





(C) 2012 Luiz G. Rodrigues. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
A new species of Euragallia Oman, 1938 from Peru (Pasco Department) is described and illustrated. Euragallia batmani sp. n. can be distinguished from the other species of the genus by the very posteriorly pronounced male pygofer, with an apical hook-like projection, and by the well-developed dorsal area of the aedeagal base, resembling the open wings of a bat. With the addition of Euragallia batmani sp. n., the number of Euragallia species is increased to 21. Only one additional species of the genus is recorded from Peru (Euragallia prion Kramer, 1976). A comparison between the new species and Euragallia prion is provided. A conspicuous structure, which connects the subgenital plates to the styles, is described in detail and named.
Leafhopper, Megophthalminae, Membracoidea, morphology, South America, taxonomy
The subfamily Megophthalminae comprises approximately 53 genera and 650 species distributed in four tribes. In the Neotropical Region, about 22 genera and 280 species are known. The tribes are distributed as follows: Adelungiini has a Palearctic distribution; Megophthalmini, African and Holarctic; Evansiolini, restricted to Chile (Juan Fernandez Islands); and Agalliini (traditionally treated as a separate subfamily), cosmopolitan. The latter tribe, which is the largest within the subfamily and includes the genus herein treated, was historically studied mostly by
Euragallia Oman, 1938 (type-species Agallia furculata Osborn, 1923) appears to be limited to the tropical and subtropical regions of the Western Hemisphere, being recorded from Mexico to Brazil (
The most relevant diagnostic characters of Euragallia are as follows: (1) robust species, their coloration being brown or fuscous, never with bright spots; (2) crown shorter on median region than next to eyes; (3) eyes usually bulbous; (4) subgenital plates always fused to each other at least basally, frequently small, exposing apexes of styles; (5) styles never forked; (6) abdominal segment X usually well-developed; (7) seventh sternite of female reduced, sometimes exposing base of ovipositor (
In this paper, a new species of Euragallia is described and illustrated based on three males collected in Peru (Pasco Department). This is the second species of the genus recorded from Peru. The other recorded species is Euragallia prion Kramer, 1976. A brief discussion about similarities and differences between the new species and Euragallia prion is included. The new species has remarkable male genital features, including a peculiar structure connecting the subgenital plates to the styles, which is herein described.
Material and methodsThe studied specimens belong to the Museo de Historia Natural, Universidad Nacional Mayor de San Marcos (MUSM; Lima), Illinois Natural History Survey (INHS; Champaign), and Museu Nacional, Universidade Federal do Rio de Janeiro (MNRJ; Rio de Janeiro). Label data are given inside quotations with a reversed virgule (\) separating lines. Morphological terminology follows mainly
urn:lsid:zoobank.org:act:6FFFBBDF-C9AD-4DBE-9378-AC942304551A
http://species-id.net/wiki/Euragallia_batmani
Figs 1–11Male holotype, 6.1 mm; male paratypes, 6.4–6.5 mm (n = 2).
Head and thorax. Ground color of dorsum mostly yellow to brown. Crown, in dorsal view (Fig. 1), distinctly shorter medially than next to eyes; each side with fovea closer to inner eye margin than to median coronal line, each fovea with adjacent black macula; median portion of crown with black spot. Face (Fig. 3) with distinct inverted Y-shaped dark brown macula; surface depressed medially; distance between ocelli greater than distance between each ocellus and adjacent inner eye margin; ocelli slightly closer to median line of face than to adjacent inner eye margin, each ocellus with adjacent fovea and small, inconspicuous dark brown macula. Frons (Fig. 3) mostly yellow; lateral margins bordered by black line; disk with somewhat M-shaped figure formed by numerous small dark brown maculae. Genae (Fig. 3) mostly yellow, with one dark brown macula adjacent to lorum and another adjacent to antennal base; surface striated. Clypeus (Fig. 3) with transverse dark brown line at base and median dark brown macula; with small apical setae. Pronotum (Fig. 1) with pair of black-marked depressions on anterior margin; disk weakly punctured and finely transversely striated; with pair of large dark brown triangular maculae on posterior half, dark brown longitudinal line extending from anterior to posterior margin on middle portion, and dark brown macula between apex of each triangular macula and median line. Basal portion ofmesonotum (Fig. 1) with triangular dark brown macula located medially and pair of larger, lateral dark brown triangular maculae, these three maculae delimiting yellow, somewhat M-shaped area; pair of dark brown spots close to transverse sulcus. Forewings (Figs 1, 2) opaque on basal third and along costal area until base of first apical cell, remainder of surface translucent; venation pronounced; claval veins almost entirely marked with pale yellow; corium veins marked with pale yellow on basal third, remainder mostly dark brown; claval apex marked with pale yellow.
