





(C) 2011 M. A. Alonso-Zarazaga. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Pseudaspidapion botanicum sp. n. from China is described and figured. Its host plant is Grewia biloba G. Don var. parviflora (Bunge) Hand.-Mazz (Malvaceae: Grewioideae). The genus Harpapion Voss, 1966 is recorded as new for China and Vietnam and two comb. n. are proposed: Harpapion vietnamense (Korotyaev, 1985) (from Aspidapion) and Harpapion coelebs (Korotyaev, 1987) (from Pseudaspidapion). A key to the known species of the genus Pseudaspidapion from China is presented.
Weevils, Pseudaspidapion botanicum, Harpapion , Beijing, Shaanxi, new species, new combination, morphology, systematics, biology, key
M.
In 2008, we collected a series of specimens of a Pseudaspidapion. Further specimens were later found. We consider them to represent a species new to science that we describe below.
In addition to the description of the adult characters, we also provide some biological data for the new species after one year of survey in the Beijing area.
Material and methodsMaterials examined of new species for this study are to be deposited in the Institute of Zoology, Chinese Academy of Sciences, Beijing (IZCAS), the Museo Nacional de Ciencias Naturales, Madrid (MNCN), the Zoological Institute of Russian Academy of Sciences, Moscow (ZIN), the Museum of Natural History, University of Wrocław (MNHW) and the Beijing Botanical Garden, Beijing (BBG).
Type specimens were obtained from ZIN on loan or belong to IZCAS and their data are summarised in Table 1. Information about their condition and taxonomy is given in the Discussion section.
List of studied type specimens and depositories.
| Species | Depository | Examined type material |
|---|---|---|
| Aspidapion inarmatum Korotyaev, 1985 | IZCAS | 2 paratypes |
| Aspidapion panfilovi Korotyaev, 1985 | IZCAS | Holotype and 1 paratype |
| Pseudaspidapion coelebs (Korotyaev, 1987) | ZIN | 1 paratype |
| Pseudaspidapion kryzhanovskii (Korotyaev, 1987) | ZIN | 1 paratype |
| Pseudaspidapion medvedevi (Korotyaev, 1985) | ZIN | Holotype |
| Pseudaspidapion topali (Korotyaev, 1985) | ZIN | 1 paratype |
| Pseudaspidapion vietnamense (Korotyaev, 1987) | ZIN | Holotype |
| Pseudaspidapion yunnanicum (Korotyaev, 1985) | IZCAS | Holotype and 3 paratypes |
| Pseudaspidapion zagulajevi (Korotyaev, 1987) | ZIN | 2 paratypes |
Descriptions were made and photographs were taken with a CCD Qimagine MicroPublisher 5.0 RTV mounted on a Zeiss SteREO Discovery V.12. Extended focus images were generated with Auto-Montage Pro 5.03.0061 and edited with Adobe Photoshop CS 5.0 if required. Microscopic slides were studied under a Leica DM 2500 microscope and photos were taken with a Nikon CoolPix 5400. The map was made with the software ArcGIS 9.3. Drawings were made from the original photographs by using the software Adobe Illustrator CS5.0, or directly by using a drawing tube linked to the microscope.
Nomenclature of the rostral parts follows
The dissecting method used follows
After description, the genitalia and other parts of each
specimen were placed in DMHF on a plastic card for long term
conservation (
Labels are described as they are (in Chinese), with pinyin romanization or comments in square brackets; labels are separated by semicolons and lines by slashes.
Taxonomic treatmenturn:lsid:zoobank.org:act:88656E61-918F-4F1B-A3BD-6A3FA714A237
http://species-id.net/wiki/Pseudaspidapion_botanicum
Figs 1–28 (from paratypes)This new species resembles Pseudaspidapion yunnanicum, but it can be distinguished from the latter by the characters in the table 2.
(male holotype, except where indicated). Measurements (in mm): Standard length: 1.82. Rostrum: length: 0.71, maximum width: 0.17. Pronotum: median length: 0.49, maximum width: 0.59. Elytra: median length: 1.38, maximum width: 1.08.
With the general characters of genus Pseudaspidapion as described in
Integument. Generally piceous black (Figs 1–2).
Vestiture composed of thick, longer white piliform scales with truncate apex and thin, shorter brownish to greyish acute hairs; scales present on coxae, femora and tibiae, sides of pronotum, meso- and metaventrite, mesoventral process, base of 3rd elytral interstria, metarostrum, and weak subocular patch, sometimes intermediate types present. Antennal scape apex and pedicel with sparse short greyish hairs, club and desmomeres 2–7 covered with dark hairs only. Pronotal vestiture centrifugal, scales both on apex and base perpendicular to the margins, hairs on disc distinctly longer than those on elytra, reaching middle of preceding hair. Elytral vestiture white at base and sides, brownish on disc, interstrial scales large, longer than 2/3 of the interstrial width, in two rather regular rows per interstria, one specialized seta on apical region of 9th interstria.
