





(C) 2011 Pier Mauro Giachino. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Paralovricia gen. n. beroni sp. n., belonging to the new subtribe Lovriciina, is described from two caves in the Western Rhodopes (Bulgaria): Kraypatnata peshtera, near Smilyan Village (Smolyan Municipality) and Snezhanka, near Peshtera Town (Peshtera Municipality). Three currently known genera, Lovricia Pretner, 1979, Neolovricia Lakota, Jalžić & Moravec, 2009 and Paralovricia gen. n. represent a monophyletic unit supported by important synapomorphies that allows to propose the establishment of the new subtribe Lovriciina, characterized by the following characters: last maxillary palpomeres very long and narrow, basal angles of pronotum without seta, elytra without discal setae, elytral umbilicate series of nine pores in which the main pores are the 2nd, 6th and 9th, apical recurrent striole absent and mesotibial apex expanded outward. Its systematic position within the subfamily Trechinae (sensu Lorenz 2005) is discussed and Lovriciina is placed near Anillina. Key to the genera of the subtribe is proposed. Distribution data and zoogeographical hypotheses are discussed.
Coleoptera, Carabidae, new species, new genus, new subtribe, Lovricia, Neolovricia, Rhodopes, Bulgaria
The genera Lovricia Pretner, 1979 and Neolovricia
Lakota, Jalžić & Moravec, 2009 are sporadically and restrictedly
distributed in Dinaric Alps. To date they contain three very rare
species known only after few specimens. Lovricia jalzici Pretner, 1979 is presently known only for a single female specimen from cave Gospodska pećina in the vicinity of the village Cetina (Dinara Mt.); Lovricia aenigmatica
Lakota, Mlejnek & Jalžić, 2002, for one male and one female from
an unnamed pit near the peak Sveti Jure on the Biokovo Mt., which
recently received the name of Lovrićija Jama I (
The systematic position of Lovricia, and its related genus Neolovricia, remains controversial.
The discovery, by Petar Beron, Boyan Petrov and Pavel
Stoev, in two different caves in the Rhodopes, of two specimens (male
and female) of a new genus and new species clearly belonging to the same
phyletic lineage (originally mentioned as “undefined blind carabid
beetle” by
The following acronyms have been used for depositories of material:
NMNHS National Museum of Natural History, Sofia, Bulgaria
CGi Collection Giachino, Torino, Italia.
The following acronyms have been used for the type material:
HT Holotype
PT Paratype
The specimens, whose drawings of the habitus and genitalia were made, were previously included in Canada balsam. Genitalia were pinned beneath the specimens. The drawings were made by means of a camera lucida connected to a Leitz Dialux. Measurements in millimetres (mm).
For measurement uniformity it was decided to use the same methodology proposed by
ABL apparent body length, from clypeus to apex of elytra;
WH head width, at the level of the first orbital setae;
WPm maximum width across pronotum;
WPa width across anterior angles of pronotum;
WPp width across posterior angles of pronotum;
LP length of pronotum from base to apex along midline;
WE maximum width of elytra;
LE length of the elytra, from apex of scutellum to apex of left elytron.
Only ABL measurements are reported, the other measurements are given as 7 ratios.
General width: WH/WPm and WPm/We.
Body parts: WPa/WPp, WPm/WPp, WPm/LP and WE/LE.
urn:lsid:zoobank.org:act:AA3230A9-D4BC-45C8-BC8F-0CEF2688F108
Paralovricia beroni sp. n.
A genus closely related to Lovricia and Neolovricia by the shape of the penultimate maxillary palpomeres, narrow and elongate, by the hind angle of the pronotum lacking seta, and by mesotibiae distally expanded outwards. Paralovricia gen. n. differs from both Lovricia and Neolovricia by the less “aphaenopsian” body shape, with a more cordiform pronotum and elytra with more evident and finely denticulate shoulders. From Lovricia, the new genus differs by the shape of the penultimate maxillary palpomere not subtriangular and by mesotibiae furnished of spurs besides the inner angles. From Neolovricia, the new genus differs by the labium with a developed median tooth and by the different shape of the female gonocoxite.
Body small (ABL = mm 1.80 - 2.19), elongate, rather flattened, anophthalmous, pubescent, reddish-ferrugineous, with palpi and last tarsomeres paler.
Head relatively large, clypeus separated from the frons by a transverse impression, with two pairs of long thin setae. Frontal furrows ending with round foveae impressed posteriorly, neck distinct; two supraorbital setae not reduced in length. Labrum transverse, with a slight emargination in the middle, bearing six setae. Mandibles slender with a simple apex. Maxillae strongly prominent, penultimate segments of maxillary palpi longer than broad, slightly clavate, terminal palpomeres protracted, needle-shaped and pointed. Mentum without a suture between labium and pre-basilar; labium with a large median tooth. Antennae moniliform. Surface fine, microreticulate, pubescence relatively dense, recumbent, as long as that of pronotum and elytra. Cephalic capsule covered by yellowish, recumbent, relatively long and sparse hairs.
