





(C) 2011 Victor H. Gonzalez. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Two distinctive new species of the Neotropical stingless bee genus Paratrigona Schwarz from Ecuador and Paraguay are described and figured. The Ecuadorian species, Paratrigona scapisetosa sp. n., belongs to the haeckeli-lineatifrons group and is easily distinguished from its congeners by the unique shape and pubescence of the antennal scape, which is distinctly convex on its outer margin and bears thick, long, simple hairs along its inner margin. The Paraguayan species, Paratrigona wasbaueri sp. n., belongs to the lineata group and is easily distinguished by the pattern of body pubescence in both sexes and male genitalic characters. The male of the Mesoamerican species Paratrigona ornaticeps (Schwarz) is described and figured. New geographical records for Paratrigona impunctata and Paratrigona opaca, and an updated key to the haeckeli-lineatifrons and lineata species groups are provided.
Anthophila, Ecuador, Meliponini, Neotropics, Paraguay, stingless bees, taxonomy
Stingless honey bees (Meliponini)
are among the most commonly encountered bees in the tropics of the
world. In the Americas, they are found from Mexico to Brazil and
northern Argentina,
inhabiting a diverse variety of ecosystems, including both humid and
xeric lowlands to cloud forests and Páramos in the Andes reaching up to
4000 m of altitude (
There are about 500 species of stingless bees worldwide, most of them (80%) in the Western Hemisphere (
Summary of generic and subgeneric classification of New World Meliponini including fossils. It follows
| Taxa | Species | Distribution | |
|---|---|---|---|
| Genus Cephalotrigona Schwarz | 5 | CA, SA | |
| Genus Cretotrigona Engel‡ | 1 | NA | |
| Genus Duckeola Moure | 2 | SA | |
| Genus Frieseomelitta Ihering | 16 | CA, SA | |
| Genus Geotrigona Moure | 21 | CA, SA | |
| Genus Lestrimelitta Friese | 21 | CA, SA | |
| Genus Melipona Illiger | 68 | CA, SA | |
| Genus Meliwillea Roubik, Segura & Camargo | 1 | CA | |
| Genus Nannotrigona Cockerell | 10 | CA, SA | |
| Genus Nogueirapis Moure | 4* | CA, SA | |
| Genus Oxytrigona Cockerell | 11 | CA, SA | |
| Genus Paratrigona Schwarz | 33 | CA, SA | |
| Genus Paratrigonoides Camargo & Roubik | 1 | SA | |
| Genus Partamona Schwarz | |||
| Subgenus Parapartamona Schwarz | 7 | SA | |
| Subgenus Partamona Schwarz | 34 | CA, SA | |
| Genus Plebeia Schwarz | 40 | CA, SA | |
| Genus Proplebeia Michener† | 4 | CA | |
| Genus Scaptotrigona Moure | 21 | CA, SA | |
| Genus Scaura Schwarz | 7 | CA, SA | |
| Genus Schwarziana Moure | 2 | SA | |
| Genus Tetragona Lepeletier de Saint Fargeau & Serville | 19 | CA, SA | |
| Genus Tetragonisca Moure | 4 | CA, SA | |
| Genus Trichotrigona Camargo & Moure | 1 | SA | |
| Genus Trigona Jurine | 32 | CA, SA | |
| Genus Trigonisca Moure | 39 | CA, SA | |
The purpose of this paper is to describe two distinctive new species of Paratrigona, one each from Ecuador and Paraguay, and the male of the Mesoamerican species Paratrigona ornaticeps
(Schwarz). We also provide new geographical records and an updated key
to these species. Bees of this genus are small (4–6 mm in body length),
often with a dull, dark brown to black integument and conspicuous,
contrasting maculae on the head and mesosoma. There are 32 currently
recognized species ranging from Mexico to southern Brazil, most of them
known from the worker caste (Table 2). Although Meliponini
taxonomy is based on workers, the study of the nesting behavior,
