





(C) 2011 Yuanyuan Peng. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
A new genus and species, Undulopsychopsis alexi gen. et sp. n., is described from the Early CretaceousYixian Formation of western Liaoning Province, China. This genus is probably most closely related to the Asian Cretaceous genus Kagapsychops Fujiyama, 1978. The family affinity of Undulopsychopsis gen. n. is discussed. The genus is preliminarily assigned to Psychopsidae, although it shares some character states with Osmylopsychopidae (e.g., crossveins are very scarce; Rs1 and 1A are multi-branched).
Psychopsidae, Osmylopsychopidae, fossil, Mesozoic, Huangbanjigou, China
The extant Psychopsidae
is a small family (five genera and 27 described species), currently
restricted to disjunct regions in southern Africa, southeastern Asia
and Australia (
Fossil psychopsids were much more widely distributed than
the extant taxa. Since the early 20th century, fossil psychopsids have
increasingly been found from all over the world, with species ranging
in age from the Triassic to the Tertiary. The earliest fossil record of
the Psychopsidae is Triassopsychops Tillyard, 1922 from the Late Triassic of Australia (
Fossil species currently ascribed to the family Psychopsidae.
| Species | Age | Locality | |
|---|---|---|---|
| 1 | Triassopsychops superbus Tillyard, 1922 | Late Triassic (Carnian) | Denmark Hill, Queensland, Australia |
| 2 |
Archepsychops triassicus |
Late Triassic (Carnian) | Denmark Hill, Queensland, Australia |
| 3 | Apeirophlebia grandis Handlirsch, 1906 | Early Jurassic (Early Toarcian) | Dobbertin, Germany |
| 4 | Cretapsychops decipiens Penget al., 2010 | Middle Jurassic (Aalenian/Bajocian) | Daohugou, Inner Mongolia, China |
| 5 | Beipiaopsychops triangulatus Hong, 1983 | Middle Jurassic (Aalenian/Bajocian) | Haifanggou, Liaoning, China |
| 6 | Sinopsychops chengdeensis Hong, 1982 | Middle Jurassic (Aalenian/Bajocian) | Chengde, Hebei, China |
| 7 | Calopsychops extinctus Panfilov, 1980 | Late Jurassic (Oxfordian/Kimmeridgian) | Karatau, Kazakhstan |
| 8 | Propsychops karatavicus Panfilov, 1980 | Late Jurassic (Oxfordian/Kimmeridgian) | Karatau, Kazakhstan |
| 9 | Angaropsychops sinicus Hong in Wang, 1980 | ?Early Cretaceous | Heishangou, Liaoning, China |
| 10 | Kagapsychops aranea Fujiyama, 1978 | Early Cretaceous (Valanginian/Barremian) | Kuwajima, Japan |
| 11 | Angaropsychops turgensis Martynova, 1949 | Early Cretaceous (Hauterivian) | Turga, Transbaikalia, Russia |
| 12 | Psychopsites rolandi Jepson et al., 2009 | Early Cretaceous (Hauterivian) | Lower Weald Clay, Wealden, England |
| 13 | Valdipsychops minimus Jepson et al., 2009 | Early Cretaceous (Hauterivian) | Lower Weald Clay, Wealden, England |
| 14 | Baisopsychops lambkini Makarkin, 1997 | Early Cretaceous (pre-Barremian) | Baissa, Transbaikalia, Russia |
| 15 | Epipsychopsis fusca Makarkin, 2010 | Early Cretaceous (pre-Barremian) | Baissa, Transbaikalia, Russia |
| 16 | Epipsychopsis variegata Makarkin, 2010 | Early Cretaceous (pre-Barremian) | Baissa, Transbaikalia, Russia |
| 17 | Undulopsychopsis alexi gen. et sp. n. | Early Cretaceous (Barremian) | Huangbanjigou, Liaoning, China |
| 18 | Cretapsychops corami Jepson et al., 2009 | Early Cretaceous (Barremian) | Upper Weald Clay, Wealden, England |
| 19 | Micropsychops parallelus Jepson et al., 2009 | Early Cretaceous (Barremian) | Upper Weald Clay, Wealden, England |
| 20 | Valdipsychops brigidae Jepson et al., 2009 | Early Cretaceous (Barremian) | Upper Weald Clay, Wealden, England |
| 21 | Valdipsychops logunovi Jepson et al., 2009 | Early Cretaceous (Barremian) | Upper Weald Clay, Wealden, England |
| 22 | Valdipsychops proudlovei Jepson et al., 2009 | Early Cretaceous (Barremian) | Upper Weald Clay, Wealden, England |
| 23 | Valdipsychops maculosus Jepson et al., 2009 | Early Cretaceous (Barremian) | Upper Weald Clay, Wealden, England |
| 24 | Pulchroptilonia espatifata Martins-Neto, 1997 | Early Cretaceous (Aptian) | Araripe Basin, Brazil |
| 25 | Putzneura parcimoniosa Martins-Neto & Rodrigues, 2010 | Early Cretaceous (Aptian) | Araripe Basin, Brazil |