Pronotum subcordiform, hardly explanate sides usually narrowed at posterior angles, with the maximum width at the anterior third. Anterior edge arcuate, with angles entirely rounded. Lateral margin hardly sinuated before rectangular posterior angles. Disc slightly convex, with longitudinal median furrow impressed. Basal transversal furrow deep and conspicuous. Surface with distinct puncturation and long, somewhat sparse, fine erected pubescence. Anterior sixth of their length with pair of marginal setae; basal setae lacking. Scutellum subtriangular.
Elytra longer than their combined width, rounded apically near the suture, wholly covering the abdomen, dorsoventrally slightly convex without a distinct marginal groove. Humeral angles rounded but evident; lateral margins with edges finely denticulate. Sculpture of elytra distinctly microreticulate consisting of wrinkled lines; striae missing; recurrent striola lacking. Elytral disc without discal setiferous punctures, with pubescence relatively dense, recumbent and long, not arranged in rows. Umbilicate series consisting in 9 setiferous pores, with 2nd, 6th and 9th ones bearing a long seta; a geminate pair is made by 5th and 6th pores, with the 5th placed after the 6th one.
Abdominal sterna smooth, with fine and sparse pubescence.
Legs relatively short. Femora robust. Protarsomeres not dilated in the male. Mesotibiae distally expanded outwards and fringed with dense bristles, inner angles with additional spurs. Metafemora smooth. Tarsal claws simple.
Aedeagus with the median lobe stout and poorly
arcuate, with a simple narrowly and irregularly sub-squared apex. Basal
bulb with the orifice greatly expanded dorsally and delimiting two
subequal lateral lobes as in Anillini (
Female genitalia with the gonocoxite separated from the subgonocoxite (Fig. 11). The latter narrow and elongated, drop-like shaped, totally free from setae or thorns. Gonocoxite stocky, angular and slightly curved, not pointed at the apex; bearing 3 stocky ensiform setae on the external-proximal edge and a dorsal one at the internal edge. The apical part of the gonocoxite shows, at the inner edge, a sensorial dimple bearing two fine and short nematiform setae. Spermatheca ((Fig. 10) short and stocky, membranous, not sclerotized; spermathecal gland not found.
urn:lsid:zoobank.org:act:FFDDD4DB-5B2E-4036-9DD2-C30C3A14A347
http://species-id.net/wiki/Paralovricia_beroni
Figs 1-11Bulgaria, Western Rhodopes, Smolyan Municipality, near the village of Smilyan, Kraypatnata peshtera cave, 41.5123° N; 24.7600° E, 780 m.
HT ♂, Bulgaria, Western Rhodopes, Smolyan Municipality, near the village of Smilyan, Kraypatnata peshtera cave, 41.5123° N; 24.7600° E, 780 m, 11.VII.1997, Boyan Petrov leg. (NMNHS). PT: 1 ♀, Bulgaria, Western Rhodopes, Peshtera Municipality, near the town of Peshtera, Snezhanka cave, 42.0092° N; 24.2720° E, 860 m, 17.VI.2005, Petar Beron & Pavel Stoev leg. (CGi).
Note: Male HT was completely dismembered and lacking of abdominal sternites and left metathoracic leg. The drawing of the habitus of this specimen is therefore entirely reconstructed on the basis of individual anatomical parts that are included now in Canada Balsam.
Body small (ABL = mm 1.80 ♂ 2.19 ♀), elongate, rather flattened, anophthalmous. Pubescence very sparse, short, yellow, recumbent.
Head relatively large but narrower than pronotum (WH/WPm = 0.97 ♂, 0.95 ♀), clypeus truncate with the frontoclypeal sulcus distinct. Frontal furrows with posterior round foveae, occiput coarsely and densely punctate. Mandibles slender with a simple apex. Maxillae strongly prominent, penultimate segments of maxillary palpi longer than broad, bearing 4 setae, terminal palpomeres protracted, needle-shaped, with an apical tuft of sensillae (Figs 3-4). Labium (Figs 5-6) with a large median tooth, showing two small basal setae; mentum with a large, rounded, depressed fovea, latero-posteriorly surrounded by a ring of 10-12 setae. Antennae moniliform from the fourth antennomere onwards, long, markedly exceeding the humeral portion of the elytra when stretched backwards. Cephalic chaetotaxy as in the description of the genus.
Pronotum slightly convex, subcordiform (WPa/WPp = 1.42 ♂, 1.54 ♀), with the maximum width at the anterior third (WPm/LP = 1.09 ♂, 1.10 ♀). Anterior angles obtuse and broad. Lateral margin hardly sinuated before the posterior angles, which are rectangular and slightly projecting laterally. Punctures of the disc nearly equal to those of the occiput. Anterior sixth of their length with a pair of marginal setae; basal setae lacking. Scutellum subtriangular, pointed apically, with distinct transverse cells.