males, and queens may provide additional characters useful in
recognizing cryptic species and in phylogenetic analyses. Unfortunately,
the nest and the male of Paratrigona are known for less than half of the species while the queen is only known for two species (
Summary of currently included species in Paratrigona
with information on the known sexes, nest and distribution. Sex/caste:
♀ = worker; ♂ = male; ‡ = queen. Nest: A = aerial; C = inside tree
cavity; F = inside ant nest; S = subterranean; T = inside termite nest; -
= unknown. The distribution and nesting site are based on
| Species | Sex | Nest | Distribution |
|---|---|---|---|
| “guatemalensis species group” | |||
| Paratrigona anduzei (Schwarz, 1943) | ♀♂ | A | Colombia, Venezuela |
| Paratrigona eutaeniata Camargo & Moure, 1994 | ♀♂‡ | A | Colombia |
| Paratrigona guatemalensis (Schwarz, 1938) | ♀ | A | Costa Rica, Guatemala, Mexico |
| Paratrigona lophocoryphe Moure, 1963 | ♀♂ | - | Colombia, Costa Rica, Panama |
| Paratrigona rinconi Camargo & Moure, 1994 | ♀ | - | Colombia |
| “haeckeli-lineatifrons species group” | |||
| Paratrigona compsa Camargo & Moure, 1994 | ♀ | - | Brazil |
| Paratrigona euxanthospila Camargo & Moure, 1994 | ♀ | - | Brazil |
| Paratrigona femoralis Camargo & Moure, 1994 | ♀ | - | French Guiana |
| Paratrigona guigliae Moure, 1960 | ♀ | - | Bolivia |
| Paratrigona haeckeli (Friese, 1900) | ♀♂ | - | Brazil, Peru |
| Paratrigona lineatifrons (Schwarz, 1938) | ♀ | T | Brazil |
| Paratrigona melanaspis Camargo & Moure, 1994 | ♀ | - | Brazil |
| Paratrigona onorei Camargo & Moure, 1994 | ♀ | - | Bolivia, Ecuador |
| Paratrigona ornaticeps (Schwarz, 1938) | ♀♂ | C | Costa Rica, Panama |
| Paratrigona scapisetosa Gonzalez & Griswold sp. n. | ♀ | - | Ecuador |
| Paratrigona uwa Gonzalez & Vélez, 2007 | ♀ | - | Colombia |
| “impunctata species group” | |||
| Paratrigona impunctata (Ducke, 1916) | ♀♂ | T | Brazil, Colombia, Ecuador, French Guiana, Guyana, Peru, Suriname |
| Paratrigona isopterophila (Schwarz, 1934) | ♀♂ | - | Colombia, Panama |
| “lineata species group” | |||
| Paratrigona crassicornis Camargo & Moure, 1994 | ♀ | - | Brazil |
| Paratrigona incerta Camargo & Moure, 1994 | ♀ | - | Brazil |
| Paratrigona glabella Camargo & Moure, 1994 | ♀♂ | - | Argentina, Bolivia |
| Paratrigona lineata (Lepeletier, 1836) | ♀♂ | S | Bolivia, Brazil |
| Paratrigona nuda (Schwarz, 1943) | ♀ | F | Bolivia, Peru |
| Paratrigona prosopiformis (Gribodo, 1893) | ♀♂ | - | Bolivia, Brazil, Colombia, Ecuador, Peru |
| Paratrigona subnuda Moure, 1947 | ♀♂ | S | Brazil |
| Paratrigona wasbaueri Gonzalez & Griswold sp. n. | ♀♂ | - | Paraguay |
| “peltata-opaca species group” | |||
| Paratrigona catabolonota Camargo & Moure, 1994 | ♀ | - | Brazil |
| Paratrigona lundelli (Schwarz, 1938) | ♀ | - | Belize, Guyana |
| Paratrigona myrmecophila Moure, 1989 | ♀ | F | Brazil |
| Paratrigona opaca (Cockerell, 1917) | ♀♂ | F | Colombia, Costa Rica, Ecuador, Guatemala, Mexico, Panama |
| Paratrigona pacifica (Schwarz, 1943) | ♀♂‡ | A | Bolivia, Brazil, Colombia, Ecuador, Peru |
| Paratrigona pannosa Moure, 1989 | ♀ | - | Brazil, French Guiana |
| Paratrigona peltata (Spinola, 1853) | ♀ | - | Brazil |
| Incertae sedis | |||
| Paratrigona permixta Camargo & Moure, 1994 | ♀ | - | Venezuela |
Morphological terminology follows that of
Institutional acronyms used herein are:
BBSL U.S. National Pollinating Insects Collection, Bee Biology and Systematics Laboratory, Utah State University, Logan, UT, USA.