| 26 | Litopsychopsis burmitica Engel & Grimaldi, 2008 | Early Cretaceous (Albian) | Burmese amber |
| 27 | Embaneura vachrameevi G. Zalessky, 1953 | Late Cretaceous (Cenomanian) | Emba, Kazakhstan |
| 28 | Grammapsychops lebedevi Martynova, 1954 | Late Cretaceous (Cenomanian) | Kem’ River, Siberia, Russia |
| 29 | Kagapsychops continentalis Makarkin, 1994 | Late Cretaceous (Turonian) | Kzyl-Zhar, Kazakhstan |
| 30 | Arctopsychops zherikhini Makarkin, 1994 | Late Cretaceous (Turonian) | Arkagala, NE Siberia, Russia |
| 31 | Propsychopsis helmi Krüger, 1923 | Eocene (Lutetian/Bartonian) | Baltic amber |
| 32 | Propsychopsis hageni MacLeod, 1971 | Eocene (Lutetian/Bartonian) | Baltic amber |
| 33 | Propsychopsis lapicidae MacLeod, 1971 | Eocene (Lutetian/Bartonian) | Baltic amber |
| 34 | Miopsychopsis relicta Makarkin, 1991 | Late Eocene/Early Oligocene | Amgu, Sikhote-Alin, Russia |
| 35 | Miopsychopsis sikhotensis Makarkin, 1991 | Late Eocene/Early Oligocene | Amgu, Sikhote-Alin, Russia |
The specimen described herein is from the Yixian
Formation of Huangbanjigou Village, Shangyuan County, Beipiao City,
western Liaoning Province, northeastern China. The principal
fossil-bearing layers in Huangbanjigou locality are silty mudstone,
yellowish to grayish, rich in insects, fish and plants (
The material was examined using a Leica MZ12.5 dissecting microscope. The photographs were taken using a Nikon D100 digital camera coupled to a Nikkor 105mm macro lens, and final photographs were adjusted by using Adobe Photoshop 4.0 image-editing software. All line drawings were drawn made directly using CorelDraw 12 graphic software.
We follow the traditional (sensu
Family Psychopsidae Handlirsch, 1906
Undulopsychopsis alexi sp. n.
Forewings: costal gradate series absent; branches of Rs dichotomously branched; Rs1 multi-branched, pectinate with branches directed anteriorly; M forked far distal to origin of Rs; CuP dichotomously branched. Hind and outer margins of both wings undulate.
The generic name is derived from the Late Latinundula (meaning a small wave, refers to its undulate wing margins) and Psychopsis (the type genus of the family). The gender is feminine.
This new genus differs from all other psychopsids by possessing undulate outer and hind margins of both wings. The combination of the following forewing character states is also characteristic: no costal gradate series; branches of Rs dichotomously branched; the basal-most branch of Rs multi-branched, and M forked far distal to the origin of Rs. The new genus has scarce costal crossveins, which are not arranged in gradate series, in contrast to the genera Grammapsychops, Miopsychopsis Makarkin, 1991, Baisopsychops Makarkin, 1997, Cretapsychops Jepson et al., 2009 and Epipsychopsis Makarkin, 2010. Undulopsychopsis gen. n.possesses the dichotomously branched branches of Rs; this condition is also present in the following psychopsid genera: Triassopsychops, Angaropsychops Martynova, 1949, Psychopsites Jepson et al., 2009, Valdipsychops Jepson et al., 2009, Epipsychopsis, Pulchroptilonia, Putzneura Martins-Neto & Rodrigues, 2010, Kagapsychops, Grammapsychops, and Embaneura. Among these the new genus is most similar to those genera which have the multi-branched Rs1 and M forked far distal to the origin of Rs. This combination is present only in the genus Kagapsychops. The type species of this genus (Kagapsychops aranea Fujiyama, 1978) is fragmentary and poorly preserved, but Kagapsychops continentalis Makarkin, 1994 is well-preserved (although incomplete). Undulopsychopsis gen. n.clearlydiffers from Kagapsychops by being a much smaller size (the forewing of the former is approximately twice shorter than that of the latter), and the absence of the gradate series of crossveins in the radial space. Other fossil psychopsids, for example Propsychopsis Krüger, 1923, Litopsychopsis Engel & Grimaldi, 2008 and Micropsychops Jepson et al., 2009 differ strongly from the new genus by having mostly unbranched veins of Rs before end-twigging and several long gradate series of crossveins in the radial space.
