





(C) 2011 Jakub Prokop. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Three new palaeopteran insects are described from the Middle Permian (Guadalupian) of Salagou Formation in the Lodève Basin (South of France), viz. the diaphanopterodean Alexrasnitsyniidae fam. n., based on Alexrasnitsynia permiana gen. et sp. n., the Parelmoidae Permelmoa magnifica gen. et sp. n., and Lodevohymen lapeyriei gen. et sp. n. (in Megasecoptera or Diaphanopterodea, family undetermined). In addition the first record of mayflies attributed to family Syntonopteridae (Ephemeroptera) is reported. These new fossils clearly demonstrate that the present knowledge of the Permian insects remains very incomplete. They also confirm that the Lodève entomofauna was highly diverse providing links to other Permian localities and also rather unique, with several families still not recorded in other contemporaneous outcrops.
Insecta, Palaeoptera, Syntonopteridae, Alexrasnitsyniidae fam. n., Parelmoidae gen. n., sp. n., Middle Permian, palaeodiversity
Before the tremendous effort of collect of Dr Jean
Lapeyrie who brought together a large collection of thousands of
fossils, the ‘red’ Middle Permian Salagou Formation (Lodève Basin) was
considered devoid of fossils. Palaeoenvironment of Lodève Basin could be
characterized as climate controlled playa with ephemeral pools
colonized by aquatic clam shrimps (Conchostraca) and tadpole shrimps
(Notostraca) together with insects (
We describe herein four new fossil insects from the
Salagou Formation those correspond to important new taxa. Unfortunately
the great majority of this entomofauna is represented by isolated wings.
The insect bodies have been destroyed by the abundant necrophagous
animals (mainly Triops Schrank, 1803) and the decay due to taphonomic process (
The material described in the present paper comes from
the Middle Permian deposits of Salagou Formation found in several
localities in the Lodève Basin, Hérault, France (
The material was observed under stereomicroscope Olympus SZX-9 and venation pattern drawings were drawn directly through stereomicroscope by camera lucida. Photographs were made using digital camera Nikon D80 with macro lens Nikon AF-S VR Micro-Nikkor 105 mm by single sided cross-light exposure.
We follow the wing venation nomenclature of
Family Syntonopteridae Handlirsch, 1911
Figs 1A–B
Specimen Ld LAP 483 (Lapeyrie collection, imprint of proximal part of forewing), stored at the Musée of Lodève, France.
Middle Permian, Guadalupian, Lodève Basin, Salagou Formation, Lodève, Hérault, France (
Counter-imprint of a fore wing without trace of preserved coloration, strongly developed corrugation of longitudinal veins; basal part of fore wing 22.7 mm long and 11.6 mm wide, estimated total length about 45 mm; area between ScP and C rather broad with numerous simple cross-veins; concave ScP straight and basally running close to radial and medial veins; RA nearly straight; RA and RP extremely approximate between wing base and a point situated 17.5 mm distally at which RP strongly diverges from RA; convex MA diverging from MP nearly at right angle and directed towards radial veins, 11.3 mm from wing base; MA distally closely parallel to RA and RP for 5.4 mm, then nearly touching RP at one point; RP emerging 6.4 mm distal of base of MA; concave MP nearly straight; convex CuA diverging from CuP close to wing base and running parallel to medial veins, CuA with two visible terminal branches; simple CuP strongly concave and only weakly curved; anal area partly preserved, first anal vein of neutral polarity ending with two main branches, second anal vein convex and distally pectinate with several branches connected by rather dense network of cells; along posterior wing margin a broad area between CuP and first anal vein; a small pentagonal elongate cell below second anal vein, near wing base.
