





(C) 2011 Yuri M. Marusik. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Three species of Xerolycosa: Xerolycosa nemoralis (Westring, 1861), Xerolycosa miniata (C.L. Koch, 1834) and Xerolycosa mongolica (Schenkel, 1963), occurring in the Palaearctic Region are surveyed, illustrated and redescribed. Arctosa mongolica Schenkel, 1963 is removed from synonymy with Xerolycosa nemoralis and transferred to Xerolycosa, and the new combination Xerolycosa mongolica (Schenkel, 1963) comb. n. is established. One new synonymy, Xerolycosa undulata Chen, Song et Kim, 1998 syn. n. from Heilongjiang = Xerolycosa mongolica (Schenkel, 1963), is proposed. In addition, one more new combination is established, Trochosa pelengena (Roewer, 1960) comb. n., ex Xerolycosa.
Wolf spider, Asia, new combination, new synonymy
This paper is the first in a series of reviews of the Palaearctic Evippinae
Specimens were photographed using either a JEOL JSM-5200 scanning electron microscope or an Olympus Camedia E-520 camera attached to an Olympus SZX16 stereomicroscope at the Zoological Museum, University of Turku. Digital images were montaged using a “CombineZM” image stacking software. Photographs were taken in dishes of different sizes with paraffin at the bottom. Different sized holes were made in the bottom to keep the specimens in the correct position. Figures had been made previously and in some cases we were unable to generate scale bars for the digital photographs. All measurements are given in mm. Drawings we made either by using a grid method with a MBS-9 stereomicroscope or a Leitz stereomicroscope with a camera lucida. The bleached epigyne of the holotype female was temporarily coloured with Chlorazol Black. Epigynes were macerated using KOH solution. In the tables of leg spination, apical and dorsal spine data are omitted.
Terminology of the copulatory organs follows (
Abbreviations used in the text: AME, ALE, PME, PLE – anterior median, anterior lateral, posterior median and posterior lateral eyes respectively; pv – proventral; rv – retroventral; v – ventral; p – prolateral; r – retrolateral.
Acronyms:
IBPN Institute for Biological Problems of the North, Magadan, Russia
MNHN Muséum National d'Histoire Naturelle, Paris, France
SMFM Senckenberg Museum, Frankfurt am Main, Germany
SZMN Siberian Zoological Museum RAS, Novosibirsk, Russia
TNU Zoology Department, Taurida National University, Simferopol, Ukraine
ZISP Zoological Institute, St.-Petersburg, Russia
ZMMU Zoological Museum of the Moscow State University, Russia
ZMUT Zoological Museum, University of Turku, Finland
Taxonomic surveyhttp://species-id.net/wiki/Xerolycosa
Members of this genus can be easily separated from otherEvippinae genera by the fewer number of ventral tibial spines on leg I (3pv & 2rv, or 2–2v), carapace lacking transverse depression (present in Evippa Simon, 1882) and lack of pseudo-articulation of tarsi (Fig. 13). Xerolycosa can be differentiated by the shape of their copulatory organs. Females have a short droplet-shaped septum (about as long as wide), while in Evippa the septum is long and has a well developed septal stem. The male palp in Xerolycosa has a shorter course of the seminal duct and a shorter embolus, which is only partly hidden by the tegulum.
Medium-sized (5.5–7.5) dark coloured or spotty lycosids. Carapace spotty or dark coloured with lighter median band and two lateral stripes. Cephalic region not elevated. Chelicerae with 3 promarginal and 2 retromarginal teeth. Inner side of chelicerae with a kind of stridulatory file (Fig. 12). Femora with 3 dorsal spines, tibia and metatarsus with 2 dorsal spines, sometimes poorly developed, tibia and metatarsi with four or five ventral spines (3pv-2rv or 2–2v). Tarsi without transverse furrow.
Male palp: cymbium with several apical spines; tegular apophysis shifted retrolaterally, with bill-like extension directed ventrally. Palea absent, embolus forming almost a circle, only partly hidden by tegulum. Epigyne: fovea (depression) absent, septum droplet-shaped, covered with hairs, almost as wide as high; stem short. Weakly sclerotized parts of epigyne are referred to here as windows (Wi).
Saitocosa was synonymised with Xerolycosa by
In Platnick's catalogue (
Because of the burrowing behaviour in Xerolycosa mongolica
(Schenkel, 1963), believed to be absent in the other species, we
first followed A.A. Zyuzin's (personal communication) opinion that it
may belong to a separate genus. However, females of Xerolycosa nemoralis are known to excavate shallow depressions in soil (
Xerolycosa species can be distinguished by the shape of the copulatory organs. In addition Xerolycosa mongolica can be recognized by the variegated (spotty) pattern of the carapace and abdomen, and by lacking a light median band. The spination of leg I may help to distinguish males of Xerolycosa mongolica, and females of all species.
The male palps in all three species are rather similar in general appearance. The species can be relatively easy recognized in retrolateral view by the profile of the tegular apophysis (Figs 22–24, 25, 27, 29) and by the shape of the embolic region following dissection, notably the course and length of the embolus, and the seminal duct position (Figs 26, 28, 30). The males of Xerolycosa miniata and Xerolycosa nemoralis have the same spination pattern on leg I (Table 1), but the females have different leg spine formulae (Table 2). The epigynes in the three species are very similar and can be distinguished by the shape of the septum and the “windows” (Figs 31, 33, 35, 37, 39, 41). Additional differences can be found in the spermathecae (Figs 32, 36, 40, 34, 38, 42).
