





(C) 2011 Agnieszka Homan. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
The family Rasnitsynaphididae fam. n. has a unique combination of characters: 9-segmented antennae; rhinaria arranged in many transverse rows, surrounding the antennal segments; segment IX narrower than other segments of flagellum, always without rhinaria; cubitus branches separated; ovipositor present; siphuncular pores absent. The new family comprises the genus Rasnitsynaphis gen. n. with three species, Rasnitsynaphis ennearticulata sp. n., Rasnitsynaphis coniuncta sp. n., and Rasnitsynaphis quadrata sp. n., all from the Lower Cretaceous of Transbaikalia.
Hemiptera, aphids, fossil insects, Lower Cretaceous, Baissa, new family, new genus, new species
Baissa is one of the most important Lower Cretaceous insect fossil sites. The outcrop is located in the Asian part of Russia, in western Transbaikalia, on the left bank of the Vitim River. The exact age of lacustrine sediments of the Zaza Formation at Baissa is disputable, usually estimated as Early Cretaceous (Neocomian–Aptian), however most paleoentomologists date them as Valanginian–Hauterivian (
More than 20 000 fossil insects (including above 2 500 aphid specimens), often of excellent preservation state, have been collected there. Nearly all aphids from Baissa belong to three families: Ellinaphididae Kania & Wegierek, 2008, Szelegiewicziidae Wegierek, 1989 (both in Palaeoaphidoidea), and Oviparosiphidae Shaposhnikov, 1979 (Aphidoidea). The new taxa described below show an even higher morphological diversity of aphids in the Lower Cretaceous.
Material and methodsThe material comes from the collection of the Laboratory of Arthropods, Paleontological Institute (PIN), Russian Academy of Sciences, Moscow, where it is currently stored. The research methods did not differ substantially from those widely used in paleoentomological research (
Sternorrhyncha Amyot & Serville, 1843
Aphidomorpha Becker-Migdisova & Aizenberg, 1962
urn:lsid:zoobank.org:act:94C0708C-7F79-426A-B9FB-1DD3D53BAF67
Rasnitsynaphis gen. n.
Antennae 9-segmented, shorter than hind tibia. Rhinaria arranged in many transverse rows, surrounding antennal segments. Segment IX narrower than other segments of flagellum, blunt at apex, always without rhinaria. Cubitus branches separated. Vein Rs separates from pterostigma in one third of its length. Media originates below the base of pterostigma. Ovipositor present. Siphuncular pores absent.
urn:lsid:zoobank.org:act:2466FD65-63B1-4F19-9BC1-576C84461B21
Type species. Rasnitsynaphis ennearticulata sp. n.
Named to honour Alexandr P. Rasnitsyn, who is not only eminent researcher, but also a teacher of many paleoentomologists. P.W. is honoured to be one of his students.
As for family.
Body massive. Front edge of head convex. Head with epicranial suture. Rostrum shorter than body, reaching to the middle of abdomen. Antennae longer than one third of the body length but shorter than its half. Antennal segment III 2–4 times as long as wide; segments IV-VIII of the same length or nearly so. Surface of all segments of flagellum sculptured in form of transverse ribs. Rhinaria ellipsoidal, arranged in more or less dense rows which are sometimes joined. Cubital vein CuA1 2.5 times longer than CuA2. Pterostigma big, spindle-shaped, pointed, 3–3.5 times as long as wide. Vein Rs slightly bent at base. Media with two or three branches. Bifurcation of vein M into M1 and M2 or into M1+2 and M3 just beyond the base of vein Rs.
| 1 | Antennae short, about 1/2 of thorax height, segment III two times as long as wide, with ca. 9 rows of rhinaria; segments IV–VIII as long as wide; M with 2 branches | Rasnitsynaphis quadrata sp. n. |
| – | Antennae longer, segment III three to four times as long as wide; segments IV–VIII longer than wide | 2 |
| 2 | Segment III four times as long as wide, with ca. 11 rows of rhinaria | Rasnitsynaphis coniuncta sp. n. |
| – | Segment III three times as long as wide, with ca. 17 rows of rhinaria; M with 3 branches | Rasnitsynaphis ennearticulata sp. n. |
urn:lsid:zoobank.org:act:9040D3E2-3EFE-474D-889A-2408FD52A5BC
http://species-id.net/wiki/Rasnitsynaphis_ennearticulata
Fig. 1Holotype: PIN 3064/2109(2211); Baissa, Transbaikalia; Zaza Formation, bed 31.
From the Greek term ennea for “nine” and from the Latin term articulum for “segment” in reference to the 9-segmented antennae.