urn:lsid:zoobank.org:act:29E097D3-A80A-48B2-88D7-8240349F17D0
http://species-id.net/wiki/Undulopsychopsis_alexi
Figs 1–3Holotype CYNB044, a well-preserved specimen, with body partially preserved and four wings overlapping pairwise.
As for the genus.
Body: only partial thorax preserved. Pronotum sub-rectangular, 1.2 mm long, 2.8 mm wide, suffused with many long hairs. Mesonotum 3 mm long, 3.5 mm wide, with some long hairs laterally.
Forewing (Fig. 3) subtriangular, 21.5 mm long, 12.3 mm wide. Costal space broad throughout; subcostal veinlets forked; humeral veinlet slightly recurrent, branched. Subcostal space much broader than R1 space. R1 space narrow. Sc and R1 close distally but not fused. Rs with 10 primary branches, branches of Rs dichotomously branched; Rs1 pectinately branched with branches directed anteriorly. M appears originating from R, forked far from origins of Rs1. MA and MP probably simple (their terminal parts not preserved). Cu forked near wing base. CuA pectinately branched distal to fork of M. CuP multi-branched, dichotomous. Anal area well-developed. 1A long, dichotomously branched. 2A multi-branched (incompletely preserved). Only few crossveins detected: costal space basally with scarce crossveins, not forming gradate series; subcostal space with 4 crossveins preserved; R1 space with 5 crossveins preserved; medial space with 2 crossveins preserved. Veins covered with dense hairs, particularly long basally. Trichosors distinct. Wing membrane in general brownish; colour pattern consists mainly of two pale transverse zigzagged bands which are proximally darker than other portions of wing; indistinctly mottled basally and apically. Wing margin haired; hind and outer margins undulate.
Hind wings almost entirely hidden under forewing, about 16.5 mm long as preserved, 10 mm wide. Venation very poorly preserved; no details visible. Outer margin undulate.