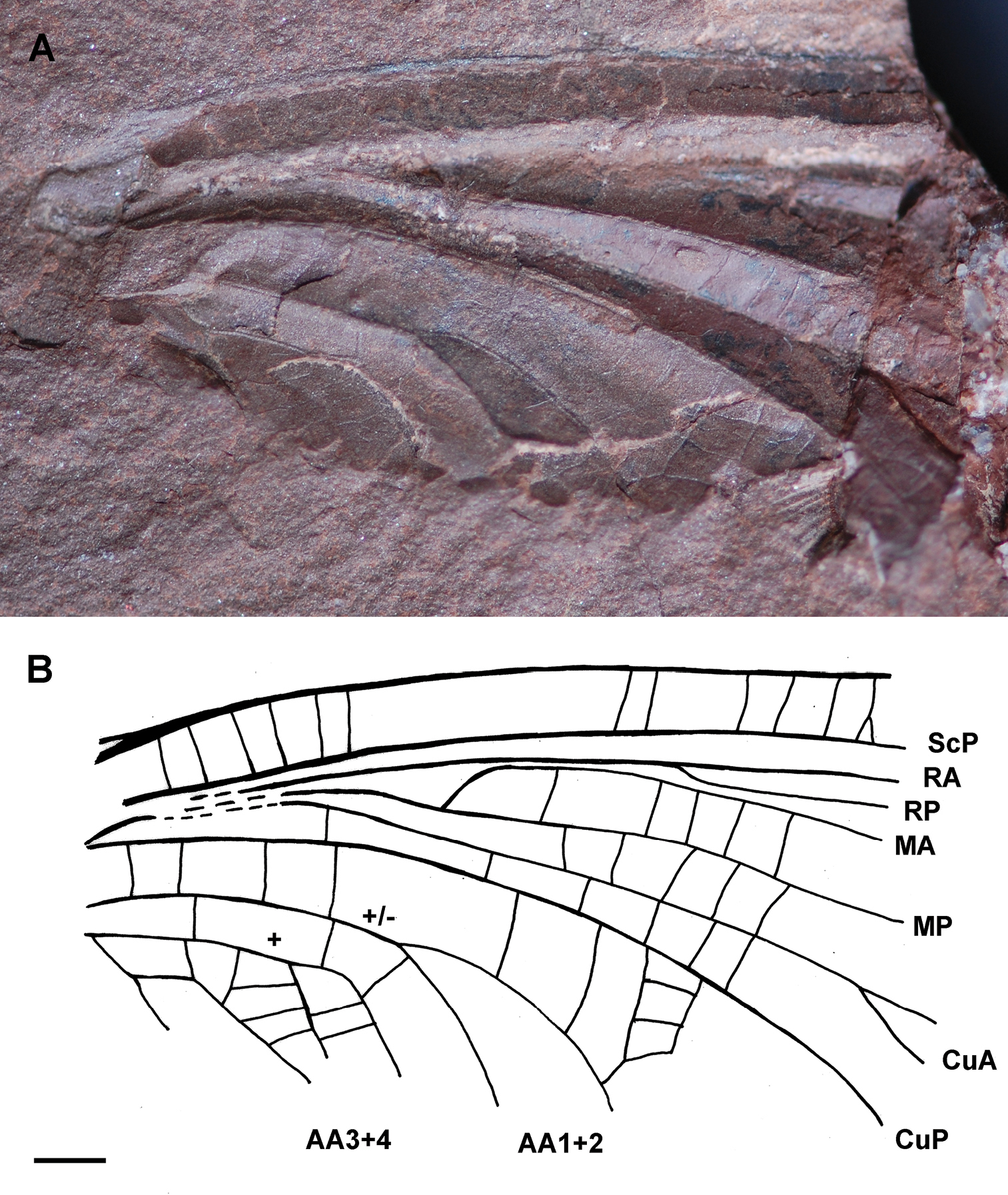
Genus and species indet.(Ephemeroptera: Syntonopteridae), specimen Ld LAP 483: A photograph of forewing B drawing of forewing (Scale bar represents 2 mm).