Antennae rather long; segment III three times as long as wide;segments IV–VIII of about the same length, rectangular, longer than wide. Media with three branches.
Length of the body 2.1 (Fig. 1c). Width of head 0.43 (Fig. 1a). Lateral sutures join in the middle of epicranium in the four fifth of the head length. On the dorsal side of head capsule there are three diagonal strips, running from the middle part of epicranium to the frontolateral edge of head. The distance between ocelli (situated on the back of head) 0.27. Length of the apical segment of rostrum 0.18 (Fig 1e). Antennae 0.93 long, ca. 0.8 of thorax width (Figs 1b, d). Length of antennal segments: I 0.06; II 0.08; III 0.23; each of IV–VII 0.09; VIII 0.08; IX 0.07. Antennal segment III with about 17 rows of rhinaria, with at most 8 rhinaria arranged in one row. Segments IV–VIII at most with 7 rows of rhinaria. Tibia of fore legs 0.75 long, middle tibiae 0.83 to 0.86. Mesothoracic sternite 0.93 wide, 0.53 long. Fore wings about 3.8 long. The distance from the base of the wing to the end of pterostigma 2.8. Distance between bases of cubital veins CuA1 & CuA2 0.14. The length of M1+2 more or less equals the length of the common stem of M. The posterior part of abdomen strongly sclerotized (Fig. 1f).
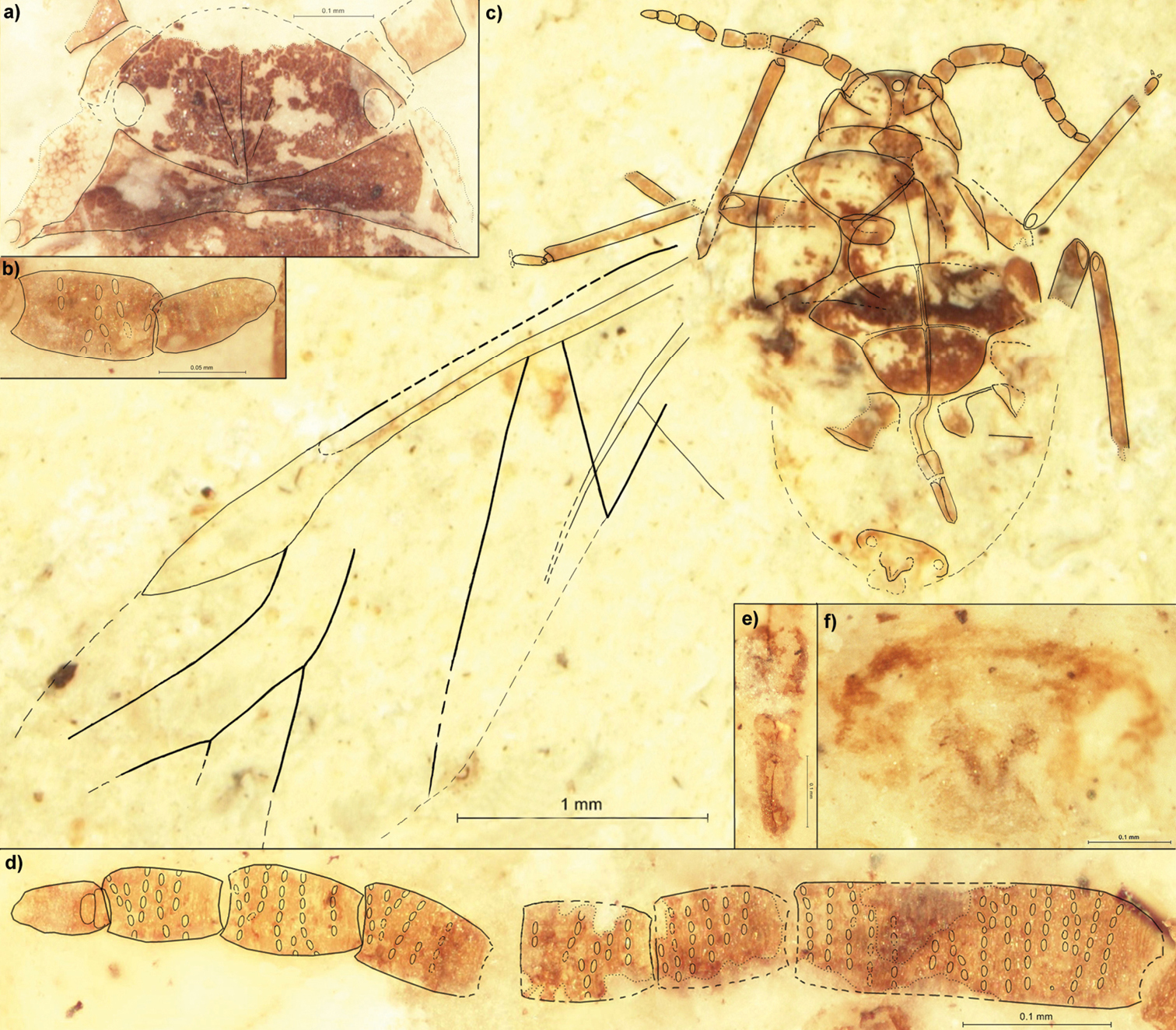
Rasnitsynaphis ennearticulata sp. n., PIN 3064/2109(2211) (holotype): a head, dorsal view b VIII and IX segment of right antenna, ventral view c body, ventral view d flagellum of left antenna, ventral view e apical segment of rostrum, ventral view f apical part of abdomen with ovipositor, ventral view.