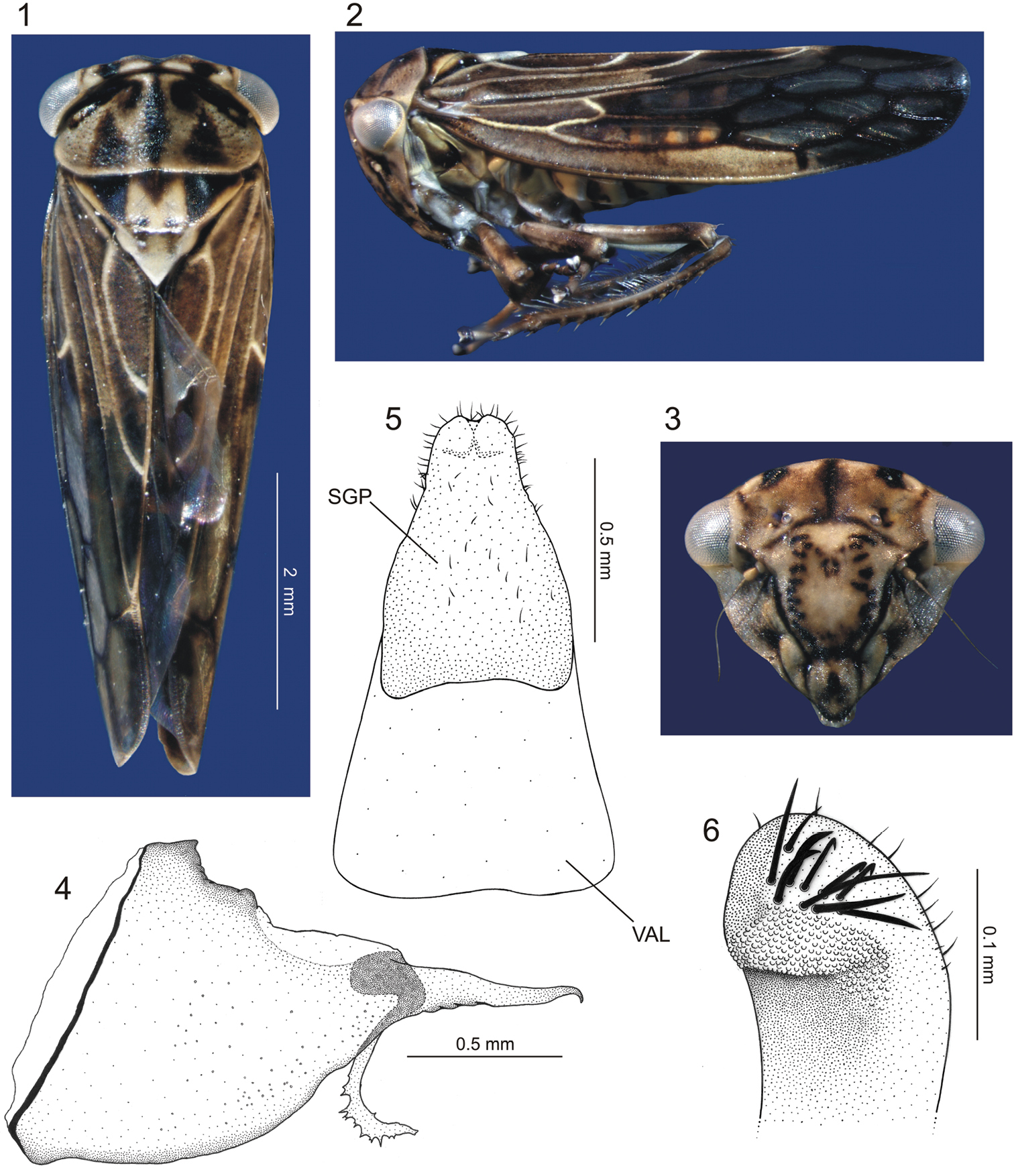
Euragallia batmani sp. n., body and external male genitalia. 1 body, dorsal view 2 body, lateral view 3 face, frontal view 4 pygofer, lateral view 5 valve (VAL) and (fused) subgenital plates (SGP), ventral view 6 apical portion of subgenital plate, laterodorsal view.
