





(C) 2011 Alexei S. Bashkuev. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
The family Mesopanorpodidae is revised. Most taxa referred to this family are not related to the type genus Mesopanorpodes Tillyard, 1918. The type species of the latter, Mesopanorpodes wianamattensis, is closely related to Mesopsyche Tillyard, 1917. Therefore Mesopanorpodes is transferred to Mesopsychidae Tillyard, 1917 (= Mesopanorpodidae Tillyard, 1918, syn. n.).The earliest Mesopsychidae are described from the Upper Permian of European Russia (Severodvinian; Isady locality, Vologda Province): Permopsyche issadensis gen. et sp. n. (type species) and Permopsyche rasnitsyni sp. n. Two species described under Mesopanorpodes from the Upper Permian of Australia are also included into Permopsyche: Permopsyche belmontensis (Riek, 1953), comb. n., Permopsyche robustus (Riek, 1953) comb. n. The first pre-Triassic Mesopsyche, Mesopsyche incompleta sp. n. is described from the uppermost Permian (the town of Vyazniki, Vladimir Province). Bittacopanorpa javorskii Zalessky, 1935 from the uppermost Permian or basal Triassic of Kuznetsk Basin is identified as a hindwing of Mesopsyche: Mesopsyche javorskii (Zalessky, 1935) comb. n. The origin, evolutionary history, and stratigraphic occurrence of Mesopsychidae are discussed.
Mesopsychidae, Mesopanorpodes, Mesopsyche, Permopsyche, Bittacopanorpa, Permian, Triassic, new taxa, revision
Confusingly, this remark passed completely unnoticed by
all the subsequent authors who discussed the composition and taxonomy of
the Mesopanorpodidae.
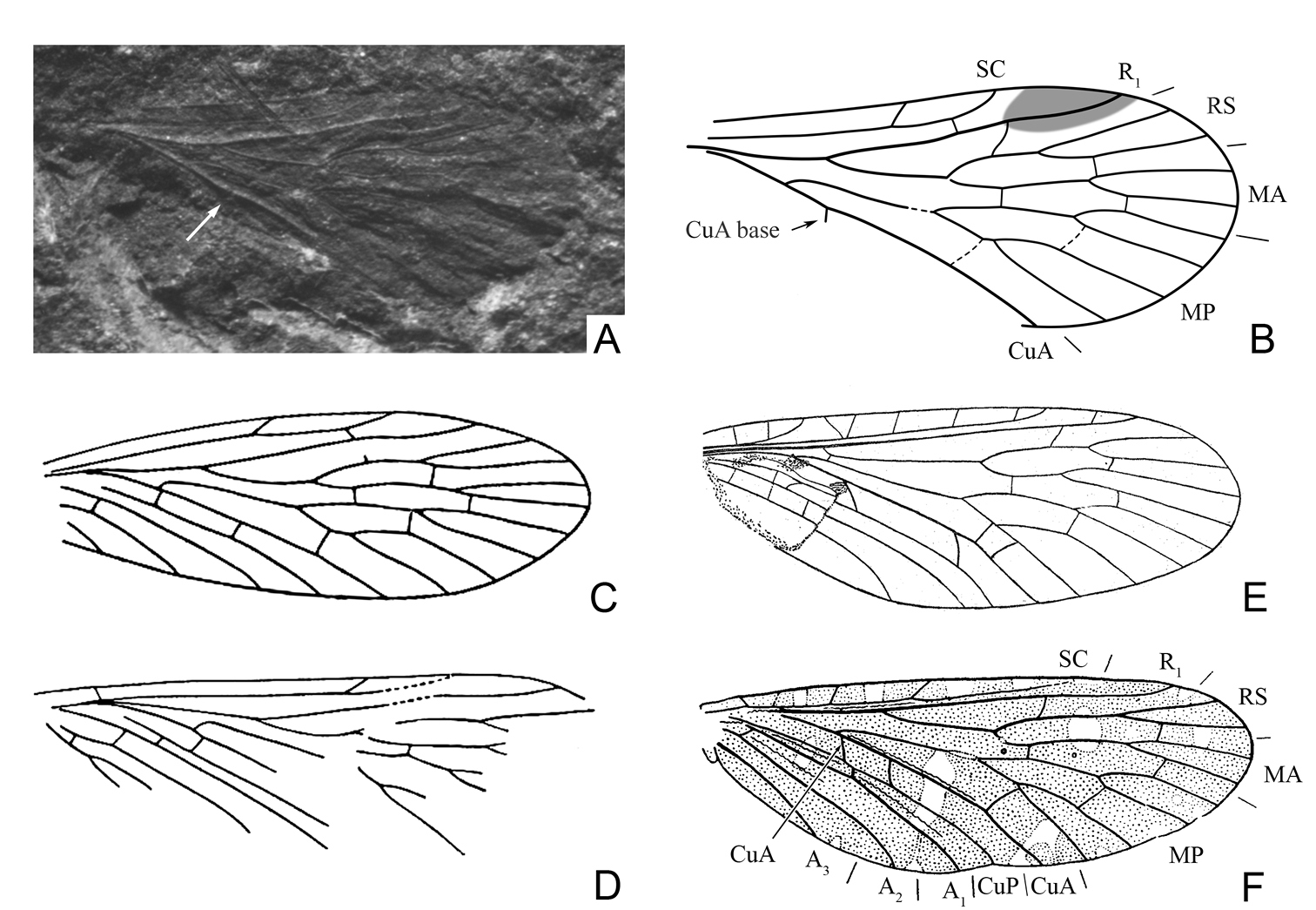
Wings of Mesopanorpodes and Mesopsyche, for comparison A photograph of Mesopanorpodes wianamattensis Tillyard (position of CuA base marked with arrow) B Mesopanorpodes wianamattensis, line drawing (orig.) C Mesopanorpodes belmontensis: from
In fact, species of Prochoristella (with the possible exception of Prochoristella belli Riek, 1953, known only from a hindwing) display characters typical of Permochoristidae, that is, first of all, a deep fork on MA vein and an oblique base of CuA, either fused with MP for a distance, or forming cubito-median Y-vein. Prochoristella and Mesopanorpodes are similar only in the number of RS+MA and MP endings being reduced to four. However, this is characteristic of many independent lineages, e.g. Permotanyderidae + Liassophilidae, Nannochoristidae, or Bittacidae, and cannot be accepted as a reliable apomorphy. I consider Prochoristella, including its possible junior synonyms (or at least closely related genera) Afristella Riek, 1974, Mesotanyderus Riek, 1955, and Austrochoristella Willmann, 1989, as belonging to Permochoristinae (Permochoristidae) and not related to Mesopanorpodes.
