





(C) 2011 Charlene Janion. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Two new species of Neanurinae (Collembola) are described from the Western Cape, South Africa: Ectonura monochaeta sp. n. and Ectonura barrai sp. n. Ectonura monochaeta sp. n. differs from other species in the genus by its strongly reduced chaetotaxy, and the lateral shift of dorso-internal chaetae on Abd. V and their integration in the tubercles (De+DL). Ectonura barrai sp. n. is similar to Ectonura natalensis (Womersley, 1934), but differs in chaetotaxic details and chaetal group arrangement. A key to the seven species of Neanurinae recorded from South Africa is given.
Taxonomy, new species, Western Cape, Fynbos
Neanurinae Collembola are represented in tropical Africa by a large number of species in the tribe Paleonurini, and a single representative of the tribe Neanurini, the parthenogenetic species Neanura muscorum (Templeton, 1835). However, only a few areas have been sampled outside the mountain ranges of Eastern Africa (
Among the large amount of samples recently collected in
the Western Cape Province in the frame of the Franco-South African
PROTEA project “Uncovering Springtail Diversity in the South African
Cape Floristic Region: a combined taxonomic and barcoding approach”, we
retrieved representatives of the three cited genera, including new Ectonura species, as well as a single species of a fourth genus, Paleonura Cassagnau, 1982. This confirms that South Africa fauna of Neanurinae is particularly poor, compared to that of East African mountains or Madagascar (
Ectonura is therefore the most diversified genus of Neanurinae in southern Africa. Several undescribed species were present in our samples, mostly as isolated specimens. Two of them were collected in sufficient number and are described in this paper: Ectonura monochaeta sp. n. from Table Mountain and Ectonura barrai sp. n. from Grootvadersbosch, both located in the Western Cape Province.
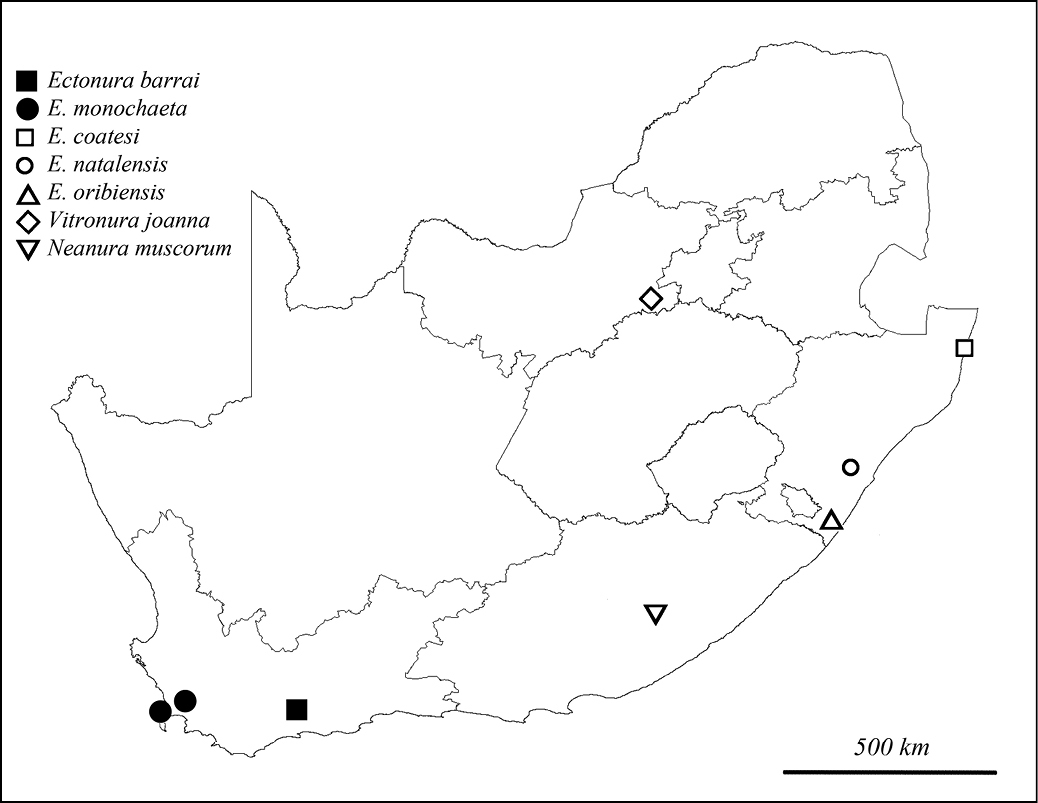
Distribution of Neanurinae recorded from South Africa.
Type deposit – IM, Iziko Museum (Cape Town, South Africa); MNHN, Museum National d'Histoire Naturelle (Paris, France).
