





(C) 2011 Wilfried Wichard. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Palerasnitsynus ohlhoffi gen. et sp. n. is described fromBurmese amber of late Albian (Lower Cretaceous) age. This is the first record of the family Psychomyiidae from Burmese amber, and the earliest fossil record of the family. The genus Palerasnitsynus gen. n. differs from all other known psychomyiid genera by the absence of fork III in the forewings.
Fossil Trichoptera, fossil insects, aquatic insects, fossil taxonomy, palaeoenvironment
Trichoptera are very rare in Burmese amber and previously only two species, Burminoptila bemeneha Botosaneanu, 1981 (Hydroptilidae) and Wormaldia myanmari Wichard & Poinar, 2005 (Philopotamidae), have been named. Burmese amber from Myanmar is late Albian (late Lower Cretaceous) in age (
Only seven fossil psychomyiid species have been described so far: Arkharia oblimata Sukatsheva, 1982 comes from Upper Cretaceous of south-eastern Siberia, originally placed in family Philopotamidae but belonging to the Psychomyiidae (
urn:lsid:zoobank.org:act:FD7212F5-2A0A-41AF-834B-B429CEF89188
Palerasnitsynus ohlhoffi sp. n.
The new genus is named after Prof. Dr. Alexandr P. Rasnitsyn on the occasion of his 75th birthday, in recognition of his extensive work on Palaeoentomology.
Genus Palerasnitsynus gen. n. differs from all extant genera (Paduniella Ulmer, 1913, Psychomyia Latreille, 1829, Psychomyiella Ulmer, 1908, Lype Mclachlan, 1878, Padangpsyche Malicky, 1993, Tinodes Curtis, 1834, Trawaspsyche Malicky, 2004, Eoneureclipsis Kimmins, 1955, and Zelandoptila Tillyard, 1924) and the extinct genus (Arkharia Sukatsheva, 1982) by the absence of apical fork III in the forewing.
Head: Ocelli absent. Antennae filiform, almost as long as forewings; each scapus only slightly longer than each pedicel and individual flagellomeres; flagellomeres cylindrical. Maxillary palps 5-segmented in both sexes, terminal 5th segment flexible, annulated, and at least twice as long as each of the other segments. Labial palps 3-segmented with flexible, annulated, long terminal segments.
Thorax: Mesoscutum with one pair of setal warts. Legs with tibial spurs 2/4/4. Wings elongate, rounded at apex, hind wings narrower than forewings. Forewing subcosta (Sc) simple, terminating into costa (C) at about three quarters the length of wing. Radius R1 simple; R2+3 unforked, R4+5 forking into R4 and R5 (fork II) close to potential crossvein r; discoidal cell (Dc) short. Media M1+2 unforked, M3+4 forking into M3 and M4 (fork IV); crossvein m indistinct; median cell (Mc) apparently open. Cubitus Cu1 bifurcated into Cu1a and Cu1b (fork V); Cu2 simple; crossvein m-cu apparently present, closing thyridial cell (Tc). Forewings with forks II, IV, and V.
urn:lsid:zoobank.org:act:7DBC53F5-0678-4639-8D36-C698DA87655C
http://species-id.net/wiki/Palerasnitsynus_ohlhoffi
Figs 1 –3Male embedded in Burmese amber, late Albian, Myanmar, deposited in the Staatliches Museum für Naturkunde Stuttgart (ex coll. Rainer Ohlhoff, Saarbrücken). The holotype is well preserved in slightly cloudy amber. The forewings cover the hindwings in dorsal view; the hindwing venation is difficult to analyze due to the overlapping hind wings and forewings. The male genitalia are visible in ventral view. The antennae and legs are completely preserved but the left mid leg is broken.
The species is named after Rainer Ohlhoff, who kindly made the fossil available for this study.
The only species of the genus Palerasnitsynus which differs from all other known psychomyiid genera by the absence of apical fork III in the forewings. Length of male 1.9mm (intraspecific variation not known).
