 mm), 3♀ (1 ovig. 11.8 mm, 2 non-ovig. 9.1, 9.8 mm), Tighremt, Bejaïa, Algerie, 36°52'0.60"N, 4°51'25.29"E, 4 Aug 2009, intertidal seaweeds (CIBIO-UP, SstTit4, SstTit2, SstTit18, SstTit1, SstTit15, SstTit17, respectively). 2♂ (13.2, 7.9 mm), Sidi Khaled, Tigzirt, Tizi-Ouzou, Algerie, 36°53'48.52"N, 4°10'52.46"E, 28 Jul 2009, intertidal seaweeds (CIBIO-UP, SstTiz16, SstTiz17). 3 mancas (3.8, 4.1, 4.3 mm), Alboran Island, Spain, 35°56'58.06"N, 3°01'48.57"W, 12 Feb 2005, intertidal seaweeds (CIBIO-UP, SstAlb1-3).
The species is characterised by a smooth and domed cephalon, with a prominent dorsal boss in lateral view; pereonites smooth, lacking lateral tubercles; pereopods II–VII robust, with merus and carpus 1.2 and 1.1 times as wide as long, respectively; pleotelson margins parallel or subparallel, curving regularly towards distal extremity at one third of its length; pleotelson with three pairs of lateral sutures only visible in ventral view; appendix masculina long, extending beyond apical margin of the endopod by more than one fifth of its length, but not beyond apical spines of endopod.
Body elongate, five times as long as wide (Figure 1). No secondary sexual dimorphism observable. Length of specimens in type series: 4.3–13.2 mm. Colour light brown to pale yellow, lightly pigmented.
Cephalon 1.3 times as wide as long, posterior margin immersed in pereonite I, smooth (no signs of mid-dorsal tubercle) but domed, with a prominent dorsal boss in lateral view; eyes dark, triangular or round, on lateral edge of cephalon; supra antennal line straight, anterolateral angles acute. Pereonites smooth, without dorsal carina. Coxal plates small, present on pereonites II–VII and hardly visible in dorsal aspect. All pleonites medially fused, with three pairs of small antero-lateral sutures in ventral view only. Pleotelson 2.4 times as long as wide, approximately one third of total body length.
Antennule: peduncle of three articles, article 1 ovoid, articles 2–3 cylindrical, similar in size; flagellum bearing seven pairs of aesthetascs. Antenna: peduncle of five articles, article 1 reduced, article 2 as wide as long, articles 3–5 progressively longer; flagellum of 17 articles, the distal one with minute vestigial apical article bearing a brush of short setae; flagellum varying from 14 to 17 articles on type series.
Mandible: Right mandible incisor 4 toothed; lacinia mobilis with one or two incisors; spine row with seven curved serrate spines; molar process truncate, without tooth. Maxillule: inner lobe with three distal plumose spines, inner margin with thin simple setae; outer lobe 1.8 times longer than inner lobe, with eight stout spines, four of them serrate; outer margin with small simple setae. Maxilla: trilobate, endopod with seven recurved plumose spines and eight simple setae; inner and outer lobes of exopod with five and four pectinate spines, respectively. Maxilliped: palp 4–articulate; exopod round; endite with a single coupling hook, five spines and a few simple setae along the distal margin.
Pereopods I–VII ambulatory (Figure 2), robust, with merus 1.2 times as wide as long, and carpus 1.1 times as wide as long, terminating in a biungulate dactyl with simple setae; pereopod I with simple spines on inner surface of propodus, and weak setation on ventral margin; pereopods II–VII subsimilar; pereopods II and VI with 8–12 palmate setae on distal margin of propodus.
Ventral penis smooth. Pleopods I–II rami with plumose marginal setae (Figure 3); pleopod II with long appendix masculina, extending beyond endopod by more than one fifth of its length, but not beyond its apical spines, apex distal inner margin serrated, with five minute spines; pleopods III–V 1.1 times longer and 1.2 times wider than I–II, without setae. Uropod: uniramous, with small plumose seta on lateral distal angle of peduncle.
The epithet honours Knud Hensch Stephensen (1882–1947), former curator of the crustacean collections at the ZMUC, who first noticed that some specimens he placed in Stenosoma acuminatum were likely to be a new species from the Mediterranean (Stephensen, 1915).