Rostrum cylindrical and moderately robust, in dorsal view 4.18× as long as maximum width, 1.45× as long as pronotum in midline, widest at mesorostrum, pro- and metarostrum with sides almost parallel, metarostrum with two very fine, punctulate dorsal submedial sulci shortened at mesorostral level, and two weak dorsal sublateral sulci, dorsal submedial sulci prolonged on frons, separated, dorsal sublateral sulci prolonged close to ocular margins, metarostrum microreticulate, matt, pro- and mesorostrum smooth, shining, sparsely punctulate; in lateral view weakly and evenly curved, ventral margin forming a weak angle at mesorostrum, each side with a very thin low dorsal sublateral keel running from front margin of eye to upper margin of scrobe and beyond, limiting ventrally the dorsal sublateral sulcus, prorostrum with a marked ventral sublateral sulcus under this keel.
Head transverse, frons weakly convex with four rows of relatively deep punctures running from metarostrum and hardly surpassing hind margin of eyes, occiput almost reaching hind eye level, subocular keel reaching middle of eyes, the area between subocular keels microreticulate and impunctate, with a low, fine median keel. Eyes round, moderately convex.
Antennae inserted at basal 0.26 of rostral length, scape 5.00× as long as wide, as long as club, 1.18× as long as mesorostral width. Pedicel 2.14× as long as wide, as long as desmomeres 2+3, desmomere 2 1.67×as long as wide, desmomeres 3–5 1.33× as long as wide, desmomere 6 1.14× as long as wide, desmomere 7 1.25× as long as wide, shortly obconical. Club oblong, compact, 2.5× as long as wide, as long as the last 5.5 desmomeres, sutures well marked (Fig 7).
Pronotum campaniform, transverse, 0.83× as long as wide, widest just behind middle, constrictions weak, sides weakly dilated at middle, base 1.31× as wide as apex, bisinuate with moderate medial rounded projection towards scutellum, basal flange moderately developed. Prescutellar fovea shallow, as broad as one puncture’s diameter, as long as 2–3 diameters, prolonged in a very fine sulcus reaching middle of pronotum. Discal punctures relatively deep, ca. 0.5–1× diameter apart, interspaces slightly convex, microreticulate.
Scutellum large, elongated, triangular, 1.63× as long as wide, with two basal tubercles separated by a weak median depression, in lateral view subacutely prominent, in front view tubercles fused basally; apex constricted and moderately raised, distinctly visible in lateral view.
Elytra 1.23× as long as wide, 2.82× as long as pronotum, widest almost at the middle, humeri distinct; striae deep, about as wide as interstriae at base and apex, about half as wide at elytral disc, distinctly catenulate-punctate, punctures round to oblong, space between punctures about 2.00× as long as puncture length, striae apically connected 1+2+9, 3+4, 5+6, 7+8. Interstriae evidently convex with small punctures, surface distinctly wrinkled, not microreticulate, shining. Macropterous (wing of paratype, Fig 8).
Ventral areas. Mesocoxae and metacoxae narrowly separated by a distance of 0.26× and 0.38× of their transverse diameter, respectively. Metaventrite ca. as long as mesocoxae. Mesoventral process more prominent than metaventral process. Anterior metaventral rim present, weak. Abdominal ventrites microreticulate, with length ratios along midline: 31-17-6-7-16. Ventrites 1–2 coarsely and densely punctate, 3–4 and base of 5 very sparsely and minutely punctate, apical half of 5 densely punctate with a median convexity. Suture I distinct, distance from hind margin of metacoxae, as long as ventrite 2. Ventrite 5 subsemicircular, transverse, 0.41× as long as wide.
Pygidium (from paratype) semicircular, 0.69× as long as wide, apical edge of arc with a flat and glabrous side, punctate medially with interspace microreticulate (Figs 9–10).
Legs. Metafemora a little more robust than pro- and mesofemora, profemur 2.15× as long as wide, widest at middle, minutely punctate. Protibia almost straight, 6.50× as long as wide (Fig. 5). Protarsomere 1 2.29× as long as wide, tarsomere 2 1.22× as long as wide, tarsomere 3 bilobed, 0.8× as long as wide, lobes narrow, not much dilated outwards, onychium 3.25× as long as wide, projecting from lobes of tarsomere 3 for 0.54× its length. Meso- and metatibiae similarly mucronate, mucro subconical (Fig. 6), about 0.50× as long as tibial apical width. Tarsal claws with conspicuous obtuse basal tooth.