Length of male (tip of head to tip of abdomen) 1.9 mm; forewing length 1.9 mm; hind wing length 1.7 mm.
Head: Eyes large and bulbous; ocelli absent; antennae filiform, almost as long as forewings, with cylindrical scapes, pedicel and flagellomeres. Maxillary palps 5-segmented, segment 1 short, segment 2 longer than segments 1 and 3, as long as segment 4, segment 5 flexible, annulated, twice as long as segment 4. Labial palps 3-segmented with long terminal segments, nearly as long as segments 1 + 2.
Thorax: Mesoscutum with pair of setal warts. Tibial spurs 2/4/4. Forewings: Subcosta (Sc) simple, Radius: R1 simple, R2+3 unforked, R4+5 forked at cross-vein r; discoidal cell short. Media: M1+2 simple, M3+4 forked. Crossvein m indistinct, median cell (Mc) apparently open. Cubitus: Cu1 forked; Cu2 unforked. Crossvein m-cu apparently present, defining thyridial cell (Tc). Forewings apically rounded; venation with apical forks II, IV and V. Light-coloured hindwing venation indistinct. Rs apparently forked into R2+3 and R4+5; forks I and II apparently absent. Crossvein r apparently absent. M branched, M1+2 unforked, M3+4 indistinct.
Genitalia: In ventral view, sternite IX with sinusoidal apical margin, bearing pair of two-segmented gonopods; coxopodite short, broad at basis; harpago long. Harpagones regularly curving mesad, widening at apex. Coxopodites with row of strong hairs along median margins; each coxopodite with brush of short, dark setae at apex. In dorsal view two small black spines about half as long as gonopods, visible through hyaline wings. External structures of male genitalia indistinct in lateral and dorsal view.

Palerasnitsynus ohlhoffi sp. n. holotype (male) in ventral view. Scale bar 1mm.
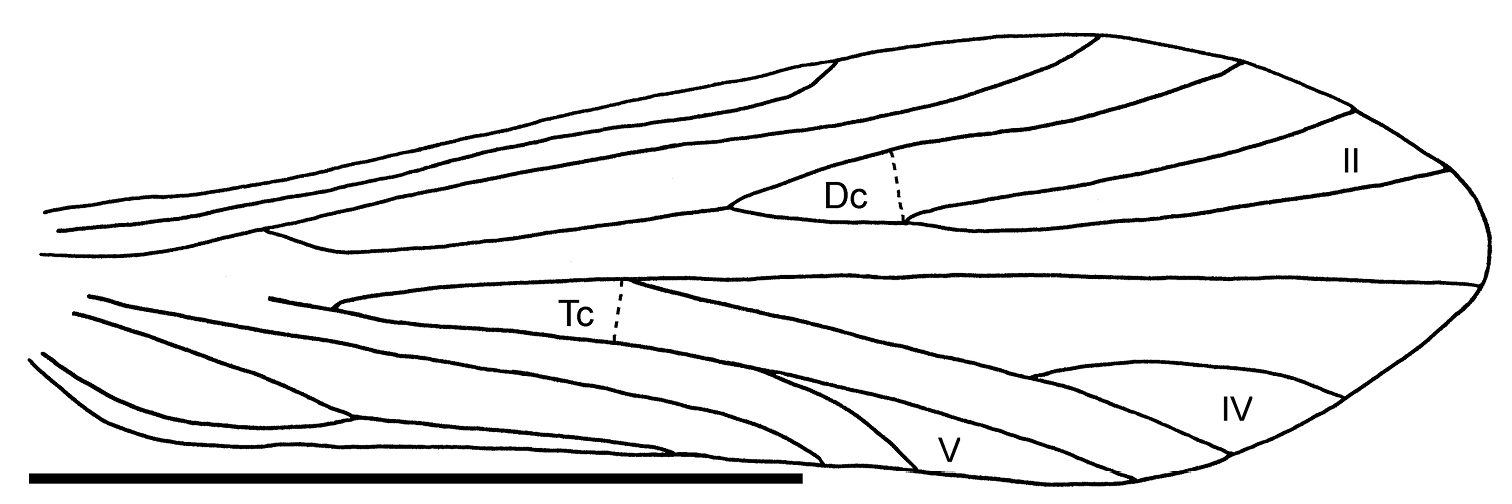
Palerasnitsynus ohlhoffi sp. n. male right forewing. Scale bar 1mm.
