





(C) 2012 Mauro Rampini. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
A description of four new species of Dolichopoda Bolivar, 1880 (Orthoptera, Rhaphidophoridae) from Eastern Aegean region (Southern Sporades), including Western Turkey, is reported. This brings to a total of 11 the number of Dolichopoda species recorded for caves of the Aegean area. Overall, these species show a high degree of morphological homogeneity and they are very close to Dolichopoda paraskevi Boudou-Saltet, 1973 from Crete and Dolichopoda naxia Boudou-Saltet, 1972 from Cyclades (Naxos Island). The Western Turkish species are morphologically not closely related to the other Anatolian species; this suggests an independent origin for the taxa occurring in the Southern Taurus and Black Sea regions. These new data help to better define the already high level of diversity of the Hellenic Dolichopoda and strengthen the hypothesis that the central area of dispersal for the genus would correspond to the ancient Aegean plate.
Cave crickets, Dolichopoda, new species, Eastern Aegean, Western Turkey
The subfamily Dolichopodainae Brunner von Wattenwyl, 1888 (Orthoptera, Rhaphidophoridae) is limited to the Northern hemisphere; several species belonging to the genus Dolichopoda Bolivar, 1880 inhabit caves of Southern Europe and Asia Minor. To date the genus includes a total 48 species (
In recent years several Dolichopoda were collected by us for the first time in caves of the Southern Sporades and Aegean Turkey. At a first examination these new populations did not show remarkable morphological differences resulting all very similar and close to one other. However, a genetic study based on mitochondrial DNA (mtDNA) revealed a noticeable level of genetic divergence, comparable to that usually found among morphologically divergent species of the genus (
All these new species are attributable to the sub-genus Dolichopoda s. str. (
The material was brought together by M.S. Taylan, M. Rampini and C. Di Russo for collective study and deposition in: Museum of Zoology, University of Rome “La Sapienza” (MZUR); Akdeniz University, Science Institute, Biology Department, Antalya, Turkey (AUZM). The localities of the studied species are presented on the distribution map shown in Figure 22.
Systematicsurn:lsid:zoobank.org:act:92E36710-6542-4B84-A645-F34534D3D40E
http://species-id.net/wiki/Dolichopoda_sutini
Figures 1–5The Sütini cave is situated on the road from Selçuk to Sirince (İzmir Province, Turkey).
Thirty-three specimens.
Holotype male. Turkey, İzmir, Selçuk, Sütini cave, 27.06.2008, M.S. Taylan leg. (AUZM).
Paratypes: 5 males, 5 females, 1 nymph, same data and collector as for holotype; 1 male, 12 females, 8 nymphs, Turkey, Aydın, Söke, Aşıkali cave, 13.07.2009, M.S. Taylan leg. (AUZM).
The size is relatively small with the hind legs strongly elongated. This species is close to Aegean species Dolichopoda naxia from Naxos, and Dolichopoda paraskevi from Crete but differs from them for the number of spines on the hind tibia (19); these are 25 in Dolichopoda naxia and 16 in Dolichopoda paraskevi. The tenth tergite differs from that of Dolichopoda naxia and Dolichopoda paraskevi for the shape and size of the lateral lobes and for the deeper medial incision. The male subgenital plate appears close to that of Dolichopoda paraskevi while it differs from Dolichopoda naxia; here the lateral lobes are trapezoidal. The median process of the epiphallus, flattened and rather enlarged at the base, is very close to that of Dolichopoda paraskevi, while in Dolichopoda naxia it is narrow and more elongated. From a lateral view the median process differs from that of the other two species being more curved. The female subgenital plate, with rounded lobes, is similar to that of Dolichopoda paraskevi but differs from Dolichopoda naxia for the deeper medial incision between lobes. The ovipositor, similar in length and shape, is different only for the number of denticles on the inferior valves.
Male (holotype). Body colour pale-testaceous, uniform with the exception of the posterior margins of the tergites, which are darker. Legs long, slender and yellow-testaceous in colour with the femora unarmed. Fore tibia armed with 4/5 spines on both sides of the inferior edge and a pair of spurs of equal length on the apex. Mid tibia with 3/4 short spines on both sides of the upper edge, 4/5 spines on the lower edge and two apical spurs similar to those of the fore tibia. The hind tibia is longer with 18/19 spines of varying length on both sides of the upper edge and 1/3 homogeneous spines on the lower edge. Tenth tergite on the posterior edge with two prominent lateral lobes triangular in shape with rounded apex separated from one another by a deep median incision (Figure 1). Subgenital plate globular at the bottom, with a deep median incision that runs for half of the total length; the symmetrical lateral lobes, triangular in shape, hold two evident styli cylindrical in shape (Figure 2). The epiphallus is sclerotized and shows a median process relatively long, almost flattened and with an acute apex; it appears large at the base and without lateral constrictions; from the side, the median process is uniformly thickened and curved; the basal processes are rather developed and slightly divergent (Figure 3 a, b). The accessory apparatus is sclerotized and composed by an uneven Y shaped piece, showing strong spines at the base, and by even partially trapezoidal valves.
Length(mm): body 14.0; pronotum 3.0; fore femora 16.2; middle femora 16.5; hind femora 22.4; fore tibia 16.8; middle tibia 17.3; hind tibia 30.2; hind tarsus 10.2; 1st article of hind tarsus 5.0.
Female. The length of the body ranges between 14.8–15.9 mm (ovipositor excluded) and the general form of the female is similar to the male. The subgenital plate is triangular (Figure 4), with two prominent lobes deeply incised and rounded at the posterior edges. The ovipositor has an average length of 10 mm, rather enlarged at the base and little curved on the superior edge (Figure 5).