Genitalia and terminalia (from topotypic paratypes). Ninth sternite (spiculum gastrale) Y-shaped and slightly winged near arm base, manubrium ca. 2.00× as long as arms (Fig. 15). Penis depressed, moderately curved, pedon with apical plate straight in side view, truncate and slightly constricted apicad in side view; temones about 0.50× as long as pedon; endophallus without large structures (Figs 13–14). Tegminal plate articulated with free ring, laterally developed, enveloping, with parameroid lobes not notched apically, membranous area rhombic in general outline, microsetose apically, reaching middle of parameroid lobes, basal sclerotized area large, each side with 5 short macrochaetae, 2 latero-apical and 3 latero-medial, several sensilla distributed on middle part and posterior margin; fenestrae short, tranverse, narrowly separated; linea arquata visible; prostegium bidentate, apex of projection rounded; median unsclerotized strip moderately elongate, reaching hind margin of fenestrae; manubrium with apex not broadened (Figs 11–12).

Pseudaspidapion botanicum Alonso-Zarazaga & Wang, sp. n. 1 male paratype, dorsal view 2 male paratype, lateral view 3 female paratype, head and rostrum, dorsal view 4 female paratype, head and rostrum, lateral view. Scales: 1–2: 1000 μm; 3–4: 500 μm.
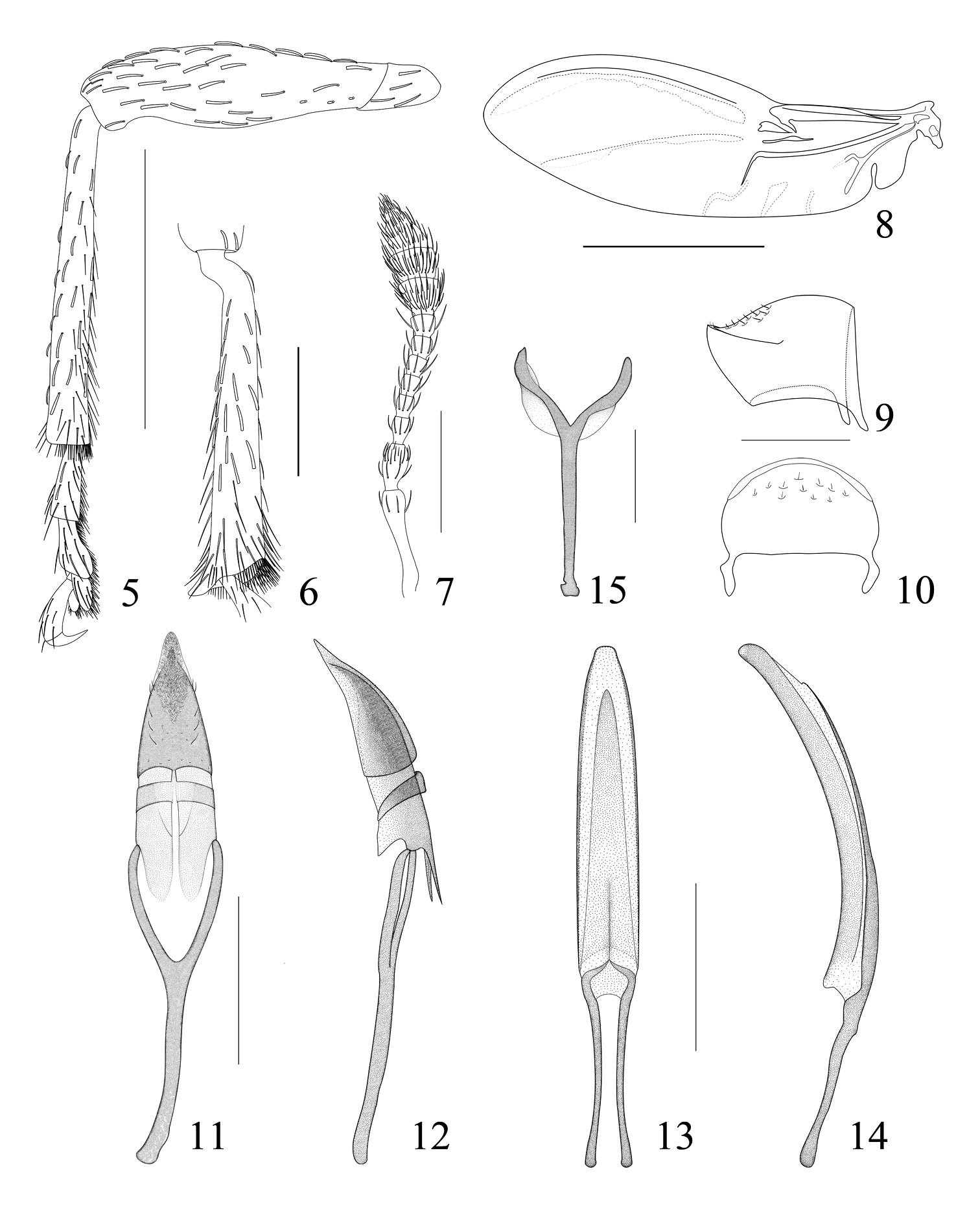
Pseudaspidapion botanicum Alonso-Zarazaga & Wang, sp. n., male paratype 5 front leg 6 hind tibia 7 antenna 8 hind wing 9 pygidium, lateral view 10 pygidium, dorsal view 11 tegmen, dorsal view 12 tegmen, lateral view 13 penis, dorsal view 14 penis, lateral view 15 spiculum gastrale. Scales: 5: 500 μm; 6–7, 9–15: 200 μm; 8: 1000 μm.