Chaetotaxy of leg I in Xerolycosa males.
| Species | Segments of leg I | |||
|---|---|---|---|---|
| femur | patella | tibia | metatarsus | |
| miniata | 2p+2r | 1p+1r | 1p+2r+3-2v | 2p+1r+2-2v |
| mongolica | 1 or 2p+2r | 1p | 0 or 1p+2-2v | 1p+2-2v |
| nemoralis | 2p+2r | 1p+1r | 1p+2r+3-2v | 2p+1r+2-2v |
Chaetotaxy of leg I in Xerolycosa females
| Species | Segments of leg I | |||
|---|---|---|---|---|
| femur | patella | tibia | metatarsus | |
| miniata | 2p | 0 | 1p+2-2v | 2p+2-2v |
| mongolica | 2p | 0 | 0 or 1p+3-2v | 2p+2-2v |
| nemoralis | 2p+2r | 1p | 1p+3-2v | 2p+2-2v |
| 1 | Carapace with wide whitish median band | 2 |
| – | Carapace without whitish median band | Xerolycosa mongolica |
| 2 | Tegular apophysis with well developed ridge, and lower part as high as upper part (Fig. 25), free part of embolus bent (Fig. 26), epigynal windows wider than high (Figs 39, 42) | Xerolycosa nemoralis |
| – | Tegular apophysis has no developed ridge and lower part is higher than upper part (Fig. 27), free part of embolus gradually rounded, epigynal windows droplet-shaped, higher than wide (Figs 31, 33) | Xerolycosa miniata |
http://species-id.net/wiki/Xerolycosa_miniata
Figs 6–7, 22, 27–28, 31–34For a complete list of references see
FINLAND (selected records from different parts of the range): 15♂♀ (ZMUT), Nauvo Seili (60°15'N, 21°58'E), sandy sea shore, 16.05.-11.08.1974 (R. Mannila); 1♂ 1♀ (ZMUT), Virolahti Siikasaari (60°28'N, 27°35'E), sandy sea shore, 07.05.-13.09.1970 (S. Kännö); 17♂♀ (ZMUT), Pori Yyteri (61°33'N, 21°32'E), among Empetrum in sand dune, 14.07.1968 (P.T. Lehtinen); 1♀ (ZMUT), Hailuoto Marjaniemi (65°02'N, 24°36'E), Elymus sandy shore, 12.7.1973 (P.T. Lehtinen). RUSSIA:Adygeya: 1♂ 1♀ (TNU-2657/18), Caucasian State Reserve, 12 km SE kordon Guzeripl, Pastbishche Abago Mt. Range (43°53'N, 40°12'E 43°56'N, 40°16'E, 1727–2010 m a.s.l.), 18–23.08.2009 (M.M. Kovblyuk). Tuva: 13♂ 11♀ (IBPN), Uyuk River mouth, 52°04'N, 94°22'E, 600–700 m, 3–5.06.1995 (Y.M. Marusik). UKRAINE: Crimea: 1♀ (TNU-2187/1), Simferopol Distr., Kesslers' Forest, 8.08.2000 (M.M. Kovblyuk).
The species differs distinctly from Xerolycosa mongolica by the carapace pattern, having a light longitudinal band and stripes. Males can be distinguished from those of Xerolycosa nemoralis by the shorter seminal duct, a bent free part of the embolus and a bent tip, a rounded (not pointed) process of the tegular apophysis, basal part higher than apical (equal in Xerolycosa nemoralis), and the lack of a tegular ridge. Females can be distinguished by the proportions of the epigyne (windows longer than wide, whereas in Xerolycosa nemoralis they are wider than long).
Male. Total length 5.0 (4.7–6.2).
Carapace: 2.85 (2.52–3.09) long, 2.1 (1.79–2.22) wide. Carapace
length/femur IV ratio 1.2. Habitus and pattern as in
Palp as in Figs 22, 27–28, cymbial spines poorly distinct, upper part of tegular apophysis with claw-like outgrowth; embolus relatively thin, following an oval course, tip modified.
Length of leg segments:
| femur | patella | tibia | metatarsus | tarsus | Total | |
|---|---|---|---|---|---|---|
| I | 1.95 | 0.85 | 1.5 | 1.6 | 1.13 | 7.03 |
| II | 1.8 | 0.8 | 1.35 | 1.55 | 1.15 | 6.65 |
| III | 1.85 | 0.8 | 1.3 | 1.85 | 1.05 | 6.85 |
| IV | 2.38 | 0.95 | 1.85 | 2.8 | 1.38 | 9.35 |
Spination of legs:
| femur | patella | tibia | metatarsus | |
|---|---|---|---|---|
| I | 2p+2r | 1p+1r | 1p+2r+3-2v | 2p+1r+2-2v |
| II | 2p+2r | 1p | 2p+2r+2-2v | 2p+1r+2-2v |
| III | 2p+2r | 1p+1r | 2p+2r+2-2v | 2p+2r+2-2v |
| IV | 2p+1r | 1p+1r | 2p+2r+2-2v | 2p+2r+3-2v |
Female. Total length 7.0 (4.8–7.4). Carapace: 3.0 (2.46–3.28) long, 2.1 (I. 71–2.42) wide. Carapace length/femur IV ratio 1.25. Habitus and pattern as in Fig. 7; pattern on carapace same as in male but with less distinct lateral light stripes.
Epigyne as in Figs 31–34, sides of stem rounded, windows shaped like inverted droplets, their upper margins almost horizontal.