This fossilbears a combination of the main characters of ephemeropterid family Syntonopteridae, i.e., a strong corrugation of the main longitudinal veins connected by mainly simple transverse crossveins also present in Odonatoptera and some Palaeodictyopterida (e.g., Breyeriidae),
MA with a strong anterior curve at its base, shortly connected with
RP distally; CuA with a short terminal twigging and anal area with
well-defined cell(s). We can argue that this fossil is a forewing
fragment for the absence of a characteristic constriction of concave
vein IN- between AA1+2 and AA3+4 (
Alexrasnitsynia permiana gen. et sp. n.
Wing venation only. ScP ending on anterior wing margin near mid part of wing; stems of CuA, M and R very close; CuA, MP and MA diverging at the same point; MA and radial stem very closely parallel; MA distally fused for a short distance with RP; a very broad area between RA and anterior wing margin with several long simple oblique crossveins; a very broad area between CuA and CuP; CuA with weak secondary posterior branches.
urn:lsid:zoobank.org:act:C0239FF9-E345-4B7A-8146-33A90B85659B
http://species-id.net/wiki/Alexrasnitsynia_permiana
Figs 2A–BHolotype LdLAP 318A (Lapeyrie collection, prints of two identical wings), stored at the Musée of Lodève, France.
Middle Permian, Guadalupian, Mérifons Member, Salagou Formation, Lodève, Languedoc, France (
That of the family.
Wing 11.8 mm long, 3.9 mm wide; ScP simple, ending in costal margin near mid part of wing, a narrow area between it and C without visible crossveins; area between C and RA relatively broad, 0.6 mm wide, with a row of simple oblique crossveins; RA simple ending on anterior wing margin 0.4 mm from wing apex; RP separating from RA 2.6 mm from wing base; RP with six-seven branches and covering a broad area with series of crossveins; CuA, MP, and MA diverging at the same point, 1.3 mm from wing base; MA very