Pseudaspidapion botanicum Alonso-Zarazaga & Wang sp. n. 16 female resting on underside of leaf 17 female boring a bud for oviposition, with guarding male.

Types of Pseudaspidapion, Aspidapion and Harpapion, in lateral and dorsal views 18–19 Aspidapion panfilovi Korotyaev, 1985, female holotype 20–21 Pseudaspidapion zagulajevi (Korotyaev, 1987), female paratype 22–23 Pseudaspidapion kryzhanovskii (Korotyaev, 1987), female paratype 24–25 Pseudaspidapion yunnanicum (Korotyaev, 1985), female holotype 26–27 Pseudaspidapion topali (Korotyaev, 1985), male paratype 28–29 Pseudaspidapion medvedevi (Korotyaev, 1985), male holotype 30–31 Harpapion coelebs (Korotyaev, 1987), male paratype 32–33 Harpapion vietnamense (Korotyaev, 1987), male holotype. Scales: 18–33: 1000 μm.
Male paratypes. Measurements (in mm) (n=10): Standard length: 1.38–2.05 (mean= 1.735). Rostrum: length: 0.52–0.82 (mean= 0.722), maximum width: 0.13–0.19 (mean= 0.164). Pronotum: median length: 0.36–0.53 (mean= 0.479), maximum width: 0.44–0.68 (mean= 0.594). Elytra: median length: 1.06–1.54 (mean= 1.406), maximum width: 0.78–1.18 (mean= 1.038).
Rostrum 1.44–1.58× as long as pronotum, 4.00–4.63× as long as wide. Ventrite 5 sometimes slightly subtriangular. Otherwise as in holotype.
Female paratypes (Figs 3–4). Measurements (in mm) (n=10): Standard length: 1.56–2.05 (mean= 1.831). Rostrum: length: 0.52–0.76 (mean= 0.669), maximum width: 0.130–0.155 (mean= 0.143). Pronotum: median length: 0.37–0.53 (mean= 0.460), maximum width: 0.48–0.65 (mean= 0.571). Elytra: median length: 1.24–1.62 (mean= 1.444), maximum width: 0.88–1.14 (mean= 1.032).
Sexual dimorphism rather weak in this species, females differ from males by a weakly thinner and more evenly curved rostrum with a brighter and less sculptured metarostrum, and all the tibiae without mucros. Rostrum 1.35–1.53× as long as pronotum, 4.00–5.07× as long as wide. Antennae inserted at ca. basal 0.27 of rostrum. Ventrite 5 almost flat, hardly punctured. Otherwise practically as in male.