DZUP Departamento de Zoologia, Universidade Federal do Paraná, Brazil.
SEMC Snow Entomological Collection, Division of Entomology, University of Kansas Natural History Museum, Lawrence, Kansas, USA.
USNM National Museum of Natural History, Washington, DC, USA.
Systematics Tribe Meliponini Lepeletier de Saint Fargeau, 1836.Genus Paratrigona Schwarz, 1938
urn:lsid:zoobank.org:act:E7B29A75-9CEE-4245-877C-276952559457
http://species-id.net/wiki/Paratrigona_scapisetosa
Figs 1–4♀, Ecuador: Mor. [Morona] Santiago, Miazal, 50 km. SE Macas, 1-4/7-1-1993, 300 m., M & J. Wasbauer Coll. (BBSL).
This species belongs to the haeckeli-lineatifrons species group sensu

Female holotype of Paratrigona scapisetosa Gonzalez and Griswold, sp. n. (1–4), and Paratrigona haeckeli (5). 1–3 Lateral, dorsal, and frontal views, respectively 4, 5 Antennal scape in profile view.
Worker: Total body length 4.3 mm; head
width 1.7 mm; forewing length 3.5 mm. Head slightly wider than long;
inner orbits of compound eyes converging below (Fig. 3);
malar area subequal to width of third flagellomere; clypeus about twice
as broad as long; interalveolar distance equal to median ocellar
diameter, shorter (0.8x) than alveolar diameter; alveolocular distance
1.7 times alveolar diameter, 2.1 times interalveolar distance;
interocellar distance about two times median ocellar diameter, 1.2
times longer than ocellocular distance; ocelloccipital distance about
1.3 times median ocellar diameter; scape 3.3 times longer than wide,
1.5 times broader than width of third flagellomere, distinctly convex
on outer margin, abruptly narrowed on basal one-fourth; pedicel about
as long as broad, slightly shorter than first flagellomere;
flagellomeres slightly broader than long, except apical flagellomere
much longer than broad (Fig. 4);
compound eye 2.7 times longer than broad; gena 0.7 times narrower than
compound eye in profile. Scutum with anterior margin rounded, 1.2 times
broader than long, about twice as long as scutellum; scutellum 1.4
times broader than long, narrowly rounded on distal margin
(long-paraboloid sensu
Integument in general finely and densely punctate, dull, as in other species of the genus; propodeum without impunctate midline.
Color predominantly ferruginous, except: black on upper two-thirds of frons, apex of scape, pedicel, flagellum; dark reddish brown on subantennal area, epistomal sulcus, malar area, mandible distally (darker on teeth), mesepisternum ventrally, hind tibia distally, outer surfaces of middle and hind basitarsi; yellow maculations as follows: mandible basally, labrum, scape, face (Fig. 3), gena and vertex with complete, thin band (at most as wide as median ocellar diameter), pronotum, pronotal lobe, anterior one-third of tegula, costal sclerite, mesepisternum laterally, metepisternum entirely, propodeum laterally, scutum laterally with broad band (as wide as median ocellar diameter), axilla, lateral and distal margins of scutellum with narrower band than on scutum, apices of middle and hind femora, outer surfaces of fore and middle tibiae with narrow band, hind tibia with band on posterior margin, fore and middle basitarsi basally. Wing membrane subhyaline with weak greenish and coppery reflections; veins and stigma dark brown (Figs 1, 2).
Body sparsely covered with minute, appressed, simple yellowish to whitish hairs except as follows: inferior margin of mandible, labrum, and hypostomal area with long (about as long as median ocellar diameter), erect hairs; scape with stiff, erect, simple hairs (0.5 times width of third flagellomere) along inner margin (Fig. 4); gena laterally with few stiff, erect hairs as those on scape; mesepisternum ventrally, middle and hind coxae, sterna with much longer, erect simple hairs (about twice as long as median ocellar diameter); sides of propodeum with dense, branched, semierect hairs (0.5 times median ocellar diameter) partially obscuring integument.
Male: Unknown.
Queen: Unknown.
The specific epithet is a combination of the Latin words “scāpus” and “saetae”, in reference to the distinctly long hairs along the inner margin of the antennal scape.