Chaetal arrangement and notation follow
Abbreviations used in species descriptions and tables:
Tubercles and chaetal groups – Af, antenno-frontal; CL, clypeal; De, dorso-external; Di, dorso-internal; DL, dorso-lateral; L, lateral; Oc, ocular; So, subocular; VL, ventro-lateral; Ve, ventro-external; Vi, ventro-internal; Ag, antegenital; An, anal.
Appendages – Cx, coxa; Fe, femur; Fu, furcal rest; Scx2, subcoxa 2; Tita, tibiotarsus; Tr, trochanter; VT, ventral tube.
Types of chaetae – M, macrochaeta; me, mesochaeta; mi, microchaeta; S, S-chaeta (“sensillum” auct.) ; ms, s-microchaeta.
Others – omma, ommatidia; Abd., abdominal segment; Ant., antennal segment; Th., thoracic segment.
Key to South African species of Neanurinae| 1 | Three eyes on each side of the head, body bluish in colour | Neanura muscorum (Templeton, 1835) (temperate cosmopolite) |
| – | Two eyes on each side of the head, body not blue | 2 |
| 2 | Central area of head with 3 tubercles | Vitronura joanna (Coates, 1968) (uncertain status) |
| – | Central area of head with 2 tubercles | Ectonura 3 |
| 3 | Dorso-internal tubercle of tergites from Th. II to Abd. IV with at least two chaetae | 4 |
| – | Dorso-internal tubercle of tergites with one chaeta from Th. II to Abd. IV | Ectonura monochaeta sp. n. |
| 4 | Chaeta A absent on head | 5 |
| – | Chaeta A present on head | 6 |
| 5* | Modified chaetae Ag on Abd. V of the male much shorter than lateral Ag, very thick, truncated, apically ciliated | Ectonura oribiensis (Coates, 1968) (endemic) |
| – | Modified chaetae Ag on Abd. V of the male as long as lateral Ag, thin, pointed, with several unequal distal cilia | Ectonura coatesi Barra, 1994 (endemic) |
| 6 | Five chaetae on each central group of chaetae on head (1/2Af+Oc), chaeta D absent; one chaeta Di on Abd. V (Di2 absent); tubercles Di of Abd. V fused on axis | Ectonura natalensis(Womersley, 1934) (endemic) |
| – | Six chaetae on each central group of chaetae on head (1/2Af+Oc), chaeta D present; two chaetae Di on Abd. V (Di2 present); tubercles Di of Abd. V separate | Ectonura barrai sp. n. |
* The only differential character separating unambiguously Ectonura coatesi from Ectonura oribiensis is the morphology of modified chaetae Ag in the male.
TaxonomyEctonura Cassagnau, 1980
Type species: Achorutes natalensis Womersley, 1934 (Natal, South Africa)
urn:lsid:zoobank.org:act:FBCFC562-E830-4085-B711-3BCF1F4758DD
http://species-id.net/wiki/Ectonura_monochaeta
Figs 2– 3, Table 1Holotype female on slide. South Africa: Western Cape, Cape Town, Table Mountain National Park, 10/03/2009, native broadleaved forest, sieving of litter and extraction on Berlese funnel, Louis Deharveng and Anne Bedos leg (SAF-141).
8 paratypes on slides (3 males, 4 females, 1 juvenile) and more than 50 in alcohol, same data as holotype — 1 male on slide, ibid, Table Mountain, collapse of New Year cave system, 07/03/2009, native forest, litter, Berlese extraction, Louis Deharveng and Anne Bedos leg (SAF-129) — 1 male and 1 female on slides, ibid, Table Mountain, in a collapse, 10/03/2009, native forest, soil, Berlese extraction, Louis Deharveng and Anne Bedos leg (SAF-139) — 1 male and 1 female on slides, 5 specimens in alcohol, ibid, Table Mountain, Inchuk cave entrance, 10/03/2009, native forest, litter, Berlese extraction, Louis Deharveng and Anne Bedos leg (SAF-144).
Holotype, 6 paratypes on slides (3 males, 3 females) and 25 paratypes in alcohol in IM; 7 paratypes on slides (3 males, 3 females, 1 juv.) and 25 paratypes in alcohol in MNHN.
1 male on slide, 7 specimens in alcohol. South Africa: Western Cape, Stellenbosch, Jonkershoek Nature Reserve, Sosys trail, 12/03/2008, forest litter, Berlese extraction, Louis Deharveng and Anne Bedos leg (SAF-071) — 1 juvenile and 2 females on slides, ibid, Jonkershoek Nature Reserve, Sosys trail, 12/08/2010, litter, Charlene Janion leg (RSA10_JNK026 and RSA10_JNK032, 33°59.758'S, 18°57.156'E) — 1 male, 1 female and 1 juvenile on slides, about 130 in alcohol, ibid, Fish Hoek, Kalk Bay, Echo Valley forest, 05/11/2010, decaying wood of yellowwood, Berlese extraction, Louis Deharveng and Anne Bedos leg (SAF-196).
