





(C) 2012 Ricardo Pinto-da-Rocha. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
As part of an ongoing revision of the family Gonyleptidae, we have identified many species that are synonyms of previously described species or misplaced in this family. This article summarizes these findings, adding previously unavailable information or correcting imprecise observations to justify the presented taxonomic changes.
The following new familial or subfamilial assignments are proposed: Nemastygnus Roewer, 1929 and Taulisa Roewer, 1956 are transferred to Agoristenidae, Agoristeninae; Napostygnus Roewer, 1929 to Cranaidae; Ceropachylinus peruvianus Roewer, 1956 and Pirunipygus Roewer, 1936 are transferred to Gonyleptidae, Ampycinae; Gyndesops Roewer, 1943, Haversia Roewer, 1913 and Oxapampeus Roewer, 1963 are transferred to Gonyleptidae, Pachylinae.
The following generic synonymies are proposed for the family Gonyleptidae: Acanthogonyleptes Mello-Leitão, 1922 = Centroleptes Roewer, 1943; Acrographinotus Roewer, 1929 = Unduavius Roewer, 1929; Gonyleptes Kirby, 1819 = Collonychium Bertkau, 1880; Mischonyx Bertkau, 1880 = Eugonyleptes Roewer, 1913 and Gonazula Roewer, 1930; Parampheres Roewer, 1913 = Metapachyloides Roewer, 1917; Pseudopucrolia Roewer, 1912 = Meteusarcus Roewer, 1913; Haversia Roewer, 1913 = Hoggellula Roewer, 1930.
The following specific synonymies are proposed for the family Gonyleptidae: Acanthogonyleptes singularis (Mello-Leitão, 1935) = Centroleptes flavus Roewer, 1943, syn. n.; Geraeocormobius sylvarum Holmberg, 1887 = Discocyrtus serrifemur Roewer, 1943, syn. n.; Gonyleptellus bimaculatus (Sørensen, 1884) = Gonyleptes cancellatus Roewer, 1917, syn. n.; Gonyleptes atrus Mello-Leitão, 1923 = Weyhia brieni Giltay, 1928, syn. n.; Gonyleptes fragilis Mello-Leitão, 1923 = Gonyleptes banana Kury, 2003, syn. n.; Gonyleptes horridus Kirby, 1819 = Collonychium bicuspidatum Bertkau, 1880, syn. n., Gonyleptes borgmeyeri Mello-Leitão, 1932, syn. n., Gonyleptes curvicornis Mello-Leitão, 1932, syn. n., Metagonyleptes hamatus Roewer, 1913, syn. n. and Paragonyleptes simoni Roewer, 1930, syn. n.; Gonyleptes pustulatus Sørensen, 1884 = Gonyleptes guttatus Roewer, 1917, syn. n.; Haversia defensa (Butler, 1876) = Sadocus vallentini Hogg, 1913, syn. n.; Liogonyleptoides minensis (Piza, 1946) = Currala bahiensis Soares, 1972, syn. n.; Megapachylus grandis Roewer, 1913 = Metapachyloides almeidai Soares & Soares, 1946, syn. n.; Mischonyx cuspidatus (Roewer, 1913) = Gonazula gibbosa Roewer, 1930 syn. n.; Mischonyx scaber (Kirby, 1819) = Xundarava holacantha Mello-Leitão, 1927, syn. n.; Parampheres tibialis Roewer, 1917 = Metapachyloides rugosus Roewer, 1917, syn. n.; Parapachyloides uncinatus (Sørensen, 1879) = Goyazella armata Mello-Leitão, 1931, syn. n.; Pseudopucrolia mutica (Perty, 1833) = Meteusarcus armatus Roewer, 1913, syn. n.
The following new combinations are proposed: Acrographinotus ornatus (Roewer, 1929), comb. n. (ex Unduavius); Gonyleptellus bimaculatus (Sørensen, 1884), comb. n. (ex Gonyleptes);Gonyleptes perlatus (Mello-Leitão, 1935), comb. n. (exMoojenia);Mischonyx scaber (Kirby, 1819), comb. n. (ex Gonyleptes); and Neopachyloides peruvianus (Roewer, 1956), comb. n. (ex Ceropachylus).
The following species of Gonyleptidae, Gonyleptinae are revalidated: Gonyleptes atrus Mello-Leitão, 1923 and Gonyleptes curvicornis (Roewer, 1913).
Agoristenidae, Cranaidae, Gonyleptidae, Neotropics, taxonomy
Opiliones is currently divided into four monophyletic suborders (
From recent phylogenetic studies on the Gonyleptidae subfamilies Gonyleptinae, Pachylinae, and Metasarcinae, as well as visits to such European museums as the Senckenberg Museum (Frankfurt), Muséum National d’Histoire Naturelle (Paris), and Zoologisches Institut and Zoologisches Museum (Hamburg), we were able to precisely identify many gonyleptids, resulting in the detection of numerous synonymies and the misplacement of several species relative to modern family and subfamily concepts. Thus, we decided to publish these systematic findings in this format instead of many revisions of the respective families/subfamilies, which would take several years to finish. The aim of this article is to propose nomenclatural changes in an attempt to resolve the taxonomic nightmare in Neotropical harvestmen systematics.
Material and methodsThe following abbreviations were adopted (curators in parentheses) to refer to the depositories:
NHM The Natural History Museum [formerly British Museum], London, England (Janet Beccaloni).
HEMS Hélia Eller Monteiro Soares private collection, now included in MNRJ.
IBSP Instituto Butantan, São Paulo, São Paulo, Brazil (Darci M. Barros Battesti).
ISNB Institut Royal des Sciences Naturelles de Belgique, Brussels, Belgium (Léon Baert).
MNRJ Museu Nacional do Rio de Janeiro, Rio de Janeiro, Rio de Janeiro, Brazil (Adriano Brilhante Kury).
MZLQ Museu de Zoologia Luiz de Queiroz, now housed in IBSP.
MZSP Museu de Zoologia da Universidade de São Paulo, São Paulo, São Paulo, Brazil (Ricardo Pinto-da-Rocha).
NHMW Naturhistorisches Museum, Vienna, Austria (Jürgen Gruber).
SMF Naturmuseum Senckenberg, Frankfurt am Main, Germany (Peter Jäger).
ZMB Institut für Systematische Zoologie, Museum für Naturkunde der Humboldt-Universität zu Berlin, Germany (Jason Dunlop).
ZMH Zoologisches Museum Hamburg, Germany (Hieronymus Dastich).
ZMUC Zoologisk Museum, Universität København, Zoological Museum, University of Copenhagen, Denmark (Nikolaj Scharff).
We did not exhaustively cite literature in synonymic listings (for this purpose see
http://species-id.net/wiki/Nemastygnus
Nemastygnus was transferred from Cranaidae to Gonyleptidae, Metasarcinae by
Ocularium unarmed, saddle-shaped. Areas and posterior margin of dorsal scutum and free tergites unarmed (Fig. 1A). Scutal area I undivided. Lateral margin of dorsal scutum coriaceus. Chelicera dimorphic. Pedipalp with slender articles; femur without dorsoapical spine, with row of three ventral large setae and two ventroapical large setae; patella with prolateral large setae; tibia–tarsus with ectal-mesal large setae. Leg I filiform, three-segmented. Penis: basal setae of truncus and basal setae of ventral plate very long and bifid; stylus with a large, longitudinal keel.
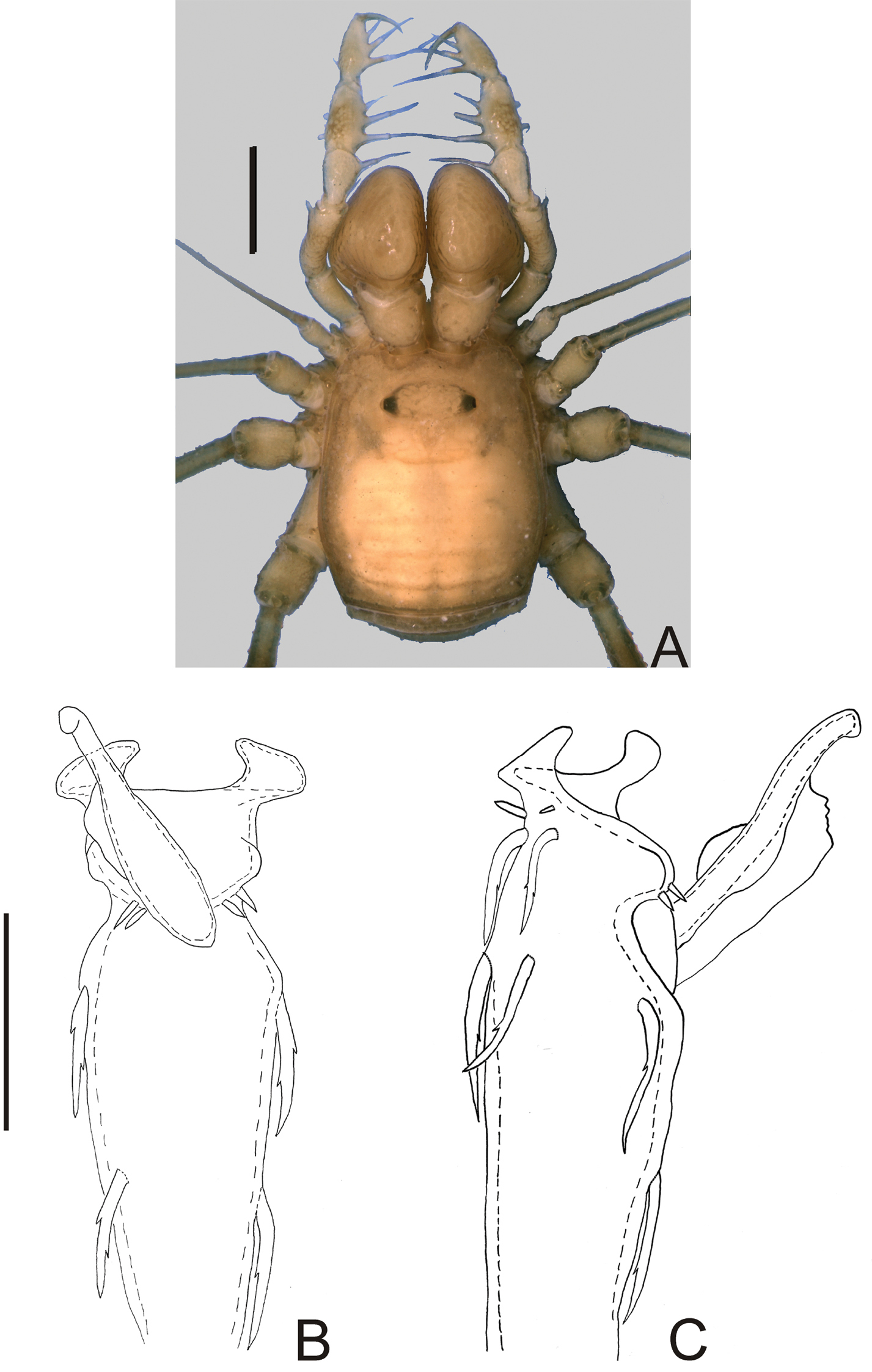
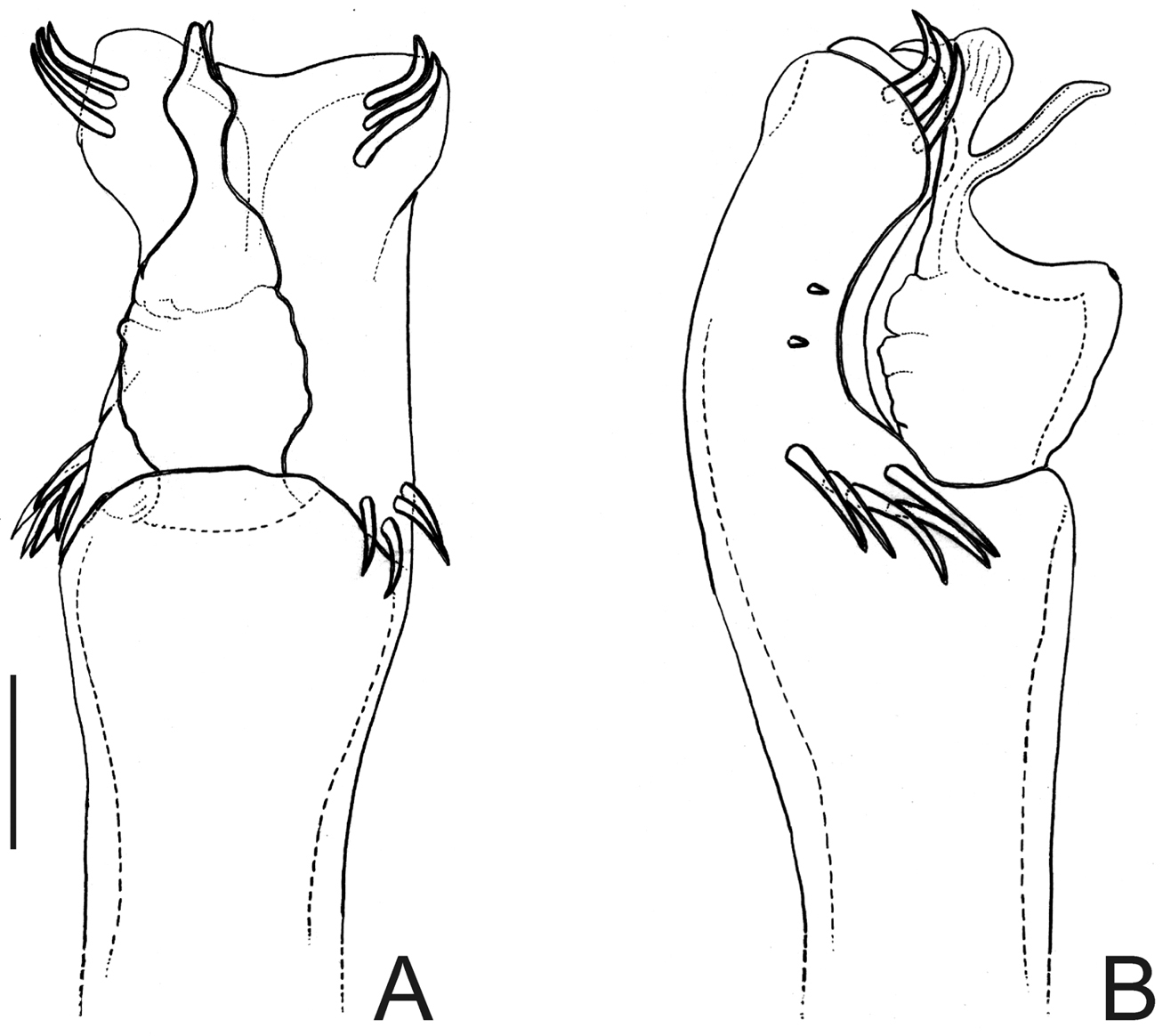
Nemastygnus ovalis Roewer. Male (holotype): A habitus, dorsal view. Distal part of penis B in dorsal view C ditto, left lateral view. B, C at same scale. Sale bars: A = 1 mm; B = 0.1 mm.