The material from Thor campaigns in 1908–1810, originally described by
There are three sympatric species with which Stenosoma stephenseni sp. n. can be confounded: Stenosoma nadejda (Rezig, 1989), Stenosoma mediterraneum (Rezig, 1989) and Stenosoma capito (Rathke, 1837). Stenosoma stephenseni sp. n. can be easily distinguished from all three species, as these have a mid-dorsal tubercle on the cephalon, one pair of lateral tubercles on the first two (Stenosoma capito) or three (Stenosoma nadejda and Stenosoma mediterraneum) pereonites, and more slender pereopods, with carpus and merus at least 1.5 times as long as wide. The appendix masculina does not extend beyond the apical margin of the endopod in Stenosoma nadejda (see
As discussed below, the inclusion of Stephensen’s specimens labeled “Stenosoma acuminatum” in Stenosoma stephenseni sp. n. has implications for the distribution of Stenosoma acuminatum. According to (
After the description of Stenosoma acuminatum by Leach (1814), many authors opted to synonymise it with Stenosoma appendiculatum (Risso, 1826) or Stenosoma capito (Rathke, 1837).
Neither
By including the two specimens from the campaigns of the Thor (
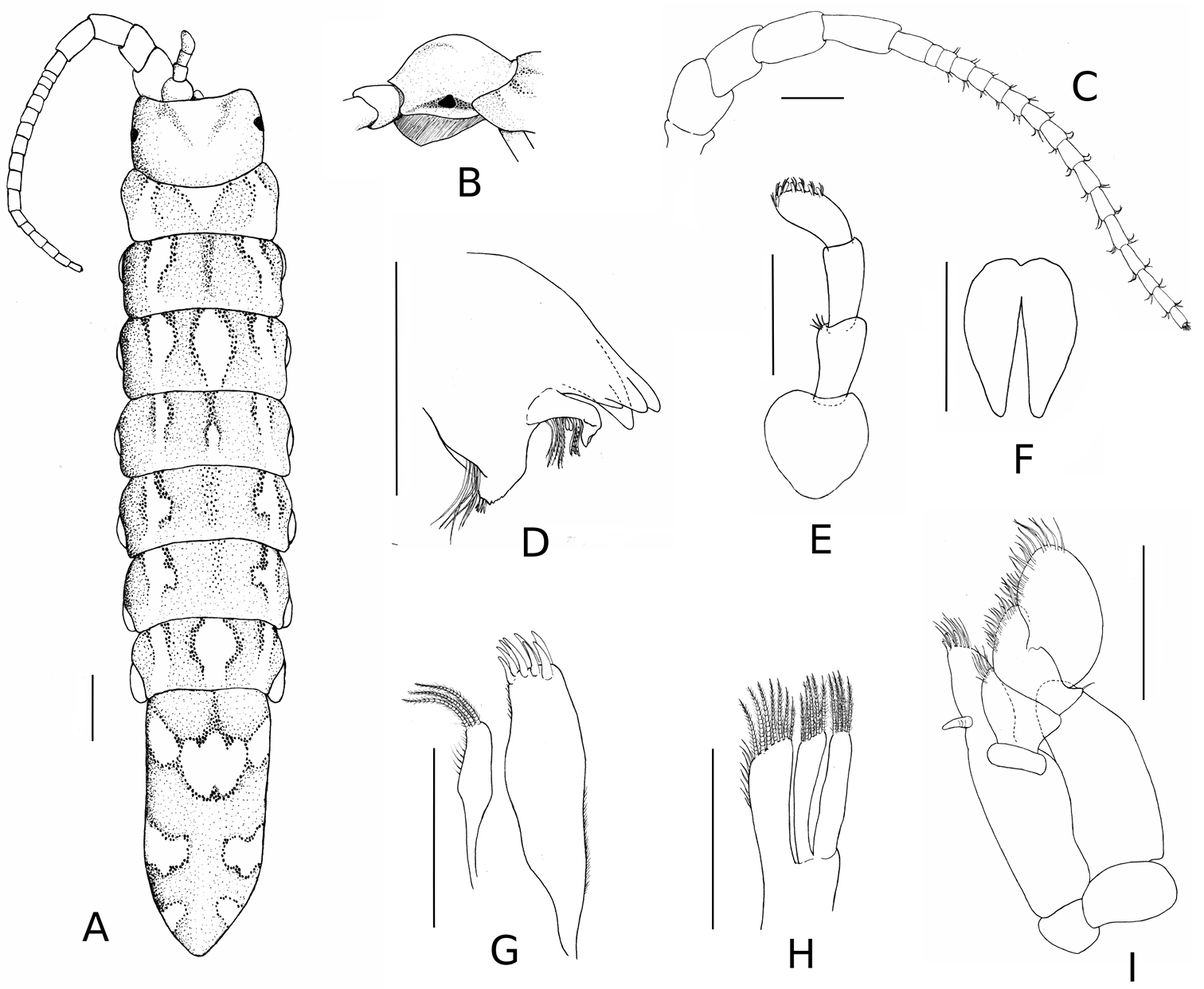
Stenosoma stephenseni, sp. n., holotype: A dorsal view B detail of cephalon C antenna D left mandible E antennula F penis G maxillule H maxilla I maxilliped. Scale bars are 500 µm, except for whole specimen (1 mm).