Holotype: ♂: (white, printed): 北京植物园樱桃沟 [Běijīng zhíwùyuán Yīngtáogōu] / 2008.VI.1 / Leg. 王志良 [Wáng Zhìliáng]; (white, printed): IOZ(E)1638667, deposited in IZCAS. This is the Beijing Botanical Garden at Yingtaogou (40°0'51.41"N, 116°12'14.34"E), Haidian district, Beijing (P. R. of China). Paratypes (132♂137♀): 4♂7♀: (white, printed and handwritten): 北京三堡 [Běijīng Sānpù]; (white, printed and handwritten): 1964.VII.20 / leg. 马文珍 [Mă Wénzhēn]; (white, printed): IOZ(E)1638668-1638678; 3♀: (white, printed and handwritten): 北京三堡 [Běijīng Sānpù] / 600m; (white, printed and handwritten): 1964.VIII.21 / leg. 李铁生 [Lí Tiéshēn]; (white, printed): IOZ(E)1638679-1638681; 1♂1♀: (white, printed and handwritten): 北京三堡 [Běijīng, Sānpù] / 1979.VIII.8; (white, handwritten): leg. 廖素柏 [Liào Sùbăi]; (white, printed): IOZ(E)1638682-1638683; 5♂5♀: (white, printed and handwritten): 北京上方山 [Běijīng Shàngfāngshān]; (white, printed and handwritten): 1980.VIII.2 / leg. 廖素柏 [Liào Sùbăi]; (white, printed) IOZ(E)1638684-1638693; 2♂: (white, printed and handwritten): 北京上方山 [Běijīng Shàngfāngshān]; (white, printed and handwritten): 1979.VII.25 / leg. 陈元清 [Chén Yuánqīng]; (white, printed): IOZ(E)1638694-1638695; 2♂2♀: (white, printed and handwritten): 北京居庸关 [Běijīng Jūyōngguān] / 500m; (white, printed and handwritten): 1964.VIII.20 / leg. 李铁生 [Lí Tiéshēn]; (white, printed): IOZ(E)1638696-1638699; 1♂: (white, printed and handwritten): 北京八达岭 [Běijīng Bādálīng] / 700m; (white, printed and handwritten): 1963.VII.25 / leg. 李铁生 [Lí Tiéshēn]; (white, printed): IOZ(E)1638700; 8♂8♀: (white, printed): 北京香山 [Běijīng Xiāngshān]; (white, printed and handwritten): 1963.V.30 / leg. 李铁生 [Lí Tiéshēn]; (white, printed): IOZ(E)1638701-1638716; 1♀: (white, printed and handwritten): 北京香山 [Běijīng Xiāngshān]; (white, printed and handwritten): 1964.V.5 / leg. 马文珍 [Mă Wénzhēn]; (white, printed): IOZ(E)1638717; 1♀: (white, handwritten): 北京香山 [Běijīng Xiāngshān] / 1957.VIII.16; (white, printed): IOZ(E)1638718; 1♂: (white, printed and handwritten): 北京卧佛寺 [Běijīng Wòfósì] / 50m; (white, printed and handwritten): 1962.VI.25 / leg. 王春光 [Wáng Chūnguāng]; (white, printed): IOZ(E)1638719; 1♀: (white, printed and handwritten): 北京卧佛寺 [Běijīng Wòfósì]; (white, printed and handwritten): 1963.IX.3 / leg. 姜胜巧 [Jiāng Shèngqiáo]; (white, printed): IOZ(E)1638720; 1♀: (white, printed and handwritten): 北京卧佛寺 [Běijīng Wòfósì] / 50m; (white, printed and handwritten): 1962.VIII.31 / leg. 谢汝忠 [Xieruzhong]; (white, printed): IOZ(E)1638721; 1♂: (white, printed and handwritten): 北京潭柘寺 [Běijīng Tánzhèsì] / 1975.VII.24; (white, printed and handwritten): leg. 王书永 [Wang Shuyong]; (white, printed): IOZ(E)1638722; 1♀: (white, printed and handwritten): 1964.VII.20 / leg. 马文珍 [Mă Wénzhēn]; (white, printed): IOZ(E)1638723; 2♂3♀: (white, printed and handwritten): 北京圆明园 [Běijīng Yuánmíngyuán] / 1980. VII.7 / leg. 姜胜巧 [Jiāng Shèngqiáo]; (white, printed): IOZ(E)1638724-1638728; 2♂10♀: (white, printed): 北京门头沟军庄西杨坨村 [Běijīng Méntóugōu Jūnzhuāng Xīyángtún] / 2008.VII.27 / leg. 王志良 [Wáng Zhìliáng]; (white, printed): 寄主: 小花扁担木 [Jìzhū: Xiăohuābiăndànmù] / Grewia biloba G. Don var. parviflora (Bunge) Hand.-Mazz; (white, printed): IOZ(E)1638729-1638740; 5♂8♀: (white, printed): 北京怀柔九渡河镇怀九河 [Běijīng Huáiróu Jiūdùhézhèn Huáijiūhé] / 2008.VI.15 / leg. 王志良 [Wáng Zhìliáng]; (white, printed): IOZ(E)1638741-1638753; 1♀: (white, printed): 北京怀柔三渡河 [Běijīng Huáiróu Sāndùhé] / 2008.V.24 / leg. 王志良 [Wáng Zhìliáng]; (white, printed) IOZ(E)1638754; 46♂24♀: (white, printed): 北京香山植物园樱桃沟 [Běijīng Xiāngshān Zhíwùyuán Yīngtáogōu] / 2008.VIII.19 / leg. 王志良 [Wáng Zhìliáng]; (white, printed): IOZ(E)1638755-1638770, IOZ(E)1638784-1638790, IOZ(E)1638804-1638850; 28♂41♀: (white, printed): 北京植物园樱桃沟 [Běijīng zhíwùyuán Yīngtáogōu] / 寄主:小花扁担木 [Jìzhū: Xiăohuābiăndànmù] / 2008.VI.1 / Leg. 王志良 [Wáng Zhìliáng]; (white, printed): IOZ(E)1638851-1638900, IOZ(E)1638911, IOZ(E)1638928-1638945; 7♂7♀: (white, printed): 北京海淀百望山 [Běijīng Hăidiàn Băiwàngshān] / 2009.VI.21 / leg. 杨干燕 [Yáng Gànyàn]; (white, printed): IOZ(E)1639912-1638918, IOZ(E)16398920-1638926; 2♂1♀: (white, printed and handwritten): 陕西华阴县孟塬 [Shaănxi Huáyīn Mèngyuán] / 450m / 1972.VIII.9; (white, printed): IOZ(E)1639562-1639563, IOZ(E)1639540, deposited in IZCAS.