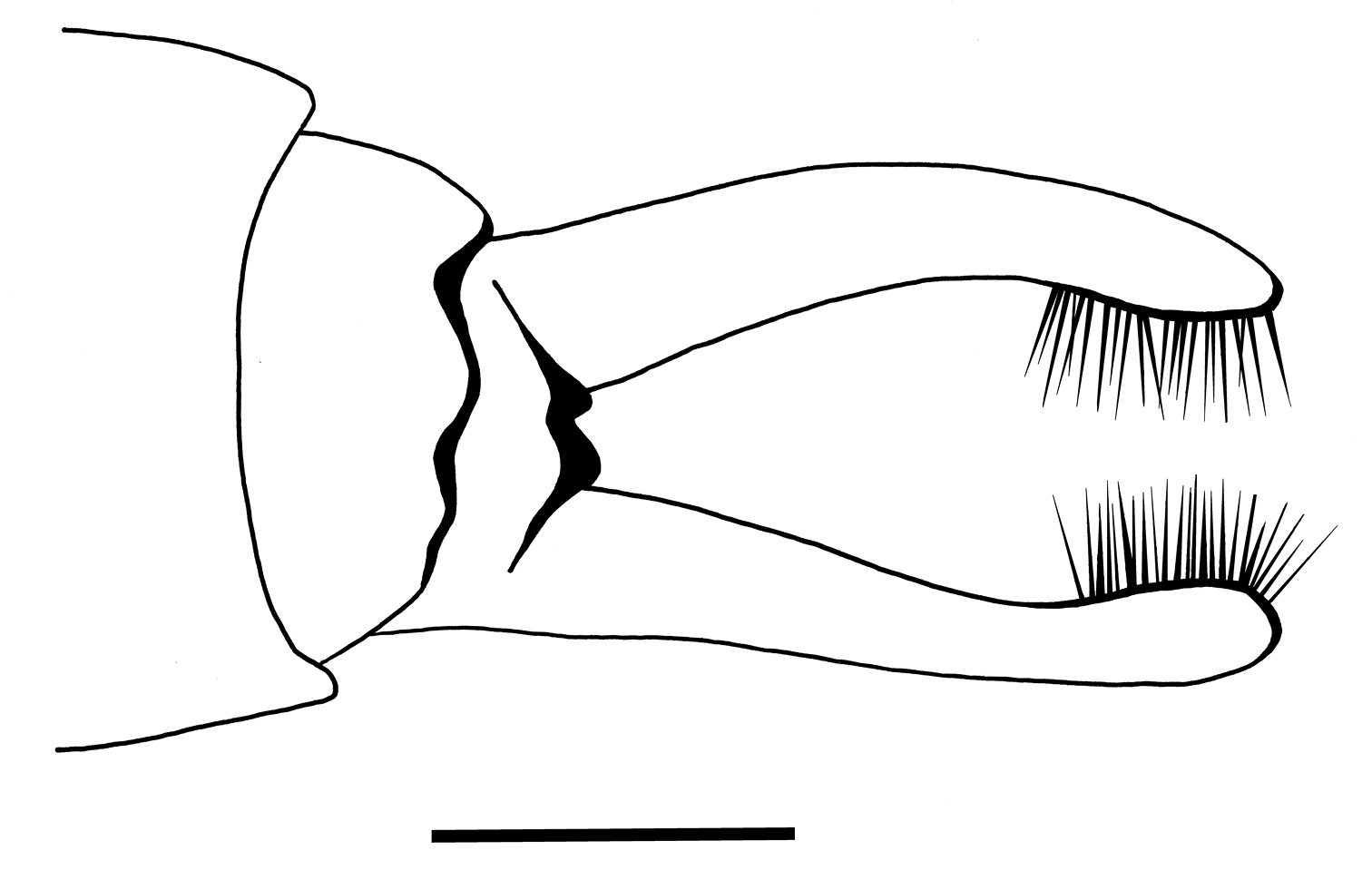
Palerasnitsynus ohlhoffi sp. n. male genitalia in ventral view. Scale bar 0.1mm.