Length 0.82 – 1.1 mm (males) and 0.75 – 0.85 mm (females). Colour white in alcohol, yellow alive (SAF-196 sample). Eyes 2+2, unpigmented, small (diameter about 1.5–1.8 times that of Ocm socket, Fig. 3A). Habitus similar to Paleonura (Fig. 2A). No cryptopygy. Secondary granules rather large (the size of a mesochaeta socket). Dorsal tubercles visible but poorly delimited except on Abd. V-VI, indicated by secondary granules enlarged and irregularly arranged, without clearly developed tertiary granules. No reticulations. No plurichaetosis. Most ordinary dorsal chaetae are macrochaetae of similar length and morphology, basally swollen, straight, cylindrical, long, thick, covered in their 2/3 distal of numerous minute scales, distally sheathed, rounded apically (Figs 2A, 3A, B, C). Some dorsal mesochaetae shorter, bent, acuminate, smooth or weakly rugose on the lateral area of head, on tubercles L of tergites and on Abd. VI. No dorsal microchaetae. S-chaetae thin, 2/3 as long as or slightly shorter than closest macrochaeta (Figs 2A, 3B).
Ant. I with 7 chaetae, Ant. II with 11 chaetae, Ant. III with 16 or 17 chaetae (chaetae d4 and d5 or only d4 absent). Ant. IV organite as a short thick rod; apical bulb simple, low and fused to Ant. IV tip (Fig. 2B). Buccal cone moderately elongated. Maxilla styliform, mandible thin and bidentate with distal tooth subdivided in 2 or 3 minute cilia. Labrum elongate, rounded apically, with ventral sclerifications arc-like. Labral formula 0/2, 4. Labium with 4 basal, 3 distal and 3 lateral chaetae, and 2 minute sphaerical x papillae (as in Ectonura barrai sp. n., Fig. 5B).
Head chaetotaxy as in Table 1A and Fig. 2A. Head with 6 chaetal groups: CL, 2 (½ Af + Oc), (2 Di, 2 De), 2 (DL, L, So). Central area with B, F, G, Ocm and Ocp. Macrochaeta Ocm internal to ocular line, equally distant to omma or slightly closer to anterior omma; Ocp macrochaeta internal to and at level of posterior omma; Oca absent (Fig. 3A). Posterior area with a very faint tubercle and only 2+2 macrochaetae (Di1 and De1). Five chaetae Vi ventrally on head (Vi5 absent).
Tergite chaetotaxy as in Table 1B and Fig. 2A. Chaeta Di absent on Th. I. Tubercles De and DL separate on Abd. IV. Tubercle L of Abd. IV shift ahead the tubercle line Di-De-DL. Tubercles Di, De and DL fused on Abd. V on each side of axis. Tubercle Di of Abd. V with Di1 macrochaeta, Di2 and Di3 absent. Abd. VI not or hardly bilobed, with strong secondary granules, present even on the axis. S-chaetotaxic formula: 2+ms, 2/11111. Ventral chaetotaxy similar to that of Ectonura barrai sp. n., except the furcal rest in some specimens (Fig. 4C). Secondary sexual characters well developed in the adult male consists of chaetae Ag1 and Ag2 of Abd. V strongly thickened and serrated (Figs 2E, 3F), and chaetae of furcal rest, some Ve of Abd. IV (Fig. 3D), sometimes Ag3 of Abd. V (Fig. 3F), and 3+3 Ve of Abd. VI serrated but less strongly. In a male juvenile from Jonkershoek, chaetae Ag1 were bifid (Fig. 3E).
Microchaetae of furcal rest smaller than secondary granules, often unconspicuous (Fig. 2D). Leg chaetotaxy given in Table 1C. Tita without chaeta M and with chaetae B4-B5 short, not longer than other long chaetae of Tita (Fig. 2C). Claw untoothed, not striated in its basal part, and devoid of secondary granulation.
The species name refers to its reduced chaetotaxy of dorso-internal tubercles of tergites, which bear only one chaeta from Th. II to Abd. IV (2 or 3 in other species of the genus).
All known localities of Ectonura monochaeta sp. n. belong to the Southern Afrotemperate Forest vegetation type. The species is common in this habitat, typically found in the Western Cape, but absent in shrub formations of the fynbos. The distribution ranges from Table Mountain National Park to Jonkershoek Nature Reserve, Stellenbosch. The species is mixed in Stellenbosch with another undescribed species of Ectonura.
