





(C) 2011 Zhi-Shun Song. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
The monotypic genus Pibrocha Kirkaldy, 1902, known only from Sri Lanka in the Oriental region, is closely related to Dorysarthrus Puton, 1895 from southwestern Asia and northern Africa (Palaearctic region). The genusis revised to include a first description of the male genital structures and a discussion of relationships between Pibrocha, Dorysarthrus and Dichoptera Spinola, 1839. A diagnostic key to the three genera and photos of their type species are provided for better comparison in these taxa. Pibrocha is assigned tentatively from Dictyopharidae to the subfamily Dorysarthrinae (Fulgoridae).
Fulgoridae, Dictyopharidae, Dorysarthrinae, Pibrocha, redescription
The monotypic genus Pibrocha was established by
Traditionally, the genus Pibrocha, along with other genera Awaramada Distant, 1914, Daridna Walker, 1858, Dichoptera Spinola, 1839 and Dorysarthrus Puton, 1905, was placed in the subtribe Dichopterina (Dictyopharidae: Dictyopharinae: Dichopterini) for the presence in the forwings of a short claval crossvein between CuP and Pcu (
While attempting to clarify the distinction between Fulgoridae and Dictyopharidae,
The speces Pibrocha egregia possesses a very elongate cephalic process, which is furrowed and constricted at its basal 1/3, and appears to be ‘fractured’ and separated into two portions by an articulation (Figs 1, 4). In many dead dried specimens, the distal portion of cephalic process is easily broken, so the species may be easily misidentified. As an example the monotypic genus Awaramada Distant was established based on Pibrocha specimen that had lost the distal portion of the cephalic process. Its type species Awaramada fryeri Distant, 1914 was synonymized with Pibrocha egregia by Liang based on examination of type material in the Natural History Museum, London, UK (BMNH) (
This study provides a review of the genus Pibrocha, including a first description of the male genital structures and a discussion of relationships between Pibrocha, Dorysarthrus and Dichoptera. A key to three genera and photos of their type species are also provided for better comparison in these taxa. Pibrocha is assigned tentatively to the subfamily Dorysarthrinae (Fulgoridae) from Dictyopharidae.
Materials and methodsThe male genitalia were cleared in 10% KOH at room temperature for ca. 12 hours, rinsed in distilled H2O, then transferred to glycerol for examination.
Morphological characters were observed with a Zeiss (Stemi SV II) optical stereomicroscope and illustrated with the aid of a drawing tube; measurements were made with the aid of an eyepiece micrometer.
The specimens studied in the course of this work are deposited in the following institutions whose names are abbreviated in the text as follows:
BMNH - the Natural History Museum, London, UK;
MNHN
NCSU - Department of Entomology Insect Collection, North Carolina State University, Raleigh, North Carolina, USA;
USNM - the National Museum of Natural History, Washington, D.C., USA.