Length of leg segments:
| femur | patella | tibia | metatarsus | tarsus | Total | |
|---|---|---|---|---|---|---|
| I | 1.75 | 0.93 | 1.35 | 1.35 | 1.05 | 6.43 |
| II | 1.75 | 0.9 | 1.25 | 1.38 | 1.03 | 6.3 |
| III | 1.8 | 0.88 | 1.2 | 1.7 | 1.03 | 6.6 |
| IV | 2.4 | 1.05 | 1.85 | 2.7 | 1.3 | 9.3 |
Spination of legs:
| femur | patella | tibia | metatarsus | |
|---|---|---|---|---|
| I | 2p | 0 | 1p+2-2v | 2p+2-2v |
| II | 2p | 1p | 1p+2-2v | 2p+2-2v |
| III | 2p+2r | 1p+1r | 2p+2r+2-2v | 2p+2r+2-2v |
| IV | 2p+1r | 1p+1r | 2p+1r+2-2v | 2p+2r+3-2v |
Xerolycosa miniata has a Euro-Mongolian boreo-nemoral range (
http://species-id.net/wiki/Xerolycosa_mongolica
Figs 1–3, 10, 18–21, 23a-b, 29–30, 35–38.Holotype ♀ (MNHN) “Urga-Tsitsikar, Chaffanjon” [1896] (can refer either to Mongolia or China). RUSSIA, Tuva: 17♂ 2♀ (IBPN & ZMUT), SE Tuva, Erzin Town environs, 50°14'N, 95°09'E, 1165 m, dry steppe, 9.06.1995 (Y.M. Marusik & S. Koponen); 13♂ 1♀ (SZMN), SE Tuva, Erzin environs, 50°14'N, 95°09'E, 1165 m, Artemisia-Stipa steppe, 9.06.1995 (D.V. Logunov); 4♂ (ZMMU), SE Tuva, Tes-Khem Valley, 50°19'N, 95°01'E, 10.06.1995 (Y.M. Marusik); 3♂ (ZMMU), environs of Kyzyl, Nanophyton erinaceus semidesert steppe, 6.06.1995 (Y.M. Marusik).
The holotype female of Xerolycosa mongolica is very pale and the pattern is not visible. The figure in
Xerolycosa undulata
was described on the basis of the holotype male from Heilongjiang, not
far from Tsitsikar. According to the text (Chen et al. 1999), the type
was deposited in the Institute of Zoology in Beijing. However, the
type was not found in the collections (Li, personal communication).
Comparison of our figures of the male palp of Xerolycosa mongolica and figures of Xerolycosa undulata provided by
Xerolycosa mongolica differs distinctly from its congeners by its spotty pattern and lack of longitudinal bands or stripes on the carapace, widely spaced anterior median eyes (more than one diameter of AME), long filiform embolus, shape of the tegular apophysis, and structure of the epigyne and vulva.
Male. Total length 6.1 (5.6–6.25). Carapace: 3.05 (2.8–3.1) long, 2.1 (1.9–2.1) wide. Carapace length/femur IV ratio 1.07 (1.03–1.12). Habitus and pattern as in Figs 2–3.
Palp as in Figs 18–21, 23, 29–30, cymbium with distinct spines, apical part of tegular apophysis with triangular extension, embolus filiform along its entire course.
Length of leg segments:
| femur | patella | tibia | metatarsus | tarsus | Total | |
|---|---|---|---|---|---|---|
| I | 2.3 | 1.05 | 2.0 | 1.85 | 1.25 | 8.45 |
| II | 2.2 | 1.0 | 1.75 | 1.85 | 1.15 | 7.95 |
| III | 2.15 | 0.9 | 1. 5 | 2.1 | 1.05 | 6.2 |
| IV | 2.85 | 1.05 | 2.25 | 3.2 | 1.45 | 10.8 |
Spination of legs:
| femur | patella | tibia | metatarsus | |
|---|---|---|---|---|
| I | 1 or 2p+2r | 1p | 0 or 1p+2–2v | 1p+2–2v |
| II | 2p+2r | 1p | 2p+2–2v | 2p+2–2v |
| III | 2p+2r | 1p+1r | 2p+2r+2–2v | 2p+2r+2–2v |
| IV | 2p+2r | 1p+1r | 2p+2r+2–2v | 2p+2r+3–2v |
Female. Total length 6.6 (5.7–6.6). Carapace: 2.35 (2.35–2.7) long, 1.7 (1.7–1.85) wide. Carapace length/femur IV ratio 1.18 (1.1–1.18). Habitus and pattern as in Fig. 2.
Epigyne as in Figs 35–38, septum almost triangular in shape, upper margins of windows inclined.
Length of leg segments:
| femur | patella | tibia | metatarsus | tarsus | Total | |
|---|---|---|---|---|---|---|
| I | 1.7 | 0.75 | 1.3 | 1.2 | 0.85 | 5.8 |
| II | 1.6 | 0.7 | 1.1 | 1.1 | 0.8 | 5.3 |
| III | 1.5 | 0.7 | 0. 9 | 1.4 | 0.85 | 4.45 |
| IV | 2.0 | 0.75 | 1.5 | 2.35 | 1.2 | 7.8 |
Spination of legs:
| femur | patella | tibia | metatarsus | |
|---|---|---|---|---|
| I | 2p | 0 | 0 or 1p+3–2v | 2p+2–2v |
| II | 2p+0 or 1r | 0 | 1p+2–2v | 2p+2–2v |
| III | 2p+1r | 1p+1r | 2p+1r+2–2v | 2p+2r+2–2v |
| IV | 1 or 2p+2r | 1p+1r | 2p+2r+2–2v | 2p+2r+3–2v |
It seems that
Xerolycosa mongolica females make burrows in the ground in places with sparse steppic vegetation. The burrows are relatively deep 7–10.5 cm and 4–6 mm in diameter (Logunov, personal communication). Apparently males do not construct burrows. These observations were first made by Dmitri Logunov in Tuva. Subsequently we (Koponen and Marusik) witnessed this behaviour. It is worth mentioning that Xerolycosa mongolica seems to be the smallest burrowing wolf spider (Logunov, personal communication).
The exact distribution of this species is unknown because the type locality is uncertain (Urga-Tsitsikar), and because of incorrect synonymisation its distribution in China is unclear. Xerolycosa mongolica is well documented from Tuva only.