10♂10♀: (white, printed): 北京香山植物园樱桃沟 [Běijīng Xiāngshān Zhíwùyuán Yīngtáogōu] / 2008.VIII.19 / leg. 王志良 [Wáng Zhìliáng]; (white, printed): IOZ(E)1638781, IOZ(E)1638801, IOZ(E)1638771-1638779, IOZ(E)1638791-1638799, to be deposited in MNCN.
1♂1♀: (white, printed): 北京香山植物园樱桃沟 [Běijīng Xiāngshān Zhíwùyuán Yīngtáogōu] / 2008.VIII.19 / leg. 王志良 [Wáng Zhìliáng]; (white, printed): IOZ(E)1638780, IOZ(E)1638800, to be deposited in BBG.
1♂1♀: (white, printed): 北京香山植物园樱桃沟 [Běijīng Xiāngshān Zhíwùyuán Yīngtáogōu] / 2008.VIII.19 / leg. 王志良 [Wáng Zhìliáng]; (white, printed): IOZ(E)1638783, IOZ(E)1638803, to be deposited in ZINM.
1♂1♀: (white, printed): 北京香山植物园樱桃沟 [Běijīng Xiāngshān Zhíwùyuán Yīngtáogōu] / 2008.VIII.19 / leg. 王志良 [Wáng Zhìliáng]; (white, printed): IOZ(E)1638782, IOZ(E)1638802, to be deposited in MNHW.
The male holotype has not been dissected to avoid any damage, since there are many specimens collected with it.
The new species is named after the first locality where it was found during a collecting visit: the Beijing Botanical Garden. It is a Latin adjective.
The species is known for the moment only from the municipality of Beijing and the province of Shaanxi.
Pseudaspidapion botanicum sp. n. was collected from Grewia biloba G. Don var. parviflora (Bunge) Hand.-Mazz (Malvaceae: Grewioideae) (
During this research, we were able to study types of several of the species placed in the genera Pseudaspidapion and Aspidapion in China in the recent Palaearctic catalogue (
The systematic placement of Aspidapion inarmatum is unclear at the moment, but it belongs neither to Aspidapion nor to Pseudaspidapion or Harpapion.
Aspidapion panfilovi, known from females until now, possesses several coincident characters with some Pseudaspidapion: its more elongate and apparently glabrous elytra, the shallow striae with interstriae about twice as broad as striae on the disc of the elytra, etc. Although it resembles the female of Pseudaspidapion zagulajevi very much, we think it is better to postpone any decision on its placement until the male is discovered. This highlights the difficulty of placing in their correct genera the species of Apionidae described only from females, a practice that should be avoided, as that of designating females as name bearing types.
Previously, the genus Pseudaspidapion was not known to exceed the Tropic of Cancer. However, the new species Pseudaspidapion botanicum reaches 40ºN latitude, an area clearly belonging to the Eastern Palaearctic region (Fig. 34).