Metathoracic leg chaetotaxy. Row I of tibia with ten primary setae, equal in size and shape, bases slightly elevated. Row II with seven primary setae, bases elevated. Row III with eight primary setae, most proximal one almost aligned with third seta of row II. Row IV with approximately 25 primary elongate setae, increasing in size towards tibia apex; intercalary setae present; distal setae indistinct from the others of the row. Apex of tibia with distal transverse row formed by conspicuous spurs. Plantar surface of first tarsomere with small setae uniformly distributed; apex bearing one conspicuous platella.
Male genitalia. Pygofer (Fig. 4), in lateral view, pronounced posteriorly, forming very long projection with about half of pygofer length; apex of projection hook-like; inner dorsal margin with elongate process extending ventrally and then gradually curved posteriorly, armed apically with irregular dentiform projections. Valve (Fig. 5) trapezoidal, fused to subgenital plates. The latter (Fig. 5), in ventral view, completely fused to each other, subtriangular, with slight apical emargination; surface with small scattered setae; apical portion, in lateral view, curved dorsally, with some well-developed setae and scale-like sculpture (Fig. 6). Connective membranous. Styles (Fig. 7), in dorsal view, expanded on apical third, club-shaped, bearing small apical claw-like projection directed inward; basal portion of style with elongate sclerotized projection, which is fused apically to the dorsal apical portion of subgenital plate and lays on a groove on outer style margin (Fig. 7 – PSC [plate-style connective]). Aedeagus (Fig. 8) symmetrical; shaft, in lateral view, strongly flattened dorsoventrally, base directed ventrally, then gradually curved dorsally; apical portion directed posterodorsally, bearing pair of subapical, slender lateral spiniform processes (Fig. 10) extending anteriorly for short distance; base of aedeagus, in dorsal view, with dorsal region well-developed, bearing lateral sclerotized projections resembling the open wings of a bat (Fig. 9 – WLP); base of aedeagus (Fig. 8), in lateral view, with spiniform projection directed dorsally. Segment X (Fig. 11), in lateral view, strongly sclerotized, bearing pair of small apical processes on each side.
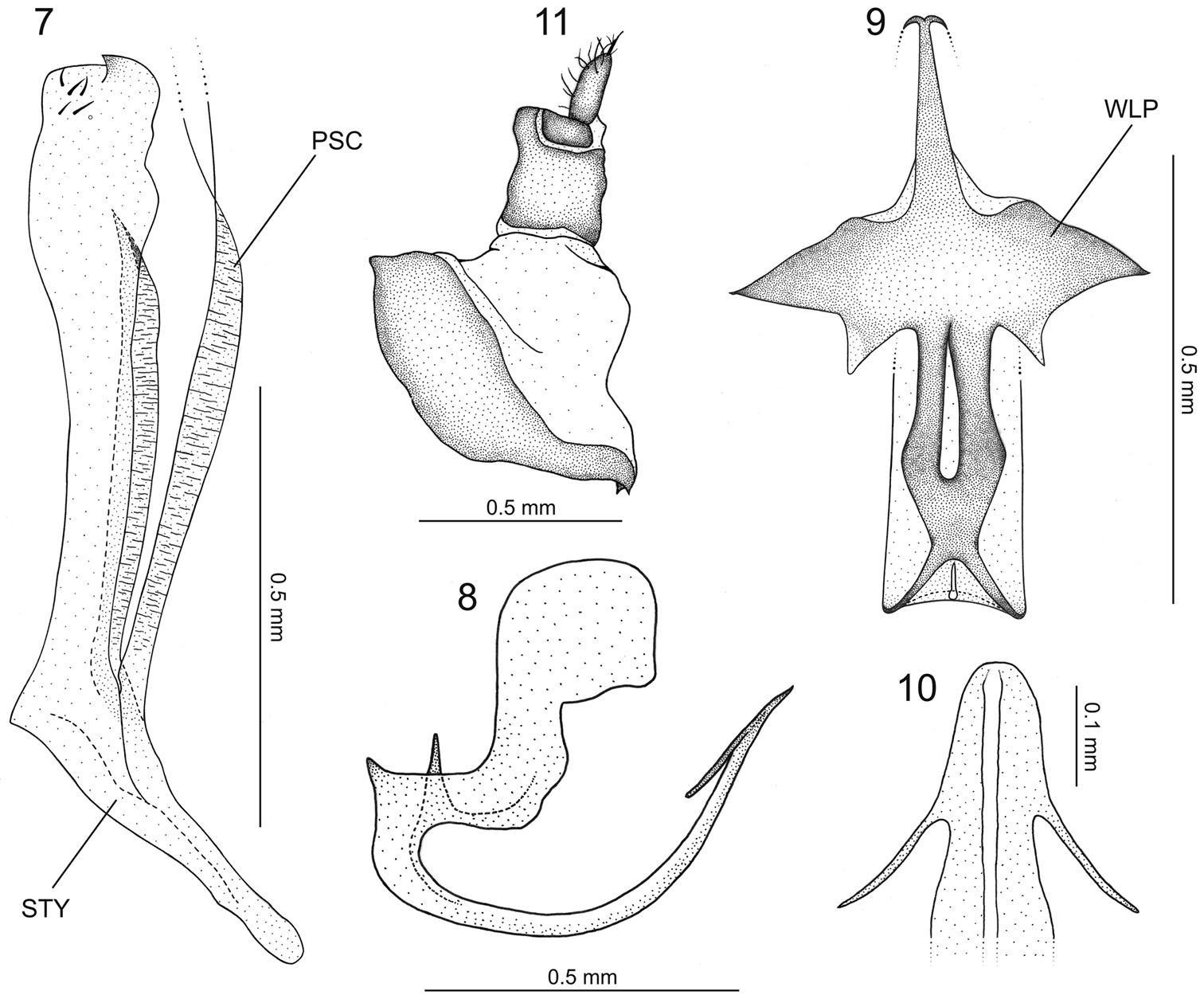