http://species-id.net/wiki/Nemastygnus_ovalis
Fig. 1Penis (Fig. 1B–C; holotype): truncus with three pairs of bifid setae (basal, subdistal lateral and distal ventral). Ventral plate with rounded lobe on corners, two dorso-basal pairs of small single branched setae, a ventral distal pair of small single branched setae and a ventral median bifid setae. Glans dorsally projected, slender, with a small ventral crest and a long dorsal crest. Stylus smooth.
Nemastygnus was originally placed in Gonyleptidae, Prostygninae.
http://species-id.net/wiki/Taulisa
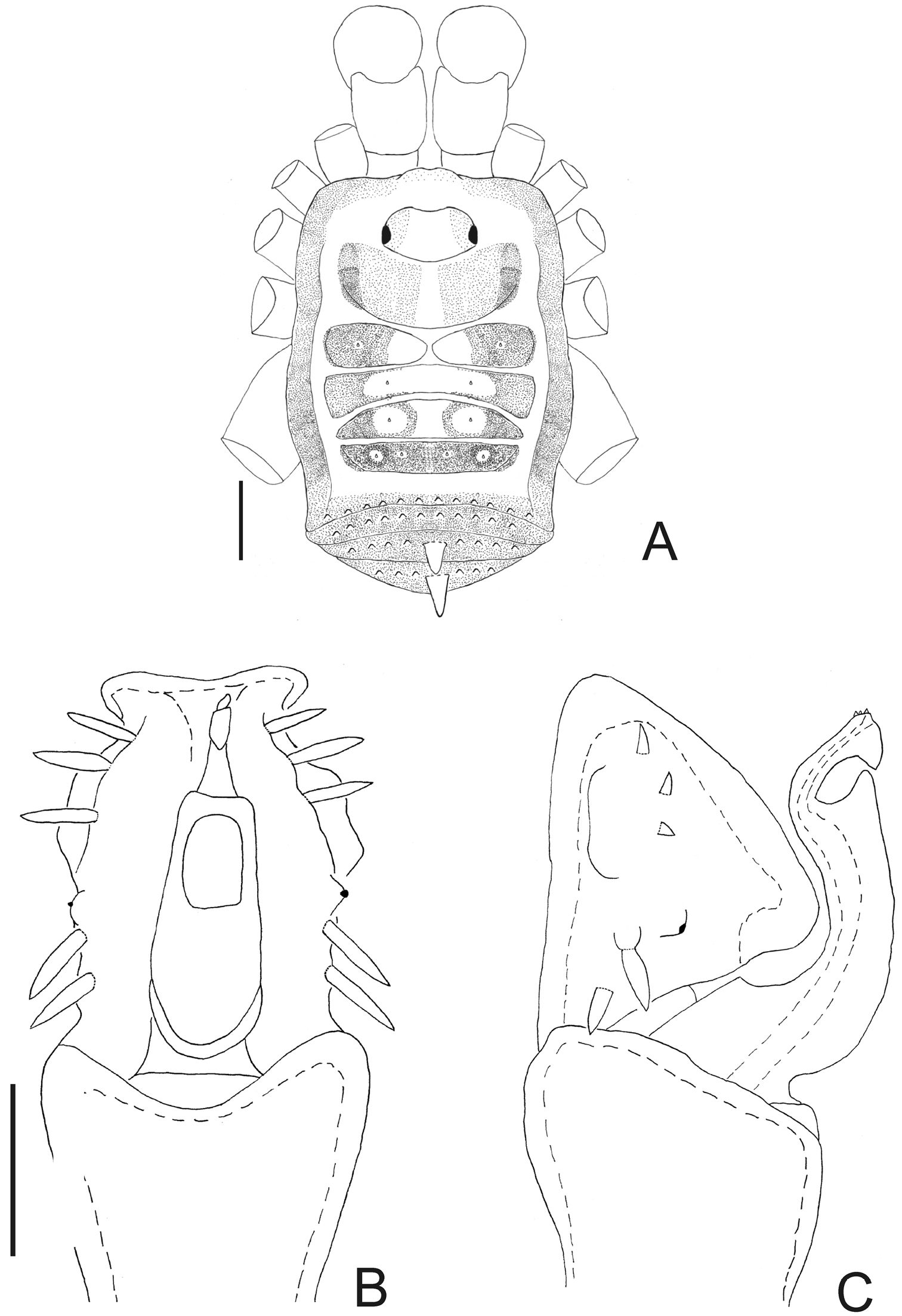
Taulisa (Fig. 2) differs from the other 10 genera of Leiosteninae by the following combination of characters: ocularium unarmed, saddle-shaped; areas of dorsal scutum with two large tubercles each, posterior margin of dorsal scutum and free tergites, each one with a central large tubercle; area I undivided; lateral margin of dorsal scutum coriaceus; pedipalp with slender articles, its femur with dorsoapical spine, with row of four ventral large setae; pedipalpal patella with prolateral large setae; pedipalpal tibia–tarsus with ectal-mesal large setae; leg I filiform, three-segmented. Male unknown.
Taulisa was originally placed in Phalangodidae, Tricommatinae.
Monotypic.

Taulisa koepckei Roewer. Female (holotype): Habitus, dorsal view. Scale bar: 1 mm.
http://species-id.net/wiki/Taulisa_koepckei
Fig. 2As for genus.
http://species-id.net/wiki/Napostygnus
Napostygnus was originally placed in Gonyleptidae, Prostygninae and transferred to the Gonyleptidae, Metasarcinae by
Monotypic.
http://species-id.net/wiki/Napostygnus_bispinosus
Fig. 3ECUADOR. Napo: Valley of Rio Napo, 1 female holotype (SMF RI 1004/3); Cantón Quijos, Parroquira Cozanga, Yanayacu Research Station (0°35'S, 78°57'W, 2128 m), 1 ♂ & 2 ♀ (MZSP 36132); ditto, 1 ♀ (IBSP 10550).
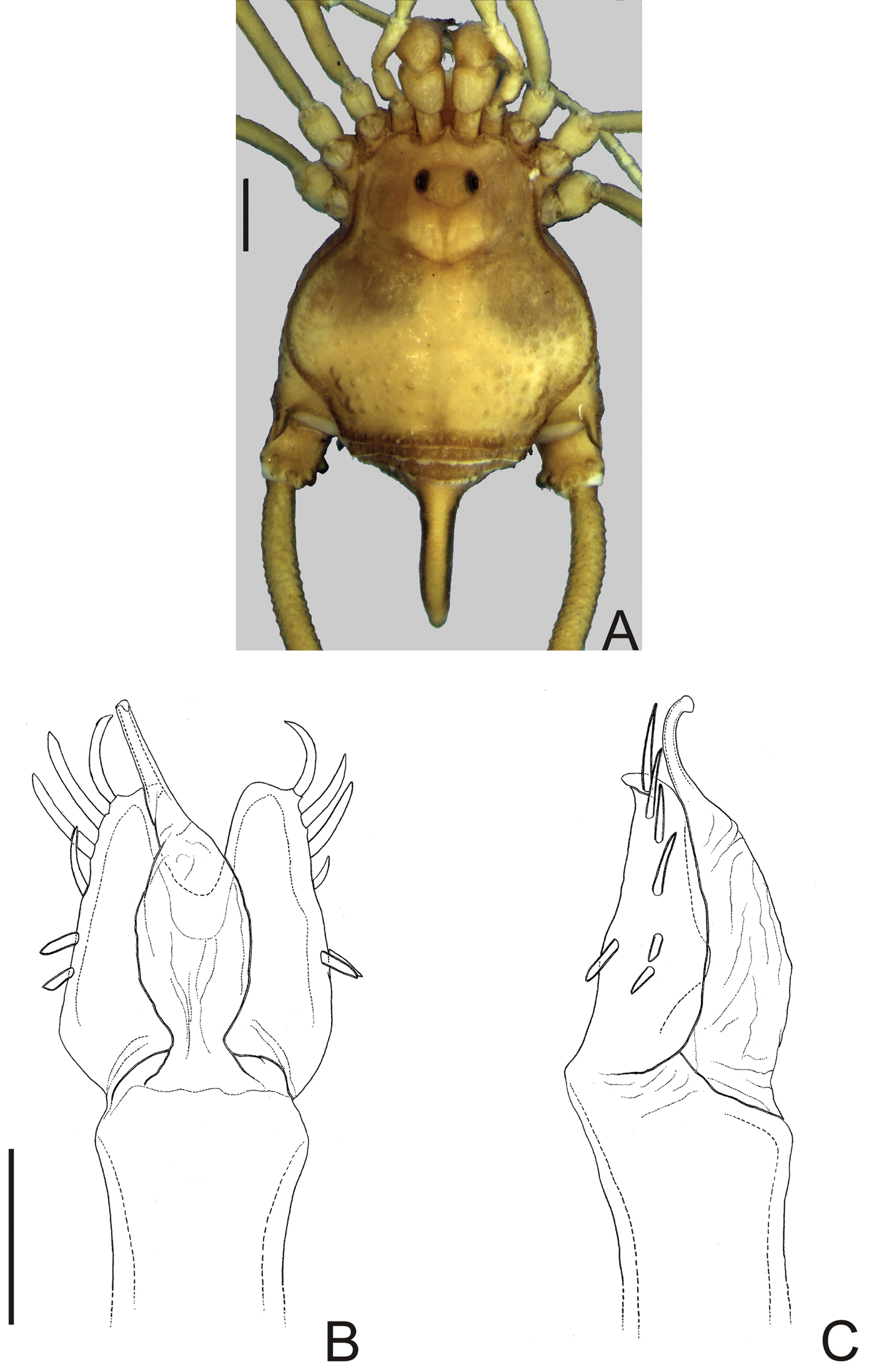
Male (MZSP 36132). Dorsum (Fig. 3A). Measurements: dorsal scutum length 4.1; dorsal scutum maximum width 3.5; carapace length 2.1; carapace maximum width 2.9; femur IV length 11.2. Body outline nearly subrectangular. Anterior margin of dorsal scutum with a median frontal hump small-granulate. Ocularium near middle of carapace, saddle shaped, with small granules near the eyes, unarmed. Carapace higher than the rest of dorsal scutum, with 4 tubercles behind ocularium. Scutal areas I–III with 2 small median tubercles on each area; IV with 4 tubercles. Lateral margin of dorsal scutum with a low density of small granules. Posterior margin of dorsal scutum and free tergite I with a row of small granules, unarmed. Free tergites II–III each with a median spine, small granulate.
Chelicera: segment I unarmed. Segment II swollen, finger II with 3 teeth, III with 4 teeth.
Pedipalpus: trochanter with 2 ventral tubercles, femur and patella smooth. Tibial setation: retrolateral iiIii; prolateral iiiIiii. Tarsal setation: retrolateral IiIi; prolateral IiiiIi.
Legs: legs I–IV unarmed and without granules, except for trochanters, which are small granulate. Basitarsus I slightly inflated. Tarsal process present. Tarsal segmentation: 6(3); 16(3); 6; 7.