General appearance of Xerolycosa mongolica 1–3 Xerolycosa nemoralis 4–5 and Xerolycosa miniata 6–7 1, 5, 7 female, dorsal 2, 4, 6 male, dorsal 3 male, ventral.
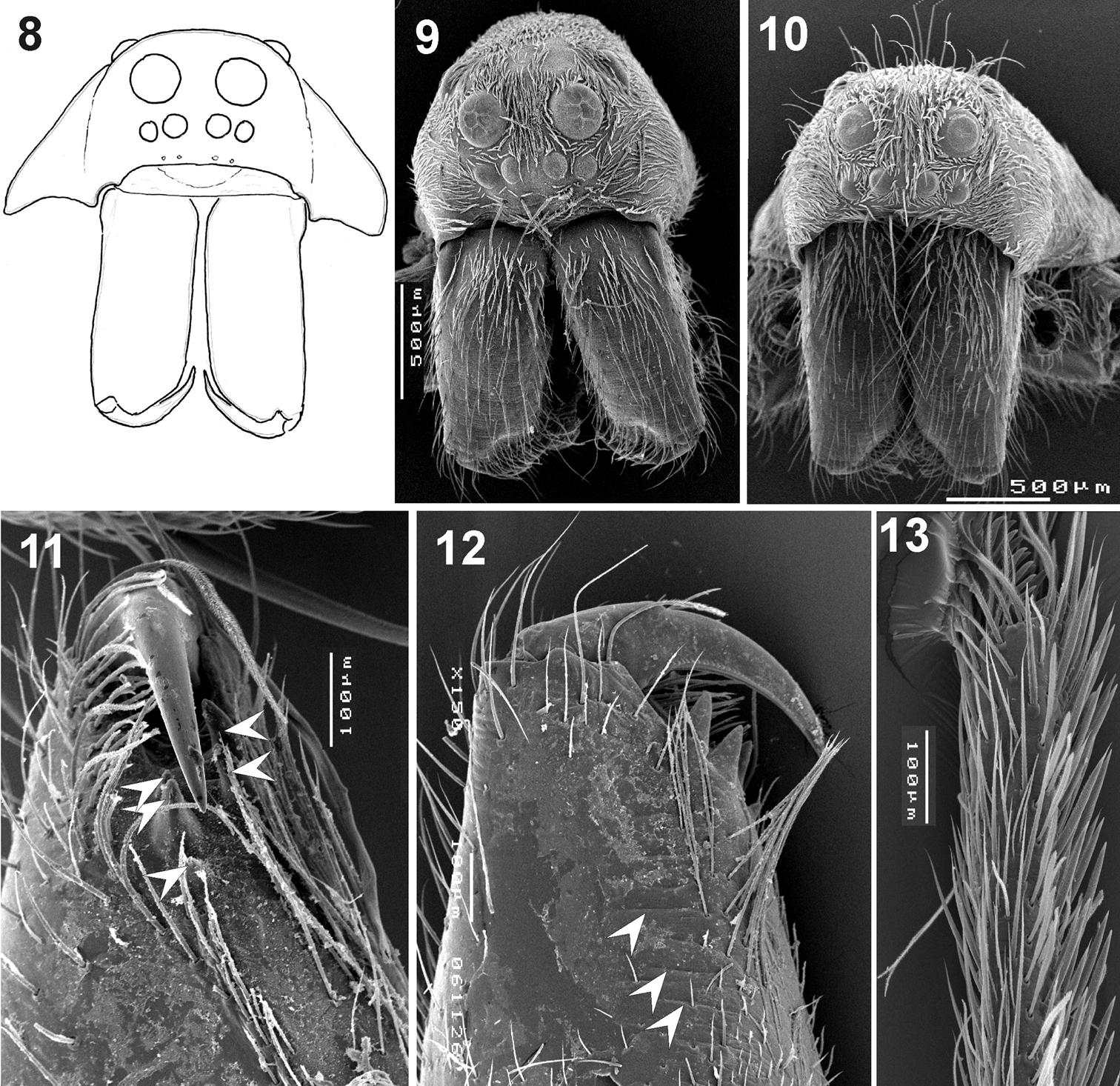
Somatic characters of Xerolycosa nemoralis 8–9, 11–13 and Xerolycosa mongolica 10 8–10 prosoma, frontal 11–12 – chelicerae, median and inner view 13 tarsus IV, retrolateral. Arrows show cheliceral teeth and stridulatory files.