Euragallia batmani sp. n., internal male genitalia and anal tube. 7 style (STY), dorsal view (PSC = plate-style connective) 8 aedeagus, lateral view 9 dorsal region of basal portion of aedeagus, dorsal view (WLP = wing-like projection) 10 apical portion of aedeagus, caudal view 11 anal tube, lateral view.
Female unknown.
The number of primary setae on the rows of the metathoracic tibia may vary slightly; for instance, the number of setae on row III may be eight (holotype) or nine (one paratype). Additional cross veins may be present on the forewings; for instance, the median anteapical cell may be divided by a cross vein. The basal portion of the forewings may be more translucent than in the holotype.
The specific epithet, batmani, is a reference to the dorsal region of the aedeagal base in dorsal view (Fig. 9), which closely resembles the open wings of a bat, like those of the Batman symbol.
Peru, Pasco Department. Holotype: male, “Peru: Pasco, Yanachaga-Chemil- \ lén N.P., Huampal Stn. 10°11'9"S, 75°34'27"W, 1050m, 6–9 X 2002 \ D. Takiya, C. Peña, R. Rakitov \ Malaise trap acr. R. Huancabamba” (MUSM). Paratypes: two males with same data as the holotype (INHS, MNRJ).
Among the Euragallia species, Euragallia prion seems to be the most similar to Euragallia batmani sp. n. In
The basal portion of the styles of the new species bears a conspicuous, sclerotized elongate projection, which lays on a groove on the outer style margin. This projection, which is fused apically to the dorsal apical portion of the subgenital plate, is herein named the plate-style connective (Fig. 7 – PSC). This structure was previously recorded in only three species of the genus, Euragallia goemansi Gonçalves & Zanol, 2010, Euragallia adelinae Gonçalves & Zanol, 2010, and Euragallia nervata (Oman, 1934), being described and illustrated in the first two by
The authors are thankful to Christopher Dietrich (INHS) for making the specimens under his care available for study. Fellowships from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) to LGR, ACG, and GM are acknowledged. This study was supported in part by Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ) (grant number E-26/171.281/2006 to Márcia Couri). The manuscript benefited from the useful comments of Michael Webb and one anonymous reviewer.