The superior valves have a pointed apex and curves upwards, whereas the inferior valves are a little shorter than the superior ones, are rounded at the apex and have 16–17 denticles.
The new species name refers to the Sütini cave in Selçuk (İzmir Province).
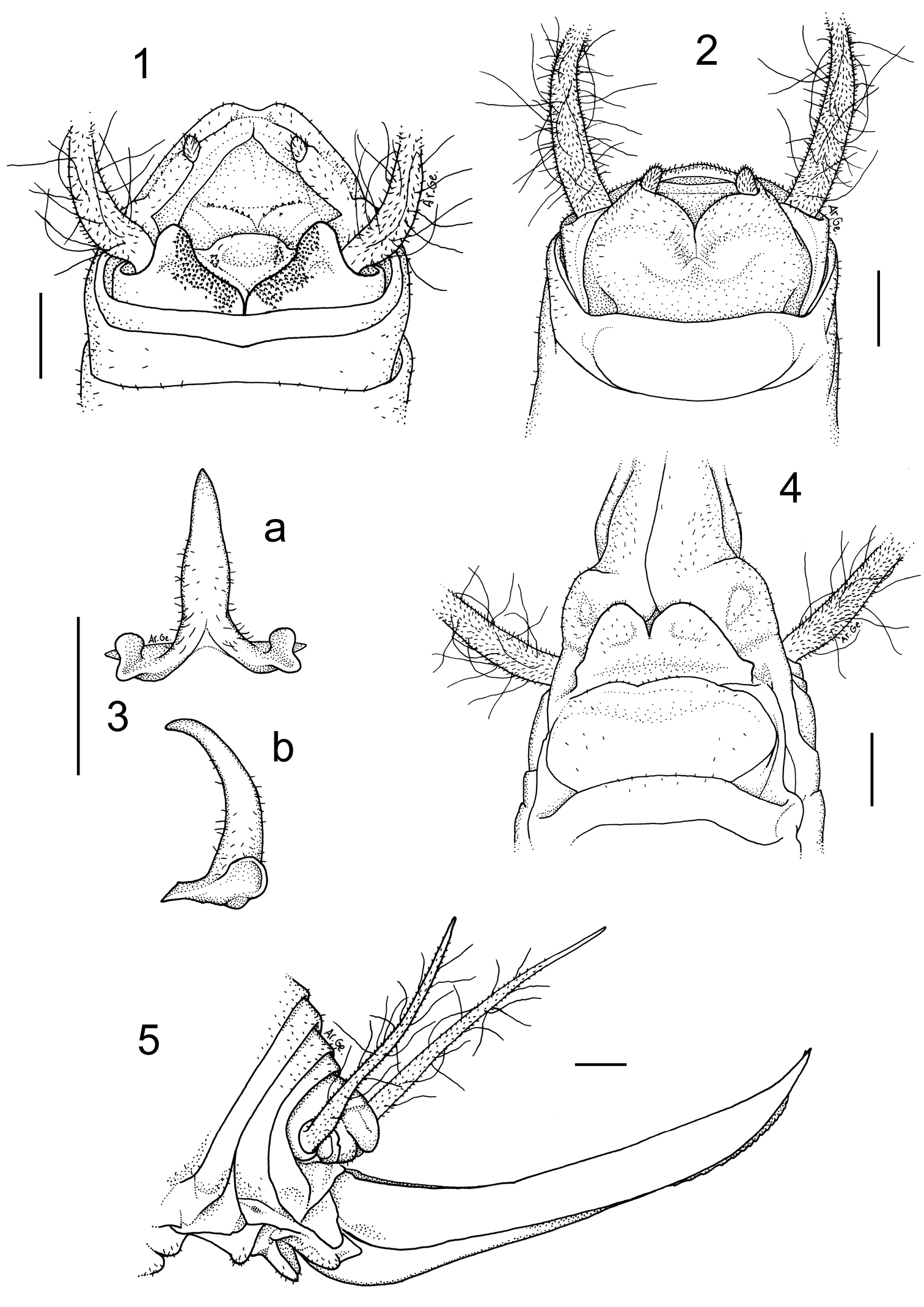
Dolichopoda sutini sp. n. Holotype male, 1 X tergite, dorsal view 2 subgenital plate, ventral view 3 epiphallus: a- dorsal view, b- lateral view. Female 4 subgenital plate, ventral view 5 ovipositor, lateral view. Scale bars: 1 mm.
urn:lsid:zoobank.org:act:CAEFB8D0-2038-48CE-912B-F8FBDA87CB85
http://species-id.net/wiki/Dolichopoda_giulianae
Figures 6–10In the North-West part of Pythagorion (Samos Island) is located the monastery of Panaghia Spiliani; here, 95 steps lead down into a big cave with a church that is dedicated to the Virgin Mary. The caves was originally used to extract blocks of massive limestone to build walls and many buildings for the town of Samos.
Sixteen specimens.
Holotype male, Greece, Samos Isl., Pythagorion, Panaghia Spiliani cave, 04.04.2008, M. Rampini, C. Di Russo leg. (MZUR).
Paratypes: 4 males, 2 females, 3 nymphs, same locality date and collectors as holotype (MZUR). 1 female, 5 nymphs, same locality as holotype 21.08.2002, F. Gasparo leg. (MZUR).
The overall appearance of this new species is very similar to that Dolichopoda sutini from İzmir. Differences are found in the squared and divergent lobes of the tenth male tergite, in the shape of the epiphallus (less enlarged at the base) and in the shorter styli of the subgenital plate. The female subgenital plate is similar to that of Dolichopoda paraskevi but differs from Dolichopoda sutini for the more rounded lobes. The ovipositor has fewer apex denticles on the inferior valves.