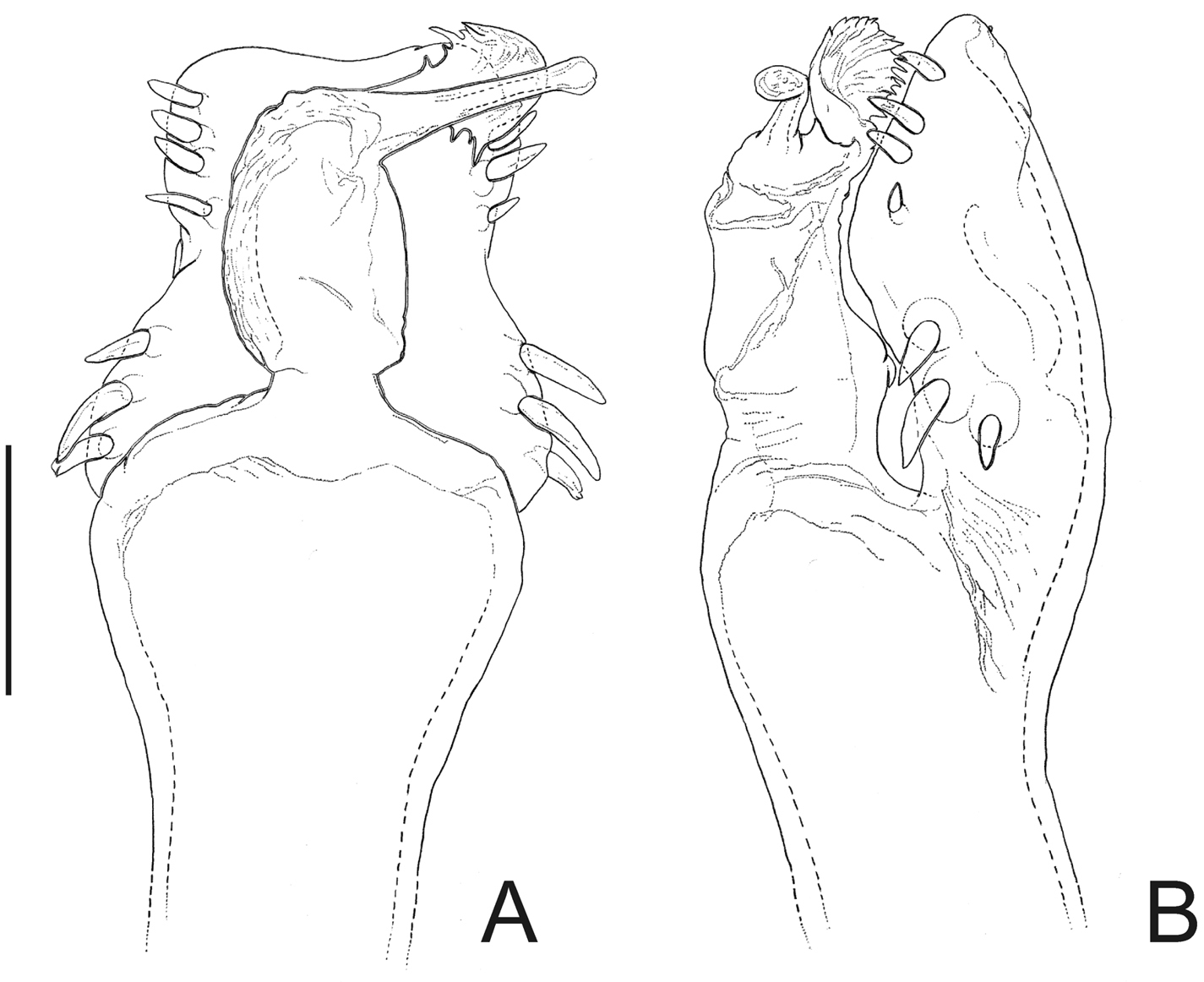
Penis (Fig. 3B–C): ventral plate with almost straight distal margin, thick median lobe and folded ventrally to the distal setae, 3 pairs of distal setae and 2 pairs of basal setae. Glans with thumb-like dorsal process. Stylus with dorsal apical projection and ventral apical small trichomes.
Coloration (in ethanol) (Fig. 3A): body background yellow with brown spots mainly on carapace, scutal areas, lateral and posterior margins of dorsal scutum and free tergites. Mesotergum with one longitudinal yellow stripe surrounded by blackish pigment at grooves I–V. Pedipalpus and chelicera yellowish brown with a brown reticulate pattern. Legs yellowish brown.
Napostygnus was originally placed in Gonyleptidae, Prostygninae.
Napostygnus bispinosus Roewer. Male (MZSP 36132): A habitus, dorsal view. Distal part of penis B in dorsal view C ditto, left lateral view. B, C at same scale. Scale bar A = 1 mm. Scale bar B = 0.1 mm.
http://species-id.net/wiki/Neopachyloides_peruvianus
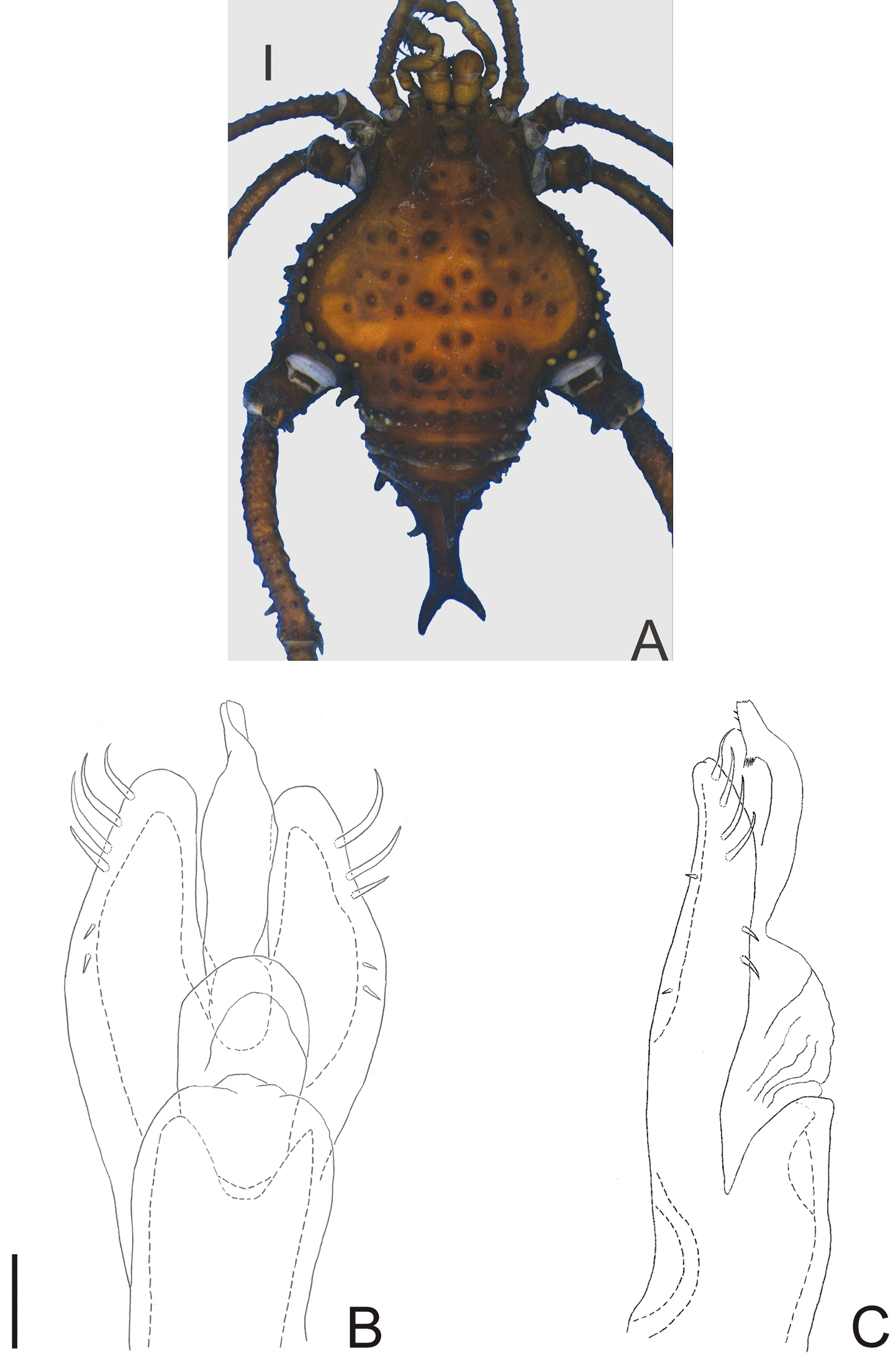
Fig. 4Neopachyloides was a hitherto monotypic genus that resembles Ampycinae genera with a paired armature on ocularium and free tergite III with a median, long spiniform apophysis asin Ampycella Roewer, 1929, Ampycus Simon, 1879, Hutamaia Soares & Soares, 1977 and Sibollus Roewer, 1929. Neopachyloides can be distinguished from these genera by the following combination of characteristics: scutal area III with a paramedian pair of enlarged, pointed tubercles, scutal area IV undivided and free tergite II unarmed. Neopachyloides peruvianus can be distinguished from Neopachyloides spinipes Roewer, 1913 by the dorsal scutum covered by granules and scutal areas I–II unarmed (Fig. 4A).
Penis (Fig. 4B–C; holotype): ventral plate with sub hexagonal shape, deep cleft on distal margin, 4 pairs of distal setae (distalmost curved and basalmost small), 3 pairs of basal setae straight. Glans very long (⅔ of ventral plate length), stylus smooth and curved dorsally, without dorsal and ventral processes.
The assignment of Neopachyloides peruvianus to Neopachyloides is based on overall similarity and should be considered tentative. Neopachyloides peruvianus is the only Ampycinae which presents just one scutal area armed with a paramedian enlarged pair of tubercles (most genera present 3 scutal areas armed with a paramedian pair of enlarged tubercles, as Ampycus, Hexabunus and Pirunipygus or all of them unarmed). We preferred to place Neopachyloides peruvianus under Neopachyloides instead of proposing a new genus because monophyly of Ampycinae genera (most of them monotypic) are doubtful. See taxonomical note of Pirunipygus paradoxus for the reasons of the new placement of Neopachyloides peruvianus.
Neopachyloides peruvianus (Roewer) comb. n. Male (holotype): A habitus, dorsal view. Distal part of penis (holotype) B in dorsal view C ditto, left lateral view. B, C at same scale. Scale bars: A = 1 mm; B = 0.1 mm.
http://species-id.net/wiki/Pirunipygus
Pirunipygus is a monotypic genus and resembles Ampycinae genera with a paramedian pair of enlarged tubercles on scutal areas I–II as Ampycus Simon, 1879, Hexabunus Roewer, 1913, Neopachyloides Roewer, 1913 and Parahernandria Goodnight & Goodnight, 1947. Pirunipygus paradoxus can be distinguished from these genera by the following combination of characters: ocularium with a median spiniform apophysis (Fig. 5A); dorsal scutum with secondary tubercles (sensu
Monotypic.
Pirunipygus paradoxus Roewer. Male (holotype): A habitus, dorsal view. Distal part of penis (holotype) B in dorsal view; C ditto, left lateral view. B, C at same scale. Scale bar: A = 1 mm. Scale bar: B = 0.1 mm.
http://species-id.net/wiki/Pirunipygus_paradoxus
Fig. 5Penis (Fig. 5B–C; holotype): ventral plate sub hexagonal, deep U-cleft on distal margin, 3–4 distal pairs of setae (2–3 distalmost curved), 2 pairs of dorsal median setae of moderate size, 2 pairs of ventral median small setae. Stylus wide on basal ¾, apex twisted, with a patch of ventral subapical trichomes and some scattered apically. Glans without dorsal and ventral processes.
Both Neopachyloides peruvianus and Pirunipygus paradoxus were formerly placed in Gonyleptidae, Pachylinae. These two species show the following diagnostic characteristics of Ampycinae (
http://species-id.net/wiki/Acanthogonyleptes
As in
The monotypic genus Centroleptes is a subjective synonym of Acanthogonyleptes, since the type examined material of Centroleptes flavus and Acanthogonyleptes singularis are identical and synonymized here.
Acanthogonyleptes alticola (Mello-Leitão, 1922); Acanthogonyleptes editus (Roewer, 1943); Acanthogonyleptes fallax (Mello-Leitão, 1932); Acanthogonyleptes fulvigranulatus (Mello-Leitão, 1922); Acanthogonyleptes marmoratus (Mello-Leitão, 1940); Acanthogonyleptes pictus (Piza, 1942); Acanthogonyleptes singularis (Mello-Leitão, 1935); Acanthogonyleptes soaresi (Mello-Leitão, 1944); Acanthogonyleptes variolosus (Mello-Leitão, 1940).
http://species-id.net/wiki/Acanthogonyleptes_singularis
BRAZIL. São Paulo: São José do Barreiro (Serra da Bocaina, Fazendas Águas de Santa Rosa), female holotype of Bocaina marmorata Piza (MZSP 810); Ribeirão Pires, male holotype of Adelphobunus singularis Mello-Leitão (IBSP 17); São José do Barreiro, 1 ♂ (HEMS 761); Santo André, 1 ♂ & 1 ♀ (MZSP 21257). Santa Catarina: Seara (Nova Teutônia), ♀ holotype of Centroleptes flavus Roewer (SMF 6430/63).
Acanthogonyleptes singularis and Acanthogonyleptes variolosus can be distinguished from the remaining species of the genus by the presence of a median single elevation on scutal area III. Males of Acanthogonyleptes singularis can be distinguished from those of Acanthogonyleptes variolosus by the presence of a robust curved retrolateral apophysis on trochanter IV.