Equally unacceptable is the assignment to Mesopanorpodidae
of a number of species from the mid-Triassic Tongchuan Fm of Shaanxi
Province, China, described by Hong You-Chong and co-authors (
The Jurassic genera Itaphlebia Sukatcheva, 1985 and Chrysopanorpa Ren, 1995 were originally placed in Mesopanorpodidae as well.
Stylopanorpodes eurypterus, Netropanorpodes decorosus, and Netropanorpodes sentosus Sun, Ren & Shih, 2007 (
Mesopanorpodes mostovskii Novokshonov et al., 2004 from the basal Triassic of Northern Russia has been transferred to the related family Nedubroviidae (
Therefore, with the above-mentioned taxa discarded, Mesopanorpodidae remain composed of only three species of Mesopanorpodes from the Upper Permian and Middle Triassic of Australia. But are they actually congeneric?
Re-examination of available photographs of Mesopanorpodes wianamattensis reveals that its CuA is distinctly bent distal to the origin of MP, and the base of CuA is shifted distad and inclined backwards, like in Mesopsyche (Fig. 1A–B). Thus, Mesopanorpodes appears venationally indistinguishable from Mesopsyche. Here I provisionally retain Mesopanorpodes as a separate genus until the complete revision of Mesopsyche. Unlike Mesopsyche wianamattensis and Mesopsyche, both species described by Riek have the base of CuA short and transverse, with the intercubital space not expanded, which is presumed to be a plesiomorphic condition. These species are included here in the new genus Permopsyche gen. n.
In this paper, two new species of Permopsyche, Permopsyche issadensis sp. n. (type species)and Permopsyche rasnitsyni sp. n., are described from the Upper Permian of European Russia. These are the oldest currently known records of Mesopsychidae. The new material includes a number of well-preserved fore- and hindwings, which allow establishing yet another difference between Permopsyche and Mesopsyche, the position of the hindwing CuA vein with respect to the membrane. In Permopsyche issadensis CuA is concave (while the R1 is convex), as is typical of Permochoristidae, while in Mesopsyche spp., including the Permian species, as well as the other Mesozoic mesopsychid genera, both CuA and R1 are convex, like in most post-Paleozoic Mecoptera.
The earliest record of Mesopsyche, and the only one known so far to be definitely pre-Triassic, Mesopsyche incompleta sp. n., is described from the uppermost Permian of the European Russia.