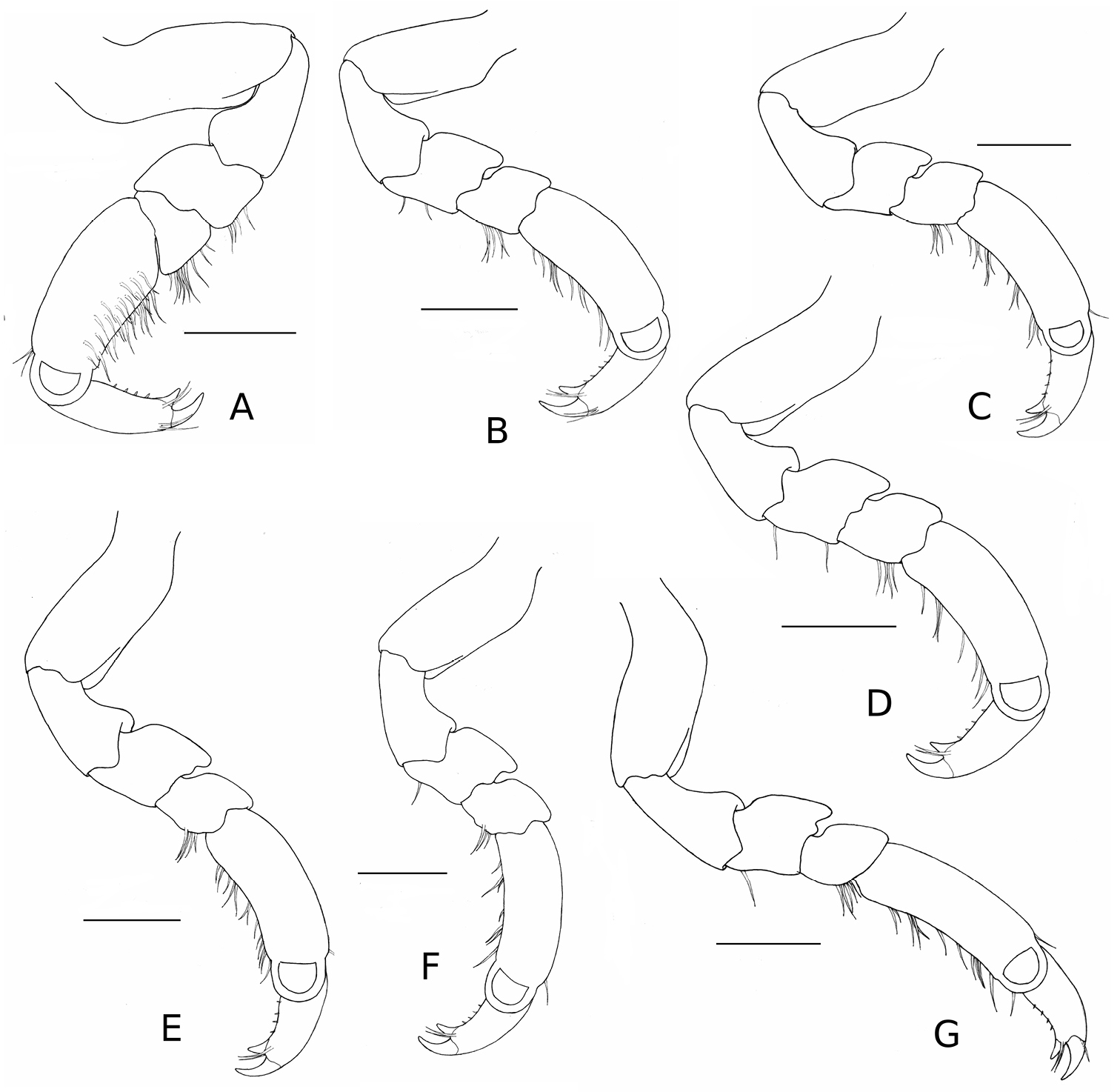
Stenosoma stephenseni, sp. n., holotype: A pereopod I B pereopod II C pereopod III D pereopod IV E pereopod V F pereopod VI G pereopod VII. Scale bars are 500 µm.