The new species Ectonura monochaeta is unique in the genus by the lateral shift of dorso-internal chaetae on Abd. V and their integration in the tubercles (De+DL). Such a lateral shift is only know in Ectonura paralata Deharveng, Weiner & Najt, 1997 from New Caledonia, but less marked and without integration of Di chaetae in (De+DL). By other chaetotaxic characters (Di2 and De2 present on head and on tergites of Th. II-Abd. IV; D, E, OcA present on head) and tubercle arrangement (tubercle Di not developed, others as large flat plates), Ectonura paralata is however only remotely related to our species. Ectonura monochaeta is also distinct from other species of the genus Ectonura by the strong reduction of its chaetotaxy: absence of several chaetae on head (A, O, C, D, E, Oca), absence of Di2, De2 and DL2 on tergites, only 2+2 dorsal chaetae on Th. I, and only 6+6 chaetae on Abd. VI.
The lateral shift of Di on Abd. V is one of the characteristic feature of two genera, the monotypic genus Zelandanura Deharveng & Wise, 1991 from Campbell Island and Pronura Delamare Debouteville, 1953 which is highly diversified in Africa and in Asia. Zelandanura differs from Ectonura by the fusion in one plate of all tubercles of the central area of head, and by the fusion of Di tubercles on the axis on Abd. IV. Contrary to Ectonura, chaetae of the central area of head are not separated in two groups on both side of the axis in Pronura.
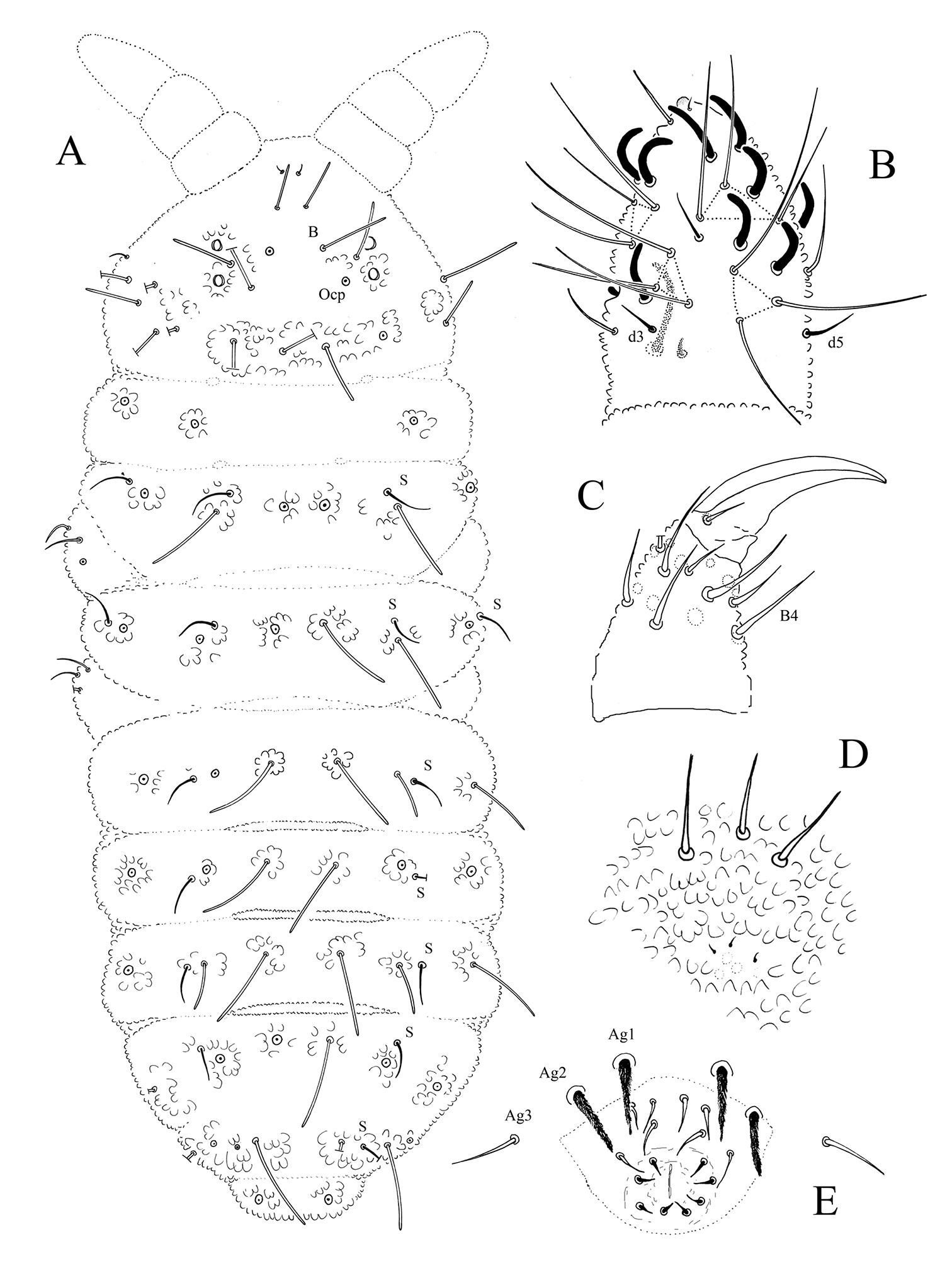
Ectonura monochaeta sp. n.; A dorsal chaetotaxy B Ant. III-IV dorsal side C tibiotarsus and praetarsus of leg I D furcal rest E male genital area.