Bittacopanorpa javorskii
Zalessky, 1936 from the transitional Permian–Triassic sequence of the
Kuznetsk Basin (Babiy Kamen’ locality, Maltsevo Formation), known only
from a wing base (Fig. 8G–H), originally placed in Neorthophlebiidae and tentatively transferred to Permotipulidae (
The study is based on examination of ca. 60 fossil specimens from the collection of Borissiak Paleontological Institute, Russian Academy of Sciences, Moscow (PIN). The holotype of Bittacopanorpa javorskii Zalessky is preserved in the collection of the Central Research Geological Prospecting Museum, Saint Petersburg (CRGPM).
Fifty-three isolated wings of Permopsyche
were collected from a lens of fluvio-lacustrine deposits within the
Sukhona River section, in Vologda Province, north-central Russia
(Isady locality). It corresponds to the Kalikino Member of the Poldarsa
Formation, and is dated the latest Severodvinian (correlated with the
Lower Wuchiapingian:
Five specimens of Mesopsyche
were collected from the Balymotikha locality, an outcrop of lacustrine
deposits of Vyazniki Permian–Triassic sequence, at the town of
Vyazniki, Vladimir Province, central Russia. It is dated the latest
Vyatkian (Late Changhsingian), somewhat below the Permian–Triassic
boundary (
The fossils were examined with a Leica M165C stereomicroscope and photographed using an attached Leica DFC 425 digital camera. Images were digitally processed with Helicon Focus v. 5.1 and Adobe Photoshop CS3 graphic software. Line drawings were made using Inkscape v. 0.48 vector graphics editor.
On the line drawings the wings are oriented with their apices to the right, while some photographs are reversed left-right for better comparison.
The wing venation terminology follows
http://species-id.net/wiki/Mesopsychidae
Mesopsyche Tillyard, 1917, Mesopanorpodes Tillyard, 1918, Vitimopsyche Novokshonov & Sukatsheva, 2001, Baissopsyche Novokshonov & Sukatsheva, 2001, Lychnomesopsyche Ren et al., 2009, Permopsyche Bashkuev, gen. n.
The genus Mesopanorpodes is restricted here to its type species, Mesopanorpodes wianamattensis. Though Mesopanorpodes appears venationally indistinguishable from Mesopsyche, it doesn’t seem appropriate to synonymize these genera prior to the complete revision of the latter. The genus Mesopsyche (sensu
urn:lsid:zoobank.org:act:57B809F6-634A-446C-AB88-79BE4E3E745C
http://species-id.net/wiki/Permopsyche
Permopsyche issadensis sp. n.; Upper Permian, European Russia.
From the Permian and Greek psyche, “soul” or “mind, ” the word often used for devising names of delicately winged insects. Gender feminine.
In forewing, SC long, bearing only inclined fore branch, connecting distally with R1 by crossvein. Costal space narrow. Both RS and MA forks not longer than their stems. MP 4-branched. CuA base oblique to transverse, not distinctly inclined backwards; M5 present or lost. Anal area not expanded. Crossveins not numerous. In hindwing, CuP concave with respect to membrane (in contrast to convex R1).
Differs from the closest genus Mesopsyche by the base of CuA being oblique to transverse (not inclined backwards), costal space narrower, and hindwing CuP concave with respect to membrane.
In addition to the type species, from the same locality, Permopsyche rasnitsyni sp. n.;and from the Upper Permian of Australia, Permopsyche belmontensis (Riek, 1953) comb. n. and Permopsyche robustus (Riek, 1953) comb. n.
The assumption that one of the key characters, the position of hindwing CuP with respect to the membrane, known in the type species only, is shared by the other species is tentative and requires further verification.