An additional female specimen of Paratrigona scapisetosa from Sierra de Chiribiquete in Colombia (Department of Caquetá) has been examined by one of us (VHG) but it was not available for study. The propodeum laterally with yellow maculations of Paratrigona scapisetosa is a character only shared with Paratrigona ornaticeps.
urn:lsid:zoobank.org:act:EB384673-259D-46E3-B2F6-825BE6088FC2
http://species-id.net/wiki/Paratrigona_wasbaueri
Figs 6–19♀, Paraguay: San Pedro, Cororo-Rio Ypane, XII-5/9-1983, Malaise Trap, M. Wasbauer coll. (BBSL).
54♀, 28♂, with same data as holotype but collected on November 24–30, and December 1–4 (BBSL, DZUP, SEMC).
This species belongs to the lineata species group sensu

Female holotype of Paratrigona wasbaueri Gonzalez and Griswold, sp. n. 6–8 Lateral, dorsal, and frontal views, respectively 9 Mesepisternum dorsally in profile 10 Apex of scutum, axilla, and scutellum in profile.
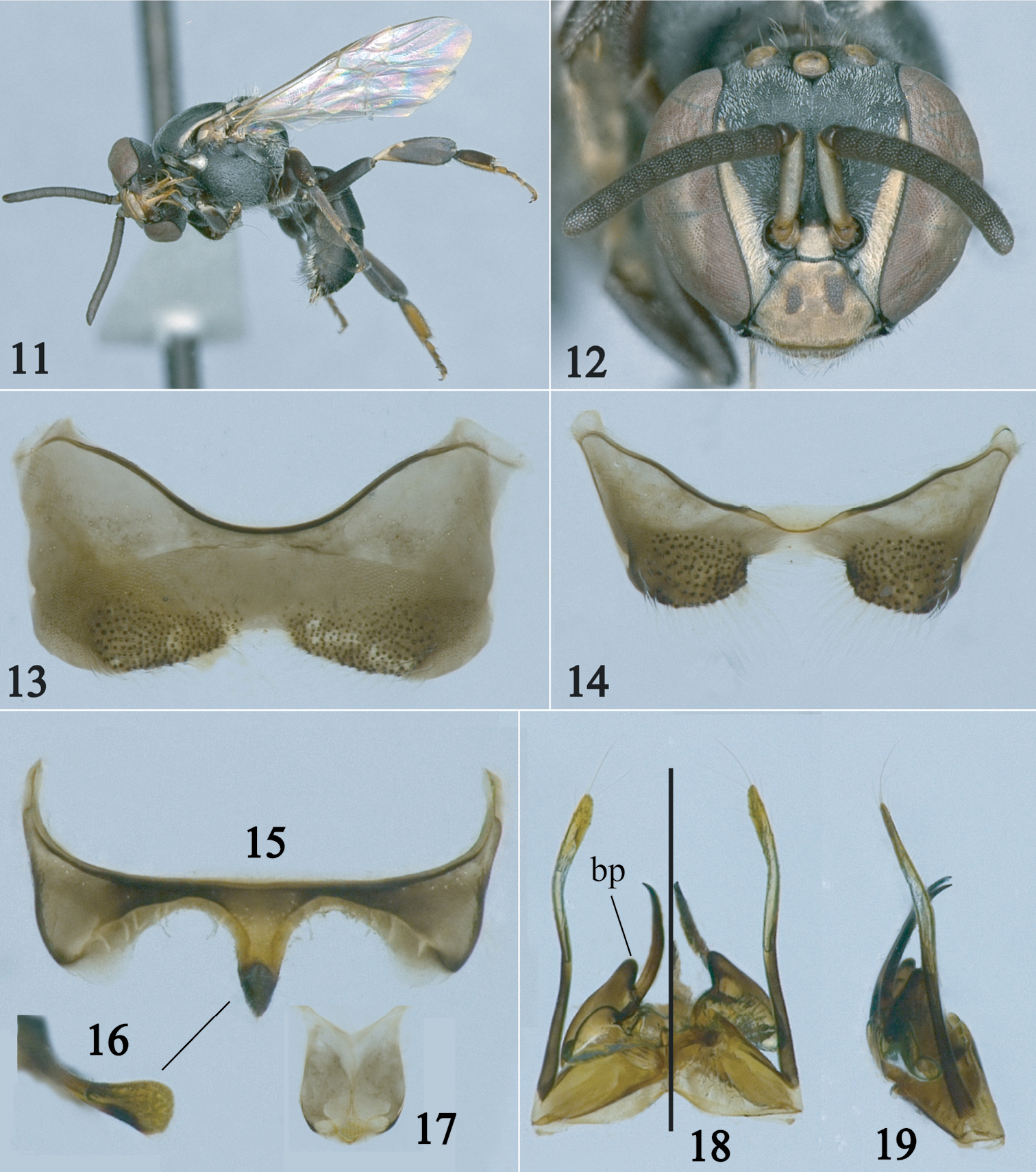
Male of Paratrigona wasbaueri Gonzalez and Griswold, sp. n. 11 Lateral habitus 12 Facial view 13 Fourth sternum 14 Fifth sternum 15 Sixth sternum 16 Apex of sixth sternum in profile 17 Seventh sternum 18 Genitalia in dorsal (left half) and ventral (right half) views 19 Genitalia in profile. bp = basal projection of penis valve.