closely parallel to radial stem and reaching RP, fused with it for 0.6 mm, and separating again distally, MA simple, slightly curved; MP simple; CuA with three short distal branches; a series of crossveins between MP and CuA; a broad area between CuA and CuP with crossveins; one anal vein preserved at least.
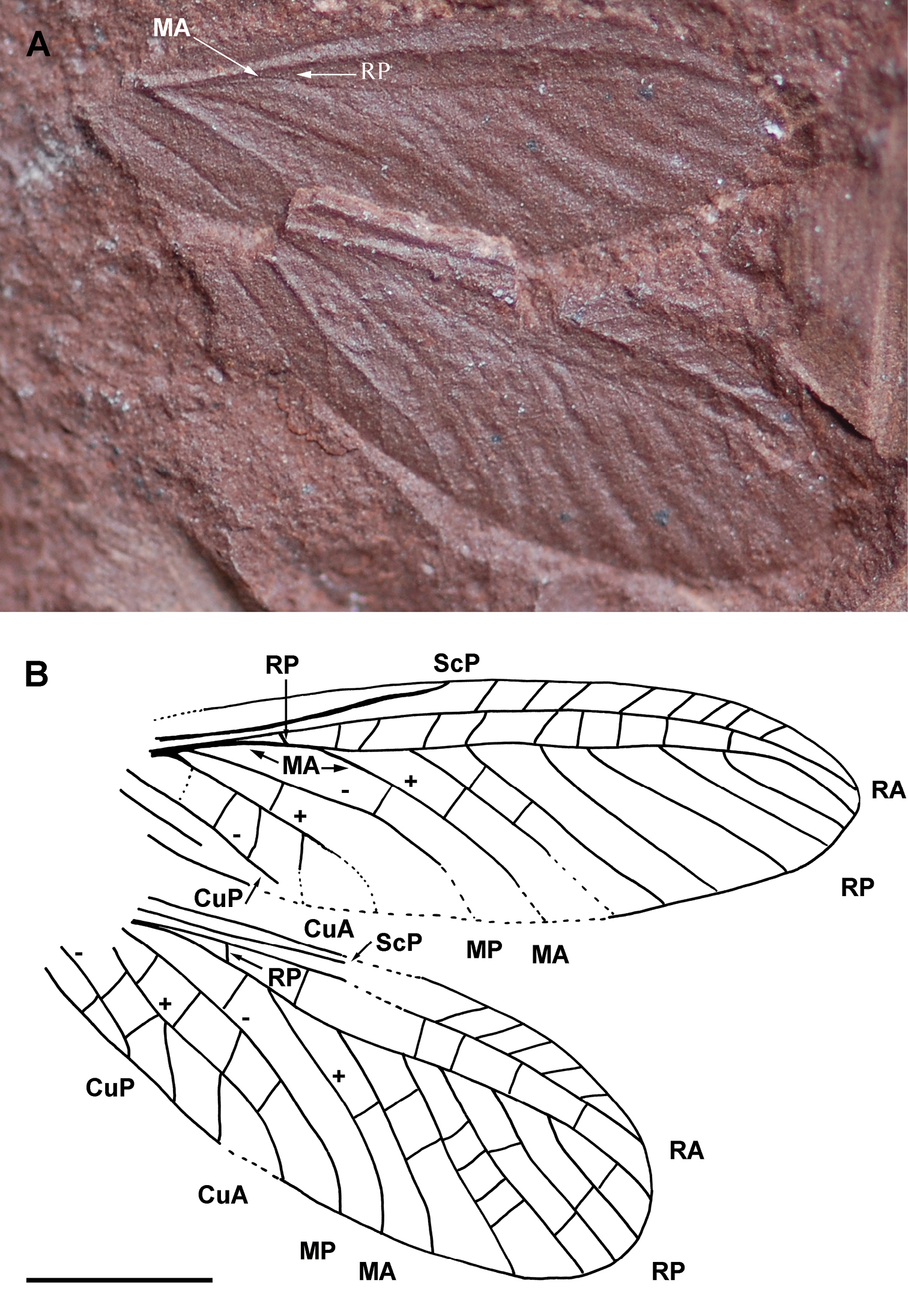
Alexrasnitsynia permiana gen. et sp. n. (Diaphanopterodea: Alexrasnitsyniidae fam. n.), holotype LdLAP 318A: A photograph of wings, B drawing of wings (Scale bar represents 3 mm).
As the two wings show the same convexity of
the veins and are very close, they are likely to be a fore- and a
hindwing of the same specimen, but it is impossible to determine which
one is the forewing. Alexrasnitsynia has a pattern of wing venation characteristic of many Diaphanopterodea (Martynoviidae, Biarmohymenidae, Asthenohymenidae, Rhaphidiopsidae,
etc.), i.e. crossveins distinct, archedictyon absent, stems of M
and R very close; CuA, MP and MA diverging at the same point; MA very
closely parallel with radial stem and distally fused for a short
distance with RP. Alexrasnitsynia differs from all the known representatives of this order, except Parelmoa Carpenter, 1947 and Permuralia
Sinichenkova & Kukalová-Peck, 1997 in the very broad area between
RA and anterior wing margin with several long simple oblique crossveins.
It differs from these two last genera in vein MA strongly approximating
radial stem, a very broad area between CuA and CuP, and CuA with weak
secondary posterior branches (
Note that the Upper Carboniferous monotypic family Velisopteridae Pinto & Adami-Rodrigues, 1997 is based on a fossil that seems to have none of the diaphanopterid characters listed above (
Alexrasnitsynia bears similar pattern of wing venation to monotypic Sypharoptera Handlirsch, 1911, based on Sypharoptera pneuma known from the Upper Carboniferous of Mazon Creek (USA).