Male (holotype). Size relatively small. Body and appendages coloration as in the previous species. Femora unarmed. Fore tibia armed with 3 spines on both sides of the upper edge and 3/5 spines on the lower edges. Mid tibia with 5 short spines on both sides of the upper edge, 4 spines on the lower edge. The hind tibia is longer with 19/22 spines of varying length on both sides of the upper edge and 0/2 homogeneous spines on the lower edge. The tenth tergite shows two prominent lobes on the posterior edge, almost squared at the apex (Figure 6). The subgenital plate, globular at the bottom, is similar to that of Dolichopoda sutini. The lateral lobes, triangular in shape, hold two short conical styli (Figure 7). The epiphallus is sclerotized and shows a long flattened median process, acute at the apex; from the side, it appears uniformly curved (Figure 8 a, b). The accessory apparatus looks similar to that of Dolichopoda sutini.
Lenght (mm): body 14.5; pronotum 3.5; fore femora 14.0; middle femora 14.0; hind femora 22.0; fore tibia 15.5; middle tibia 16.5; hind tibia 28.0; hind tarsus 11.0; 1st segment of hind tarsus 5.5.
Female. General appearance as in the male. The length of the body ranges between 15.0 and 18.0 mm (ovipositor excluded). The subgenital plate is trapezoidal with two rounded lobes (Figure 9). The ovipositor has an average length of 11.0 mm, is rather enlarged at the base and is regularly curved on the superior edge (Figure 10); the apex is pointed and curved upwards. The inferior valves have 15 apical denticles.
The new species is dedicated to our colleague and friend Giuliana Allegrucci for her useful and active collaboration in this study.
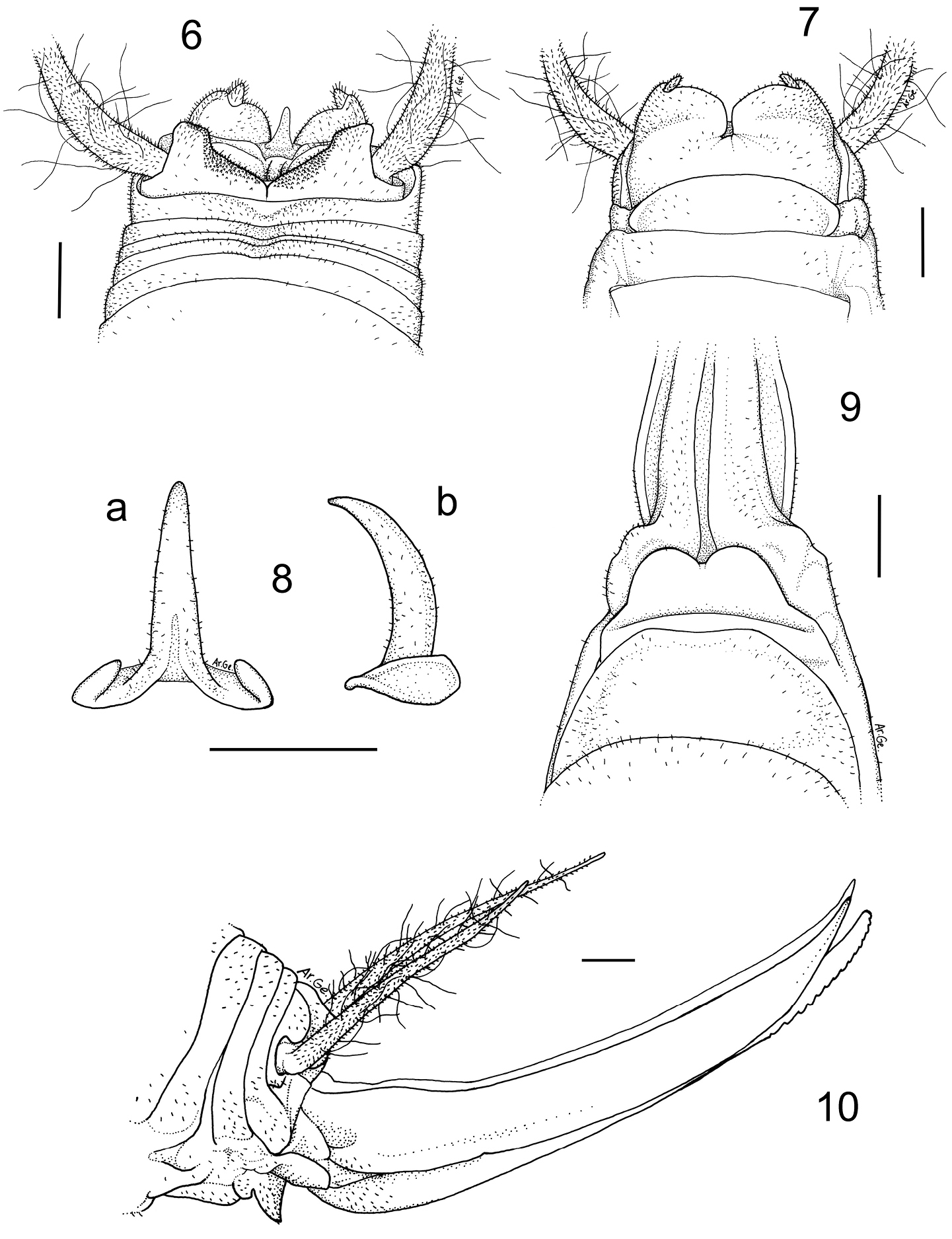
Dolichopoda giulianae sp. n. Holotype male 6 X tergite, dorsal view 7 subgenital plate, ventral view 8 epiphallus, a dorsal view b lateral view. Female 9 subgenital plate, ventral view 10 ovipositor, lateral view. Scale bars: 1 mm.
urn:lsid:zoobank.org:act:E82231B2-8FF8-4ED5-BE7D-59B803BEBDDF
http://species-id.net/wiki/Dolichopoda_kalithea
Figures 11–15The cave, with a chapel inside, is located East of Kalithea village in the Western part of Mount Kerkis, inside the canyon Kakoperato, which starts from a little monastery called Panaghias Kakoperato. The cave is known for the presence of the endemic Staphylinid rove beetle Tychobythinus brachati Besuchet, 2008.