Figs 4 –5
Female imago in Burmese amber, late Albian, Myanmar; deposited in National Museums Scotland, Edinburgh; specimen no. G.2010.20.35 (ex Scott Anderson Coll.).
Length of female (tip of head to tip of abdomen) 2.7 mm; forewing length 2.57 mm.
Head: Length 0.23 mm, width 0.47mm; filiform antennae each with 28 segments (including basal antennal segment), basal antennal segment twice as wide as other segments and twice as long as first segment; eyes large and bulbous, ocelli absent; maxillary palps 5-segmented, terminal segment flexible, annulated, and long (0.31mm), and nearly three times longer than each of the other segments (of similar length to each other); labial palps 3-segmented, also with long, flexible and annulated terminal segment.
Thorax: Mesoscutum with setal warts. Forelegs not easy to see; however, each may have 2 short spurs at end of tibia; mid legs each with 2 long spurs at end of tibia, 1 long preapical spur and possibly the base of a second; hind leg with 2 long spurs at end of tibia and 2 long preapical spurs. Tibial spurs: 2?/4?/4.
Wings with short fringe hairs on the anterior margin and long fringe hairs on posterior margin. Left forewing length 2.57mm; Sc dark and simple, terminating about three-quarters of length of anterior margin; R1 simple; Rs branching twice, first branch originating at mid-wing, R2+3 simple, R3+4 branched (= fork II); discoidal cell (Dc) present (crossvein faint); M branching twice, first branch originating at mid-wing, M1+2 simple, M3+4 branched (= fork IV); Cu branching distally (= fork V); anal veins not visible. Left hindwing apex obscured; Sc dark and simple, terminating about three-quarters of length of anterior margin; R1 simple, Rs branching once; crossvein r present, discoidal cell present; R4+5 connected to M1+2 by the crossvein r-m; M branching twice, M1+2 ending with short apical fork III, M3+4 simple; Cu branching once (= fork V), connected to M by the oblique crossvein m-cu closing thyridial cell apically; anal veins not visible. Forewings each with apical forks II, IV, and V; hind wings each with apical forks III and V.