Ectonura monochaeta sp. n.; A Ocular plate with macrochaeta Ocm B macrochaeta De1 on Th. II C chaeta B on head D chaetae Ve posterior of Abd. IV in a paratype male adult E genital plate of a male juvenile from Jonkershoek F genital plate of a male adult.
Chaetotaxy of Ectonura monochaeta sp. n.
| A-Cephalic chaetotaxy | |||||
| Group of chaetae | Tubercle | Number of chaetae | Type of chaetae | Chaetae | |
| CL | (-) | 2 | M | F | |
| 2 | me | G | |||
| 1/2Af+Oc | (+) | 3 | M | B, Ocm, Ocp | |
| 2Di, 2De | (+) | 2+2 | M | Di1, De1 | |
| DL, L, So | (-) | 5 | M | DL1, DL5, L1, L4, So1 | |
| 4 | me | So3 to 6 | |||
| B-Tergite chaetotaxy | |||||
| Di | De | DL | L | ||
| Th. I | - | 1 | 1 | - | |
| Th. II | 1 | 1+S | 1+S+ms | 3 | |
| Th. III | 1 | 1+S | 1+S | 3 | |
| Abd. I | 1 | 1+S | 1 | 2 | |
| Abd. II | 1 | 1+S | 1 | 2 | |
| Abd. III | 1 | 1+S | 1 | 2 | |
| Abd. IV | 1 | 1+S | 1 | 5 | |
| Abd. V | 3+S | 2 | |||
| Abd. VI | 6* | ||||
| C-Leg chaetotaxy | |||||
| Scx2 | Cx | Tr | Fe | Tita | |
| Leg I | 0 | 3 | 6 | 13 | 18 |
| Leg II | 2 | 7 | 6 | 12 | 18 |
| Leg III | 2 | 8 | 6 | 11 | 17 |
| D-Sternite chaetotaxy | |||||
| Abd. I | VT: 4 | ||||
| Abd. II | Ve: 4 | (Ve1 present) | |||
| Abd. III | Ve: 3 | Fu: 3-5me+3mi | |||
| Abd. IV | Ve: 7 | VL: 4 | |||
| Abd. V | Ag: 3 | VL: 1, with L | |||
| Abd. VI | Ve: 12-13 | An: 2 mi | |||
urn:lsid:zoobank.org:act:E0C421EE-4014-41F9-9A71-F23C8A7A6AEB
http://species-id.net/wiki/Ectonura_barrai
Figs 4 – 6, Table 2Holotype male on slide. South Africa: Western Cape, Grootvadersbosch Nature Reserve, Heidelberg, 24/08/2010, Southern Afrotemperate Forest vegetation, in litter, extraction on Berlese funnel, Charlene Janion leg (RSA10_GVB009, 33°59.167'S, 20°48.639'E).
1 male paratype on slide, same data as holotype — 2 paratypes on slides (1 male, 2 juveniles) and 5 paratypes in alcohol, ibid, Grootvadersbosch Nature Reserve, Heidelberg, 24/08/2010, same habitat, extraction on Berlese funnel, Charlene Janion leg (RSA10_GVB008, 33°58.964'S, 20°48.524'E)
Holotype and 4 paratypes (1 male and 1 juvenile on slides, 2 in alcohol) in IM; 5 paratypes (1 male and 1 juvenile on slides, 3 in alcohol) in MNHN.
Length 1.1–1.3 mm (males). Colour white in alcohol. Eyes 2+2, unpigmented, rather large (diameter about 3 times that of Ocm socket, Fig. 5A). Habitus similar to Paleonura (Fig. 4A). No cryptopygy. Secondary granules rather large (the size of a mesochaeta socket). Dorsal tubercles not clearly delimited, only indicated by secondary granules irregularly arranged, without tertiary granules. No reticulations. No plurichaetosis. Ordinary dorsal chaetae differentiated in macrochaetae, mesochaetae and microchaetae (Figs 4A, 6). Dorsal macrochaetae basally swollen, straight or slightly bent, subcylindrical, long, moderately thick, with minute scales sparsed apparently unilaterally, in their distal half, distally sheathed, rounded apically (Figs 5C, D, 6). Dorsal mesochaetae shorter, acuminate to blunt, smooth or weakly rugose. Dorsal microchaetae thin, smooth, less than 1/5 of macrochaetae, present on all tergites (Di2 from Th. II to Abd. V, De3 on Th. II-III, and De2 on Abd. I-V). S-chaetae thin, smooth, acuminate, 2/3 as long as or slightly shorter than closest macrochaeta (Figs 5C, 6).