Sixteen specimens.
Holotype male, Greece, Samos Isl., Mount Kerkis, Kakoperato canyon, 660 m, Kakoperato cave (or Trypa Tse-Tse cave), 05.04.2008, C. Di Russo, M. Rampini leg.
Paratypes: 6 males, 1 male, 2 nymphs, same locality, date and collectors (MZUR). South-Eastern slopes of Mount Kerkis, Marathokampos, 320 m, Sarandaskaliotissa cave (near Pythagoras cave), 1 male, 2 females, 3 nymphs, 05.04.2008, C. Di Russo, M. Rampini leg. (MZUR).
Shape and coloration as in the previous species. Size relatively large, with very long legs. This species, owing to the triangular lobes of the tenth tergite (almost fully separated by a large concavity) and to the trapezoidal lobes of the subgenital plate is close to Dolichopoda naxia. On the contrary the pronounced curve of the median process of the epiphallus differs markedly from that of Dolichopoda naxia while it resembles that of Dolichopoda sutini and of Dolichopoda giulianae. The female subgenital plate differs from that of both Dolichopoda sutini and Dolichopoda giulianae for the lower incision between the two lobes.
Male (holotype). Body and appendages coloration as in the previous species. Femora unarmed. Fore tibia armed with 1/4 spines on both sides of the upper edge and 3/5 spines on the lower edges. Mid tibia with 1/4 short spines on both sides of the upper edge, 4 spines on the lower edge. The hind tibia is longer with 13/20 spines of varying length on both sides of the upper edge and 0/3 homogeneous spines on the lower edge. The tenth tergite has two triangular lobes quite developed and separated by a large concavity (Figure 11). The subgenital plate shows two trapezoidal lobes, straight on the posterior edges and separated by a relatively short incision (Figure 12); the lobes hold two prominent cylindrical styli. The epiphallus is sclerotized and shows a quite flattened median process with an enlarged base; laterally, it appears rather thick at the base and strongly arched distally (Figure 13). The accessory apparatus is similar to that of the previous species.
Length(mm): body 16.0; pronotum 3.0; fore femora 13.0; middle femora 14.0; hind femora 21.5; fore tibia 15.5; middle tibia 16.5; hind tibia 27.0; hind tarsus 10.5; 1st article of hind tarsus 5.0.
Female. General appearance as in the male. The length of the body ranges between 19.0 and 21.0 mm (ovipositor excluded). The subgenital plate is rounded and slightly incised in the middle (Figure 14). The ovipositor has an average length of 12.0 mm, rather enlarged at the base and regularly curved on the superior edge (Figure 15). The superior valves have a pointed apex and curves upwards, the inferior valves have 14 denticles .
The new species takes its name from the Kalithea village.
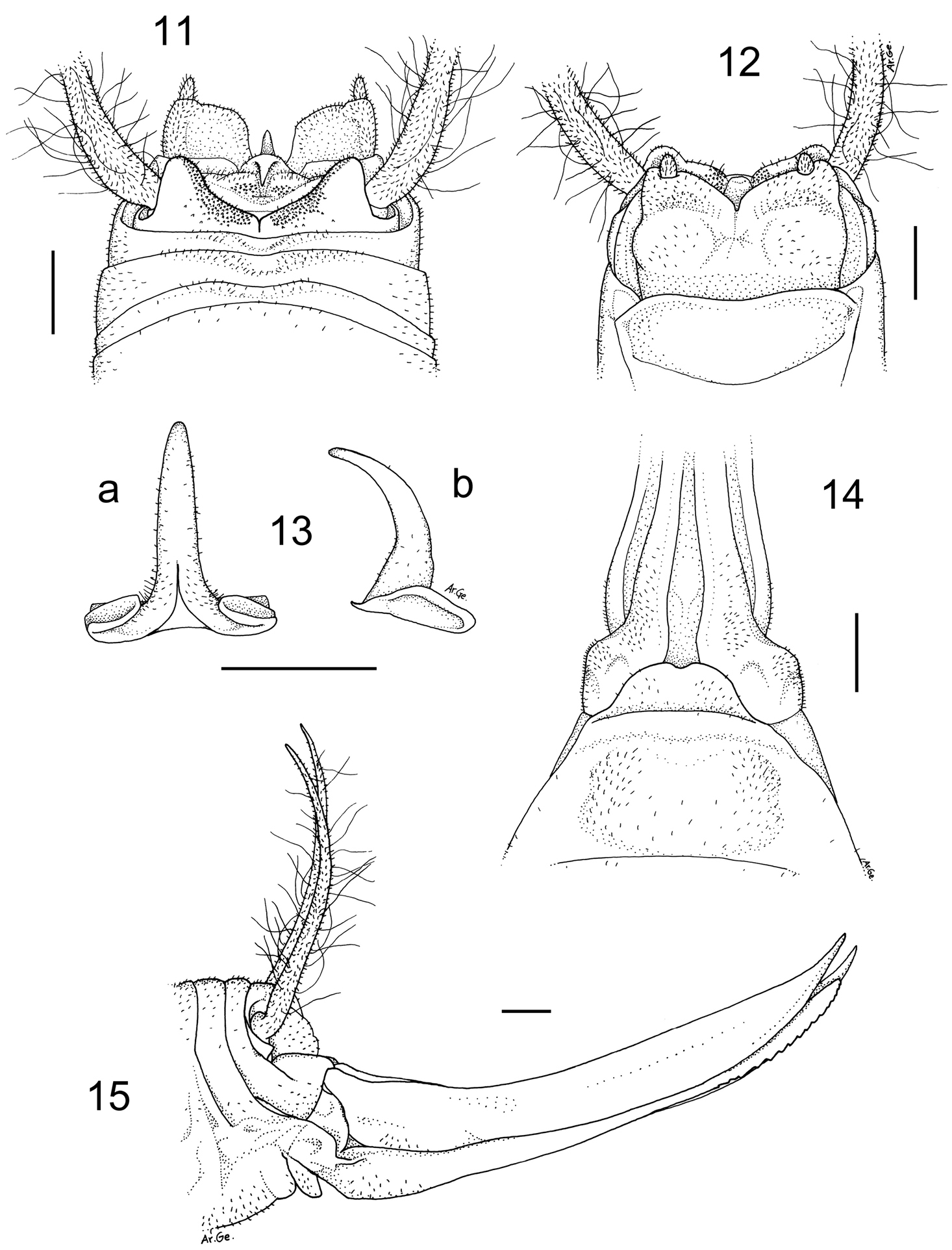
Dolichopoda kalithea sp. n. Holotype male, 11 X tergite, dorsal view 12 subgenital plate, ventral view 13 epiphallus a dorsal view b lateral view. Female 14 subgenital plate, ventral view 15 ovipositor, lateral view. Scale bars: 1 mm.
urn:lsid:zoobank.org:act:58C66299-D839-4EEC-852C-D11AC9E3E78F
http://species-id.net/wiki/Dolichopoda_calidnae