Thus this genus shows a broader distributional range extending from the
Afrotropical across the Oriental to the Palaearctic region. On the
other hand, we identified in the collection of IZCAS several specimens
of Aspidapion validum (Germar, 1817), from Altay prefecture in northern Xinjiang (China). Apart from the species Aspidapion panfilovi,
whose placement may need a revision as mentioned above, these
specimens represent the easternmost distribution record of the otherwise
Western Palaearctic genus Aspidapion. Therefore, the distribution pattern of Pseudaspidapion is mainly Palaeotropical but has expanded to Philippines (
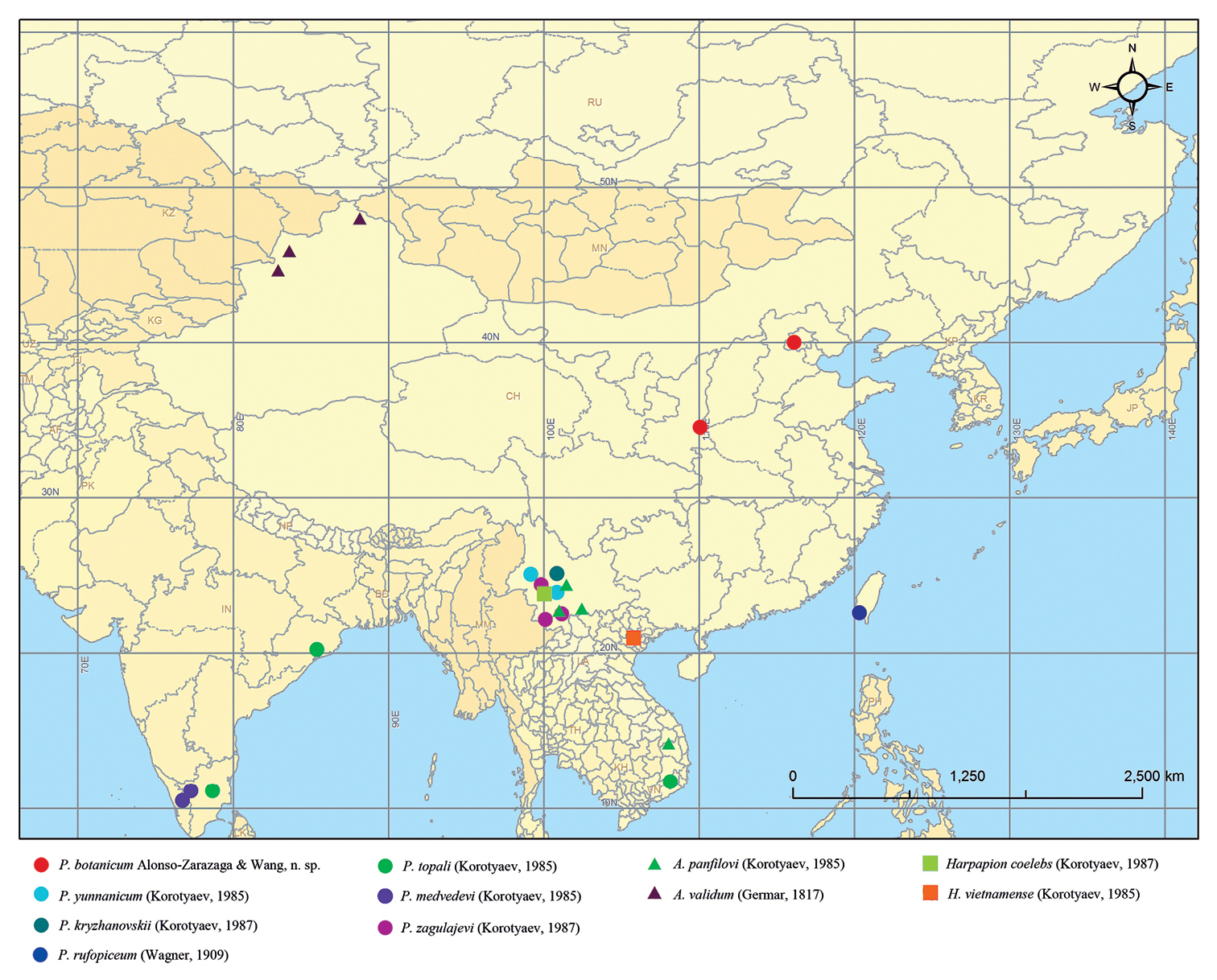
Distribution of the species of Pseudaspidapion, Harpapion and Aspidapion mentioned in the text.
Some of these species are described only from one sex, which makes it difficult to key all of the Chinese species. However, we have tried to provide a key to the known Chinese species (see below). Consequently, more field work is needed to gather specimens of both sexes, and complete our knowledge. We have not studied specimens of Pseudaspidapion rufopiceum (Wagner, 1909), a related species, which is included below from the data in its original description.