Undulopsychopsis alexi gen. et sp. n. The holotype CYNB044. Photograph. Scale bar = 5 mm.
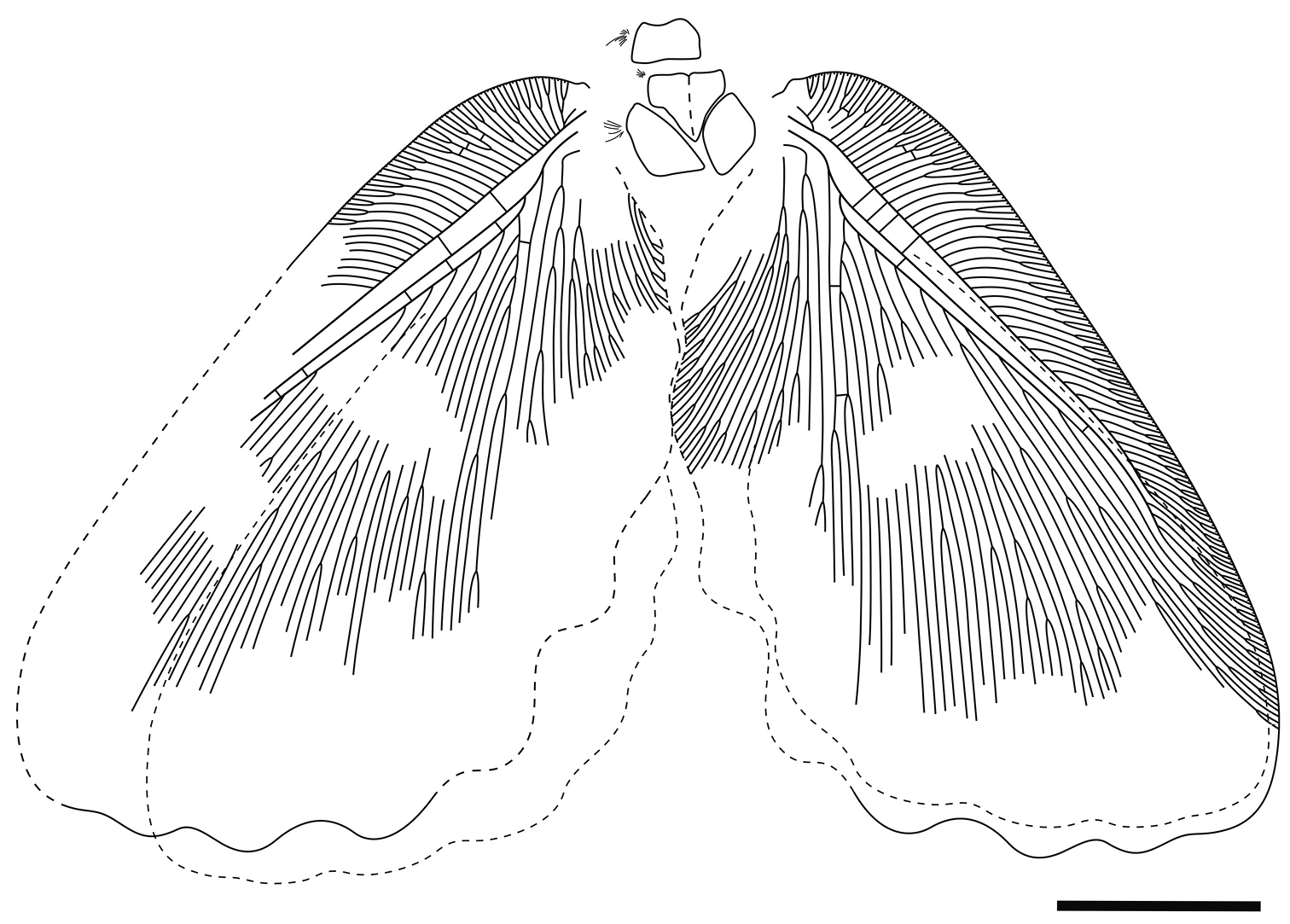
Undulopsychopsis alexi gen. et sp. n. The holotype CYNB044. Drawing. Scale bar = 5 mm.
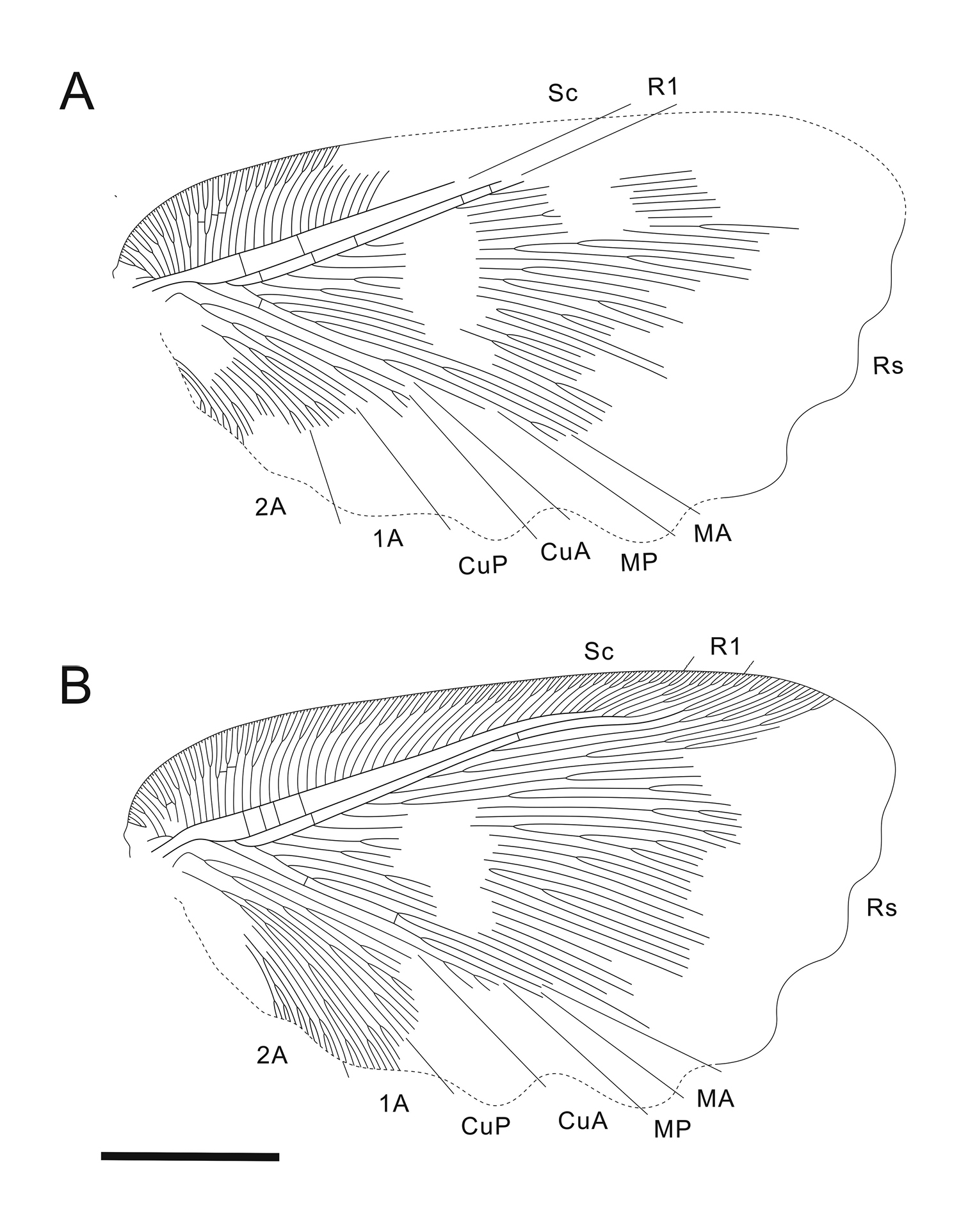
Undulopsychopsis alexi gen. et sp. n. The forewing venation of the holotype CYNB044. A left forewing (converted to the right) B right forewing. Scale bar = 5 mm.
The specific name is named in honor of the distinguished Russian paleoentomologist Prof. Alexandr (‘Alex’) Rasnitsyn.
Yixian Formation, Huangbanjigou Village, Shangyuan County, Beipiao City, Liaoning Province, China.
Based on the configuration of the venation in the radial space of the forewing, fossil psychopsids can be divided into two groups. One group includes the taxa with simple branches of Rs, the majority of which are not branched before end-twigging. This group is represented by the genera Propsychopsis, Baisopsychops, Cretapsychops, Micropsychops and Litopsychopsis. The crossveins in these genera are usually arranged in one to two gradate series in the costal space, and two to four long gradate series in the radial space. They occur in the Cretaceous and Eocene; all extant genera belong to this group as well.
The other group includes the taxa which have the branches
of Rs dichotomously branched, and often the basal-most branch of Rs
multi-branched. Representative genera of this group are the earliest
psychopsid Triassopsychops, and other Mesozoic psychopsids, e.g., Angaropsychops, Grammapsychops and Kagapsychops
(see complete list above). They possess numerous radial crossveins,
arranged in many short gradate series (often irregular), and usually no
costal gradate series. Undulopsychopsis gen. n. belongs to the latter group. It is preliminarily assigned to Psychopsidae,