Figures 16–21Foot of the Mt. Flaska, in the cave of the Nymphs, also called the cave of the Seven Virgins.
Fifteen specimens.
Holotype male, Greece, Kalymnos Isl., Pothia, Seven Virgins cave (Sanctuary of the Nimphs or Epta Parthenes cave), 28.03.2004, M. Rampini, C. Di Russo leg.
Paratypes: 1 male, 5 females, same data and collectors; Skalia village, Skalia cave (Mts Flaska), 3 males, 5 nymphs, 28.03.2004, M. Rampini, C. Di Russo leg. (MZUR).
Colour of the body uniformly pale-testaceous, legs more yellowish. The male tenth tergite shows expanded lateral lobes of triangular shape with an acute apex. The subgenital plate, with trapezoidal lobes, is similar to that of Dolichopoda naxia but with short apical styli on the posterior edge. The epiphallus is long and slender as in Dolichopoda naxia, with a little curved median process, stretched and narrower than in the previous species. The female subgenital plate is morphologically different from that of all the other species showing a triangular shape with two moderately incised lobes in the middle.
Male (holotype). Size rather large. Coloration uniformly pale-testaceus. Legs long and more light in colour. Femora unarmed. Fore tibia armed with 5 spines on both sides of the inferior edge and 3/4 spines on the lower edges. Mid-tibia with 4 short spines on both sides of the upper edge, 4/5 spines on the lower edge. The hind tibia is longer with 21/24 spines of varying length on both sides of the upper edge and 2/3 homogeneous spines on the lower edge. Eighth and ninth abdominal tergites show a sinuous posterior edge, the eighth one is hollower centrally. The tenth tergite, similar to those of the previous species, shows on the posterior edge two large lateral lobes, triangular in shape, with rather rounded apex (Figure 16). Subgenital plate globular at the bottom, with a deep middle incision that runs for half of the total length (Figure 17). Lateral lobes trapezoidal, with two short conical styli. The epiphallus is sclerotized and shows a median process relatively long, almost cylindrical and acute apically. From the side, it appears large at the base and uniformly curved. The basal processes are squared, rather developed and slightly divergent (Figure 18 a, b). The accessory apparatus is similar to that of the previous species (see photo in Figure 21 a, b).
Length (mm): body 17.5; pronotum 3.5; fore femora 14.3; middle femora 14.5; hind femora 22.5; fore tibia 15.9; middle tibia 16.7; hind tibia 26.9; hind tarsus 11.0; 1st article of hind tarsus 5.5.
Female. General appearance as in the male. The length of the body ranges between 16.0–17.0 mm (ovipositor excluded). The subgenital plate is triangular with two moderately incised lobes in the middle (Figure 19). The ovipositor has an average length of 11.0 mm, it is enlarged at the base and regularly curved on the superior edge (Figure 20). The superior valves have a pointed apex and curves upwards, whereas the inferior valves are rounded apically and have 15 denticles.
The new species takes its name from the Calidnae Islands (Kalymnos, Leros and Telendos) cited by Homer in Iliad.