urn:lsid:zoobank.org:act:FBCADCA5-E2B6-4D4E-B044-6DE29496AAFD
http://species-id.net/wiki/Permopsyche_issadensis
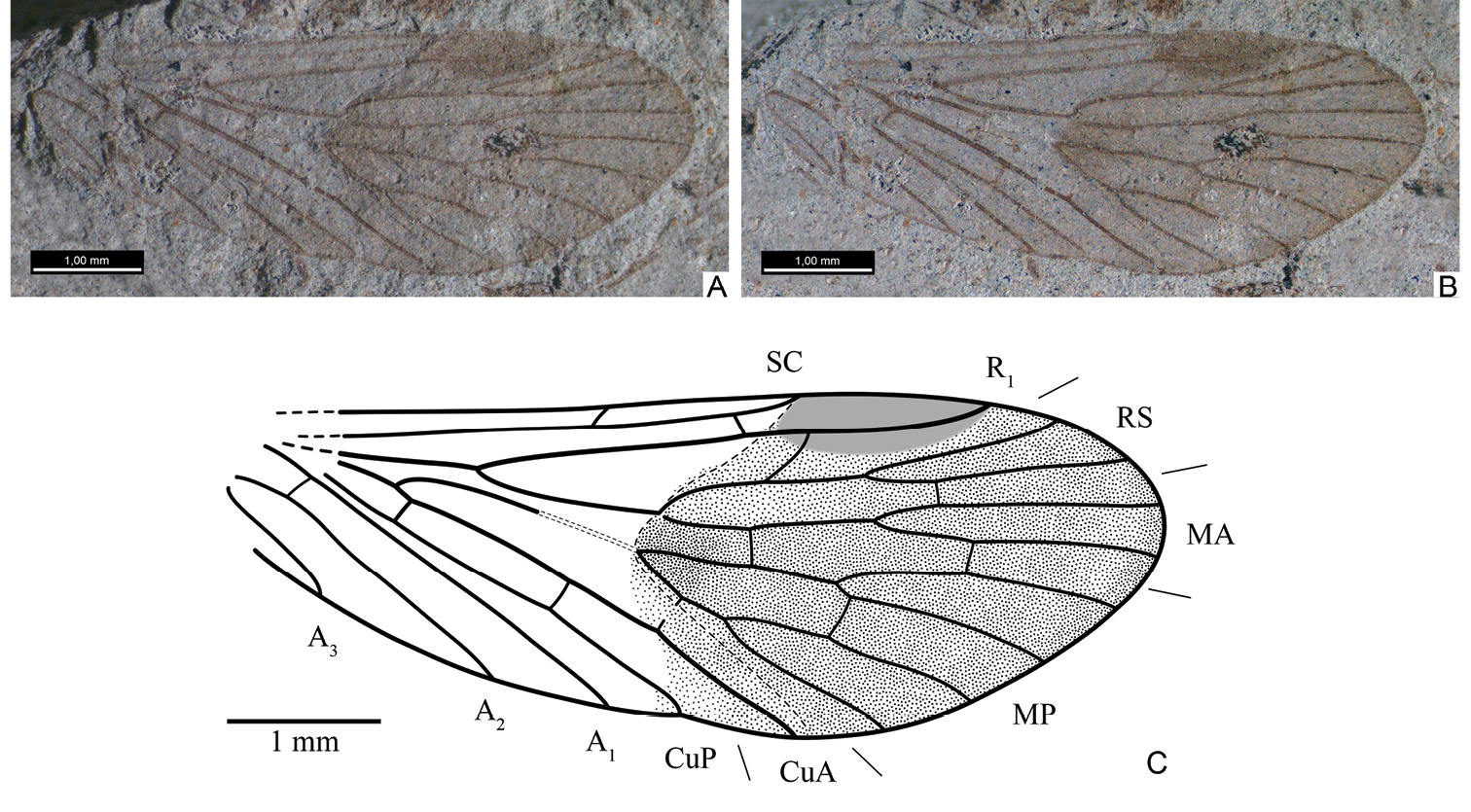
Figs 2–4, 6From the Isady locality.
PIN, no. 3840/336, well-preserved forewing (part and counterpart).
21 forewings (see Table 1) and 6 hindwings: PIN, no. 3840/725, 1381–1383, 1385, 1389a. Additionally, 8 forewings and 10 hindwings incomplete or poorly preserved, excluded from the type series.
Isady locality, Vologda Province, North European Russia; Poldarsa Formation, uppermost Severodvinian (Lower Wuchiapingian), Upper Permian.
Differs from Permopsyche belmontensis by wing shape, with anterior margin straight and posterior apical margin tending to oblique, different crossveins arrangement, and strongly unsclerotized MP stem. Differs from Permopsyche robustus by branch of SC located more basally and A3 simple. Additionally differs from both species by considerably smaller size.
Forewing. Moderately broad (length/width ratio
2.3–2.9:1); anterior margin nearly straight, slightly convex basally;
apex obtuse to rounded. SC long, reaching or almost reaching basal
margin of pterostigma; SC branch approximately at middle of RS+MA stem
(somewhat variable in paratypes). Pterostigma distinct, widened, with
oblique basal margin. R1 simple, gently bending posteriad at
pterostigma. RS and MA forks equally short, approximately as long as
their stems. Nodal line distinct, arched, running from tip of SC to
hind margin at apex of CuA. Thyridium present as unsclerotized section
on MP stem spanning from approximately 1/3–1/2 of its length down to
fork, accompanied by desclerotized spot at point of branching of RS and
MA (“thyridulum”, the term introduced by A.P. Rasnitsyn in
Permopsyche issadensis gen. et sp. n. A–B photographs of the holotype, under different illumination C line drawing of holotype.

Photographs of forewings of Permopsyche issadensis gen. et sp. n. A paratype PIN, no. 3840/717; B paratype PIN, no. 3840/1354 C paratype PIN, no. 3840/1355 D paratype PIN, no. 3840/1357. All except D reversed left-right.