as its Sc and R1 are not fused apically, and the costal space is
broad, although it almost lacks crossveins. The latter feature, and
the multi-branched Rs1 and 1A are shared by this genus with another
Mesozoic psychopsoid family Osmylopsychopidae (especially with its type genus) known from the Triassic of Australia and Central Asia (
Previously, only four species have been recorded from the Mesozoic of China, i.e., Angaropsychops sinicus Hong in Wang, 1980 (probably from the Early Cretaceous Yixian Formation), Sinopsychops chengdeensis Hong, 1982, Beipiaopsychops triangulatus Hong, 1983, and Cretapsychops decipiens Peng et al., 2010 (all from the Middle Jurassic Jiulongshan Formation). Undulopsychopsis gen. n. is the fifth representative of the Chinese psychopsids found from the different locality (Huangbanjigou). It is characterized by the undulate wing margin, a unique character state among known Psychopsidae, and the forewing venation that is not typical for this family compared with most other genera of Psychopsidae.
We thank Dr Wang YongJie (College of Life Sciences, Capital Normal University) for his valuable suggestions throughout the work. We are grateful to Dr Shih ChungKun (College of Life Sciences, Capital Normal University) for his review and improvement of our manuscript, and Dr James Jepson (University of Manchester, UK) for correction of the English. We express our gratitude to anonymous reviewers for their critical review of the manuscript. This work is supported by the National Natural Science Foundation of China (No. 40872022, 31071964), the Nature Science Foundation of Beijing (No. 5082002), the Key and PHR Program of the Beijing Municipal Commission of Education, the Chinese Municipal Education Commission Discipline Construction and Graduate Education Construction Project.