The morphological terminology used in this study follows
| 1 | Body very large and stout (large-sized species), body length (including forewings) usually more than 25 mm; head distinctly short, produced in a short or moderately long cephalic process, which is only 1/4 to half as long as pronotum and mesonotum combined (Fig. 3); cephalic process with apical portion before eyes abruptly narrowing to conic and distinctly upturned (Fig. 6); forewings with M vein first branching to MA and MP veins near base, and MP vein branching to MP1 and MP2 veins near basal 1/5 or 1/4 before nodal line; Sc+R, M and CuA veins branching to dozens of accessory veins beyond nodal line in forewings (Song and Liang, in prep.) | Dichoptera Spinola (Dichopterinae) |
| – | Body relatively much smaller and slender (medium-sized species); head very elongate and distinctly stout, produced anteriorly into a cephalic process, which is about twice as long as pronotum and mesonotum combined; cephalic process stout and cylindrical at basal 1/3, and then suddenly furrowed and constricted, which looks like being fractured and separated into two portions by an articulation; the distal remainder 2/3 turned downwards in lateral view (Fig. 8); forewings with M vein only branching to MA and MP veins near middle before nodal line; Sc+R, M and CuA veins branching to less accessory veins beyond nodal line in forewings | 2 (Dorysarthrinae) |
| 2 | Cephalic process with distal remainder 2/3 inflated and subcylindrical, which is rounded and bulbous apically in dorsal view (Fig. 2); basal 1/3 of vertex without median carina, along with a broad white median band extending over pronotum and mesonotum; frons nearly parallel before postclypeus; pronotum and mesonotum bicarinate in middle disc, lateral carinae barely visible and median carina absent; hind tibiae with 7 apical black-tipped spines | Dorysarthrus Puton |
| – | Cephalic process with distal remainder 2/3 mostly narrowed and laterally compressed, gradually expanded and dorsoventrally compressed near apex, which is truncate and clavate apically in dorsal view (Fig. 1); basal 1/3 of vertex with median carina distinct and complete; frons widest and obtusely expanded outwards before postclypeus; pronotum and mesonotum tricarinate in middle disc, median and lateral carinae distinct and complete; hind tibiae with 6 apical black-tipped spines | Pibrocha Kirkaldy |
Subfamily Dorysarthrinae Emeljanov, 1979
http://species-id.net/wiki/Pibrocha
Cephalic process twice as long as pronotum and mesonotum combined, furrowed and constricted at basal 1/3, where it appears to be ‘fractured’ and separated into two portions by an articulation; the distal remainder 2/3 mostly narrowed and laterally compressed, gradually expanded and dorsoventrally compressed near apex, which is truncate and clavate in dorsal view, and turned downwards in lateral view; vertex with basal 1/3 broad and moderately arched, median carina distinct and complete; the remainder 2/3 of vertex and frons without median carina; pronotum and mesonotum tricarinate, nearly parallel; forewings elongate and slender, nearly four times as long as broad; M vein only branching to MA and MP veins near front-middle before nodal line and firstly branched before Sc+R and CuA veins near middle; clavus with a short crossvein, connecting CuP with Pcu; legs narrow and moderately long; fore femora not flattened and dilated, hind tibiae with 6 apical black-tipped spines; aedeagus large and symmetrical, with a pair of long and slender endosomal processes extended dorsally; phallobase basally sclerotized and pigmented, without spine.
Head very elongate and distinctly stout, produced anteriorly into a cephalic process, which is about twice as long as pronotum and mesonotum combined. Cephalic process stout and cylindrical at basal 1/3, and then suddenly furrowed and constricted, where it appears to be ‘fractured’ and separated into two portions by an articulation; the distal remainder 2/3 mostly narrowed and laterally compressed, gradually expanded and dorsoventrally compressed near apex, which is truncate and clavate in dorsal view (Fig. 7), and turned downwards in lateral view (Fig. 8). Vertex with basal 1/3 broad and moderately arched, lateral carinae nearly sub-parallel and median carina distinct and complete; the remainder 2/3 narrowly sulcate, nearly parallel, gradually expanded and apically truncate, median carina indistinct in groove. Frons (Fig. 9) without median carina, intermediate carinae shallowly sulcate, nearly parallel; basal 1/3 widest and obtusely expanded outwards before postclypeus, lateral carinae slightly converging towards apex; the apical remainder 2/3 laterally compressed and abruptly narrowed. Postclypeus and anteclypeus convex medially, median carina indistinct. Rostrum long, reaching beyond abdominal segment V. Eyes oval and large. Ocelli large, reddish. Antennae with scape very small; pedicel large and subglobose, with more than 50 distinct sensory plaque organs distributed over entire surface; flagellum long, setuliform.