Ant. I with 7 chaetae, Ant. II with 11 chaetae, Ant. III with 18 chaetae (chaetae d4 and d5 present). Ant. IV organite as a very short thick rod; apical bulb feebly trilobed, fused to Ant. IV tip (Fig. 4B). Buccal cone moderately elongated. Maxilla styliform, mandible thin and bidentate with distal tooth subdivided in 2 or 3 minute cilia. Labrum elongate, rounded and finely denticulated apically, with ventral sclerifications arc-like distally. Labral formula 0/2, 4. Labium with 4 basal, 3 distal and 3 lateral chaetae, and 2 minute sphaerical x papillae (Fig. 5B).
Head chaetotaxy as in Table 2A and Fig. 4A. Head with 9 chaetal groups: CL, 2 (½ Af + Oc), 2 DL, 2 (L, So), 2 (Di, De) on very faint tubercles hardly separated on axis. Central area with A, B, D, F, G, Oca, Ocm and Ocp (alternatively, Oca might be homologous of chaeta E). Macrochaeta Ocm internal to ocular line, equally distant from omma; Ocp macrochaeta internal to and at level of posterior omma; Oca antero-internal to Ocm (Fig. 5A). Five chaetae Vi ventrally on head (Vi5 absent).
Tergite chaetotaxy as in Table 2B and Figs 4A and 6. Chaeta Di present on Th. I. Tubercles De and DL fused on Abd. IV. Tubercle L of Abd. IV slightly shift ahead the tubercle line Di-De-DL. Tubercles De and DL fused on Abd. V, separated from Di. Tubercle Di of Abd. V with Di1 macrochaeta, Di2 microchaeta and Di3 absent. Abd. VI not or hardly bilobed, with 1+1 areas of slightly enlarged and irregularly arranged secondary granules. S-chaetotaxic formula: 2+ms, 2/11111. No modified chaetae in male (Figs 4E, 5E).
Microchaetae of furcal rest minute and thick, smaller than secondary granules (Fig. 4D). Leg chaetotaxy given in Table 2C, similar to that of Ectonura monochaeta sp. n. (Fig. 2C). Tita without chaeta M and with chaetae B4-B5 short, not longer than other long chaetae of Tita. Claw untoothed, not striated in its basal part, and devoid of secondary granulation.
This species is named in honour of Jean-Auguste Barra, for his important contribution to the knowledge of South African Collembola.
This species was collected in the yellowwood forest leaf litter of Grootvadersbosch Nature Reserve. This is a remnant forest of the larger Tsitsikamma Forest Reserve situated 300 km to the south. The forest consists of indigenous trees such as yellowwood, ironwood and stinkwood.
Ectonura barrai sp. n. is similar to Ectonura natalensis in its relatively complete chaetotaxy, but different in several details of chaetal arrangement. Based on the redescription of

Ectonura barrai sp. n.; A dorsal chaetotaxy (tubercles not represented except on Abd. IV-V; x, chaeta Di2 absent unilaterally on Th. III) B Ant. III-IV dorsal side C ventral chaetotaxy of abdomen D Furcal rest E male genital plate.