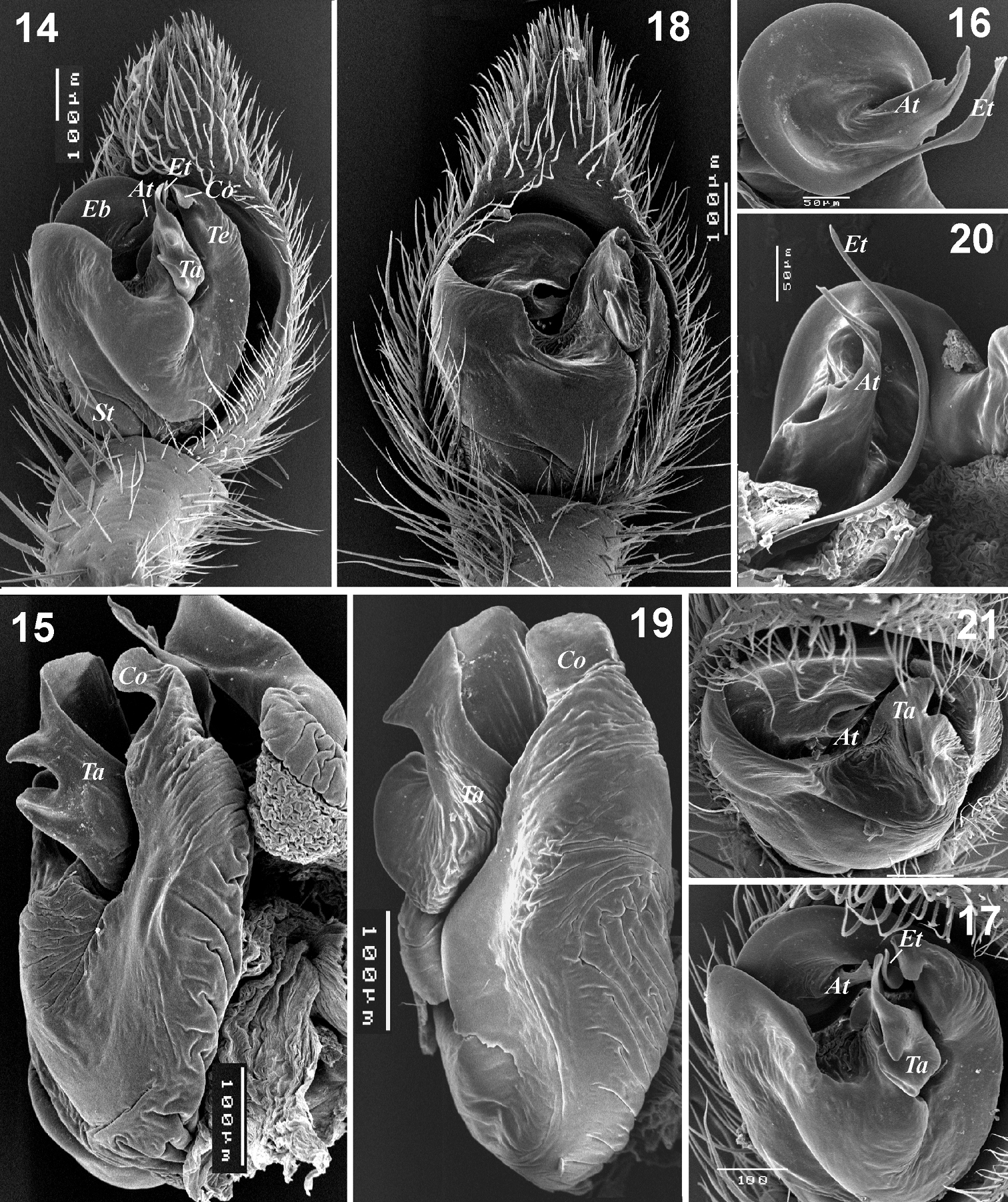
Male palp of Xerolycosa nemoralis 14–17 and Xerolycosa mongolica 18–21. 14, 18 whole palp, ventral 15, 19 – bulbus, lateral 16, 20 embolic division, ventral and ventro-retrolateral 17, 21 whole palp, apical. Abbreviations: At– terminal apophysis; Co – conductor; Eb– base of embolus; Et – tip of embolus; St– subtegulum; Ta – tegular apophysis; Te– tegular extension.
http://species-id.net/wiki/Xerolycosa_mongolica
Figs 4–5,For a complete list of references see
Xerolycosa nemoralis:
FINLAND (selected records from different parts of the range): 18♂♀ (ZMUT), Vuolijoki, Vuottolahti, Lapinniemi (64°13'N, 27°20'E), 16.07.1972 (P.T. Lehtinen); 1♂ (ZMUT), Hammarland Sålis (60°15'N, 19°44'E), dry forest, 26.06.-06.08.1971 (P.T. Lehtinen); 5♂ 4♀ (ZMUT), Turku Kärsämäki (60°30'N, 22°15'E), forest, 24.04.-04.08.1972 (I. Oksala); 12♂♀ (ZMUT), Harjavalta, Sport center (61°17'N, 22°10'E), pine forest, 09.07.-09.08.1992 (S. Koponen). RUSSIA: Adygeya:2 ♀ (TNU-2718/18), Caucasian State Reserve, env. kordon Guzeripl (44°00'N, 40°08'E, ~ 670 m), Abies & Fagus wood, 13–17.08.2009 (M.M. Kovblyuk); 2 ♀ (TNU-2719/3), Caucasian State Reserve, env. kordon Guzeripl (44°00'N, 40°08'E, ~ 670 m), Abies & Fagus wood, pitfalls, 16-23.08.2009 (M.M. Kovblyuk). Krasnoyarsk Prov.: 1♀ (IBPN), West Sayany Mts., Oiskiy Mt. Range, Buiba Riv., 52°47'N, 93°18'E, 1230 m, among stones, 20-21.06.1995 (Yu.M. Marusik). Sakhalin Island: 4♂ 4♀ 13 juv. (IBPN), SE part, Tsapko Vill. env., Zhdanko Mt. Range, 48°06.051'N, 142°31.557'E, h 220m, foothill of Vladimirovka Mt., stony debris along temporary creek, 1.08.2001 (Y.M.Marusik ); 5♂ 2♀ (IBPN), SE part, env. of Starodubskoye Vil., Naiba River mouth part, 47°24.992'N, 142°45.384'E, 23.07.2001 (Y.M.Marusik); 4♂ 5♀ (IBPN), SW part, Krilyon Peninsula, W shore, ca 5 km S of Shebunino Vil., Kitosia River mouth, 36°22.536'N, 141°52.562'E, 14-15.08.2001 (Y.M.Marusik). Magadan Area: 1♂ (IBPN), ca. 30 km N of Magadan, Dukcha River Valley, gravely bank, June 1995 (Y.M.Marusik). Kamchatka Province: 1♂ (IBPN), Kamchatka Peninsula, 10–12 km N of Paratunka Vil., Yelizovo Forestry, 53.050°N, 158.225°E, 15-28.07.2004 (A.S. Ryabukhin). No precise data: numerous males and females have been collected on five Kuril Islands: Kunashir, Iturup, Urup, Chirpoi and Paramushir.