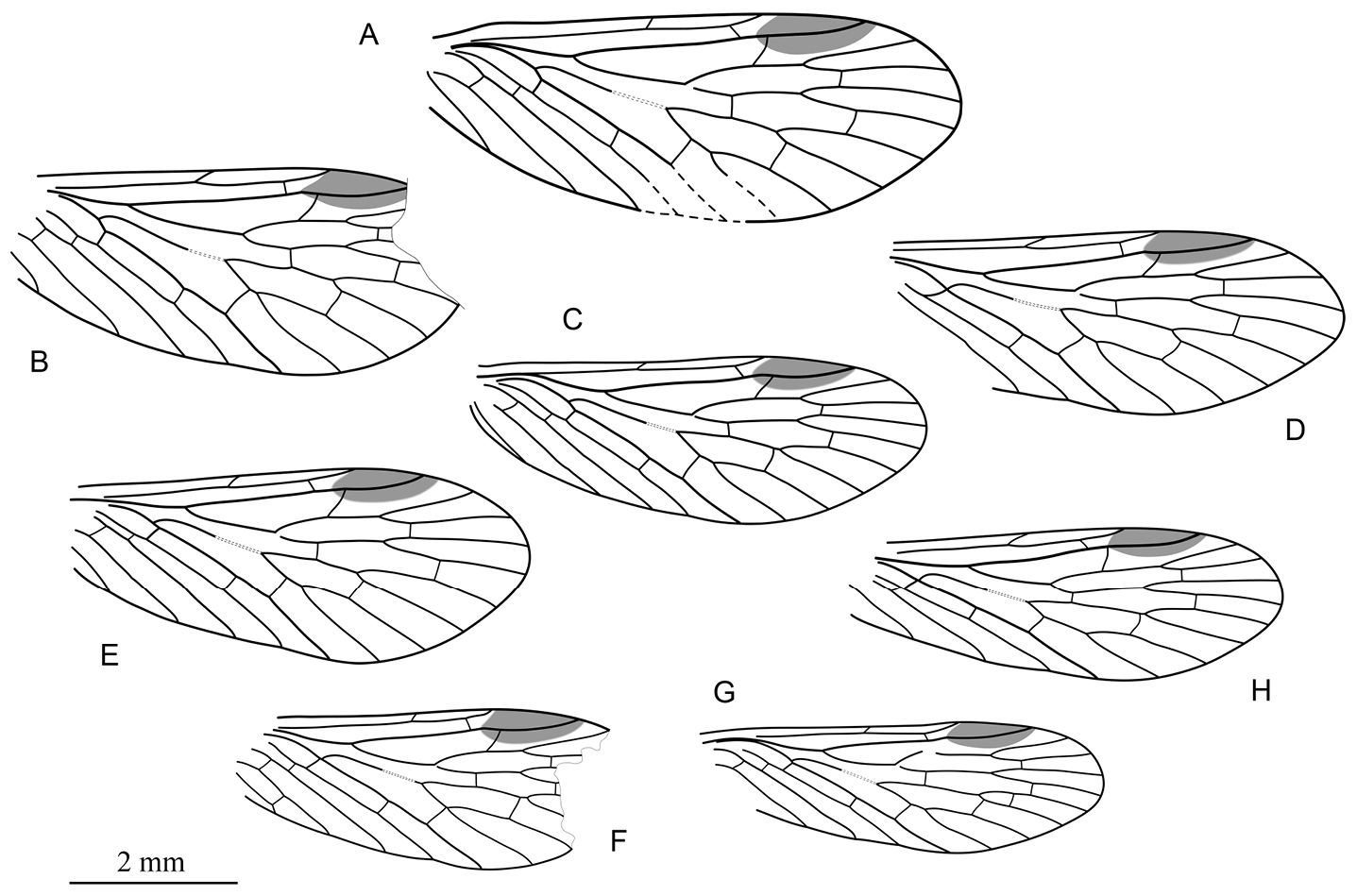
Line drawings of paratypes of Permopsyche issadensis gen. et sp. n. showing the variability of venation and wing size A PIN, no. 3840/718 B PIN, no. 3840/719 C PIN, no. 3840/105 D PIN, no. 3840/1357 E PIN, no. 3840/1352 F PIN, no. 3840/1354 G PIN, no. 3840/728 H PIN, no. 3840/729. All to the same scale.
Hindwing. Both venation scheme and coloration similar to those in forewing, adjusted for general difference between fore- and hindwings typical of Mecoptera. SC short, reaching wing midlength, variably forking apically (Fig. 6). Pterostigma distinct. R1 with inclined branch in pterostigmal area. A1 separating from CuP relatively closely to base.