Genitalia: Length 0.3mm. Typical long and simple ovipositor present; narrow cleft medio-ventrally and a pair of short apodemes apically.

Palerasnitsynus sp. female in ventral view. Scale bar 1mm.
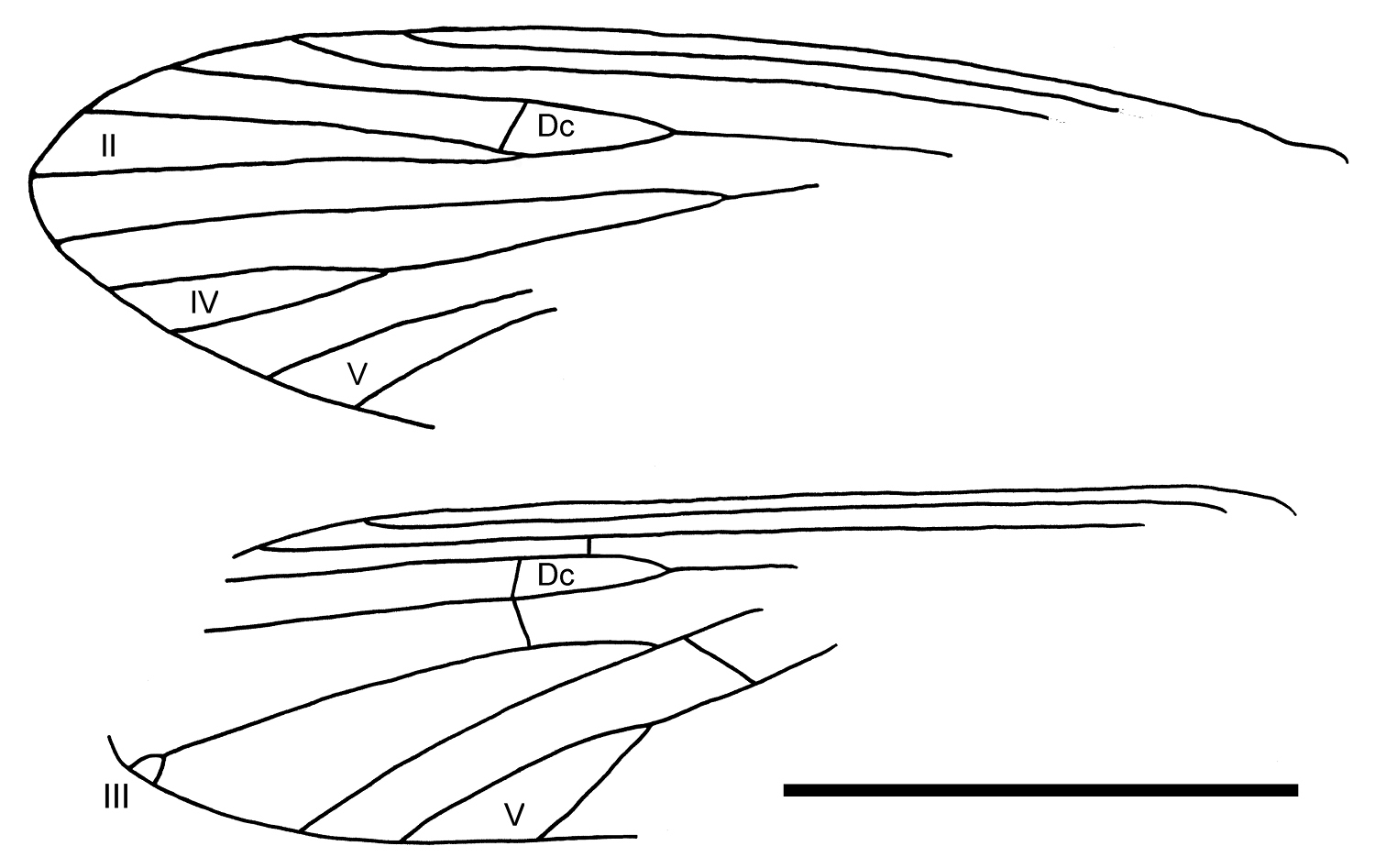
Palerasnitsynus sp. female left forewing (part) and left hind wing (part). Scale bar 1mm.
Due to the characteristic features (ocelli absent; maxillary palps 5-segmented and with terminal segment flexible and annulated; mesoscutum with one pair of setal warts; and tibial spurs probably 2/4/4), the female probably belongs to the Psychomyiidae. Due to the absence of forewing apical fork III, the specimen belongs to the genus Palerasnitsynus gen.n. The female and Palerasnitsynus ohlhoffi sp. n. agree in the forewing venation and differ in the hind wing; the discoidal cells are closed and short forks III are present in the female hind wings, but both traits are absent in Palerasnitsynus ohlhoffi sp. n. There is no evidence to identify this female specimen as Palerasnitsynus ohlhoffi sp. n.
The diversity of insects in Burmese amber is high compared with that of other Cretaceous ambers.
The low diversity of aquatic insects in Burmese amber might reflect that the late Albian palaeoenvironment had few freshwater habitats. This contrasts with Eocene Baltic amber diversity, comprising 25% amphibious aquatic insects (
Many thanks to Bill Crighton (National Museums Scotland) for taking the photograph of the female specimen (Fig. 4). Our special thanks go to John Morse (Clemson, USA) and an anonymous reviewer for constructive comments.