The species differs distinctly from Xerolycosa mongolica by the pattern of its carapace, having longitudinal bands and stripes, and by having the anterior median eyes situated more closely together (less than one diameter of AME, more than one diameter in Xerolycosa mongolica). From Xerolycosa miniata, males can be distinguished by their longer seminal duct, rounded embolus, the sharply pointed process of the tegular apophysis and by the proportions of the epigyne (windows wider than high, whereas in Xerolycosa miniata they are higher than wide). Xerolycosa nemoralis females possess 2 retrolateral spines on femur I whereas the other species have only 2 prolateral spines.
Male. Total length 6.0 (5.5–6.8). Carapace: 2.9 (2.75–2.9) long, 2.0 (1.9–2.0) wide. Carapace length/femur IV ratio 1.23 (1.12–1.23). Habitus and pattern as in Fig. 4.
Palp as in Figs 14–17, 24–26, tip of cymbium with poorly developed spines, upper part of tegular apophysis with bill-shaped extension, embolus relatively thick, free part (=embolus proper) bent, tip modified.
Length of leg segments:
| femur | patella | tibia | metatarsus | tarsus | Total | |
|---|---|---|---|---|---|---|
| I | 1.85 | 0.85 | 1.5 | 1.55 | 1.05 | 6.8 |
| II | 1.85 | 0.85 | 1.35 | 1.4 | 1.1 | 6.55 |
| III | 1.7 | 0.8 | 1.25 | 1.75 | 1.05 | 6.55 |
| IV | 2.35 | 1.0 | 1.8 | 2.8 | 1.35 | 9.3 |
Spination of legs:
| femur | patella | tibia | metatarsus | |
|---|---|---|---|---|
| I | 2p+2r | 1p+1r | 1p+2r+3–2v | 2p+1r+2–2v |
| II | 2p+2r | 1p+1r | 2p+2r+2–2v | 2p+1r+2–2v |
| III | 2p+2r | 1p+1r | 2p+2r+2–2v | 2p+2r+2–2v |
| IV | 2p+1r | 1p+1r | 2p+2r+2–2v | 2p+2r+3–2v |