urn:lsid:zoobank.org:act:197A9263-DFAA-4A61-9774-822FE7D3585D
http://species-id.net/wiki/Rasnitsynaphis_coniuncta
Fig. 2Holotype: PIN 3064/2209; Baissa, Transbaikalia; Zaza Formation, bed 31.
From the Latin term coniunctus for “joined” in reference to the joined rows of rhinaria.
Antenna rather long; segment III four times as long as wide; segments IV–VIII of about the same length, rectangular, longer than wide.
Length of the body about 2.4 (Fig. 2b). Head with epicranial suture. Antennae 0.88 long, about 2/3 of thorax height (Figs 2a, c). Length of antennal segments: II 0.12; III 0.24; IV 0.09; V 0.08; each of VI–VII 0.09; VIII 0.08; IX 0.07 to 0.08. Antennal segment III with 11 rows of rhinaria, with at most 7 rhinaria arranged in one row. Segments IV–VIII at most with 6 rows of rhinaria. Femur of fore legs 0.74 long, tibia 1.06. Middle tibia 1.14 long. Hind femur 0.87 long, tibia 1.34. The second segment of hind leg tarsus 0.24 (Fig. 2d). The distance between bases of cubital veins CuA1 & CuA2 0.13.
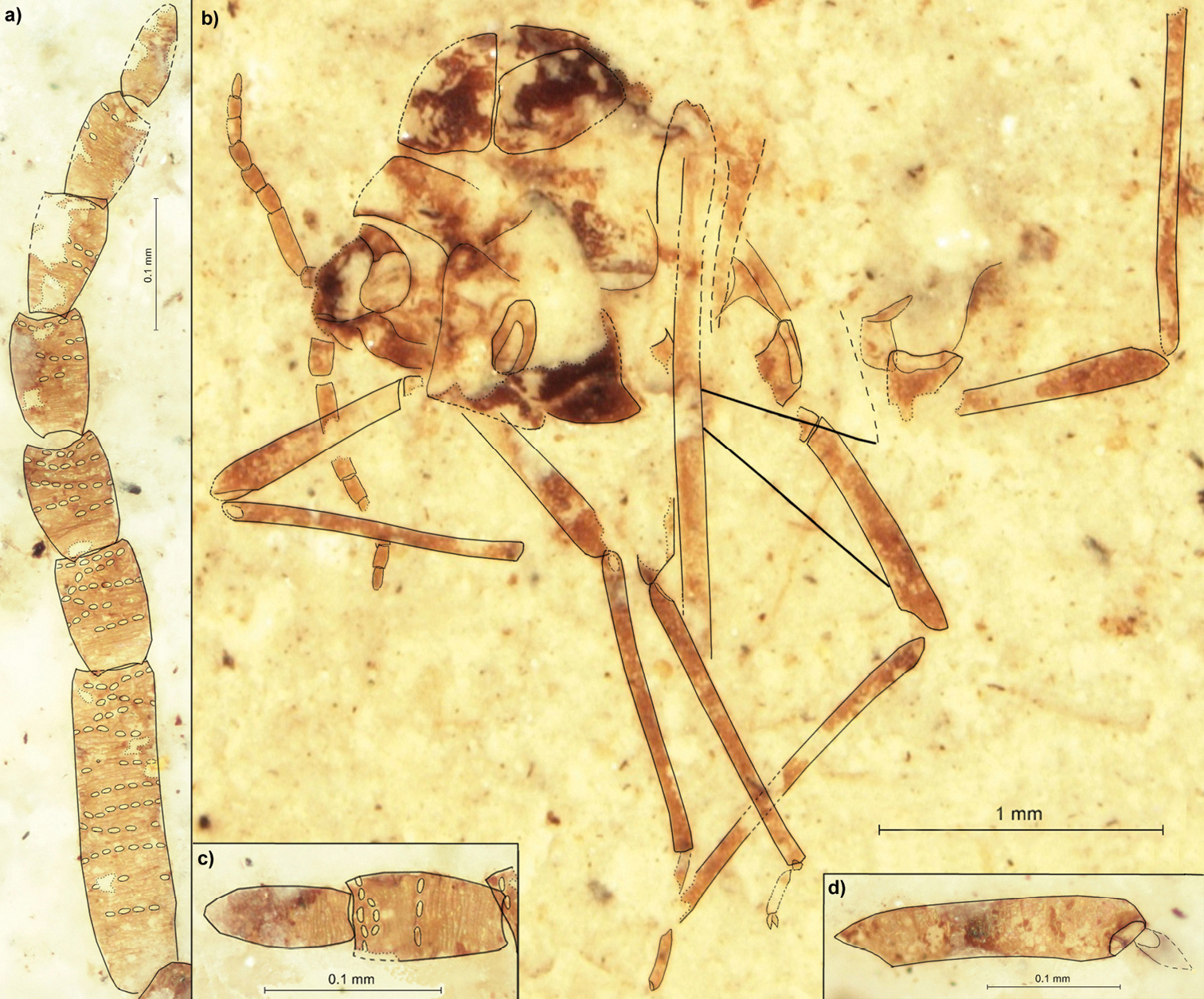
Rasnitsynaphis coniuncta sp. n., PIN 3064/2209 (holotype): a flagellum of right antenna b body, lateral view c VIII and IX segment of left antenna d second segment of hind tarsus.
urn:lsid:zoobank.org:act:7434FB55-DE6B-4C54-9086-F2E62111405D
http://species-id.net/wiki/Rasnitsynaphis_quadrata
Fig. 3Holotype: 3064/2279; Baissa, Transbaikalia; Zaza Formation, bed 31.
From the Latin term quadratus for “square” in reference to the square shape of antennal segments IV–VII.
Antennae quite short; segment III two times as long as wide; segments IV–VII of the same length, square, as long as wide. Media with two branches.