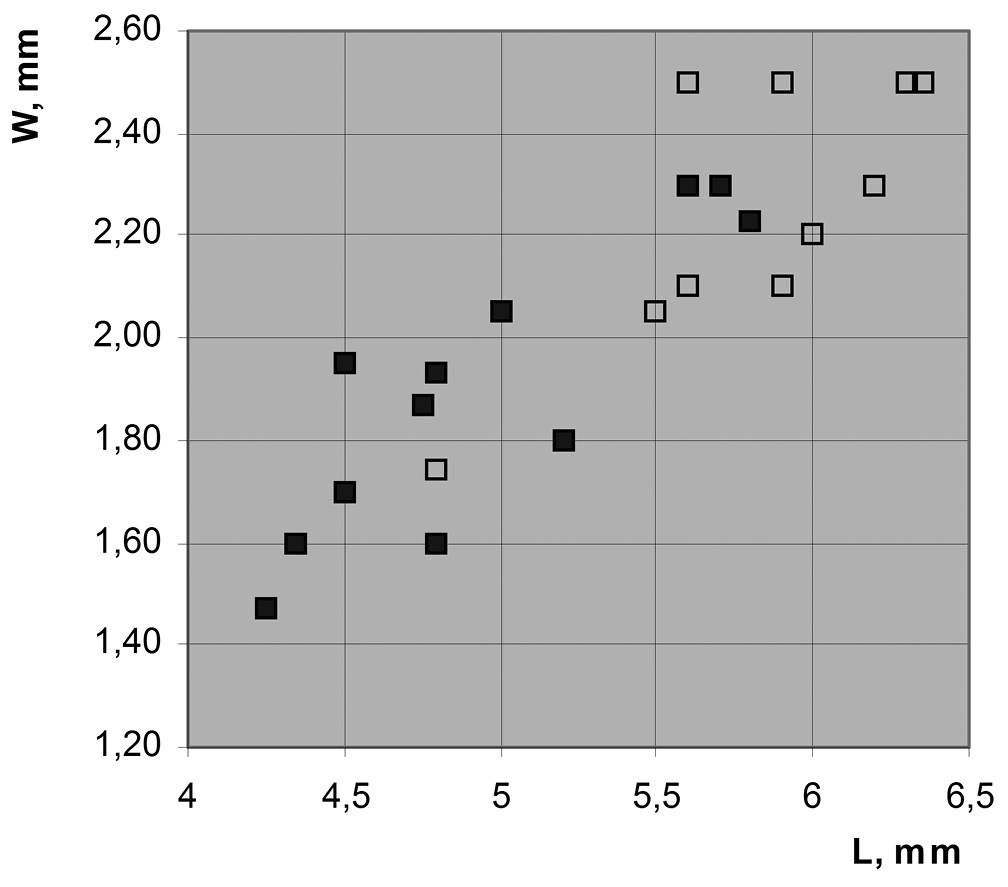
Correlation between wing size and structure of connection between CuA and MP in Permopsyche issadensis, based on Table 1; see text for explanation.

Figure 6. Hindwings ofPermopsyche issadensis gen. et sp. n. A–B paratype PIN, no. 3840/1381, photograph and line drawing C–D paratype PIN, no. 3840/1382 C details of cubito-anal structure D line drawing of hindwing A, C reversed left-right.
Forewing length, 4.25–6.35, width, ca. 1.5–2.5; hindwing length, 3.9–5.5, width, 1.5–2.25.
In Permopsyche issadensis thewing size varies conspicuously. As shown in the size distribution diagram for forewings (Fig. 5; Table 1), the presence of Y-vein and the transverse (rather than oblique) shape of CuA base (hollow squares vs filled squares) both correlate with the increased wing size (irrespectively of the length/width ratio). However, the size distributions of the two morphotypes overlap broadly. The two wing morphotypes may represent sexual dimorphism. Alternatively, they can be attributed to slight differences in flight mechanics depending on the body size. The distinctive and uniform wing color pattern makes it unlikely that the morphotypes represent two closely related species.