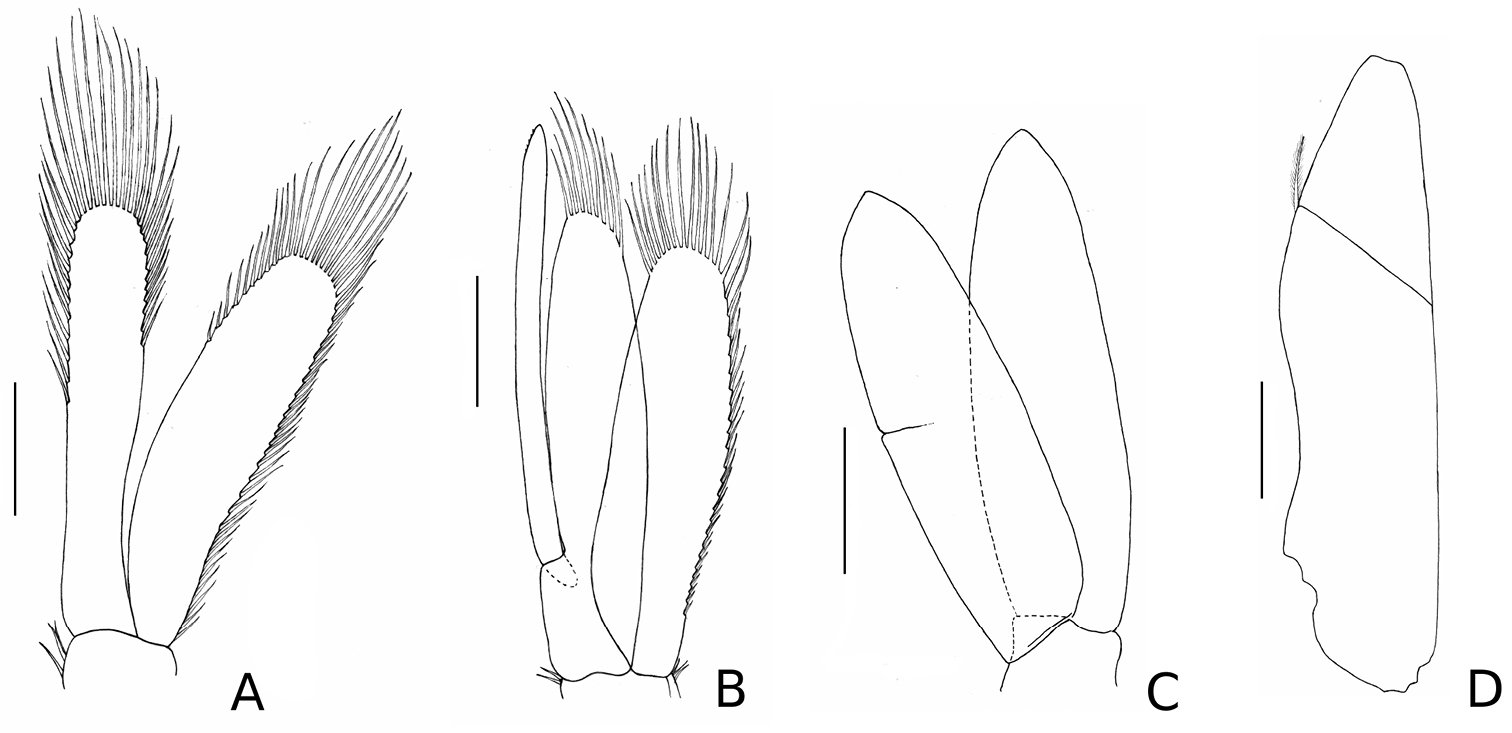
Stenosoma stephenseni, sp. n., holotype: A pleopod I B pleopod II C pleopod III D uropod. Scale bars are 500 µm.
During this work, the description of Idotea angustata Lucas, 1849 came to our attention. This species was described from Algiers (Algeria), and judging from its original description, clearly belongs to the genus Stenosoma, together with Idotea carinata Lucas, 1849.