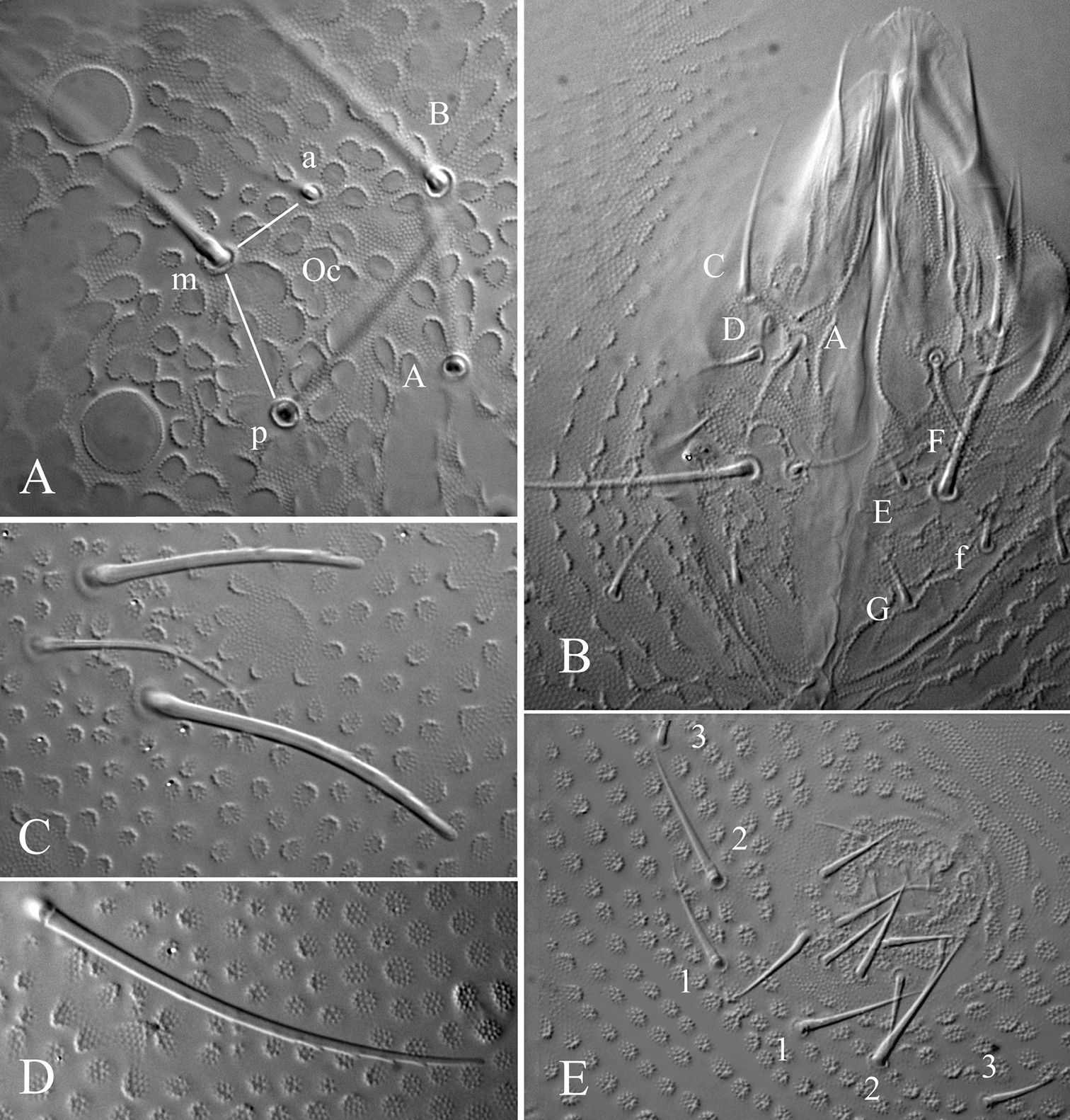
Ectonura barrai sp. n.; A ocular plate B Labium C chaetal group De on Th. III (without the De3 microchaeta) D macrochaeta DL on Abd. I E male genital plate.
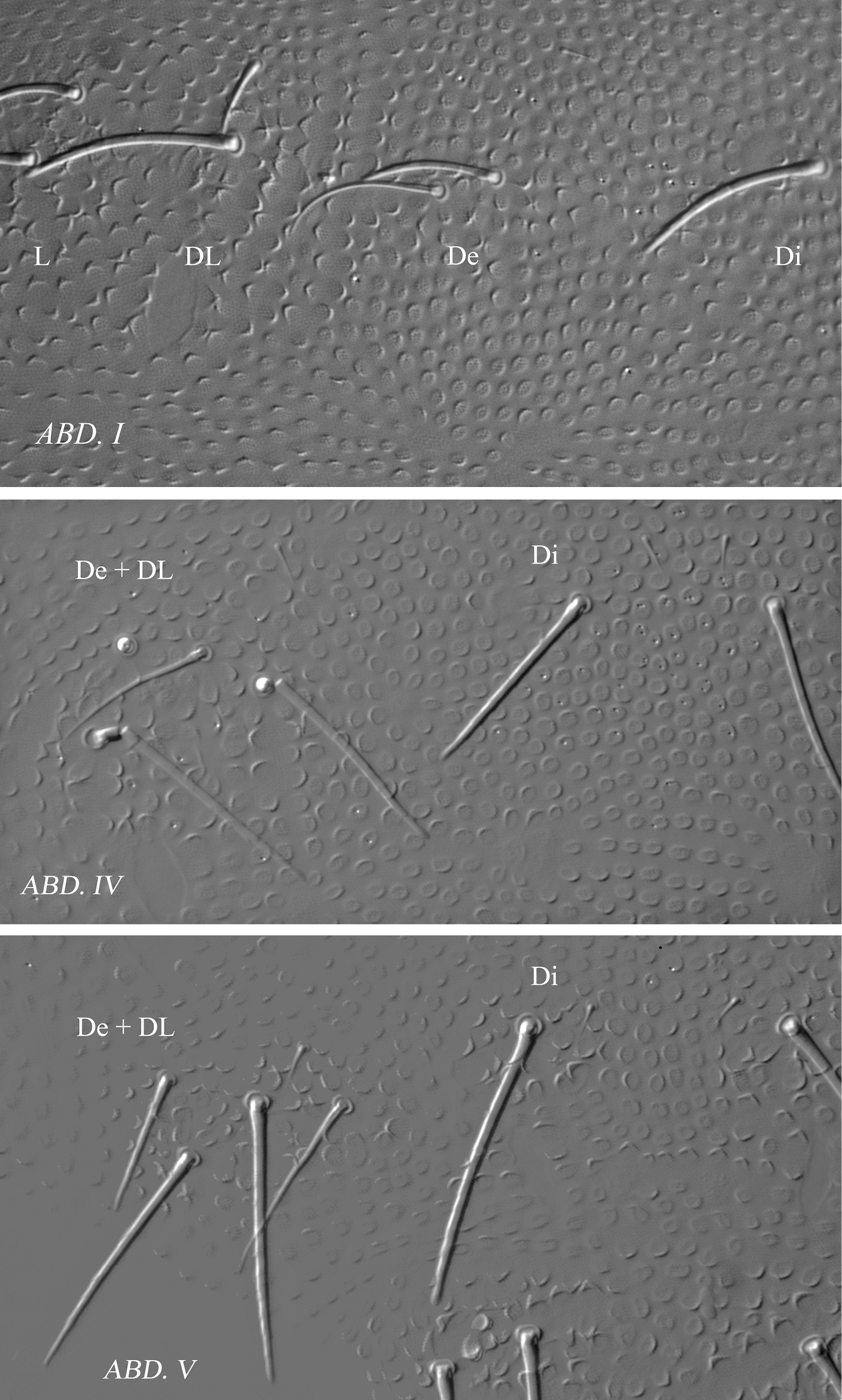
Ectonura barrai sp. n.; chaetal groups and faint tubercles Di, De and DL on Abd I, Abd. IV and Abd. V.
Chaetotaxy of Ectonura barrai sp. n.
| A-Cephalic chaetotaxy | |||||
| Group of chaetae | Tubercle | Number of chaetae | Type of chaetae | Chaetae | |
| CL | (-) | 2 | M | F | |
| 2 | me | G | |||
| 1/2Af+Oc | (+) | 4 | M | A, B, Ocm, Ocp | |
| 2 | me | D, Oca | |||
| Di, De | (+) | 2 | M | Di1, De1 | |
| 2 | mi | Di2, De2 | |||
| DL | (+) | 2 | M | DL1, DL5 | |
| 2 | me | DL3, DL4 | |||
| 1 | mi | DL2 | |||
| L, So | (-) | 3 | M | L1, L4, So1 | |
| 5 | me | L2, So3 to 6 | |||
| B-Tergite chaetotaxy | |||||
| Di | De | DL | L | ||
| Th. I | 1 | 2 | 1 | - | |
| Th. II | 2-3 | 3+S | 3+S+ms | 3 | |