Forewing sizes of Permopsyche issadensis gen. et sp. n. Some measurements are acceptably approximate due to restoration of wing base and/or apex.
| PIN, no. 3840/… | Length | Width | L:W | CuA/MP fusion |
| 730 | 4.25 | 1.47 | 2.89 | ■ |
| 1362 | 4.35 | 1.6 | 2.72 | ■ |
| 1360 | 4.5 | 1.7 | 2.65 | ■ |
| 723 | 4.5 | 1.95 | 2.31 | ■ |
| 1367 | 4.75 | 1.87 | 2.54 | ■ |
| 728 | 4.8 | 1.6 | 3.00 | ■ |
| 1355 | 4.8 | 1.74 | 2.76 | □ |
| 1354 | 4.8 | 1.93 | 2.49 | ■ |
| 1359 | 5.0 | 2.05 | 2.44 | ■ |
| 729 | 5.2 | 1.8 | 2.89 | ■ |
| 105 | 5.5 | 2.05 | 2.68 | □ |
| 1349 | 5.6 | 2.1 | 2.67 | □ |
| 1352 | 5.6 | 2.3 | 2.43 | ■ |
| 719 | 5.6 | 2.5 | 2.24 | □ |
| 1358 | 5.7 | 2.3 | 2.48 | ■ |
| 1357 | 5.8 | 2.23 | 2.60 | ■ |
| 1353 | 5.9 | 2.1 | 2.81 | □ |
| 1350 | 5.9 | 2.5 | 2.36 | □ |
| 1366 | 6.0 | 2.2 | 2.73 | □ |
| 336 | 6.2 | 2.3 | 2.70 | □ |
| 717 | 6.3 | 2.5 | 2.52 | □ |
| 718 | 6.35 | 2.5 | 2.54 | □ |
The nodal line (the line of wing flexion) in the forewing of the new species is essentially similar to that of both Mesopsyche (
Hindwings of Permopsyche are almost as abundant at Isady as the forewings. All the hindwings examined, as far as their preservation allows to tell, have the set of characters (including coloration) diagnostic of the forewings of Permopsyche issadensis and can be confidently referred to the same species.
urn:lsid:zoobank.org:act:E74CC301-D5B9-4093-9194-BCE6B1C852D2
http://species-id.net/wiki/Permopsyche_rasnitsyni
Fig. 7A–DIn honorof an outstanding paleoentomologist, Prof. Dr. Alexandr Rasnitsyn.
PIN, no. 3840/1351, well-preserved forewing (part and counterpart), with apex missing.
PIN, no. 3840/733, 1363, 1371, incomplete forewings.
Isady locality, Vologda Province, North European Russia; Poldarsa Formation, uppermost Severodvinian (Lower Wuchiapingian), Upper Permian.
Similar to Permopsyche issadensis ingeneral venation scheme, but of larger size, with MA fork shortened, intercubital space widened, and wing membrane almost uncolored.
Forewing length/width ratio about 2.8–3:1; anterior margin nearly straight; apex obtuse to rounded. Costal space very narrow along its entire length. MA fork distinctly shorter than RS fork. MP unsclerotized for more than 1/2 of its length. CuA base long, nearly transverse, M5 present or lost. Intercubital space rather expanded. Wing membrane mostly uncolored, with small diffuse dark patch at center of wing.

A–D forewings of Permopsyche rasnitsyni gen. et sp. n. A–B photograph of holotype, PIN, no. 3840/1351, part and counterpart C line drawing of holotype, PIN, no. 3840/1351 D line drawing of paratype, PIN, no. 3840/1363 E–F Permopsyche sp., photograph and line drawing of specimen PIN, no. 3840/1556.
Holotype length, as preserved, 6.7, width, 2.5; paratypes lengths, ca. 7.0–8.5, width unmeasurable due to widthwise deformation.
Fig. 7E–F
PIN, no. 3840/1556, well-preserved forewing (part and counterpart).
The wing is 6 mm long, 2.1 mm wide, subtriangular, with anal area narrowed and posterior apical margin slightly oblique; MA forking before RS. The wing is entirely uncolored (neither part, nor counterpart showing any trace of coloration). The specimen may either be an aberrant specimen of Permopsyche issadensis, or represent a different species, but describing a separate species based on a single wing does not seem well justified.
http://species-id.net/wiki/Mesopsyche
Mesopsyche triareolata Tillyard, 1917; Upper Triassic, Australia.
Forewing relatively broad. SC rather long, with single or multiple fore branches. Costal space typically expanded. Both RS and MA forks not longer than their stems. MP 4-branched. CuA base from transverse to distinctly inclined backwards, usually situated much more distally than MP origin. Crossvein cua-cup of same inclination as CuA base. Intercubital space rather expanded. Crossveins not numerous, their arrangement rather stable. In hindwing, CuP convex with respect to membrane; fused with A1 very close to base.
Differs from Permopsyche by the base of CuA being inclined backwards, costal space widened, and hindwing CuP convex.