Female. Total length 6.7 (6.4–7.1). Carapace: 3.25 (2.8–3.35) long, 2.25 (2.0–2.4) wide. Carapace length/femur IV ratio 1.18 (1.14–1.18). Habitus and pattern as in Fig. 5.
Epigyne as in Figs 39–42, windows wider than high, septum with rounded sides.
Length of leg segments:
| femur | patella | tibia | metatarsus | tarsus | Total | |
|---|---|---|---|---|---|---|
| I | 2.25 | 1.0 | 1.75 | 1.7 | 1.3 | 8 |
| II | 2.15 | 0.95 | 1.65 | 1.65 | 1.3 | 7.7 |
| III | 2.15 | 0.9 | 1. 6 | 2.0 | 1.25 | 6.3 |
| IV | 2.75 | 1.05 | 2.2 | 3.25 | 1.5 | 10.75 |
Spination of legs:
| femur | patella | tibia | metatarsus | |
|---|---|---|---|---|
| I | 2p+2r | 1p | 1p+3–2v | 2p+2–2v |
| II | 2p+2r | 1p | 1p+2–2v | 2p+2–2v |
| III | 2p+2r | 1p+1r | 2p+2r+2–2v | 2p+2r+2–2v |
| IV | 2p+1r | 1p+1r | 2p+1r+2–2v | 2p+2r+3–2v |
Judging from the figures, the record of Xerolycosa nemoralis by Yin et al. (1997: f. 3a-d) from China refers to another species and even a different genus.
Xerolycosa nemoralis has a trans-Palaearctic boreo-nemoral range (
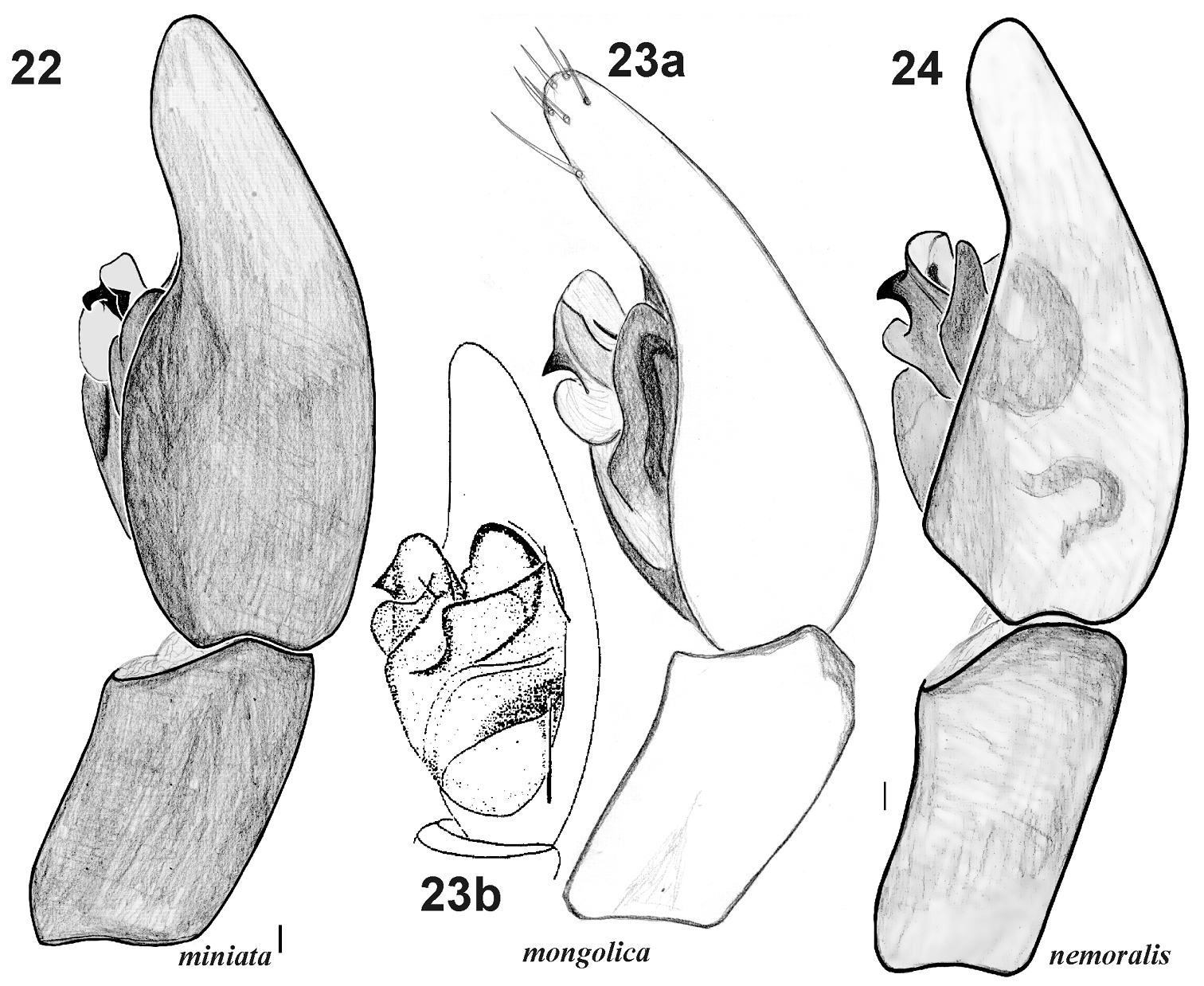
Male palp of Xerolycosa miniata 22 Xerolycosa mongolica 23 and Xerolycosa nemoralis 24 22, 23a, 24 retrolateral 23b prolateral. 23b after
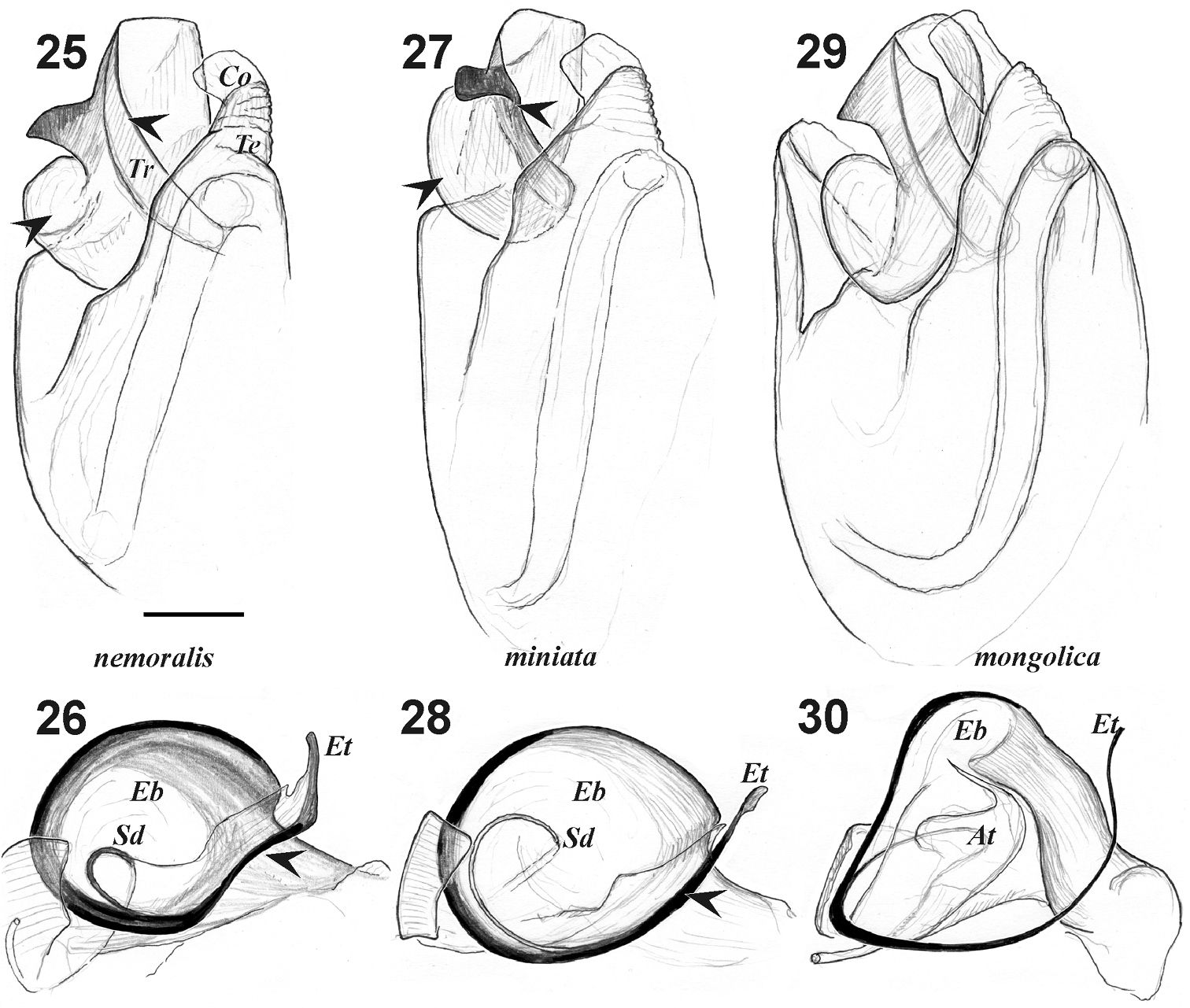
Male palp of Xerolycosa nemoralis 25–26 Xerolycosa miniata 27–28 and Xerolycosa mongolica 29–30 25, 27, 29 bulbus, retrolateral 26, 28, 30 embolic division, ventral. Arrows indicate differences between Xerolycosa miniata and Xerolycosa nemoralis. Abbreviations: At– terminal apophysis; Co – conductor; Eb –base of embolus; Et – tip of embolus; Sd – seminal duct.