Pronotum (Fig. 7) a little shorter than mesonotum medially, narrow anteriorly, broad posteriorly; anterior margin slightly arched centrally, lateral marginal areas straight and sloping with two long lateral carinae on each side between eyes and tegulae, posterior margin very broadly concave; disc tricarinate in middle, median and intermediate carinae distinct and complete, with a big lateral pit at side of median carina, respectively. Mesonotum (Fig. 7) tricarinate in disc, nearly parallel. Forewings (Fig. 10) elongate and slender, nearly four times as long as broad; anterior and posterior margins more or less parallel, apex rounded; M vein only branching to MA and MP veins near front-middle before nodal line and firstly branched before Sc+R and CuA veins near middle; apical area with at least three rows of transverse veinlets, veinlets usually not aligned, but in each field running along its length; clavus with a short crossvein, connecting CuP with Pcu; stigma broad and distinct, with 3–5 cross veins. Legs narrow and moderately long; fore femora not flattened and dilated, hind tibiae with 4 lateral and 6 apical black-tipped spines; hind tarsomeres I with about 8–9 and tarsomeres II with about 6–7 black-tipped apical spines, respectively.
Sri Lanka.
http://species-id.net/wiki/Pibrocha_egregia
Figs 1, 4, 7–16Male, narrow and elongate, body length (from apex of cephalic process to tip of forewings) 21.3–21.5 mm; length of head (including two portions: the former is from apex of cephalic process to curved part, the latter is from curved part to base of eyes) (3.2+5.5)–(3.3+5.4) mm, width (including eyes) 1.8 mm; length of forewings 11.8–12.5 mm.
Vertex, genae and frons dull brownish-ochraceous, speckled with fuscous, suffused with testaceous-red. Basal 1/3 of frons with some small fuscous spots between intermediate carinae and lateral carinae. Pronotum and mesonotum brownish-ochraceous, tens of punctate spots on each lateral area of pronotum fuscous. Thorax ventrally and legs pale ochraceous. Forewings and hindwings hyaline, venation fuscous, stigma and scattered apical maculate markings on forewings and hindwings fuscous. Abdomen dorsally brownish ochraceous, ventrally paler, with numerous small fuscous spots.
Male genitalia: pygofer slightly broad, nearly rectangular, ventrally distinctly broader than dorsally (about 3.0:1) in lateral aspect (Fig. 12); posterior margin deeply excavated apically to accommodate anal tube, with a long, fingerlike, directed posteriorly process near apex in lateral view (Fig. 12); dorsal margin deeply excavated to accommodate anal tube, dorsal-lateral margins produced posteriorly in dorsal view (Fig. 13). Segment X (anal tube) narrow and elongate, with ratio of length to width near middle about 3.0:1; apical ventral margin protruded an angle on each side, apical dorsal margin deeply excavated to accommodate anal style in dorsal views (Fig. 13); epiproct relatively robust and long. Gonostyles large and broad, without spiniform setae on inner surfaces in basal half; narrow basally, broadest medially and reduced towards apex in lateral view (Fig. 12); upper margin with a small, obtuse process near upper middle, outer upper edge with a ventrally directed, hooklike process near middle in lateral aspect (Fig. 12). Aedeagus (Figs 14–16) large and symmetrical, with a pair of long and slender endosomal processes extended dorsally: basal 2/3 sclerotized and pigmented, apical 1/3 membranous; phallobase basally sclerotized and pigmented, with a pair of ventral angular lamellar processes which its edge membranous, without spine (Figs 15, 16).

1 Pibrocha egregia (Kirby), ♂, dorsal view 2 Dorysarthrus mobilicornis Puton, holotype ♀, dorsal view 3 Dichoptera hyalinata (Fabricius), ♂, dorsal view. Scale bars: Figs 1–3 = 1 mm.
Holotype ♂ of Awaramada fryeri Distant, [Sri Lanka]: (1) Kandy, Ceylon, 7-02; (2) [red label] Type / H.T.; (3) [Distant’s handwriting] Awaramada fryeri Distant.
SRI LANKA: 1♂, Ceylon, Udawattekelle, 1966.X.30, no collector; 1♂, Udawattekelle, Kandy, 1966.XI.10–13, no collector (both in USNM); 2♀♀, 1♂, [MNHN(EC)7458, 7459, 7460], Perad (=Peradeniya), Ceylan, Coll. Bugnion, Th. Bourgoin det. 1990; 1♂, Kandy, 7.02. Ceylon, Coll. Bugnion [MNHN(EC)7461], Th. Bourgoin det. 1990; 1♂, Kandy, 6.05. Ceylon, Coll. Bugnion [MNHN(EC)7562], Th. Bourgoin det. 1990 (all in MNHN).