In addition to the type species, from the beds near the Permian–Triassic boundary of Kuznetsk Basin, Mesopsyche javorskii (Zalessky, 1935) comb. n.; from the Middle Triassic of China, Mesopsyche tongchuanensis (Hong, 2007) comb. n.; from the Middle–Upper Triassic Madygen Formation of Kyrgyzstan, Mesopsyche shcherbakovi, Mesopsyche justa, Mesopsyche ordinata, Mesopsyche tortiva, and Mesopsyche gentica (
urn:lsid:zoobank.org:act:FC61FFCE-6E4D-401F-BD62-227A6E376D1B
http://species-id.net/wiki/Mesopsyche_incompleta
Fig. 8Latin incompletus, “incomplete”.
PIN, no. 5103/286, hindwing (part and counterpart), with cubito-anal area missing.
PIN, no. 5103/122, incomplete forewing (basal half only), part and counterpart; PIN, no. 5103/123, incomplete forewing (anterior half only, with apex, basal and posterior parts missing), part and counterpart. In addition, two forewing fragments apparently belonging to the same species.
Balymotikha locality, Vyazniki town, Vladimir Province, Central European Russia; uppermost Vyatkian (Upper Changhsingian), Upper Permian.
Forewing. Anterior margin slightly convex. Costal space somewhat wider than subcostal one. Pterostigma distinct, lanceolate. SC not reaching the pterostigma, bearing one oblique distal branch; connected distally with R1 by short transverse crossvein. R1 sharply curved posteriad at origin of weak distal branch in pterostigmal area. RS forking somewhat before than MA. Thyridium at MP stem before fork. CuA base obscure, presumably curved backwards and located somewhat distal to M5. Crossvein cua-cup slightly sigmoid, distinctly curved backwards. Crossvein pattern typical of genus. Color pattern in form of sparse dark spots, mostly around crossveins.
Hindwing venation and coloration similar to those in forewing. SC slightly shortened, reaching level of RS+MA bifurcation, somewhat beyond wing midlength, forking apically. Pterostigma distinct. R1 nearly straight, turning apically towards anterior margin. R1 branch obscure. CuA distinctly convex along the whole length.

A–F, wings of Mesopsyche incompleta sp. n. A–B holotype PIN, no. 5103/286, hindwing, photograph and line drawing C–D paratype PIN, no. 5103/122, forewing, photograph and line drawing E–F paratype PIN, no. 5103/123, forewing, photograph and line drawing G–H Mesopsyche javorskii (Zalessky) comb. n., holotype hindwing, photograph (by D. Shcherbakov) and line drawing (orig.), not to scale A, C, G reversed left-right.
hindwing length of holotype, 9.8; forewing length of paratype PIN, no. 5103/122, 6.1, as preserved; paratype PIN, no. 5103/123, 7.9, as preserved.
The new species substantially differs from others by SC with a single, distally located fore branch.
The systematic position of Mesopsychidae and the entire “suborder” Paratrichoptera was discussed by
The finds described herein reveal main trends in the evolution of Mesopsychidae after their separation from the permochoristid-like ancestor. These are:
– Change in the forewing CuA base inclination and position: from proximal and inclined forwards (typical of Permochoristidae) to inclined backwards, and probably also shifted distad.
– Change in position of the hindwing CuA: from concave (Permopsyche) to convex (most Mesopsychidae).
– Loss of fusion of CuP and A1 veins in the hindwing: from quite basally fused in Permopsyche, through a rudimentary basal fusion in Mesopsyche (as indicated by
– Increase in the average wing size, from 5–6 mm in the Late Permian to 20–30 mm since the Late Triassic.
The Permian Mesopsychidae occurred together with Nedubroviidae (in Isady, Balymotikha, and probably Belmont;
New finds also cast light on the stratigraphic distribution of the Aneuretopsychina scorpionflies, which appear to be the most common group of Mecoptera during the latest Permian – early Middle Triassic time interval (viz. Mesopsychidae, Nedubroviidae, and later also Pseudopolycentropodidae). They initially replaced Permochoristidae in the end of Permian, and were in turn progressively supplanted during the Triassic – Early Jurassic by Parachoristidae and Orthophlebiidae, which gave rise to the modern mecopteran lineages. In the mid-Jurassic – Lower Cretaceous insect assemblages, Aneuretopsychina scorpionflies are rather rarely found, constituting only a minor component of the mecopteran faunas.
I am grateful to Prof. Alexandr Rasnitsyn and Dr Dmitry Shcherbakov for valuable comments and discussions, to Dr Louis A. Somma for reviewing the paper and important remarks, to Dr Yong Yi Zhen who kindly provided the photographs of Mesopanorpodes from the collection of the Australian Museum, Sydney, and to Dr Roman Rakitov for improving the English of manuscript. The research was supported by the Russian Foundation for Basic Research, projects 09-04-01241 and 10-04-01713, and by the Program 15 of the Presidium of the Russian Academy of Sciences “The Origin of the Biosphere and Evolution of the Geo-Biosystems”.