Worker: Total body length 4.2 mm
(3.8–5.2 mm); head width 1.6 mm (1.6–1.7 mm); forewing length 3.6 mm
(3.6–3.8 mm). Head slightly wider than long; inner orbits of compound
eyes converging below (Fig. 8);
malar area about half width of third flagellomere; clypeus 1.8 times
broader than long; interalveolar distance 1.2 times median ocellar
diameter, about equal to alveolar diameter; alveolocular distance 1.3
times alveolar diameter; interocellar distance 2.1 times median ocellar
diameter, 1.8 times longer than ocellocular distance; ocelloccipital
distance about 1.3 times median ocellar diameter; scape 6.3 times longer
than wide, narrower (0.8 times) than width of third flagellomere;
pedicel about as long as broad, slightly shorter than first
flagellomere; flagellomeres slightly broader than long, except apical
flagellomere much longer than broad; compound eye 2.8 times longer than
broad; gena 0.8 times narrower than compound eye in profile. Scutum with
anterior margin rounded, slightly broader than long, 2.8 times longer
than scutellum; scutellum about twice as long as broad, broadly
rounded on distal margin (short-semicircular sensu
Integument in general finely and densely punctate, dull, as in other species of the genus; propodeum without impunctate midline.
Color black, except: light reddish brown on mandible (darker on teeth), tegula, margins of middle and posterior basitarsi, distitarsi; yellow maculations as follows: labrum, scape, face (Fig. 8), pronotum dorsally with medially interrupted band, pronotal lobe, scutum laterally with narrow band (about half width median ocellar diameter), axilla, lateral and distal margins of scutellum with broader band than on scutum, tibiae basally. Wing membrane hyaline with weak greenish and coppery reflections; veins and stigma dark brown (Figs 6, 7).
Body sparsely covered with minute, appressed, simple whitish hairs except as follows: inferior margin of mandible, labrum, vertex, hypostomal area, mesepisternum dorsally, distal margin of scutellum dorsally and ventrally, sides of propodeum with long (about as long as median ocellar diameter) erect hairs; mesepisternum ventrally, middle and hind coxae, sterna with much longer, erect simple hairs (about twice as long as median ocellar diameter); sides of propodeum with dense, branched, semierect hairs (0.5 times median ocellar diameter) partially obscuring integument; inner surface of tarsi with yellowish to ferruginous hairs.
Male: As in female except for longer body pubescence, brownish legs, and the following: Total body length 4.4–5.3 mm; head width 1.6 –1.7 mm; forewing length 3.7–3.8 mm. Inner orbits of compound eyes strongly converging below (Fig. 12); malar area linear; clypeus 1.3 times broader than long; interalveolar distance subequal to median ocellar and alveolar diameters individually; alveolocular distance about as long as alveolar diameter; interocellar distance about twice median ocellar diameter, 2.8 times longer than ocellocular distance; ocelloccipital distance about 0.7 times median ocellar diameter; scape 3.6 times longer than wide, slightly narrower than width of third flagellomere; pedicel longer than first flagellomere; first flagellomere about twice as broad as long, 0.5 times shorter than second flagellomere; gena about half width of compound eye in profile. Hind tibia about 3 times longer than broad; hind basitarsus 3 times longer than broad. Genitalia and associated sterna as in Figs 13–19.