The genus Acanthogonyleptes is a morphologically homogeneous group, probably representing a monophyletic group (see Sphaerobunus in
http://species-id.net/wiki/Geraeocormobius_sylvarum
Brazil. Paraná: Cachoeirinha, 2 ♂ & 5 ♀ (MNRJ 26912); Paranavaí, 6 ♂ & 7 ♀ (MNRJ 4732).
http://species-id.net/wiki/Gonyleptellus_bimaculatus
BRAZIL. Locality not specified further, female holotype of Gonyleptes bimaculatus, examined by digital photos (ZMUC). São Paulo: Santos, ♂ holotype of Gonyleptes cancellatus (SMF 1320); Bananal (Estação Ecológica do Bananal), A. Monteiro leg., IV.2004, 10 ♂ & 10 ♀ (MZSP 27804). Rio de Janeiro: Itatiaia (Parque Nacional do Itatiaia), Equipe Biota leg., 8–15.VI.2001, 3 ♂ & 1 ♀ (MZSP 21746); Terezópolis (Parque Nacional da Serra dos Órgãos, sede), R. Pinto-da-Rocha, S. Outeda-Jorge, C. Mattoni leg., 10.II.2007, 3 ♂ & 5 ♀ (MZSP 28223); Nova Friburgo (Mury, Debossan, 950 m), R.S. Bérnils & P. Labiak leg., 29.VII.1996, 6 ♂ & 3 ♀ (MZSP 15120).
http://species-id.net/wiki/Gonyleptes
Gonyleptes is a genus composed of generally large species (dorsal scutum length longer than 8 mm), which males present femur IV with conspicuous armature and scutal areas I–III convex in lateral view. The penis presents ventral plate very convex and apex of ventral process serrate and semicircular. See taxonomical note below.
Gonyleptes possesses 23 species and its habitus resembles other genera placed in Gonyleptinae. As
Composition. Gonyleptes acanthopus (Quoy & Gaimard, 1824); Gonyleptes armatus Perty, 1833; Gonyleptes atrus Mello-Leitão, 1923, revalidated; Gonyleptes barbiellinii Mello-Leitão, 1932; Gonyleptes calcaripes (Roewer, 1917); Gonyleptes curticornis (Mello-Leitão, 1940); Gonyleptes curvicornis (Roewer, 1932), revalidated, comb. n.; Gonyleptes fragilis Mello-Leitão, 1923; Gonyleptes gertschi Soares & Soares, 1948; Gonyleptes gonyleptoides (Soares & Soares, 1945); Gonyleptes granulatus (Piza, 1940); Gonyleptes horridus Kirby, 1819; Gonyleptes parcigranulatus Soares & Soares, 1949; Gonyleptes pectinatus Koch, 1845; Gonyleptes pectinipes Roewer, 1917; Gonyleptes perlatus (Mello-Leitão, 1935), comb. n.; Gonyleptes pseudogranulatus Soares & Soares, 1946; Gonyleptes pseudoguttatus Giltay, 1928; Gonyleptes pustulatus Sørensen, 1884; Gonyleptes recentissimus Mello-Leitão, 1932; Gonyleptes saprophilus Mello-Leitão, 1922; Gonyleptes vatius Bertkau, 1880; Gonyleptes viridisagittatus Soares & Soares, 1945.
http://species-id.net/wiki/Gonyleptes_horridus
Figs 6A–D; 7BRAZIL. Locality not specified further, male holotype of Gonyleptes horridus, examined by digital photos (NHM 1863.41). São Paulo: São Paulo, male holotype of Metagonyleptes hamatus (SMF 891). Rio de Janeiro: Rio de Janeiro (Gávea), 1 male (MZSP 1370). Santa Catarina: (Serra Azul), male holotype of Paragonyleptes simoni (SMF RII 1333).
Gonyleptes horridus can be recognized by: the relatively larger size; anterior border of carapace smooth (without tubercles or spines in the corners); frontal hump on anterior border of carapace and ocularium with white small tubercles; prolateral apical apophysis of coxa IV long, with abrupt curvature backwards. Gonyleptes horridus, Gonyleptes perlatus and Gonyleptes pustulatus can be distinguished from all other species of Gonyleptes by the robust and bifid (two distal tips) retrolateral apical apophysis of coxa IV. Females of Gonyleptes horridus and Gonyleptes perlatus present dimorphic sulfur yellow spines on the free tergites, not present in females of Gonyleptes pustulatus. Gonyleptes horridus can be distinguished from Gonyleptes perlatus by the armature pattern of femur IV of the males (see note on Gonyleptes perlatus below). Gonyleptes horridus has relatively smaller tubercles on posterior part of the lateral margins of abdominal scutum than in Gonyleptes perlatus.
Dorsal scutum length 7–9 mm. Anterior margin of carapace without tubercles or spines. Median frontal hump on anterior margin of carapace and ocularium with a pair of tubercles on each one (Fig. 6A). Four scutal grooves delimiting three scutal areas. Scutal areas I, II with low white tubercles in the median region; III with high paramedian tubercles. Lateral margin of dorsal scutum with flattened white tubercles. Pedipalps approximately same length as dorsal scutum. Prolateral apophyses of coxa IV elongated, strongly curved backwards, dorsal branch longest. Retrolateral apophysis of coxa IV bifid. Femur IV straight, with dorsobasal apophyses and longitudinal rows of spines (Fig. 6B–C). Penis with typical Gonyleptinae-like pattern (
All synonyms given in
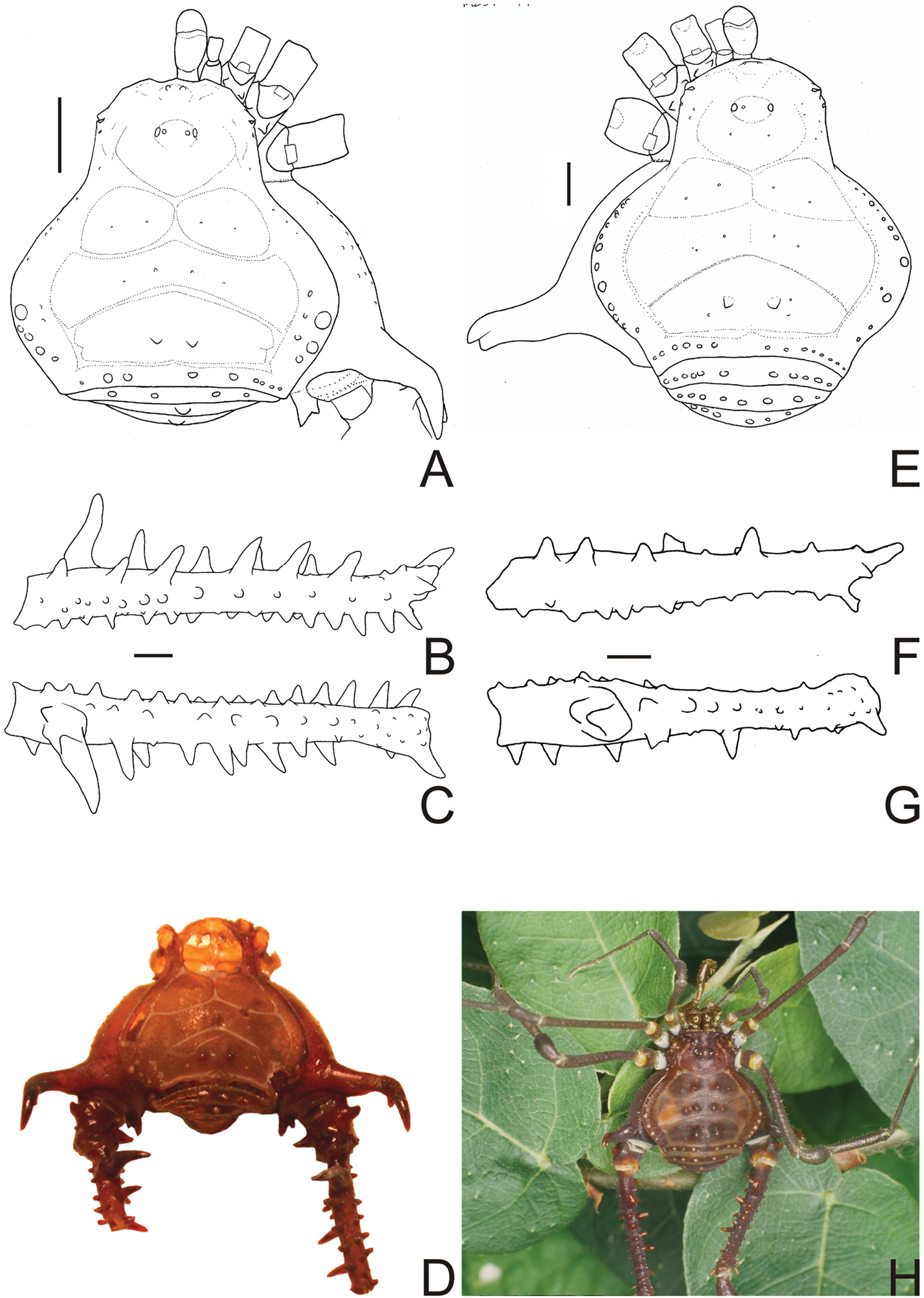
Gonyleptes spp. male dorsal habitus and femora IV: A–D Gonyleptes horridus Kirby (HEMS 103): A habitus, dorsal view B right femur IV ventral view C ditto, dorsal view D picture of the holotype (NHM 1863.41). E–H Gonyleptes curvicornis (Roewer) (MZSP 0871): E habitus, dorsal view F right femur IV ventral view G ditto, dorsal view H picture of a live specimen. B, C at same scale. F, G at same scale. Scale bars: 1 mm.
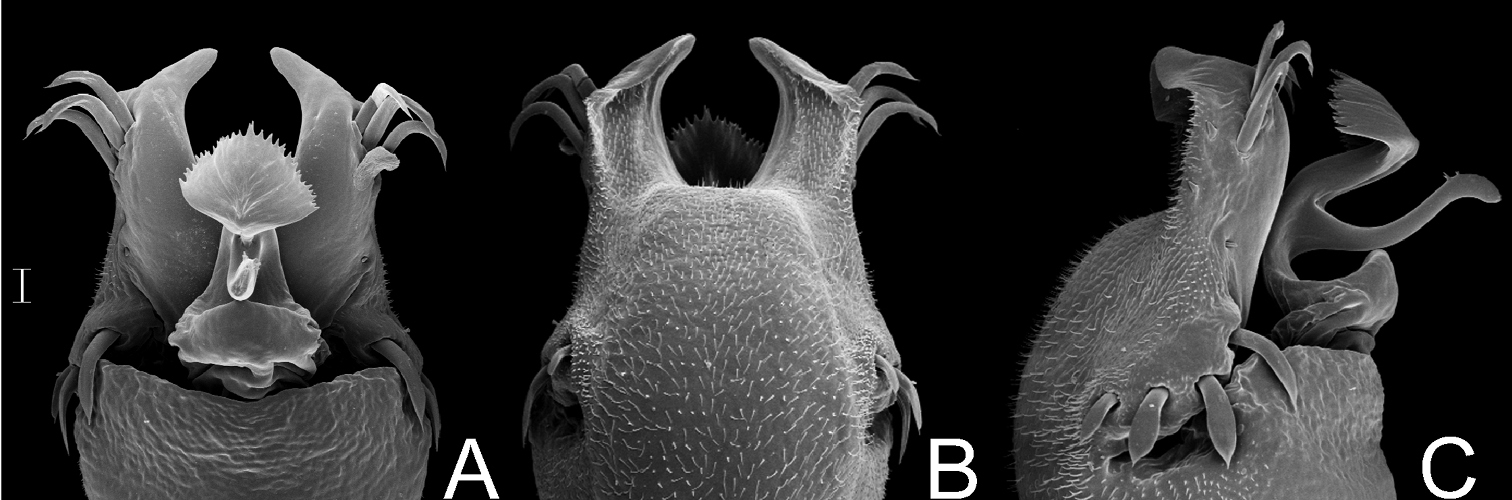
Gonyleptes horridus Kirby. Distal part of penis (MZSP 15496) A dorsal view B ditto, ventral view C ditto, left lateral view. Scale bar: 0.02 mm.
http://species-id.net/wiki/Gonyleptes_curvicornis
Figs 6E–H; 8BRAZIL. São Paulo: Locality not specified further, male holotype (SMF 979); São Paulo (Chácara Dr. J. L. Lane), male (MZSP 0871). Rio de Janeiro: Rio de Janeiro (Grajaú), 1 male (MZSP 1117).
Gonyleptes curvicornis resembles Gonyleptes horridus (see diagnosis above) but can be distinguished by the relatively shorter prolateral apical apophysis of coxa IV, lacking the abrupt backwards curvature (Fig. 6E). The retrolateral apical apophysis of coxa IV is reduced and not bifid as in Gonyleptes horridus. The male femur IV has a different pattern of armature (Fig. 6F–G) compared to Gonyleptes horridus. Carapace with a pair of tubercles on the posterior area (posterior to ocularium). Females of Gonyleptes curvicornis do not present dimorphic sulfur yellow spines on the free tergites.
Penis (Fig. 8A–B; MZSP 1117): ventral plate sub hexagonal, with a deep U-cleft on distal margin (its distal tips convergent), 3 distal, 1 median, 4 basal pairs of setae (distal ones curved, median ones shortest). Stylus sigmoid, with ventral subapical trichomes. Ventral process robust, apex flabelliform with several projections of varying sizes on distal margin.
Roewer’s (1913; 1923) redescriptions of Gonyleptes horridus and Weyhia curvicornis are the same. Following Gonyleptes horridus sensu Roewer,
Gonyleptes curvicornis (Roewer). Distal part of penis (MZSP 1117) A dorsal view B ditto, left lateral view. Scale bars: 0.05 mm.
http://species-id.net/wiki/Gonyleptes_atrus
Figs 9A–B, 10BRAZIL. Minas Gerais: Lambari (Parque Estadual Nova Baden), (MZSP 32513); Delfim Moreira (MZSP 29393); Itamonte (MZSP 21251); Poços de Caldas (MZSP 29366). Rio de Janeiro: Itatiaia, syntypes of Weyhia bisignata (MNRJ 27321). São Paulo: Campos do Jordão (MZSP 21251).