Sri Lanka.
http://species-id.net/wiki/Dorysarthrus
The genus Dorysarthrus was established by Puton in 1895 based on a single species, Dorysarthrus mobilicornis Puton, 1895 from Palestine. Now Dorysarthrus comprises four species, namely Dorysarthrus alfierii De Bergevin, 1923 (not ‘1924’ as stated by
http://species-id.net/wiki/Dorysarthrus_mobilicornis
Figs 2, 5Holotype ♀, [PALESTINE]: (1) [Puton’s handwriting] Dorysarthrus mobilicornis Put; (2) [Puton’s handwriting] Jerusalem; (3) ♀; (4) [red label] Type; (5) TH BOURGOIN det. 1990, [Bourgoin’s handwriting] Dorysarthrus mobilicornis PUTON, 1895; (6) MNHN-HF-90-106; (7) Museum Paris, MNHN(EH), 452 (MNHN).

4 Pibrocha egregia (Kirby), ♂, lateral view 5 Dorysarthrus mobilicornis Puton, holotype ♀, lateral view 6 Dichoptera hyalinata (Fabricius), ♂, lateral view. Scale bars: Figs 4–6 = 1 mm.
Palestine, Syria.
http://species-id.net/wiki/Dichoptera
The genus Dichoptera was erected by Spinola in 1939 as one of five dictyopharid genera for the family Dictyopharidae. A total of eleven species are included in the genus, which is restricted in the Oriental region. The genus Dichoptera was moved by
http://species-id.net/wiki/Dichoptera_hyalinata
Figs 3, 6INDIA: 1♂, Chittoor, 1940.IX., P.S. Nathan (NCSU).
Bangladesh, India, Sri Lanka.
According to the diagnostic key and photos of the type species of the three genera Pibrocha, Dorysarthrus and Dichoptera, it seem obvious that Pibrocha may be more closely related to Dorysarthrus than Dichoptera. Pibrocha and Dorysarthrus share some synapomorphies from the following characters: the medium-sized species, much smaller and slenderer than Dichoptera species; the very elongate, nearly fractured cephalic process and a similar forewing venation. These distinct characters support well the monophyly of Pibrocha and Dorysarthrus, and they are assigned together in the subfamily Dorysarthrinae.