Sypharoptera pneuma Handlirsch, 1911(?Diaphanopterodea: Sypharopteridae), holotype YPM No. 0064 (Upper Carboniferous, Carbondale Formation, Mazon Creek (Illinois, USA): photograph (Scale bar represents 5 mm).
Sypharoptera shares with Alexrasnitsynia rather short ScP, RA and RP widely separated and connected by a series of transversal crossveins, but the organisation of RP, MA, MP, and CuA at their bases is unknown in the former. So we cannot be sure of its possible affinities with our fossil. In particular, it is not possible to determine on the photograph of the holotype of Sypharoptera if it has a MA very closely parallel to radial stem and reaching the short oblique basal part of RP, fused with it for short distance, and separating again distally, as in Alexrasnitsynia.
The other structures of wing venation are similar in Sypharoptera and Alexrasnitsynia, with the main difference in the greater number of branches of RP in the latter (6–7 branches) than in the former (3–4 branches). One might be tempted to consider that Alexrasnitsynia and Sypharoptera belong to the same family of Diaphanopterodea. Nevertheless several important structures diagnostic to the family to which Alexrasnitsynia belongs are unknown in Sypharoptera, so we prefer to consider Alexrasnitsynia in a separate new family.
urn:lsid:zoobank.org:act:6977AE80-1AA2-49C0-92A9-2437B8A41AC7
http://species-id.net/wiki/Permelmoa_magnifica
Figs 4A–BHolotype Ld LAP 365ab (Lapeyrie collection, imprint and counterimprint of a complete wing), stored at the Musée of Lodève, France.
Middle Permian, Lodève Basin, Salagou Formation, Les Vignasses, Lodève, Hérault, France (
Complete wing without trace of coloration; wing 11.7 mm long and 3.3 mm wide; ScP simple and parallel to RA ending on costal margin close to wing apex (about 0.5 mm); stems R+M basally distinctly curved, with R and M separating about 1.8 mm from wing base; convex simple RA nearly straight, and ending in wing apex; RP separating from RA about 1/3 of wing length, RP with three long branches ending on posterior wing margin; simple convex MA shortly connected to RP by a crossvein; concave MP deeply forked 0.9 mm from division R and M; anterior branch of MP secondary bifurcated, posterior branch simple; simple convex CuA, basally close to R+M; no apparent ‘trichotomy’ R/M/CuA; simple concave CuP; anal area with three or four veins.

Permelmoa magnifica gen. et sp. n. (Diaphanopterodea: Parelmoidae), holotype Ld LAP 365ab: A photograph of wing imprint, B photograph of wing counter-imprint, C drawing of wing (scale bar represents 3 mm).
Permelmoa is clearly attributable to the Parelmoidae because of combination of the following characters: ScP terminating on C; RP ending with three long branches; MA not coalescent with RP, but shortly connected by crossvein; MP deeply forked. Permelmoa differs from Parelmoa and Pseudelmoa in the absence of trifurcation between R, M, and CuA near wing base, and in the presence of a longer ScP terminating to C close to wing apex. In Permuralia, ScP is rather short and terminating on RA, CuA is touching M, and MA and RP are fused for a short distance, unlike in Permelmoa.
Lodevohymen lapeyriei sp. n.
Named after Lodève town and Hymen.
Wing characters only. Presence of basal fusion of M with CuA; broad areas between C and ScP and between ScP and RA without apical ‘pterostigma’-like structure, RP with five posterior parallel branches; strong convexity of basally connected veins R, M, and CuA, forming a curve.
urn:lsid:zoobank.org:act:6FCC725B-B6A5-47D3-854F-04490448BDA7
http://species-id.net/wiki/Lodevohymen_lapeyriei
Figs 5A–BHolotype LdLAP 556 (imprint of nearly complete wing, basal part not well preserved, Lapeyrie collection), stored at the Musée of Lodève, France.