Dorsal scutum length 8–10 mm. Corners of anterior margin of carapace smooth. Median frontal hump on anterior margin of carapace and ocularium with one pair of tubercles each one. Posterior region of carapace with 2 median tubercles. Mesotergum densely covered by tubercles. Scutal areas I–III each with a pair of median enlarged tubercles (III with largest and round ones). Prolateral apical apophysis of male coxa IV slightly directed backwards, retrolateral apophysis of male coxa IV absent or very reduced (Fig. 9A–B). Dorsobasal apophysis of male femur IV robust, curved and retrolaterally oriented. Ornamentation of male femur IV variable, normally with two prolateral spines.
Penis (Fig. 10A–C, MZSP 29393): ventral plate with 3 distal and 3 basal pairs of large setae, 1 median pair of short setae. Stylus with ventral trichomes. Ventral process with a triangular flabellum, with digitiform ventral median projection.
Gonyleptes atrus was synonymized, without further remarks, with Gonyleptes saprophilus by
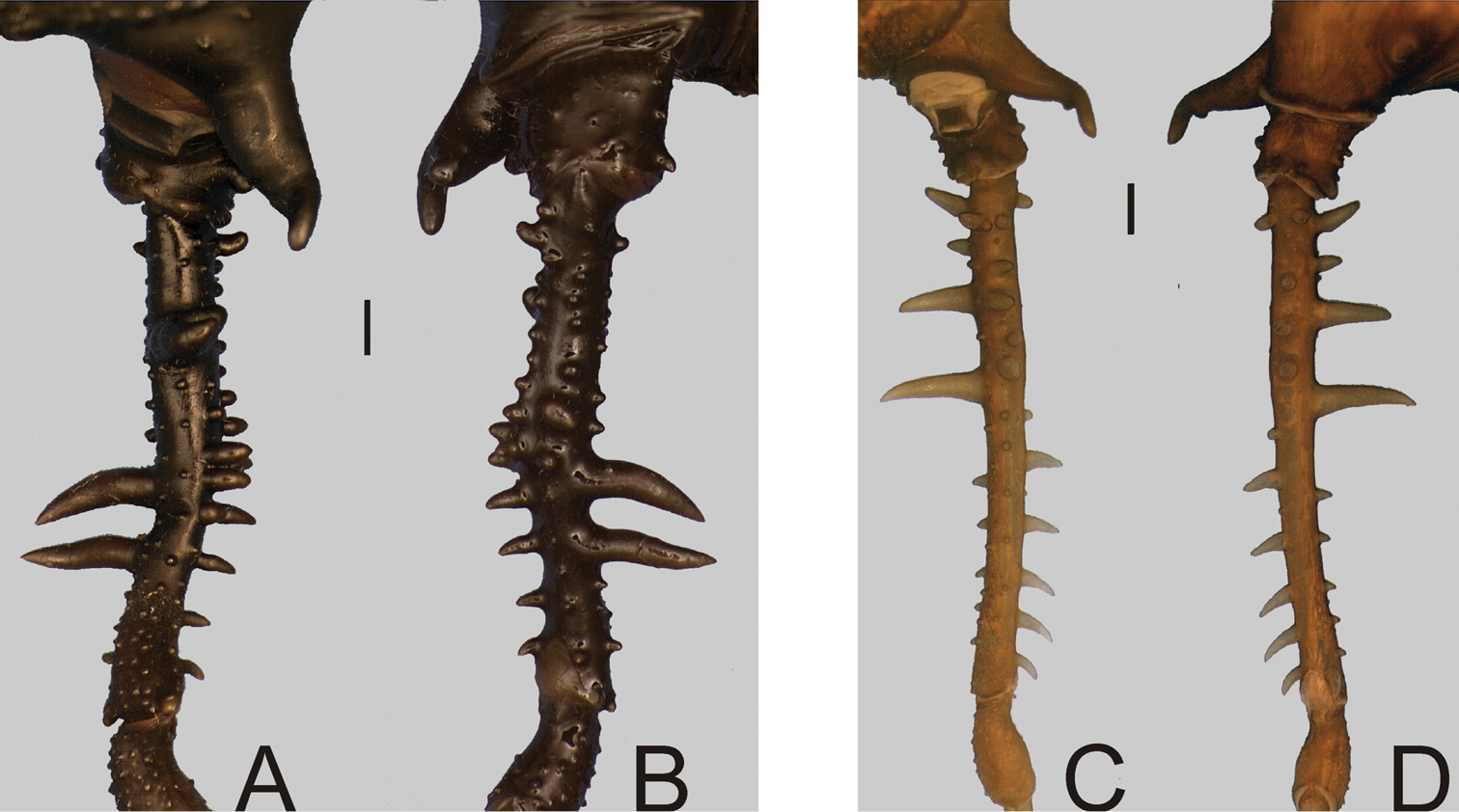
Gonyleptes spp. male femora IV: A, B Gonyleptes atrus Mello-Leitão (MZSP 32513): A right femur IV dorsal view B ditto, ventral view. C, D Gonyleptes saprophilus Mello-Leitão (MZSP 9996): C right femur IV dorsal view D ditto, ventral view. A, B at same scale. C, D at same scale. Scale bars: 1 mm.
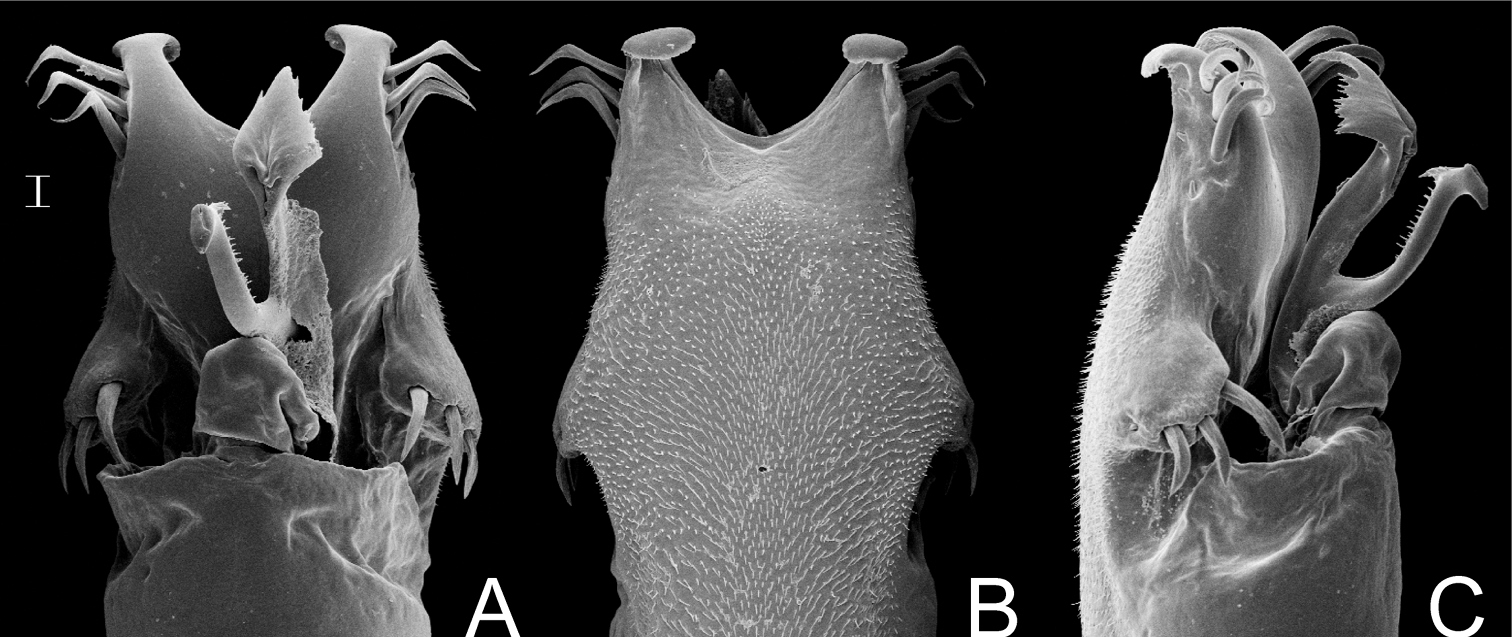
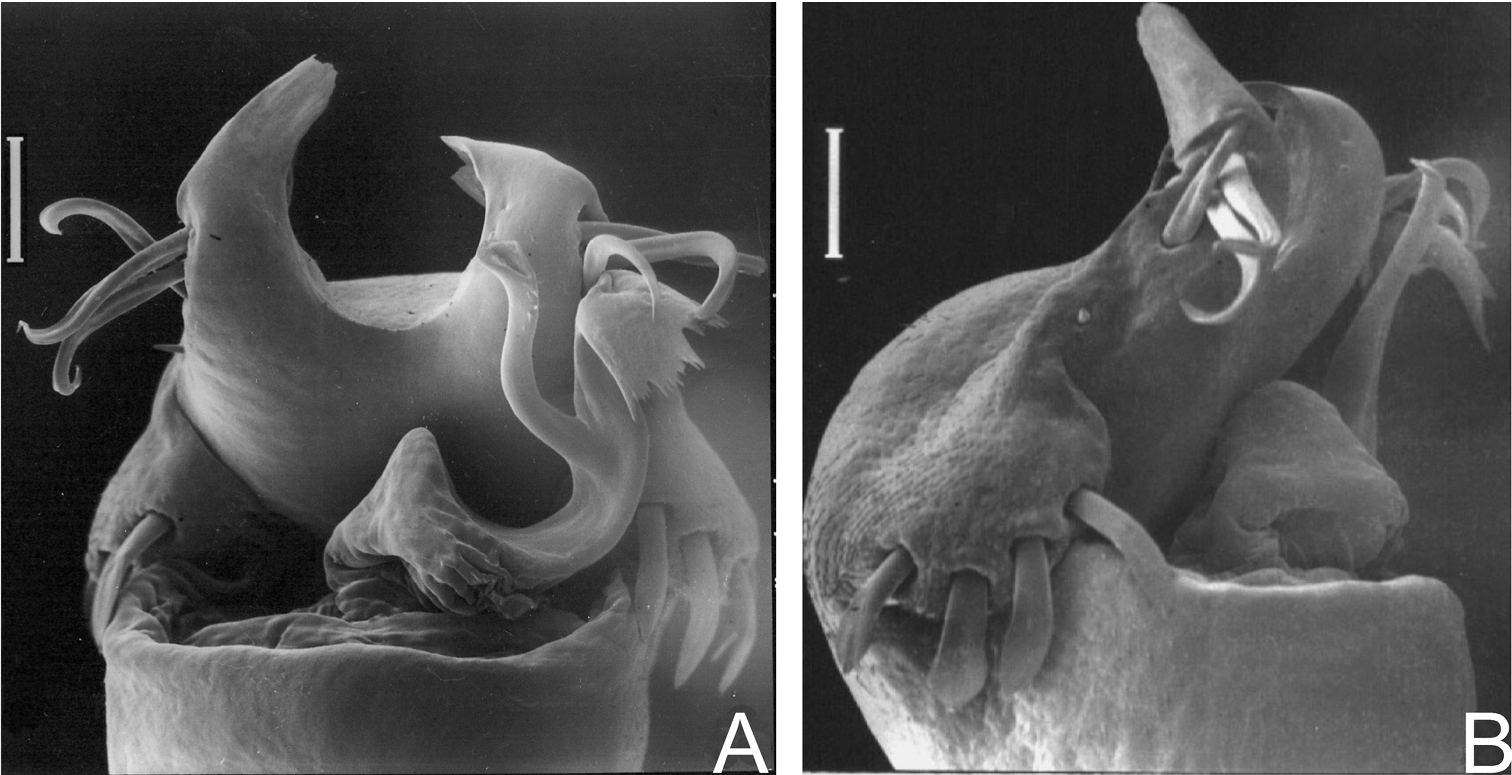
Gonyleptes atrus Mello-Leitão. Distal part of penis (MZSP 29393) A dorsal view B ditto, ventral view C ditto, left lateral view. Scale bar: 0.02 mm.
http://species-id.net/wiki/Gonyleptes_fragilis
Fig. 11Gonyleptes fragilis resembles Gonyleptes horridus and Gonyleptes curvicornis because of the overall similarities of the features of the dorsal scutum. Gonyleptes fragilis can be distinguished from the other two species by the ocularium with a pair of spines (tubercles in Gonyleptes horridus and Gonyleptes curvicornis) and the pale yellowish spots around the tubercles on abdominal scutum.
Penis (Fig. 11A–C): ventral plate inflated in the middle region, with a deep U-cleft on anterior margin (its distal tips convergent), 3 distal pairs of long, curved setae, 1 median pair of short setae, 4 basal pairs of spatulate setae. Glans sac slightly projected distally, with similar projection to a dorsal process. Stylus sigmoid, smooth. Ventral process robust, apex flabelliform with several small projections on distal margin.