| Th. III | 2-3 | 3+S | 3+S | 3 | |
| Abd. I | 2 | 2+S | 2 | 3 | |
| Abd. II | 2 | 2+S | 2 | 3 | |
| Abd. III | 2 | 2+S | 2 | 3-4 | |
| Abd. IV | 2 | 4+S | 6 | ||
| Abd. V | 2 | 4+S | 3 | ||
| Abd. VI | 7 | ||||
| C-Leg chaetotaxy | |||||
| Scx2 | Cx | Tr | Fe | Tita | |
| Leg I | 0 | 3 | 6 | 13 | 18 |
| Leg II | 2 | 7 | 6 | 12 | 18 |
| Leg III | 2 | 8 | 6 | 11 | 17 |
| D-Sternite chaetotaxy | |||||
| Abd. I | VT: 4 | ||||
| Abd. II | Ve: 4 | (Ve1 present) | |||
| Abd. III | Ve: 3 | Fu: 4me+4mi | |||
| Abd. IV | Ve: 7 | VL: 4 | |||
| Abd. V | Ag: 3 | VL: 1, with L | |||
| Abd. VI | Ve: 12-13 | An: 2 mi* | |||
We are grateful to CapeNature and SANParks for collecting permits. We thank Jacques Scheepers for field assistance and Vernon Visser for producing the map. This work was supported by the France-South Africa grant no. 68652 for L. Deharveng and A. Bedos, and the Centre for Invasion Biology for C. Janion.