Queen: Unknown.
The specific epithet is a patronym honoring Marius Wasbauer, friend and colleague, who has contributed much of the material we have studied, including the species described in this paper.
In most paratypes the yellow maculations are pale, discolored, most likely due to preservation in alcohol for an extended period.
http://species-id.net/wiki/Paratrigona_ornaticeps
Figs 20–27The female of this species can be separated from other species of the haeckeli-lineatifrons
group by the following combination of characters: frons with broad and
long inverted Y-shaped maculation; paraocular area with complete, long,
broad maculation; supraclypeus yellow; and clypeus with broad,
inverted T-shaped maculation (cf.,
The male of Paratrigona ornaticeps can be separated from that of Paratrigona haeckeli, the only other male known of the haeckeli-lineatifrons group (Table 2),
by the sixth sternum, which has a broader midapical projection, and
the shape of the gonostylus, which is more strongly curved apically in
profile (compare Figs 26 and 27 with

Male of Paratrigona ornaticeps (Schwarz). 20 Lateral habitus 21 Facial view 22 Fourth sternum 23 Fifth sternum 24 Sixth sternum 25 Seventh sternum 26 Genitalia in dorsal (left half) and ventral (right half) views 27 Genitalia in profile.
Male: Total body length 4.7–5.1 mm;
head width 1.6 –1.7 mm; forewing length 3.7–3.8 mm. Head slightly wider
than long; inner orbits of compound eyes strongly converging below (Fig. 21);
malar area very short, about one-fourth width of third flagellomere;
clypeus 1.5 times broader than long; interalveolar distance slightly
shorter (0.9x) than median ocellar and alveolar diameters individually;
alveolocular distance 1.3 times alveolar diameter, 1.2 times
interalveolar distance; interocellar distance 1.5 times median ocellar
diameter, 2.7 times longer than ocellocular distance; ocelloccipital
distance about 0.7 times median ocellar diameter; scape 2.8 times longer
than wide, slightly broader than width of third flagellomere; pedicel
about as long as broad, slightly longer than first flagellomere; first
flagellomere 1.5 times broader than long, 0.7 shorter than second
flagellomere, remaining flagellomeres slightly broader than long,
except apical flagellomere much longer than broad; compound eye 2.6
times longer than broad; gena about half width of compound eye in
profile. Scutum with anterior margin rounded, 1.1 times broader than
long, 2.3 times longer than scutellum; scutellum 1.5 times broader than
long, broadly rounded on distal margin (long-paraboloid sensu
Integument in general finely and densely punctate, dull, as in other species of the genus; propodeum with faint midline.
Color predominantly black, except: brownish on anterior surface of flagellum, tegula, legs (legs light yellow to ferruginous with pale maculations); yellow maculations as follows: mandible, labrum, anterior surface of scape, face as in Fig. 21 (frons sometimes maculate along middle line, never with inverted broad Y-shaped maculation as in female), gena basally with thin band (less than half median ocellar diameter), vertex with small spot laterally, pronotum, pronotal lobe, tegula with small spot on anterior half, costal sclerite, mesepisternum laterally with spots, metepisternum almost entirely, scutum laterally with broad band (about half median ocellar diameter), axilla, lateral and distal margins of scutellum with narrower band than on scutum, apices of middle and hind femora, outer surfaces of fore and middle tibiae with narrow band, hind tibia with band on posterior margin, fore and middle basitarsi basally. Wing membrane subhyaline with weak greenish and coppery reflections; veins and stigma dark brown (Fig. 20).
Body sparsely covered with minute, appressed, simple yellowish to whitish hairs except as follows: vertex, hypostomal area, middle trochanter with long (about half median ocellar diameter), erect hairs; hind coxa, seventh tergum with much longer erect hairs (at least as long as median ocellar diameter); sides of propodeum with dense, branched, semierect hairs (about 0.5 times median ocellar diameter) partially obscuring integument.