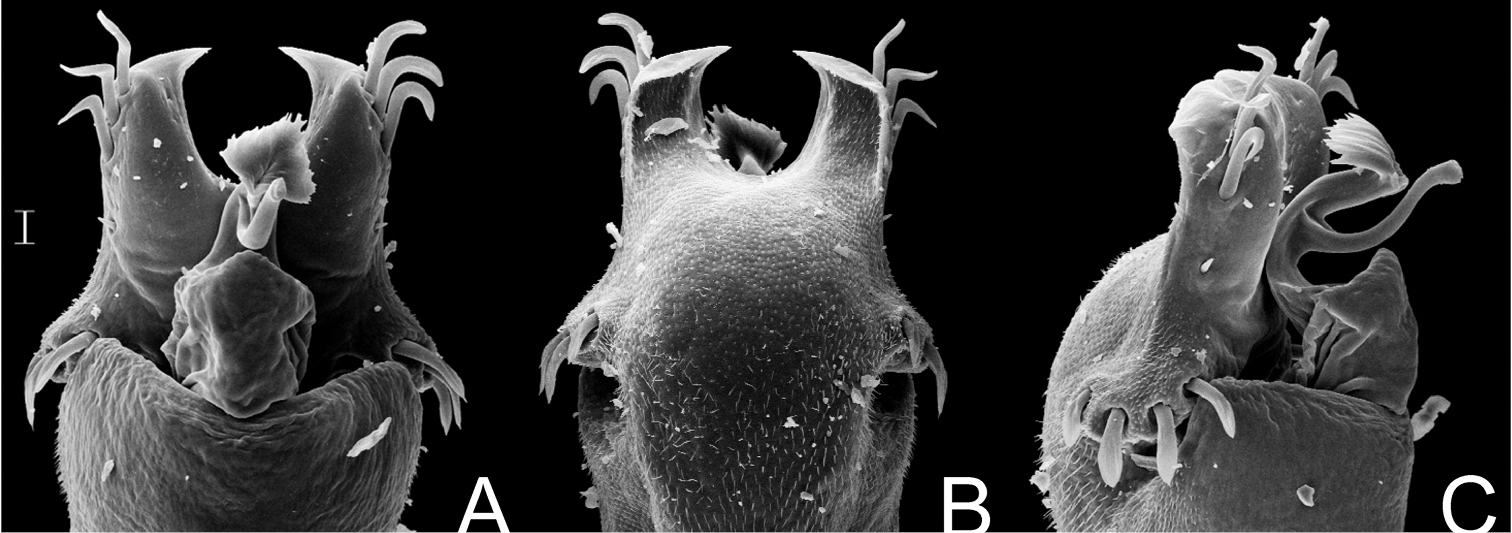
Gonyleptes fragilis Mello-Leitão. Distal part of penis (MZSP 30764) A dorsal view B ditto, ventral view C ditto, left lateral view. Scale bar: 0.02 mm.
http://species-id.net/wiki/Gonyleptes_perlatus
Gonyleptes perlatus is very similar to Gonyleptes horridus (according to
We propose the allocation of this species in Gonyleptes because it is very similar to Gonyleptes horridus (see comments in
http://species-id.net/wiki/Gonyleptes_pustulatus
Fig. 12BRAZIL. Locality not specified further, male holotype of Gonyleptes pustulatus, examined from photo (ZMUC). São Paulo: Santos, male holotype of Gonyleptes guttatus (SMF 1321). Rio de Janeiro: Casimiro de Abreu, 5 ♂ & 6 ♀ (MNRJ 17456).
Gonyleptes pustulatus (Fig. 12A–B) superficially resembles Gonyleptes bimaculatus because of the conspicuous white patches on mesotergum. Males of Gonyleptes pustulatus can be distinguished from Gonyleptes bimaculatus by: femur IV armed (unarmed in Gonyleptes bimaculatus) and presence of a bifid retrolateral apical apophysis on coxa IV.
Penis (Fig. 12C–D; MNRJ 17456): ventral plate with slightly inflated middle region, a deep distal U-cleft on anterior margin (its distal tips are divergent), 3 distal pairs of long, curved setae, 1 median pair of short setae, 4 basal pairs of spatulate setae. Stylus slightly straight, with apex slightly swollen. Ventral process robust, apex flabelliform with serrated and rounded distal margin.
Gonyleptes pustulatus is a remarkable species by body color pattern. It was described without any illustration. Probably this was the reason that kept this species virtually unknown and led Roewer to describe Gonyleptes guttatus.
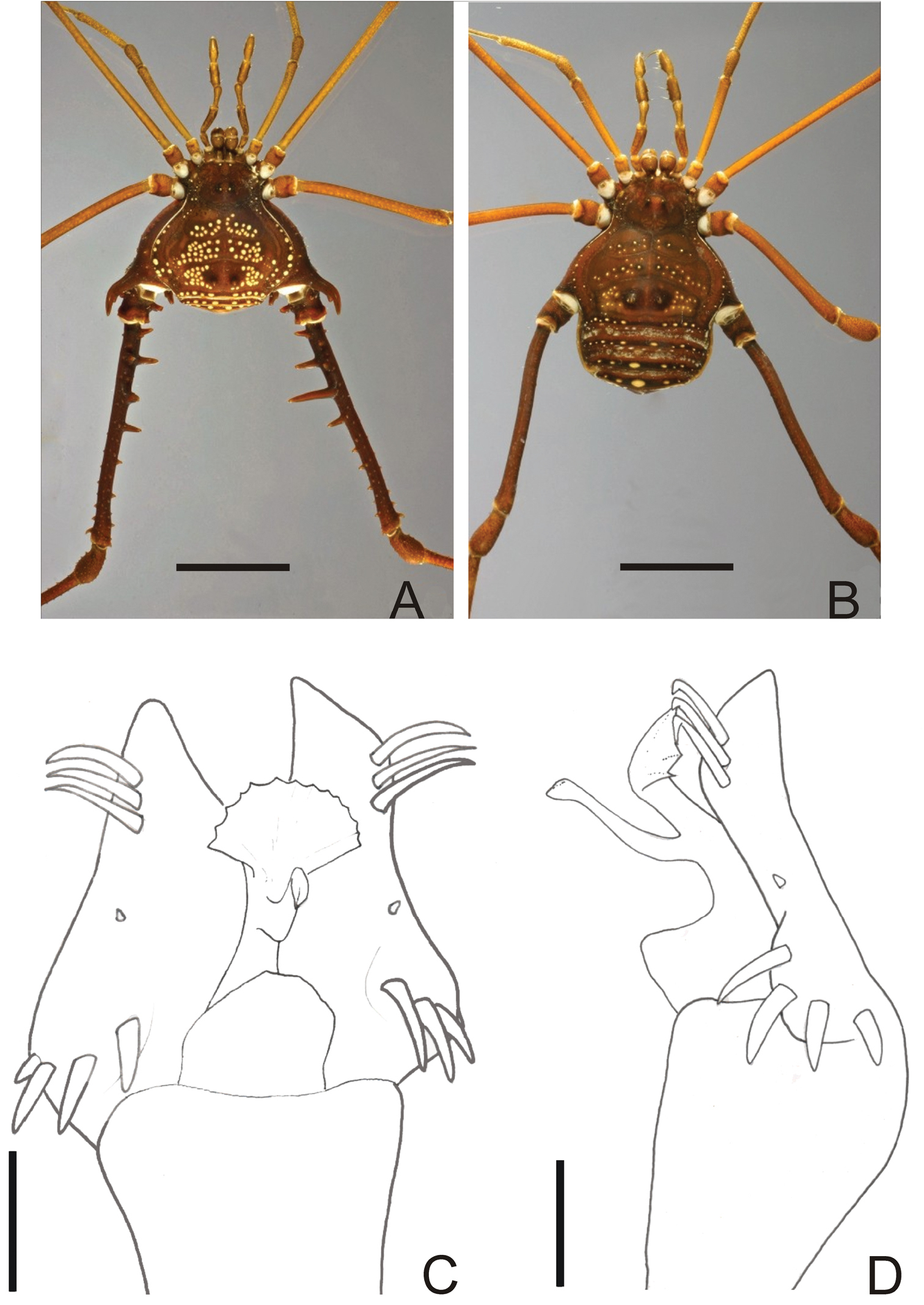
Gonyleptes pustulatusSørensen. Male (MNRJ 17456): A habitus in dorsal view. Female (MNRJ 17456): B habitus in dorsal view. Distal part of penis (MNRJ 17456) C in dorsal view D ditto, right lateral view. Scale bars: A–B = 1.0 mm; C–D = 0.01 mm.
http://species-id.net/wiki/Liogonyleptoides_minensis
Comparisons between the type material of Currala bahiensis and Liogonyleptoides minensis allowed us to conclude that they are synonymous. Liogonyleptoides minensis is a species that can be easily diagnosed among gonyleptines by the relatively larger size, body dorsally rounded, without tubercles on corners, presence of scutal area IV; femur III curved (instead of sigmoid); male femur IV with a long, robust dorsobasal apophysis and a prolateral one that is curved dorsally on distal third. The genus Currala is now composed only by its type species Currala spinifrons Roewer, 1927.
http://species-id.net/wiki/Mischonyx
Mischonyx resembles Currala Roewer, 1927 and Schenkelibunus Strand, 1932, which are gonyleptine of relatively smaller size (about 5 mm of dorsal scutum length). Mischonyx can be distinguished from those genera by the combination of the following characters: anterior border of the carapace with robust spines; frontal hump on anterior margin of carapace and ocularium with a pair of spines; scutal areas I–III with a paramedian pair of tubercles, those on I–II elliptical and widest pair on scutal area III; lateral margin of the dorsal scutum with large white tubercles (in specimens preserved in 70% ethanol).
According
Mischonyx anomalus (Mello-Leitão, 1936); Mischonyx antiquus (Mello-Leitão, 1934); Mischonyx cuspidatus (Roewer, 1913); Mischonyx fidelis (Mello-Leitão, 1931); Mischonyx insulanus (Soares, 1972); Mischonyx intermedius (Mello-Leitão, 1935); Mischonyx kaisara Vasconcelos, 2004; Mischonyx poeta Vasconcelos, 2005; Mischonyx processigerus (Soares & Soares, 1970); Mischonyx squalidus Bertkau, 1880 and Mischonyx sulinus (Soares & Soares, 1947).
http://species-id.net/wiki/Mischonyx_scaber
BRAZIL. Locality not specified further, 3 males & 1 female syntypes (NHM 1863.41). Rio de Janeiro: Niterói, female holotype of Xundarava holacantha (MNRJ 1469); ditto, male holotype of Weyhia absconsa (MNRJ 1483).
Mischonyx scaber resembles Mischonyx cuspidatus and Mischonyx poeta because of armature of the male femur IV: presence of a dorsobasal apophysis relatively short and curved; one retrolateral apophysis on distal third; and a row of high, acuminated tubercles increasing in size from the base to distal third. Mischonyx scaber can be distinguished from Mischonyx cuspidatus by the presence of a short gap between the retrolateral row and the retrolateral apophysis on femur IV (the retrolateral row is continuous to the retrolateral apophysis on femur IV in Mischonyx cuspidatus). Mischonyx scaber can be distinguished from Mischonyx poeta by the absence of enlarged tubercles (present in Mischonyx poeta) on the posterior part of the lateral margin of the dorsal scutum.
Mischonyx scaber was described from Brazil, however, many authors, including
http://species-id.net/wiki/Mischonyx_cuspidatus
BRAZIL. Rio de Janeiro: Ilha Grande, male holotype (SMF 900); Macaé, 16 ♂ & 6 ♀ (MNRJ 4601). Santa Catarina: Serra Azul, male holotype (SMF 1328); Gaspar, 1 ♂ (MNRJ 11584).
Mischonyx cuspidatus resembles Mischonyx poeta and Mischonyx scaber (see diagnosis of Mischonyx scaber). Mischonyx cuspidatus can be distinguished from them by the median high, acuminated tubercle on the free tergites I–III.
The type locality of Gonazula gibbosa, Serra Azul (in the state of Santa Catarina), is an unknown toponym from where seven species are recorded (see
http://species-id.net/wiki/Megapachylus_grandis
Fig. 13Penis (Fig. 13A–C; MZSP16837): ventral plate with slightly convergent lateral sides deep U-cleft on distal margin, its apex folded ventrally, 3 distal pairs of large setae, slightly curved at apex, 1 median pair of short setae and, 4 basal pairs of spatulate setae, directed frontwards. Glans with dorsal process. Stylus thin, long, sigmoid, with subapical ventral trichomes. Ventral process shaft folded ventrally, apex flabelliform, with serrate distal margins and long distal tip.
The male of the species described by Soares & Soares shows a moderate development of armature of leg IV but the number and arrangement of tubercles are perfectly coincident with those of Megapachylus grandis. It could be a beta male, although an extensive population study should be conducted to confirm this.