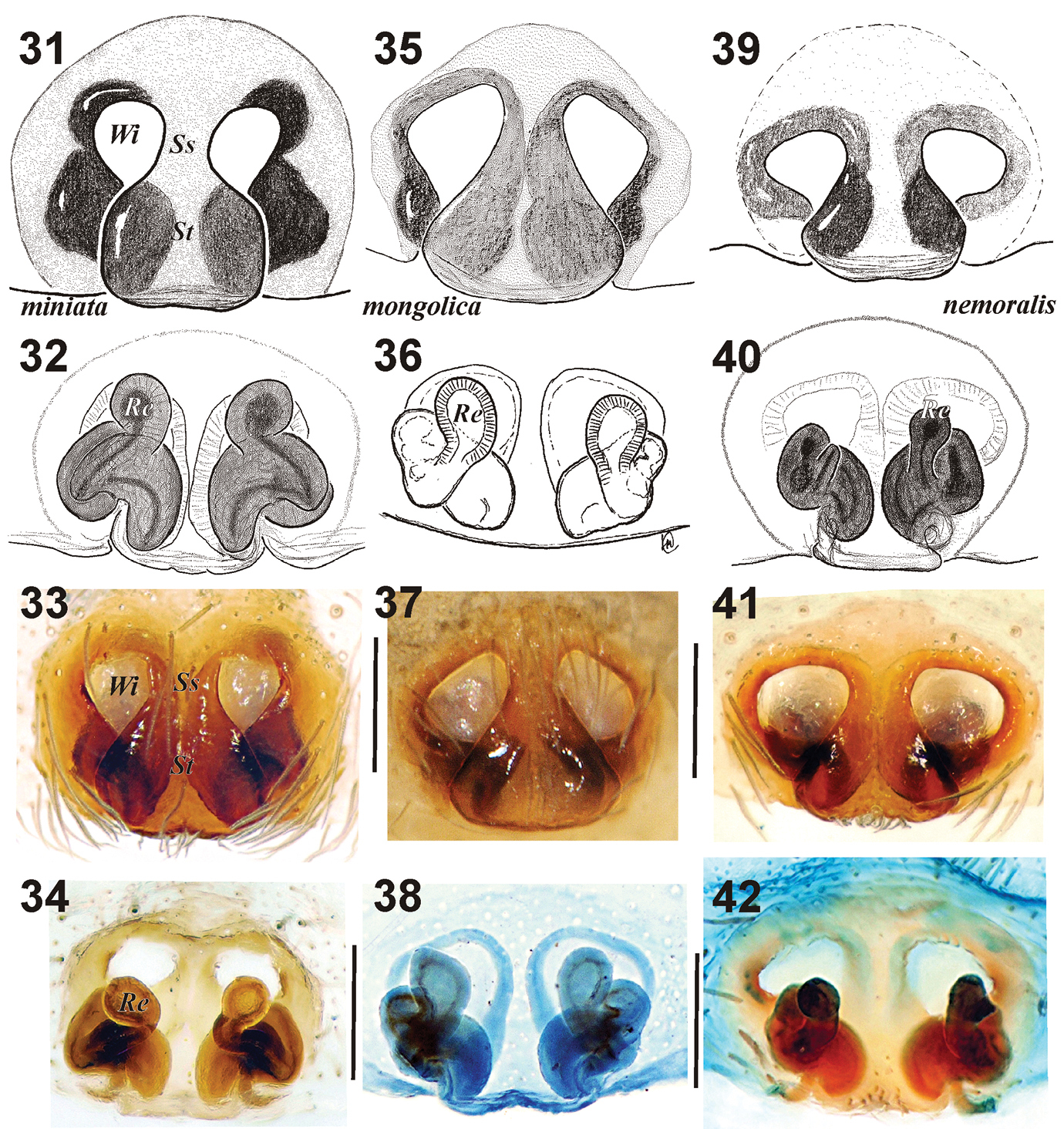
Epigyne of Xerolycosa miniata 31–34 Xerolycosa mongolica 35–38 and Xerolycosa nemoralis 39–42 31, 33, 35, 37, 39, 41 ventral 32, 36, 40, 34, 38, 42 dorsal. (36 & 38 holotype). Abbreviations: Re– receptaculum; Se– septum; Ss – septal stem;Wi – window of epigyne.
We thank the following colleagues who contributed to this study: C. Rollard (MNHN) and Shuqiang Li (Institute of Zoology, Beijing, China) for loan of type material or providing information about types; D.V. Logunov (Manchester, UK) showed us X. mongolica in the field and informed us about its burrowing ecology and A.A. Zyuzin's ideas about placement of this species; T. Kronestedt (Stokholm, Sweden) and C. Deltshev (Sofia, Bulgaria) reviewed the manuscript. N. Tsurusaki helped us with Japanese literature and N. Fritzén (Vaasa, Finland) gave us freshly collected specimens of X. miniata and X. nemoralis that were used for making digital photographs. The English of the final draft was kindly checked by D. Penney (Manchester, UK).
This work was supported in part by the RFFI grants # 09-04-01365 and 11-04-01716-a and by the Karadag Nature Reserve.