Length of the body 2.0 (Fig. 3b). Length of head 0.28. Head with lateral sutures. Length of the apical segment of rostrum 0.14 (Fig. 3c). Antennae 0.71 long, about 1/2 of thorax height (Fig. 3a, b). Length of antennal segments: I 0.06; II 0.07; III 0.16; each of IV–IX 0.06. Antennal segment III with about 9 rows of rhinaria, with at most 8 rhinaria arranged in one row. Segments IV–VIII at most with 4 rows of rhinaria. Femur of middle legs 0.54 long, tibia 0.73. Hind femur 0.57 long. Mesothoracic sternite 0.49 long. The distance from the base of the wing to the end of pterostigma 2.7. The distance between bases of cubital veins CuA1 & CuA2 0.11. The posterior part of abdomen weakly sclerotized.

Rasnitsynaphis quadrata sp. n., PIN 3064/2279 (holotype): a flagellum of right antenna, dorsal view b body, ventral view c apical segment of rostrum.
The family Rasnitsynaphididae fam. n. on the one hand possesses primitive features of Jurassic Sinojuraphididae Huang & Nel, 2008 and Genaphididae Handlirsch, 1907, but on the other hand the features typical for Early Cretaceous Oviparosiphidae and Ellinaphididae, and also certain features of recent Aphididae and Drepanosiphidae (
The body morphology and wing venation of the new family are typical for Lower Cretaceous aphids. The 9-segmented antennae recorded in Rasnitsynaphididae is the plesiomorphic feature and proves the presence of forms intermediate between Middle Jurassic Sinojuraphididae with 12-segmented antennae (
In the shape of pterostigma and place of Rs separation Rasnitsynaphis is similar to the species of Oviparosiphum Shaposhnikov & Wegierek, 1989. In the structure of CuA and M veins Rasnitsynaphis resembles Bajsaphis Shaposhnikov, 1985, Acanthotrichaphis Shaposhnikov & Wegierek, 1989, Vitimaphis Shaposhnikov & Wegierek, 1989, and other genera of the family Oviparosiphidae (
Having so many features in common with the family Oviparosiphidae, the new family differs from it by the 9-segmented antennae and lack of siphunculi.
We would like to thank Dr Ole Heie, Holte, Dr Roger Blackman, London, and Dr Dmitry Shcherbakov, Moscow for constructive comments on the manuscript.