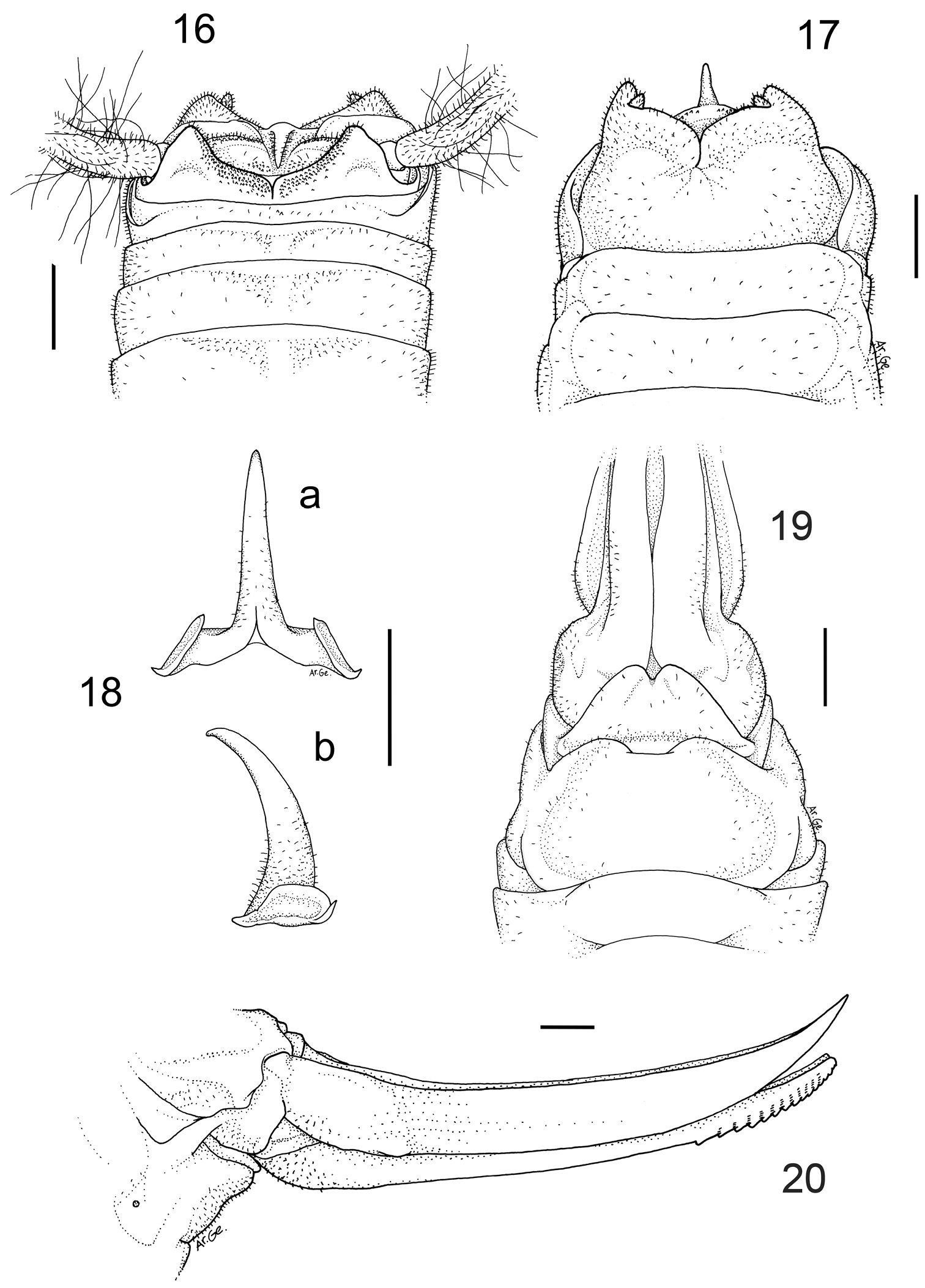
Dolichopoda calidnae sp. n. Holotype male 16 X tergite, dorsal view 17 subgenital plate, ventral view 18 epiphallus a dorsal view b lateral view. Female 19 subgenital plate, ventral view 20 ovipositor, lateral view. Scale bars: 1 mm
Here we describe three new species belonging to the cave cricket genus Dolichopoda from the Aegean Greek Islands (Southern Sporades) and one from the Western Turkish coast. Considering the additional seven species already reported for the area, there is now a total of 11 recorded species of Dolichopoda that currently inhabit caves of the region (Figure 22). These new data, therefore, document the high diversity of the genus in the Hellenic region (25 species in total) reinforcing the hypothesis of a central area of dispersal of Dolichopoda corresponding to the ancient Aegean plate (
The new species are all morphologically homogeneous, due to a strong similarity of most of the characters examined (i.e. tenth tergite, epiphallus, accessory apparatus and female subgenital plate). Based on these characters the new species show a clear affinity with Dolichopoda naxia from the Cyclades and with Dolichopoda paraskevi from Crete (see Table 1). On the other hand, they are well separated from Dolichopoda thasosensis, anendemic taxon of the Thasos Island (Tracia), showing a peculiar trilobate shape of the tenth tergite, and from the western species from Eubea and Skyros. These latter species are characterized by a bifurcate apex of the epiphallus and for this reason
The strong morphological homogeneity of this Eastern Aegean species complex, whose range extends now from the Aegean coast of Turkey to the caves of Eastern Crete, clearly suggests a common origin from an ancestor that presumably occupied the eastern part of the ancient Aegean plate. As reported by
Of particular interest is the situation on Samos Island, where two species (Dolichopoda kalithea and Dolichopoda giulianae) co-occurr. The differentiation of these two closely related species could be explained by the different geological origin of their cave habitats that could have acted as a geographic and/or ecological barrier preventing gene flow. Kakoperato and Sarandaskaliotissa caves, inhabited by Dolichopoda kalithea, open to the Eastern slopes of Mount Kerkis, a massif of dolomitic marble of Cretaceous origin. On the other hand, the Panaghia Spiliani cave, where Dolichopoda giulianae lives, is placed in a lacustrine limestone formation of Neogene origin (
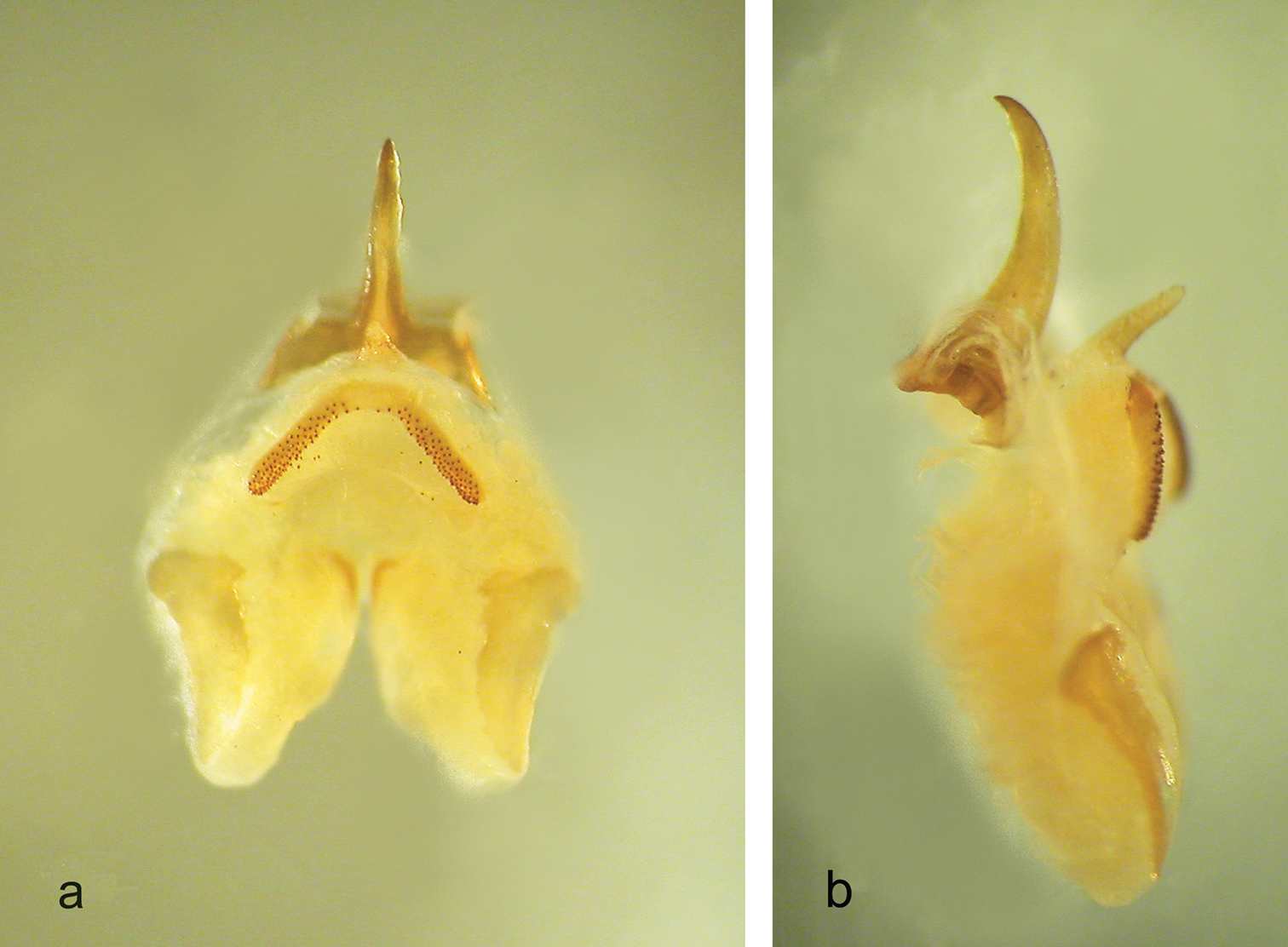
Dolichopoda calidnae sp. n., holotype male, epiphallus and accessory apparatus a dorsal view b lateral view.