Key to the Chinese species of PseudaspidapionThis key includes also Aspidapion panfilovi Korotyaev, 1985 (only known from female), which seems to be related to these species. Body length excludes rostrum.
| 1 | Meso- and metatibiae mucronate. (males) | 2 |
| – | Meso- and metatibiae unarmed. (females) | 4 |
| 2 | Body length 3.1–3.6 mm. Elytral interstrial scales minute, shorter than 1/3 width of interestria, subtransparent, elytra appearing glabrous | Pseudaspidapion zagulajevi |
| – | Body length 1.3-2.2 mm. Elytral interstrial scales larger, longer than 2/3 of the interstrial width, white or dark brown, elytra appearing pubescent | 3 |
| 3 | Body length in general smaller: 1.3–1.6 mm. Body dark reddish-brown, appendages light reddish-brown. Pedicel little longer than wide | Pseudaspidapion rufopiceum |
| – | Body length in general larger: 1.5–2.2 mm. Body and appendages dark brown to piceous black. Pedicel at least 2 × as long as wide | Pseudaspidapion yunnanicum / Pseudaspidapion botanicum sp. n. (see Table 2 for separation) |
| 4 | Body length more than 3.7 mm. Rostrum ca. 2.5 × as long as pronotum. Elytra short, rounded, in side view strongly convex. Scutellum large, base about as wide as interocular distance, triangular, sides straight; in side view completely prominent above elytral outline | Pseudaspidapion kryzhanovskii |
| – | Body length less than 3.7 mm. Elytra oblong-eliptical, in side view moderately convex. Scutellum small, base at most 2/3 as wide as interocular distance, sides concave, forming a basal subpentagonal area prolonged apically in a shaft; in side view, at most some of the tubercles prominent above elytral outline | 5 |
| 5 | Body length more than 2.5 mm. Elytral interstrial scales minute, shorter than 1/3 width of interestria, subtransparent, elytra appearing glabrous | 6 |
| – | Body length less than 2.5 mm.Elytral interstrial scales larger, longer than 2/3 width of interstria, white or dark brown, elytra appearing pubescent | 3 |
| 6 | Rostrum ca. 1.8 × as long as pronotum, robust, mesorostrum wider than interocular distance. Basal tubercles of scutellum large, acutely prominent in side view. Body length: 3.1–3.6 mm | Pseudaspidapion zagulajevi |
| – | Rostrum ca. 2.0 × as long as pronotum, fine, mesorostrum at most as wide as interocular distance. Basal tubercles of scutellum small, hardly visible in side view. Body length: 2.7 mm | Aspidapion panfilovi |
Comparative diagnostic characters for Pseudaspidapion botanicum sp. n. and Pseudaspidapion yunnanicum.
| Pseudaspidapion yunnanicum | Pseudaspidapion botanicum sp. n. | |
|---|---|---|
| Scales on pronotal disc | Finer, sparser, shorter, tips of posterior scales not or hardly reaching the base of anterior ones | Thicker, denser and longer, tips of posterior scales surpassing the anterior by almost the basal half |
| Scape | Slender, ca. 0.26 mm long, 7.20–7.60× as long as wide, 1.66–1.85× as long as mesorostral width | Robust, ca. 0.20 mm long, 5.00–5.80× as long as wide, 1.24–1.67× as long as mesorostral width |
| Rostrum (♀) | Longer, 1.91–2.25× as long as pronotum, thinner, 7.35–10.30× as long as maximum width | Shorter, 1.35–1.53× as long as pronotum, more robust, 4.00–5.07× as long as maximum width |
| Metarostrum (dorsal view) | Microreticulate | Nearly smooth |
| Pronotum | Strong subapical constriction and sides moderately rounded behind it | Weaker subapical constriction, sides weakly rounded behind it |
| Prescutellar fovea | Deep, distinct, lanceolate, not prolonged apicad | Shallow, indistinct, sublinear, prolonged apicad to middle |
| Scutellum | Two basal tubercles separated medially by a distinctly deep notch, in anterior view tubercles not fused basally | Two basal tubercles separated medially by a weak notch, in anterior view tubercles basally fused |
We thank Marek Wanat for kindly providing us references and many good advices, Boris A. Korotyaev for lending paratypes, Steven R. Davis for doing the linguistic review before submission, Yang Ganyan for collecting specimens, Zhou Dakang for providing information about the host plants, Yin Ziwei and Nico M. Franz for providing the brush-pencil Plug-ins of Adobe Illustrator. We also thank two anonymous reviewers whose suggestions considerably improved our work. This research has been supported by an Invited Professor Award (2009) of the Chinese Academy of Sciences to the senior author and by project grant CGL2010–15786 (Ministerio de Ciencia e Innovación, Spain).