In view of the problems of defining the distinctiveness between Fulgoridae and Dictyopharidae, Dorysarthrinae is tentatively preserved in Fulgoridae based on
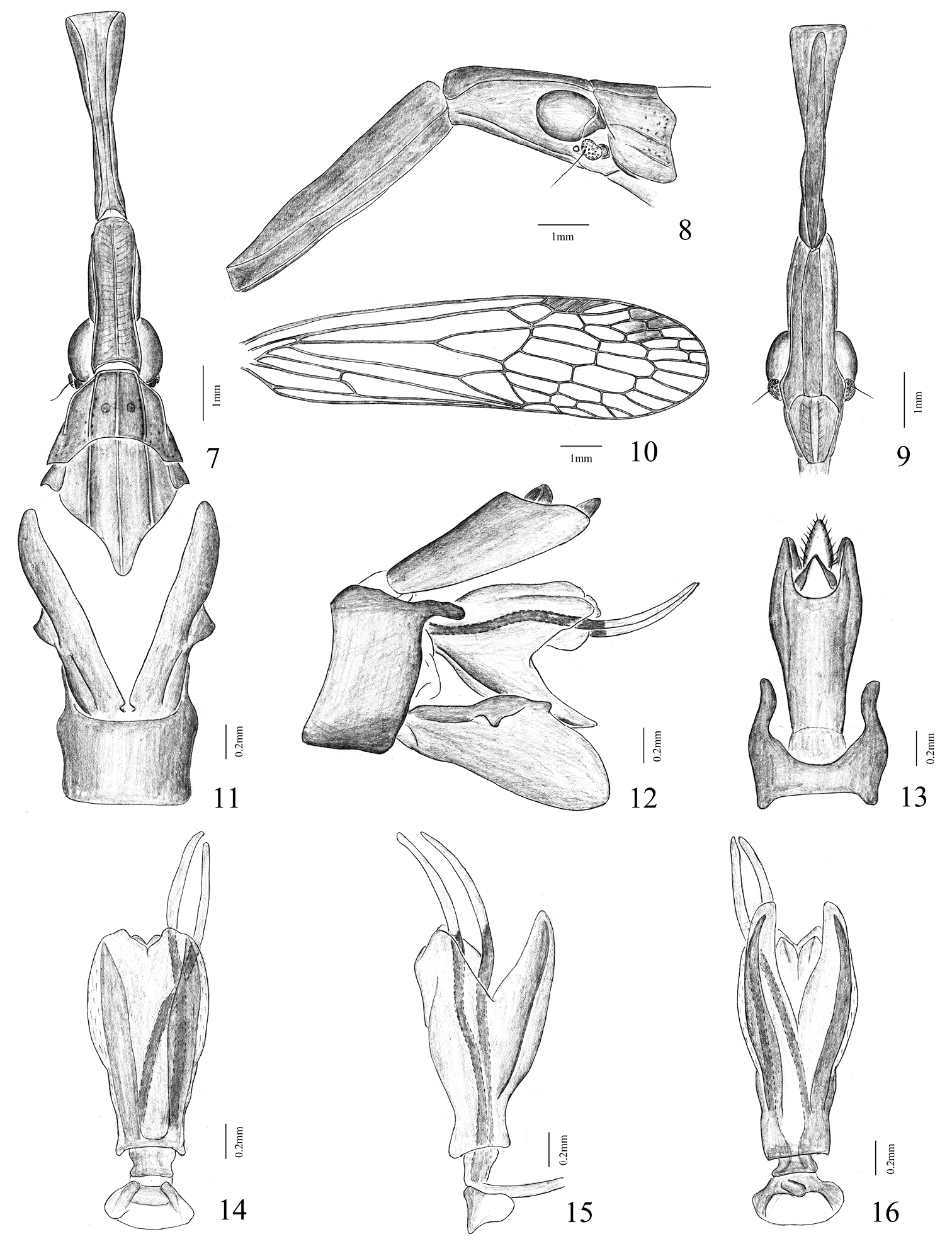
Pibrocha egregia (Kirby, 1891) 7 head, pronotum and mesonotum, dorsal view 8 head and pronotum, lateral view 9 head, ventral view 10 right forewing 11 pygofer and parameres of male, ventral view 12 genitalia of male, lateral view 13 pygofer and anal tube of male, dorsal view 14 aedeagus, dorsal view 15 aedeagus, lateral view 16 aedeagus, ventral view. Scale bars: Figs 7–10 = 1 mm, Figs 11–16 = 0.2 mm.
We are grateful to the following individuals and institutions for loans of specimens or access to collections: Drs Richard C. Froeschner and Thomas J. Henry (USNM), Mr Robert L. Blinn and Dr Lewis L. Deitz (NCSU) and Mr Mick Webb (BMNH). We thank Laurent Fauvre (MNHN) for the photos of Dorysarthrus mobilicornis Puton, 1895. We also wish to thank Dr Michael Wilson, Department of Biodiversity & Systematic Biology, National Museum of Wales, Cardiff, UK, for his very kind editorial help with this paper. Three anonymous reviewers are greatly appreciated for their efforts in improving this paper.
The work on which this paper is based was supported by the following sources: National Natural Science Foundation of China (No. 31101657), Scientific Survey on the Middle- and Lower-reaches of Lancang (Mekong) River and Grand Shangri-La Area (No. 2008FY110300), National Science Fund for Fostering Talents in Basic Research (Special subjects in animal taxonomy, NSFC-J0630964/J0109) and a grant (No. O529YX5105) from the Key Laboratory of the Zoological Systematics and Evolution of the Chinese Academy of Sciences.