(n = 33♀, 39♂) Costa Rica: 1♀, 1♂, Turrialba, VII-15-19-65, P.J. Spangler; 2♀, 5♂, S.J. [San José], San Isidro General, Feb. 1993, F. D. Parker; 1♀, 13♂, Alaj. [Alajuela], 20 km S Upala, collected on 1–10 April, 20–31 March, 22–31 May, 3–21 June 1991, and 1–10 August 1991, F. D. Parker; 25♀, 18♂, Alaj., Bijagua, 20 km S Upala, collected on 6–29 January, 12 February, 5–28 March, 10 April–6 May, 1–11 June, 1–15 July, 1–12 Aug, 1–15 Sep, 9–19 October, 1–20 November, and 25–30 December 1991, F. D. Parker; 4♀, Cart. [Cartago], Turribalba, CATIE, 26–29 June 1986, W. Hanson, G. Bohart ; 2♂, Guan [Guanacaste], 14 km S Cañas, collected on 17 March, 11–12 June 1991, F. D. Parker (BBSL, USMN).
Unlike the female, the yellow maculations in the male are usually reduced on gena, vertex, pronotum, frons, and sides of propodeum.
http://species-id.net/wiki/Paratrigona_impunctata
(n = 5♀, 2♂) Ecuador: 2♀, 2♂, Napo Province, Huahua Sumaco, km. 45 on Hollin-Loreto road, XII-16-20, 1989, Malaise trap, M. & J. Wasbauer. Collrs; 1♀, Misahualli nr. Tena, 3–8 Oct., 1999, Steven R. Keller; 1♀, Yasuni Res. Sta. 19–30 Oct. 1998, W.J. Hanson, 250 m / 6°36'W, 0°38'S; idem, 1♀, 30 Sept.-11 Oct. 2002, C. Brammer, 250 m M.T [Malaise trap]., 0°40.566'S, 076°23.851W (BBSL).
These new records expand the distribution of this species from its previously known range: Brazil, Colombia, French Guiana, Guyana, Peru, and Suriname (Table 2).
http://species-id.net/wiki/Paratrigona_opaca
Ecuador: 3♀, 5♂, Napo Province, Huahua Sumaco, km. 45 on Hollin-Loreto road, XII-20, 1989, Malaise trap, M. & J. Wasbauer, H. Real (BBSL).
This new record expands the range of this species from its previously known distribution: Colombia, Costa Rica, Guatemala, Mexico, and Panama (Table 2).
| 1 | Maculations absent on frons, gena (except for some small spots in some specimens of Paratrigona subnuda), mesepisternum, and metepisternum (lineata species group) | 2 |
| – | Maculations present on frons, gena, mesepisternum, and metepisternum (haeckeli-lineatifrons species group) | 9 |
| 2(1) | Antennal scape broad, about 1.3 times width of third flagellomere; submarginal angle (between Rs and Rs+M) obtuse (>100°); malar area longer than half width of third flagellomere | Paratrigona prosopiformis (Gribodo) |
| – | Antennal scape, at most, slightly broader than width of third flagellomere; submarginal angle acute (<90°); malar area narrower than half width of third flagellomere | 3 |
| 3(2) | Scutellum short-paraboloid, length about 0.55 times its width; paraocular band usually sinuous at the level of antennal alveolus | Paratrigona nuda (Schwarz) |
| – | Scutellum short-semicircular, length between 0.50 and 0.51 times its width; paraocular band not sinuous at the level of antennal alveolus | 4 |
| 4(3) | Scape narrower than width of third flagellomere | 5 |
| – | Scape about as broad as, or broader than, width of third flagellomere | 8 |
| 5(4) | Disc of scutum with distinctly long, erect, simple hairs | Paratrigona lineata (Lepeletier) |
| – | Disc of scutum without long, erect, simple hairs | 6 |
| 6(5) | Mesepisternum dorsally with long, erect, simple hairs (Fig. 9); scutellum dorsally with long (about as long as median ocellar diameter), erect, simple hairs on distal half (Fig. 10) (Paraguay) | Paratrigona wasbaueri sp. n. |
| – | Mesepisternum dorsally without long, erect, simple hairs; hairs on scutellum short (at most half median ocellar diameter) or absent | 7 |
| 7(4) | Scutellum with erect hairs on distal margin and some on dorsal surface; face with pale yellow maculations; northern Argentina to Bolivia | Paratrigona glabella Camargo & Moure |
| – | Scutellum with erect hairs on inferior surface only, absent on distal margin; face with bright yellow maculations (northern Minas Gerais, Brazil) | Paratrigona incerta Camargo & Moure |
| 8(4) | Scape distinctly broader (1.2–1.3 times) than width of third flagellomere (Pará, Amazonas, Brazil) | Paratrigona crassicornis Camargo & Moure |
| – | Scape about as broad as or slightly broader than width of third flagellomere (southeastern Brazil) | Paratrigona subnuda Moure |
| 9(1) | Scape thick, about 1.5 times width of third flagellomere; body ferruginous | 10 |