Lucas refers that “La tête est légèrement gibbeuse” [the head is slightly convex] and that “Les organes de la locomotion sont courtes et assez robustes” [the organs of locomotion are short and rather robust], but the lack of any reference to the presence/absence of lateral tubercles in the first pereonites, and the exact shape of the pleotelson and the protuberance of the cephalon make this description ambiguous. Hence the name Idotea angustata which, according to the rules of the ICZN, is available from
Key to the species of the genus Stenosoma
| 1 | Antenna with multiarticulated flagellum | 2 |
| – | Antenna with single clavated flagellar article | 12 |
| 2 | Pleotelson without anterolateral sutures in either dorsal or lateral views | 3 |
| – | Pleotelson with one or three anterolateral sutures in dorsal or lateral views | 8 |
| 3 | Pleotelson with three suture lines in ventral view | 4 |
| – | Pleotelson without suture lines in ventral view | 5 |
| 4 | Cephalon with mid-dorsal tubercle; pereopods II–VII slender, with carpus and merus longer than wider; pereonites I–III with lateral tubercles | Stenosoma nadejda (Rezig, 1989) |
| – | Cephalon smooth, domed; pereopods II–VII robust, with carpus and merus slightly wider than longer; pereonites I–III smooth | S. stephenseni sp. n. |
| 5 | Cephalon with a mid-dorsal tubercle or spine | Stenosoma wetzerae (Ormsby, 1991) |
| – | Cephalon smooth | 6 |
| 6 | Pereon sides straight and parallel, coxal plates barely visible from above; pleotelson sides narrowing fairly evenly to an acute terminal projection | 7 |
| – | Pereon sides appearing serrated, coxal plates triangular in dorsal view; pleotelson shape like an ink pen nib | Stenosoma lancifer (Miers, 1881) |
| 7 | Antenna large, flagellum with more than seven articles | Stenosoma acuminatum Leach, 1814 |
| – | Antenna short, flagellum with 5–7 articles | Stenosoma pacificum (Nunomura, 1974) |
| 8 | Pleotelson with one anterolateral suture in dorsal or lateral views | 9 |
| – | Pleotelson with three anterolateral sutures in dorsal or lateral views | 10 |
| 9 | Cephalon with a bilobed mid-dorsal tubercle; pereonites bearing a mid-dorsal spine | S spinosum (Amar, 1957) |
| – | Cephalon smooth; body with dorsal carina | Stenosoma appendiculatum (Risso, 1826) |
| 10 | Dorsal surface of anterior pereonites with tubercles; pleotelson narrow | 11 |
| – | Dorsal surface of anterior pereonites smooth, not tuberculate; pleotelson wide, shield-shaped (about 1.2 times as long as wide) | Stenosoma carinatum (Lucas, 1849) |
| 11 | Pereon with a mid-dorsal carina; one pair of lateral tubercles on the first two pereonites; pleotelson shape like an ink pen nib, with three anterolateral sutures visible laterally only | Stenosoma capito (Rathke, 1837) |
| – | Pereon without carina; one pair of lateral tubercles on the first three pereonites; pleotelson sides narrowing fairly evenly to an acute terminal projection, with three anterolateral sutures visible in dorsal view | Stenosoma mediterraneum (Rezig, 1989) |
| 12 | Pleotelson wide (length about 1.2 times width), shield-shaped, first suture larger than others | Stenosoma bellonae (Daguerre de Hureaux, 1968) |
| – | Pleotelson narrow (length equal to or more than 1.5 times width), lateral sutures short, all of the same length | 13 |
| 13 | Cephalon smooth; body without dorsal carina; maxilliped with one coupling hook | Stenosoma raquelae (Hedo & Junoy, 1999) |
| – | Cephalon with a bilobed mid-dorsal tubercle; body with dorsal carina; maxilliped with two coupling hooks | Stenosoma albertoi (Castellanos & Junoy, 2005) |
This work was partly financed by research project PTDC/MAR/104169/2008 from Fundação para a Ciência e a Tecnologia (FCT). M. Branco and R. Xavier have, respectively, postdoc and PhD grants from Fundação para a Ciência e a Tecnologia, (SFRH/BPD/40073/2007 and SFRH/BD/29370/2006). We would like to thank to: J.M. Guerra-García, for kindly providing the specimens from Alboran Island; M. Lowe (The Natural History Museum, London) for providing the relevant information on the specimens deposited at the BMNH, especially those in W. E. Leach’s collection; M.J. Santos, for providing access to her laboratory facilities, especially the imaging systems; T. Schiotte (ZMUC) for the help with Stephensen’s material. Our thanks are also due to V. McAtear (Librarian of the Royal Entomological Society, St. Albans) and F. P. Lima for their invaluable help in obtaining photocopies of some rare works.