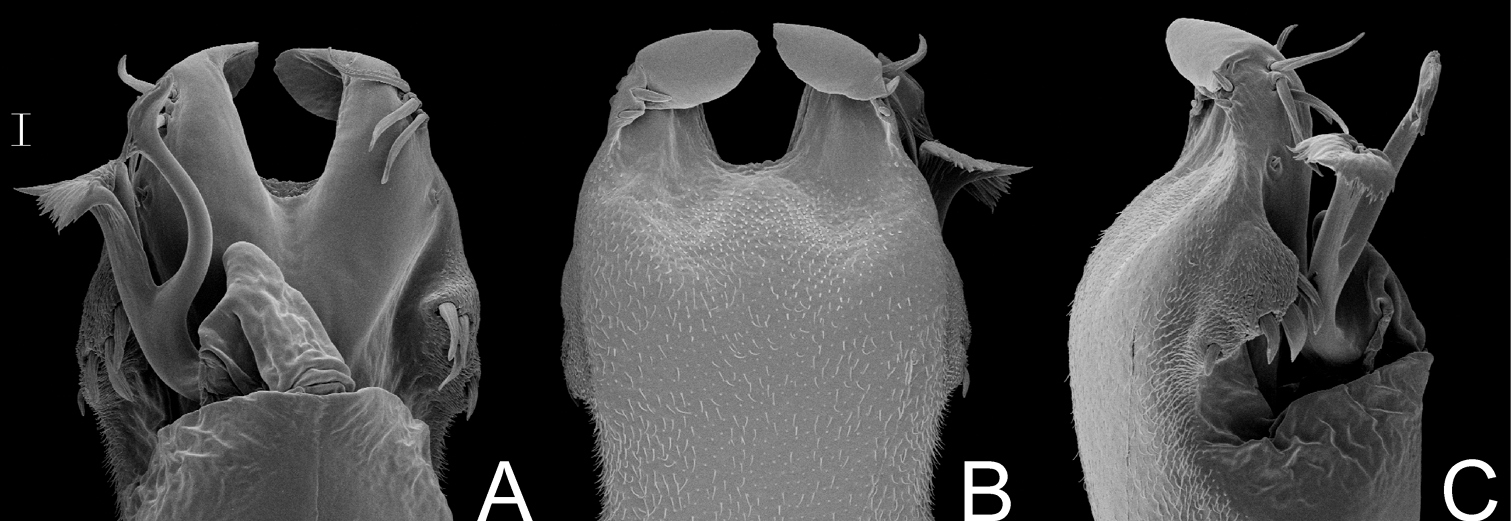
Megapachylus grandis Roewer. Distal part of penis (MZSP16837) A in dorsal view B ditto, ventral view C ditto, left lateral view. Scale bar: 0.02 mm.
http://species-id.net/wiki/Parampheres
Parampheres resembles Acanthogonyleptes, Adhynastes Roewer, 1930 and Deltaspidium Roewer, 1927 by the presence of a more elongated prolateral apical apophysis on male coxa IV (greater length/basal diameter ratio of the apophysis) than that in Gonyleptes and related Gonyleptinae genera. Parampheres can be distinguished from Adhynastes and Deltaspidium by the scutal area III with tubercles (instead of spines). Parampheres can be distinghished from Acanthogonyleptes by the absence of a clearly defined dorsobasal apophysis on the male femur IV. Parampheres species bear a row of apophyses on the dorsal basal area of the femur IV.
Metapachyloides is a monotypic genus and its type species is junior synonym of Parampheres tibialis Roewer, 1917.
http://species-id.net/wiki/Parampheres_tibialis
The monotypic genus Metapachyloides was formerly placed in Gonyleptidae, Pachylinae. Instead of a male, as stated in the original description, the holotype of Metapachyloides rugosus is a female, and its general aspect is similar to that of Parampheres Roewer, 1913. Surprisingly, in the same year, Roewer described Parampheres tibialis in Gonyleptinae, based on a male collected in the same locality (Santos), the most important harbor in Brazil. Roewer described many harvestmen species indicating Santos as their type locality, but those were never collected again there or in the area of endemism to which Santos belongs (Serra do Mar of São Paulo, see
http://species-id.net/wiki/Parapachyloides_uncinatus
Fig. 14BRAZIL. São Paulo: Porto Cabral, 4 ♂ (MZSP 787); Usina Hidrelétrica de Rosana, 2 males & 2 females (MZSP 14571). Mato Grosso do Sul: Taquaruçu, 2 ♂ & 1 ♀ (MZSP 19113); ditto, (Canal do Rio Baía), 1 ♂ (MZSP 22084).
Penis (Fig. 14A–C; MZSP 14571): ventral plate with the lateral sides slightly convergent, deep U-cleft on distal margin, its apex folded ventrally and convergent, 3 distal pairs of setae curved on apex, 1 pair of short median setae, 4 basal pairs of setae arranged in a row, 2 pairs of ventral distal short setae. Glans with small dorsal process. Stylus slender, sigmoid, smooth. Ventral process apex flabelliform with serrate distal margin.
Males of this species show polymorphism in the posterior tubercle on the lateral margin of dorsal scutum (from low, small and blunt to high, enlarged, curved and pointed apex), posterior margin of dorsal scutum (from low tubercles to two high tubercles) and armature of free tergite I (with one large median tubercle without enlarged lateral tubercles, or with 2–4 high lateral tubercles); retrolateral basal tubercle on male femur IV of same width as femur to 1.5 times wider, short (1.3 times femur width) to long (3.2 ×).
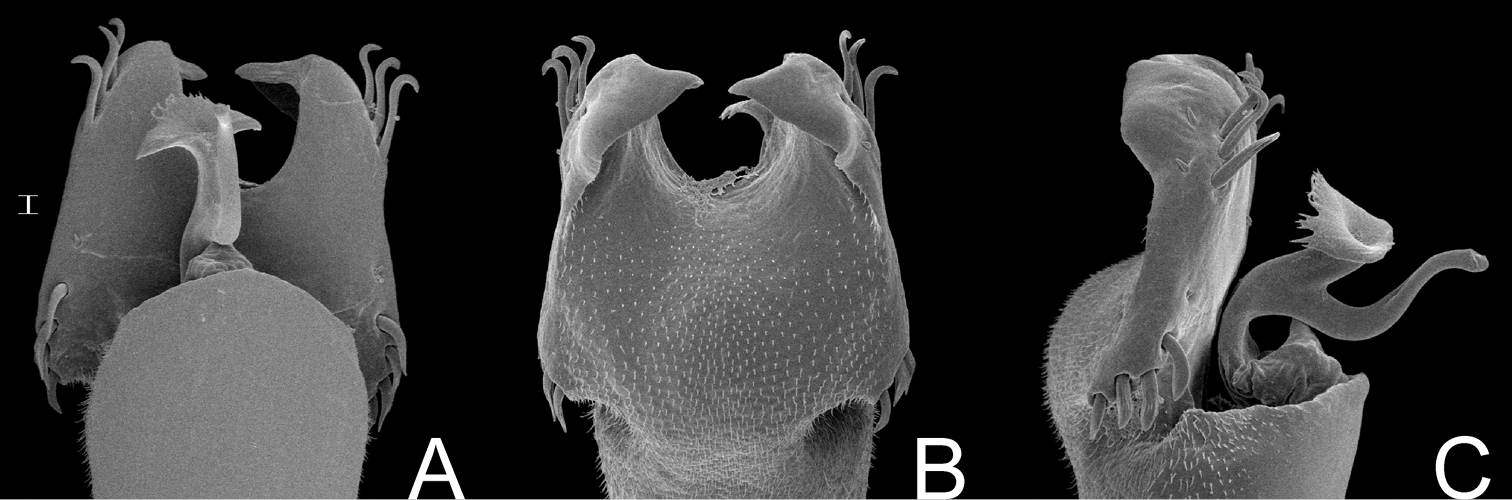
Parapachyloides uncinatus (Sørensen). Distal part of penis (MZSP 14571) A in dorsal view B ditto, ventral view C ditto, left lateral view. Scale bar: 0.01 mm.
http://species-id.net/wiki/Schubartesia_singularis
Fig. 15BRAZIL. Bahia: (Os Gerais, Rio Branco Valley), male holotype of Schubartesia singularis (MZSP). Minas Gerais: Januária (Parque Nacional Cavernas do Peruaçu), 1 ♂ & 1 ♀ (MZSP 29075); ditto, 2 ♂ (MZSP 29839).
Penis (Fig. 15A–C; MZSP 29075): ventral plate with the sides subparallel, with deep cleft on distal margin (U-cleft in ventral view and V-cleft in dorsal view; its apex convergent), 3 pairs of long helycoidal setae, 2 pairs of ventral distal short setae, 1 pair of median short setae, 4 pairs of spatulate, large setae arranged in an oblique row in lateral view, basal lobe short and projected dorsad. Stylus sigmoid, smooth. Ventral process shaft short and wide, apex flabelliform with slightly serrated distal margin.
Schubartesia singularis and Parapachyloides armatus were both formerly placed in Pachylinae and transferred recently to Gonyleptinae by
Schubartesia singularis B. Soares. Distal part of penis (MZSP 29075) A in dorsal view B ditto, ventral view C ditto, left lateral view. Scale bar: 0.02 mm.
http://species-id.net/wiki/Pseudopucrolia
See
Composition. Pseudopucrolia discrepans (Roewer, 1943); Pseudopucrolia incerta (Mello-Leitão, 1928); Pseudopucrolia mutica (Perty, 1833) and Pseudopucrolia rugosa (Roewer, 1930).
Meteusarcus armatus was formerly placed in Gonyleptidae, Pachylinae. The examination of the female holotype revealed it was a female of the heteropachyline Pseudopucrolia mutica.
http://species-id.net/wiki/Pseudopucrolia_mutica
See
See genus “Taxonomical note”.
http://species-id.net/wiki/Acrographinotus
See
Acrographinotus ceratopygus (Soares & Bauab-Vianna, 1972); Acrographinotus curvispina Roewer, 1929; Acrographinotus erectispina Roewer, 1929; Acrographinotus mitmaj Acosta, 2002; Acrographinotus niawpaq Acosta, 2001; Acrographinotus ortizi (Roewer, 1957); Acrographinotus ornatus (Roewer, 1929), comb. n.
The genus Unduavius is monotypic, but the remarkable ventral process of the penis, with its “ibis head” shape, fits perfectly in the generic concept presented by
http://species-id.net/wiki/Acrographinotus_ornatus
Fig. 16BOLIVIA. La Paz: Nor Yungas (Unduavi), 1 male & 6 females (SMF 2881/79; syntypes according to
Acrographinotus ornatus can be easily distinguished from other species of the genus by the blister-like, enlarged, brown tubercles on whitish scutal areas I–IV; posterior margin of dorsal scutum and free tergites I–II with a row of enlarged tubercles and white spots on lateral and posterior margin of dorsal scutum and coxa IV.
Penis (Fig. 16A–B; SMF 2881): ventral plate with 3 distal pairs of long setae, 0–2 median pairs of short setae, 4–5 basal pairs of setae. Stylus almost straight (apex slightly bent), smooth. Distal ventral process shaped as an ibis head.
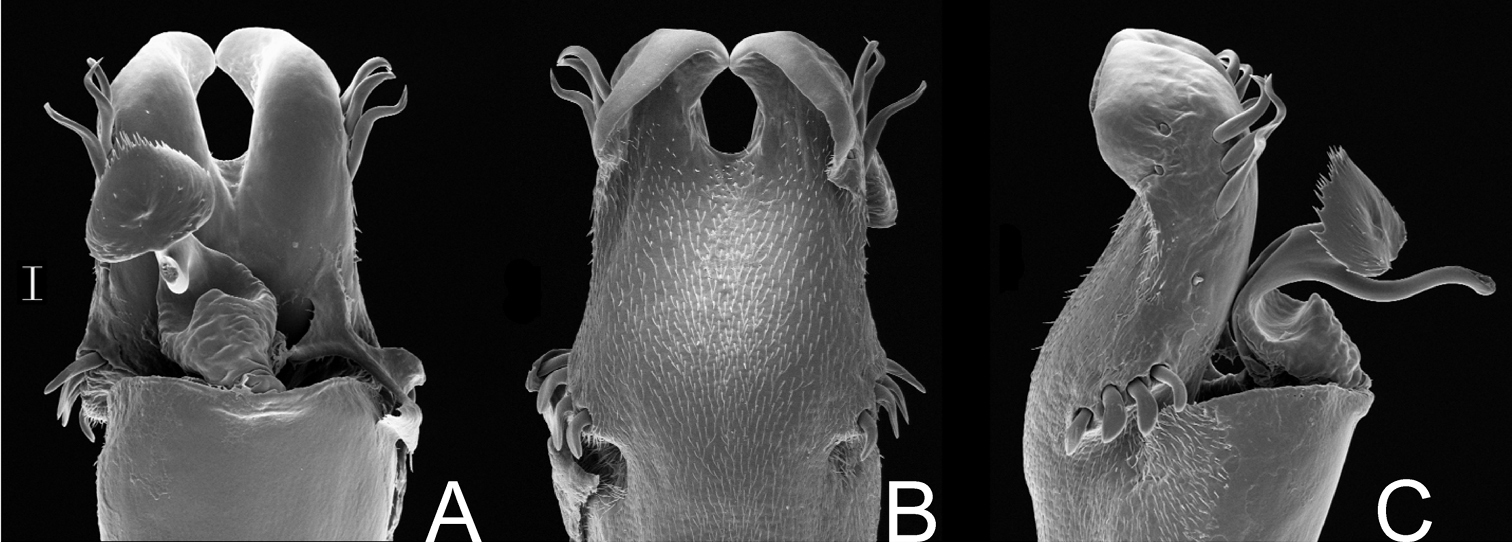
Acrographinotus ornatus (Roewer) comb. n. Distal part of penis (SMF 2881) A in dorsal view B ditto, left lateral view. Scale bar: 0.1 mm.
http://species-id.net/wiki/Gyndesops
Gyndesops resembles the largest Pachylinae genus, Discocyrtus Holmberg, 1878, which presents an ocularium and scutal area III with paired armature, four scutal areas, scutal area II–IV and free tergites I–III unarmed. This combination of characters also occurs in genera such as Gyndoides Mello-Leitão, 1927, Lacronia Strand, 1942, Paraluederwaldtia Mello-Leitão, 1927 and Parapucrolia Roewer, 1917. Gyndesops can be distinguished from Lacronia and Parapucrolia by the coloration, which is uniformly auburn (Lacronia has green/yellow spots and/or stripes on dorsal scutum and Parapucrolia has white patches on dorsal scutum). It is very difficult to distinguish Gyndesops from the remaining genera, because no revision was ever made including those groups, and their monophyly is doubtful. It is possible that Gyndesops (as well as Gyndoides and Paraluederwaldtia) is a junior synonym of Discocyrtus, but we refrained ourselves from such a rash nomenclatural act.