| – | Scape, at most, slightly broader than width of third flagellomere; body black, especially head and mesosoma | 11 |
| 10(9) | Scape distinctly convex on its outer margin, abruptly narrowed on basal one-fourth, inner margin straight or nearly so, with long (0.5 times width of third flagellomere), stiff, erect, simple hairs (Fig. 4) | Paratrigona scapisetosa sp. n. |
| – | Scape straight or nearly so on its outer margin, not abruptly narrowed on basal one-fourth, inner margin gently convex, without long hairs (Fig. 5) | Paratrigona haeckeli (Friese) |
| 11(9) | Hind femur with superior margin protuberant on basal third | 12 |
| – | Hind femur unmodified, without superior margin protuberant on basal third | 13 |
| 12(11) | Paraocular band broad, 1.4 times broader than width of third flagellomere, inferiorly filling the space between the compound eye and epistomal sulcus to the level of the tentorial pit | Paratrigona euxanthospila Camargo & Moure |
| – | Paraocular band narrow, about as broad as width of third flagellomere, inferiorly not filling the space between the compound eye and epistomal sulcus to the level of the tentorial pit | Paratrigona femoralis Camargo & Moure |
| 13(11) | Maculations absent or reduced on inferior paraocular area and clypeus; paraocular band present above superior tangent of antennal alveolus; frontal maculation separated in two small spots, one below median ocellus, the other in the middle of frons; mesepisternum dorsally with spot as large as that on pronotal lobe; malar area short, slightly longer than half width of third flagellomere | Paratrigona melanaspis Camargo & Moure |
| – | Maculations of face not reduced on clypeus and supraclypeus; paraocular band reaching inferior tangent of antennal alveolus; frontal maculation usually complete; mesepisternum dorsally with or without spot; malar area variable in length | 14 |
| 14(13) | Antennal scape robust, slightly broader than width of third flagellomere, black or with small basal spot; clypeus with reduced maculations, at most yellow along middle line; paraocular band narrower than width of third flagellomere, not reaching epistomal sulcus; mesepisternum black or with small spot; malar area short, slightly shorter than width of third flagellomere | Paratrigona onorei Camargo & Moure |
| – | Antennal scape slender, at most as broad as width of third flagellomere, maculate on anterior face; clypeus with well developed maculations, not restricted to middle line; paraocular band of variable width, reaching epistomal sulcus; mesepisternum with large spot; malar area variable in length | 15 |
| 15(14) | Paraocular band narrow, slightly broader than half width of third flagellomere, about the same width along entire length; genal band narrow, not reaching vertex | Paratrigona lineatifrons (Schwarz) |
| – | Paraocular band broad, 1.4 times broader than width of third flagellomere wider in the middle or below antennal alveolus; genal band complete, reaching vertex and joining paraocular band or slightly interrupted | 16 |
| 16(15) | Paraocular band very broad inferiorly, occupying space between compound eye and epistomal sulcus below tentorial pit | Paratrigona ornaticeps (Schwarz) |
| – | Paraocular band narrowed inferiorly, not occupying space between compound eye and epistomal sulcus below tentorial pit | 17 |
| 17(16) | Malar area about as long as, or slightly longer than, width of third flagellomere; frontal maculation widest in the middle, just above bifurcation; clypeus yellow along distal margin (Andes, Colombia) | Paratrigona uwa Gonzalez & Vélez |
| – | Malar area length about 2/3 width of third flagellomere; frontal maculation not distinctly broad above bifurcation; clypeus entirely yellow, at most, darkened on disc (Amazonas, Brazil) | Paratrigona compsa Camargo & Moure |
We are indebted to B. Harris and S. Brady (USNM) for kindly arranging the loan of Paratrigona specimens and permitting its study; Amy Comfort de Gonzalez, Claus Rasmussen, Rita Vélez, Michael Engel, Mariano Lucia, and anonymous reviewers for comments and suggestions that improved this note. This study was supported in part by National Science Foundation grant DEB-0742998.