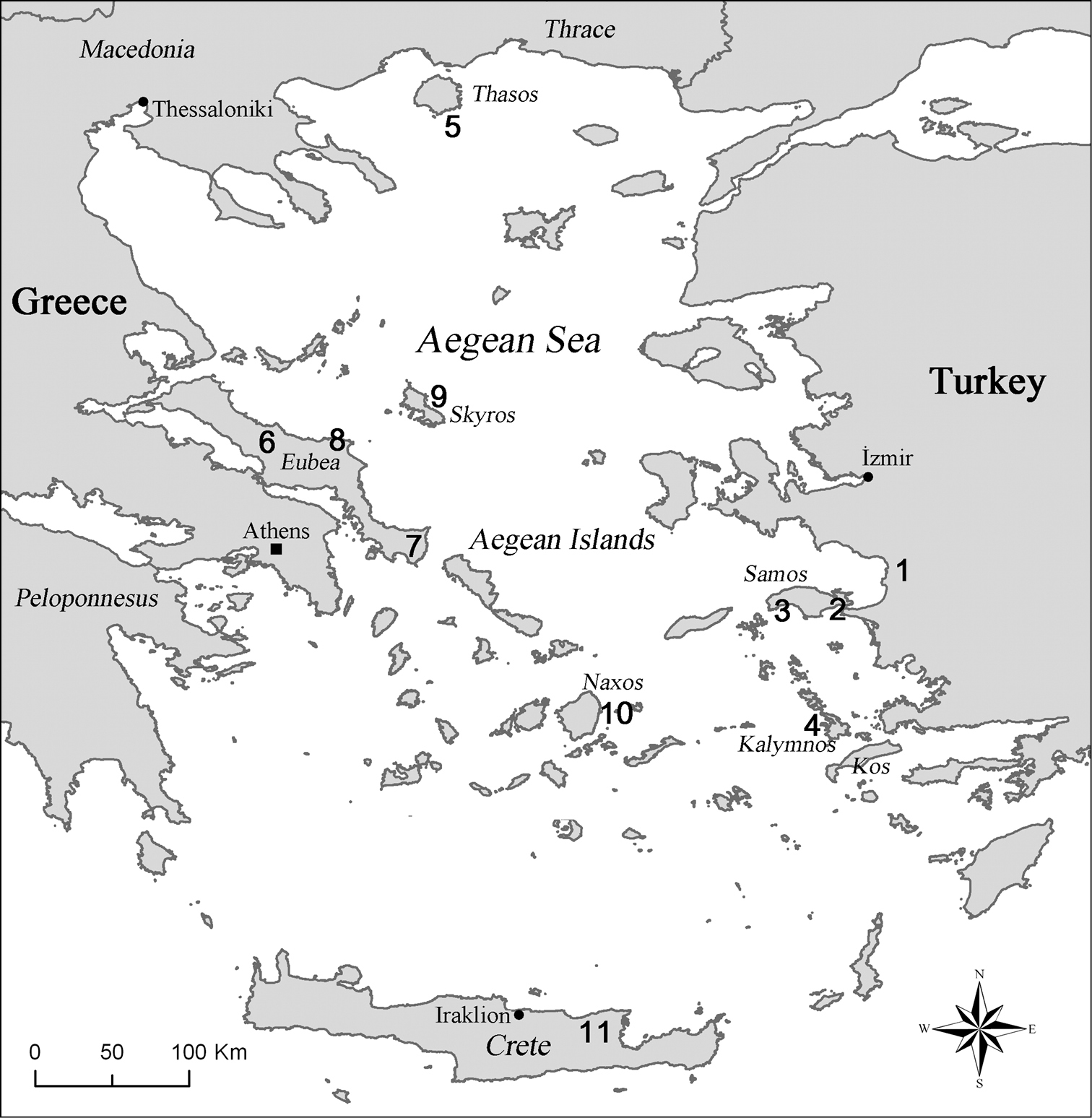
Distribution of troglophilous species of Dolichopoda in Aegean Region. 1 Dolichopoda sutini sp. n. (Sütini cave, Selçuk, İzmir) 2 Dolichopoda giulianae sp. n. (Panaghia Spiliani cave, Pythagorion, Samos) 3 Dolichopoda kalithea sp. n. (Kakoperato cave, Kalithea, M. Kerkis, Samos) 4 Dolichopoda calidnae sp. n. (Seven Virgins cave, Pothia, Kalymnos) 5 Dolichopoda thasosensis (Drakotrypa cave, Thasos) 6 Dolichopoda makrykapa (Piyi Nyphi cave, Makrykapa, Eubea) 7 Dolichopoda cassagnaui (Aghia Trias cave, Karystos, Eubea) 8 Dolichopoda ochtoniai (unnamed cave at North Est of Eubea) 9 Dolichopoda saraolakosi (Lynaria caves, Skyros) 10 Dolichopoda naxia (Za cave, Filotas, Naxos) 11 Dolichopoda paraskevi (Aghia Paraskevi cave, Skotino, Iraklion, Crete).
Mean values of 11 morphological parameters of the Aegean Dolichopoda species here studied (dimension in mm).
| Dolichopoda sutini | Dolichopoda calidnae | Dolichopoda giulianae | Dolichopoda kalithea | Dolichopoda nxia1 | Dolichopoda paraskevi2 | |
|---|---|---|---|---|---|---|
| Body | 13.50 | 18.00 | 15.00 | 16.50 | 19.00 | 14.5 |
| Pronotum | 3.00 | 3.50 | 3.50 | 3.00 | 4.00 | 3.00 |
| 1 Femora | 16.00 | 14.00 | 14.00 | 13.00 | 16.00 | 14.5 |
| 2 Femora | 15.00 | 15.00 | 14.00 | 13.50 | 16, 00 | 14.00 |
| 3 Femora | 22.00 | 23.00 | 22.00 | 21.00 | 25.00 | 23.50 |
| 1 Tibia | 17.00 | 16.00 | 16.00 | 15.00 | 17.00 | 17.00 |
| 2 Tibia | 16.50 | 17.00 | 17.00 | 16.00 | 18.00 | 17.00 |
| 3 Tibia | 30.00 | 30.00 | 29.00 | 27.00 | 33.00 | 30.50 |
| Hind tarsus | 10.00 | 11.00 | 11.00 | 10.00 | 11.50 | 10.50 |
| I°art.h.tarsus | 5.00 | 5.50 | 5.50 | 5.00 | 5.50 | 5.50 |
| Ovipositor | 10.00 | 11.00 | 11.00 | 12.00 | 11.50 | 11.50 |
1
We thank Mehmet Öz for his advices on this study and Deniz Şirin and DEUMAK (Dokuz Eylul University Cave Research Group) for his help in collecting cave samples. For the revision of English and critical reading of parts of the manuscript we thank Dr. Valerio Ketmaier (University of Potsdam, Germany and University of Rome “ La Sapienza”, Italy). This study includes data of the Project no° 2008.03.0121.002 which was supported by Akdeniz University Scientific Research Fund (Prof. M. Öz). The expeditions for collecting the cave crickets on the Aegean Islands (Samos and Kalymnos) were supported by the University of Rome ‘La Sapienza’ – Research Project 2008, n° C26F085LN7.