Middle Permian, Lodève Basin, Salagou Formation, Les Canals, Lodève Basin, Hérault, France (
Wing elongated and basally narrow with no trace of coloration preserved, wing fragment about 16.5 mm long, estimated length about 19.5 mm width 4.2 mm in widest part; anterior margin nearly straight, ScP straight ending to C close to wing apex; areas between costal margin and ScP and ScP and RA rather broad; stem of R basally connected to M diverging 1/3 of wing length; division of RA and RP about 8.5 mm from the wing base, RA simple and straight ending in wing apex; RP pectinate with five branches ending on posterior wing margin, crossveins organized in rows between branches of RP; simple MA strongly diverges from MP 1.2 mm from division of stems R and M, and well connected with RP for distance; MP, CuA separates from CuP near wing base, CuA and M basally fused and closely parallel to R, separating 3.0 mm from base of CuA; CuA and CuP simple and straight with a few crossveins between them; anal area reduced, one anal vein present close the wing base.

Lodevohymen lapeyriei gen. et sp. n.(?Diaphanopterodea), holotype LdLAP 556: A photograph of wing, B drawing of wing (Scale bar represents 3 mm).
Lodevohymen shares with the diaphanopterodean taxa Asthenohymen Tillyard, 1924 (including Karoohymen Riek, 1976) (Asthenohymenidae Tillyard, 1924) and Biarmohymen Zallesky, 1937 (Biarmohymenidae Zallesky, 1937), but also with the megasecopteran taxa Permohymen Tillyard, 1924, Ivahymen Martynov, 1932, Protohymen Tillyard, 1924 (Protohymenidae Tillyard, 1924), Scytohymen Martynov, 1937, Oceanoptera Shcherbakov, 2009 (in
Nevertheless Lodevohymen
strongly differs from all these taxa in the broad areas between C and
ScP and between ScP and RA without apical ‘pterostigma’-like structure.
Other Diaphanopterodea and Megasecoptera have not a long fusion CuA-M in basal part of wing (
The genus Sunohymen Hong, 1985, currently included into the Protohymenidae, shares with Lodevohymen
the rather broad areas between C and ScP and RA, but they differ in RP
with only two branches in the former. Also all its structures of basal
half of wing are unknown, so that its attribution to the Protohymenidae, family in which M is basally fused to CuA, remains undemonstrated (
Lodevohymen is obviously strongly different from all other Diaphanopterodea and Megasecoptera.
Lodevohymen could be better attributed to the Diaphanopterodea rather than to the Megasecoptera for the strong convexity of the basally connected veins R, M, and CuA, forming a curve. Nevertheless, as already noted by
We described in the present study two new representatives of Diaphanopterodea, including a new family, plus one enigmatic taxon that could be either related to this order or to the Megasecoptera, and the first representative of the order Ephemeroptera
in the Salagou Formation. These new fossils confirm that this fauna is
very rich, diverse and rather unique. It also confirms the general
impression that the current knowledge on the Permian insects remains
very incomplete. Interestingly, the Coleoptera
and other holometabolous orders remain nearly unrecorded in the Lodève
basin while these insects are already present in the Lower Permian and
dominate all the fossil entomofaunas after the Triassic of Vosges (
Authors are grateful to reviewers Nina Sinitshenkova (Palaeontological Institute RAS, Moscow), Nikita J. Kluge (Saint-Petersburg State University, St.-Petersburg) and journal editor Dmitry Shcherbakov (Palaeontological Institute RAS, Moscow) for their insightful comments and suggestions. First author thank to Dr. Susan Butts for access to the collection of Peabody Museum of Natural History Yale University (New Haven, USA) and possibility to take photograph of type specimen. We sincerely thank Dr Jean Lapeyrie and Stéphane Foucher from the Musée de Lodève for having made available for study the impressive collection of fossil insects from Lodève. The first author (JP) acknowledges the research support from the Grant Agency of the Czech Republic No. P210/10/0633 and Ministry of Education of the Czech Republic MSM 0021620828.