Monotypic.
http://species-id.net/wiki/Gyndesops_denisi
Fig. 17Penis (Fig. 17A–B, holotype): ventral plate subrectangular, basal half of lateral margin projected laterad, distal margin slightly concave, 3 distal pairs of cylindrical setae, 1 pair of median setae (long left seta and moderate sized right seta), 3 basal pairs of cylindrical setae (median largest). Glans sac almost reaching distal margin of ventral plate. Stylus with swollen apex, smooth. Ventral process with large shaft, apex flabelliform with serrated lateral margins.
Originally, Gyndesops denisi was allocated in Pachylinae, and later transferred to Gonyleptinae by
Gyndesops denisi Roewer. Distal part of penis (holotype) A in dorsal view B ditto, right lateral view. Scale bar: 0.1 mm.
http://species-id.net/wiki/Haversia
Haversia resembles Pachylinae with three scutal areas, such as Corralia Roewer, 1913, Diconospelta Canals, 1934, Graphinotus Koch, 1839, Huassampilia Roewer, 1913, Neogonyleptes Roewer, 1913, Oxapampeus Roewer, 1936, Spinivunus Roewer, 1943, Tumbesia Loman, 1899, Ubatubesia B. Soares, 1945, and two species of Sadocus Sørensen, 1886 (Sadocus conspicillatus and Sadocus polyacanthus). Haversia can be distinguished from those genera by the unarmed and smooth (i.e. without tubercles) scutal areas and the complex apophyses of male trochanter IV: three prolateral apophyses (anterior median, posterior median and an apical apophysis).
The two type species (Gonyleptes defensus and Sadocus vallentini)are synonymous, resulting in a generic synonymy, and the new assignment proposed here is based on penis morphology (see note on Gonyleptes denisi). The penis of Haversia follows the basic pattern of those gonyleptids placed in the large (and non-monophyletic) subfamily Pachylinae (ventral plate with straight anterior margin; glans without dorsal process; distal, median and basal setae of ventral plate short [seta length inferior to ventral plate width]; stylus and ventral process with variable shape). However, Haversia presents two features that stand out: (i) the penis glans projected dorsally and (ii) the detailed shape of apex of penis ventral process. Those features are typical of some Chilean genera, such as Fonckia, Neogonyleptes and Sadocus (unpublished data), and might be the evidence that those four genera are closely related.
Monotypic.
http://species-id.net/wiki/Haversia_defensa
Fig. 18ARGENTINA. Falklands (Islas Malvinas), 1 ♂ (SMF 1320/8).
Penis (Fig. 18A–B; SMF 1320/8): ventral plate with lateral margins slightly concave, distal margin straight, 3 distal pairs of cylindrical, straight, long setae, 1 pair of median short setae, 2 pairs of basal cylindrical, straight, long setae. Glans sac projected dorsally, ventral process and stylus arising ventrally. Stylus long, slightly curved, with apical ventral and lateral trichomes. Ventral process swollen apically, with nailhead-like apex.
The examination of detailed photos of Gonyleptes defensus type material and subsequent comparison of those with the description of Sadocus vallentini allowed recognizing them as conspecific.
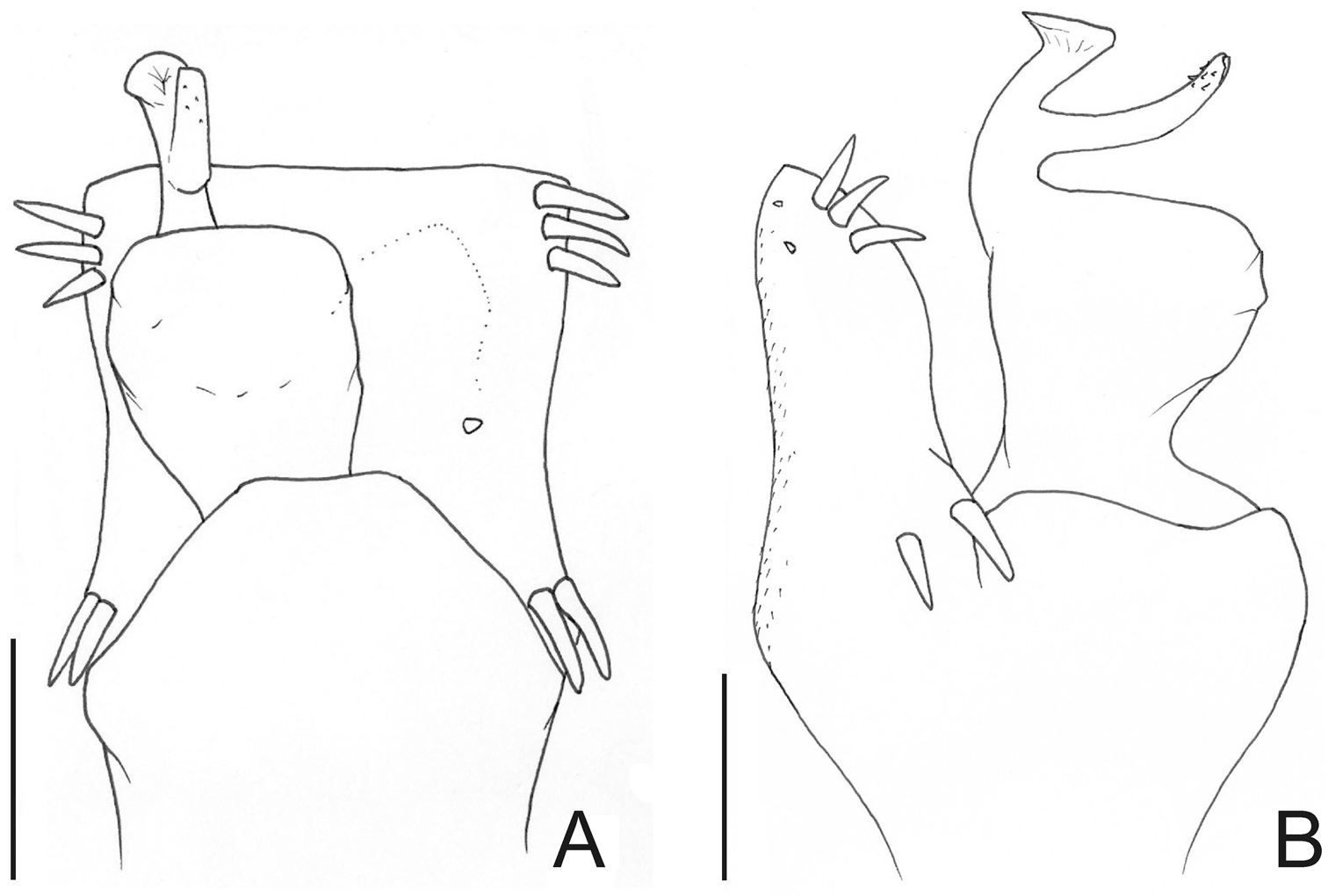
Haversia defensa (Butler). Distal part of penis (SMF 1320/8) A in dorsal view B ditto, left lateral view. Scale bars: 0.1 mm.
http://species-id.net/wiki/Oxapampeus
Based on external morphology, Oxapampeus resembles Pachylinae with three scutal areas (subfamily classical diagnosis is the presence of four areas) and scutal area III with a paramedian pair of spines, such as Corralia Roewer, 1913, Diconospelta Canals, 1934, Haversia Roewer, 1913, Huassampilia Roewer, 1913, Neogonyleptes Roewer, 1913, Spinivunus Roewer, 1943, and two species of Sadocus Sørensen, 1886 (Sadocus conspicillatus and Sadocus polyacanthus). However, Oxapampeus can be distinguished from those genera by the combination of the following characters: ocularium with a paramedian pair of pointed tubercles; frontal hump on anterior border of dorsal scutum moderately developed; scutal areas I–II each with a paramedian pair of enlarged tubercles; posterior margin of dorsal scutum straight; and free tergites I–III unarmed.
Oxapampeus has been included in Gonyleptinae due to the presence of three scutal areas on the dorsal scutum and absence of autapomorphies of other gonyleptidean subfamilies. However, its penis does not present the typical gonyleptine pattern. In addition, its ventral plate, with slightly concave lateral sides, as well as the absence of dorsally projected basal lobes, makes it resemble several Andean Pachylinae male genitalia.
Monotypic.
http://species-id.net/wiki/Oxapampeus_weyrauchi
Fig. 19Penis (Fig. 19A–B; holotype): ventral plate with lateral and distal margins slightly concave, with 3–4 distal pairs of cylindrical, straight, long setae, 0–1 median seta (visible in lateral view), 5 basal pairs of setae. Glans sac projected dorsally, ventral process and stylus arising ventrally. Stylus short and straight, with ventral subapical trichomes. Ventral process swollen apically, more elongated and thinner than stylus.
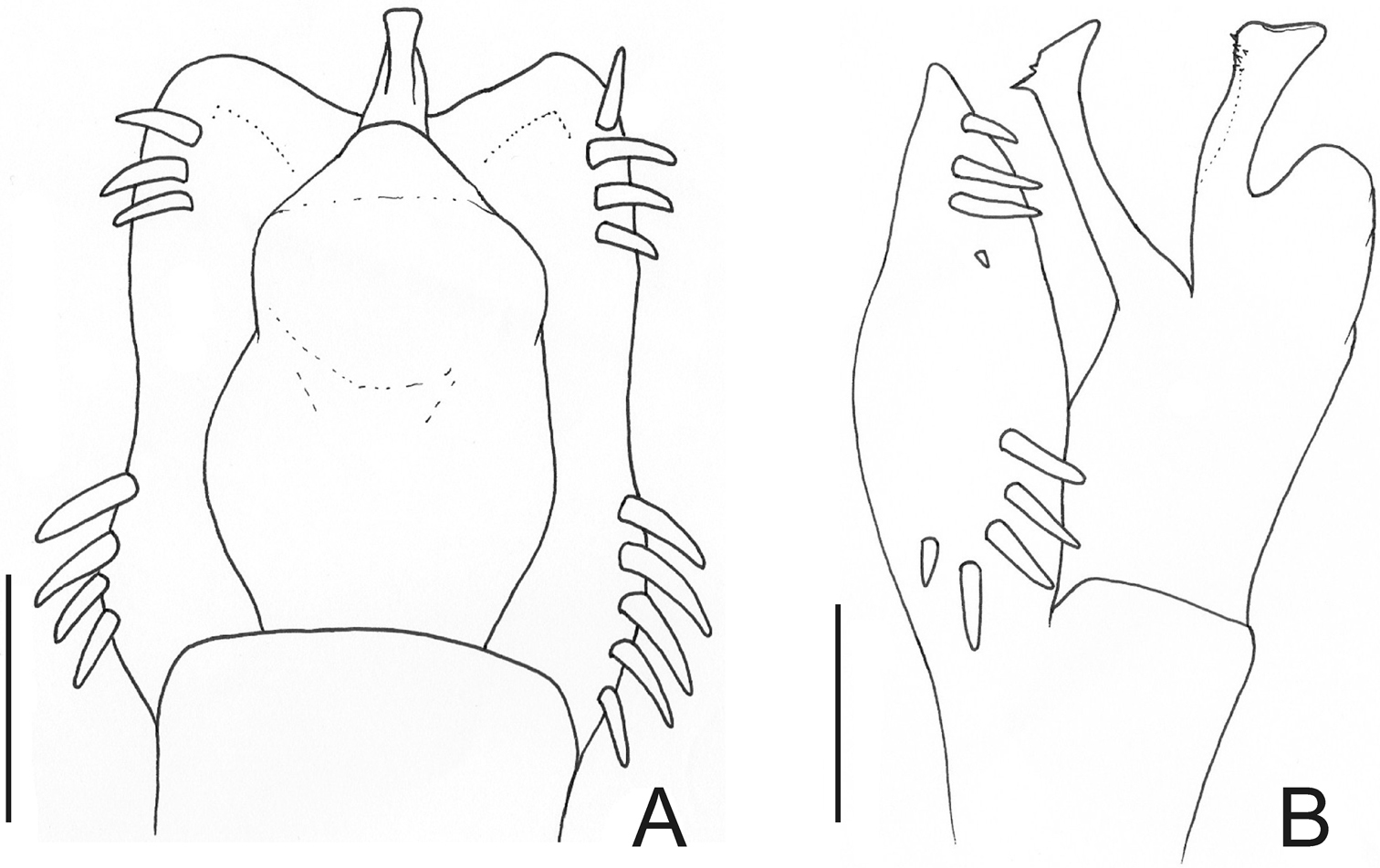
Oxapampeus weyrauchi Roewer. Distal part of penis (holotype) A in dorsal view B ditto, left lateral view. Scale bars: 0.1 mm.
We are indebted to Peter Jäger (SMF) and Adriano Brilhante Kury (MNRJ) for the loan of the examined material as well as their hospitality during visits to their institutions. These loans would not be possible without the assistance of Julia Altmann (SMF) and technical staff and students at MNRJ. Adriano Brilhante Kury is also credited for willingly making hard-to-find articles available at his own expense at “http://www.museunacional.ufrj.br/mndi/Aracnologia/pdfliteratura/pdfs%20opiliones.htm”. Cristina Rheims and Ronald M. Clouse kindly reviewed the language. Yuri Marusik and Adriano B Kury kindly improved the first version of the manuscript. The authors also thank financial support from Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP MRH #2009/3111-2, RPR #2008/06604-7) and the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq EV#; RPR #472960-2008-3).