Elytra longer than their combined width (WE/LE =
0.62 ♂, 0.66 ♀), widest closely behind one half of their length.
Humeral angles rounded but evident; lateral margins without a distinct
marginal groove but with edges finely denticulate. Sculpture of elytra
distinctly microreticulate consisting of wrinkled lines; striae missing;
recurrent striola lacking, disc without discal setiferous punctures.
Scutellar pore umbilicate and shifted from its normal position, placed
near the front edge of the elytra. Umbilicate series as in Figs 1-2, consisting in 9 setiferous pores; the main umbilicate pores bearing a long seta (sensu
Protarsomeres not dilated in the male. Mesotibiae (Figs 7-8) distally expanded on outwards and fringed with dense bristles, inner angles with additional spurs. All the last tarsomeres of pro- meso- and metatibiae hyaline and with a peculiar shape: widened at the base and narrowed at the apex.
Aedeagus (Fig. 9) with median lobe stout and poorly arcuate; apex, in lateral view, stout, and irregularly sub-squared, slightly bent downwards. Basal bulb of the median lobe small, with the basal orifice greatly expanded dorsally, reaching about one third of the length of the median lobe, delimiting two subequal lateral lobes. Shape of left and right parameres similar to each other, long, strongly widened at the base, sharply restricted, elongated and strongly curved upwards in the apical third. One large and stout coaxial seta at the apex and a second one, frail, small, ventral, in a preapical position. Inner sac with a median copulatory sclerite, clew-shaped with two dorsolateral branches.
Female genitalia as in the description of the genus.
This interesting new species is dedicated to one of its collectors, Dr. Petar Beron, a passionate biospeleologist, former Director of the National Natural History Museum of Sofia and former Vice-President of the Bulgarian Parliament, as a sign of friendship and esteem for the impetus given to the knowledge of the Bulgarian subterranean fauna.
Paralovricia beroni gen. n. sp. n. was discovered in the cave Kraypatnata peshtera (in English: “cave near the way”). The cave (Fig. 12)
is situated on the left riverbank of the river Arda, at an altitude of
780 m a.s.l. and approximately 1 km east of the village of Smilyan. It
is a diaclase cave with a total length of 38 m, -10 m in depth, and a
precipice at the end. The cave entrance is situated about 2-3 meters
above the level of the road Smilyan-Rudozem. Air temperature measured in
the last chamber is 12°C. The cave has an ascending principal gallery,
dripping water in some places and the floor covered with wet clay,
rotten logs, and some bat guano. The beetle fauna there consists of Laemostenus plasoni plasoni (Reitter, 1885) and the Leptodirine Gueorguievella petrovi Giachino & Guéorguiev, 2006 (
The second known locality (Fig. 12),
cave Snezhanka (in English: “Snow-White”) is a national tourist site.
This cave is provided with utilities and has limited access to the
interior. The cave is situated 5 km southwest of the town of Peshtera,
on the left slope over the Novomachlenska reka River, a tributary of
the Stara reka River (
It is worth mentioning that the distance between these two caves is 64-65 km by airline (Fig. 12)
and that the same species lives in such a relatively wide distance.
This is not only a remote question, but between these points are
situated the valley of Vacha River and first and third highest
elevations of the Rhodopes. Chernatitsa Mt. (with maximal point Golyam
Persenk, 2091 m a.s.l.) in the north and the Perelik Mt. (with maximal
point Golyam Perelik, 2191 m a.s.l.) in the south form united mountain
ridge with lowest points between them the col Pamporovo (1620 m a.s.l.)
and the col Prevala (1665 m a.s.l.). This seems to confirm that the
apparent rarity of one species cannot be presumed as synonymous of short
range distribution. It may be attributed instead to our lack of
bionomic knowledge. Indeed, both caves were visited several times by
biospeleologists at any time, but no more specimens from this new genus
have been found. For instance, after finding of this new species, the
Kraypatnata peshtera Cave was visited six times, and the Snezhanka
Cave more than ten times after that. According to
As already discussed in the introduction, the systematic position of the genera complex formed by Lovricia and Neolovricia, to which now Paralovricia gen. n. is added, has always been controversial. The lack of knowledge on the morphology of the aedeagus, even in a single known species, together with a too brief, too superficial, or misinterpreted description, of a number of important characters, such as the elytral chaetotaxy, helped to postpone the solution of the problem. In this way, some important phylogenetic characters could not be controlled with certainty because they were misinterpreted or omitted from the original descriptions. For example, we do not know if, even in Lovricia and Neolovricia, scutellar setiferous pores are moved toward the elytral base. While, conversely, an examination of the original drawings, although incomplete (in small specimens drawn without inclusion in Canada Balsam), allows us to say with good approximation that the umbilicate series of Lovricia and Neolovricia are similar to that of Paralovricia.
The three currently known genera, Lovricia, Neolovricia and Paralovricia represent a clear monophyletic unit supported by important synapomorphies that allow us to propose the establishment of a new subtribe.
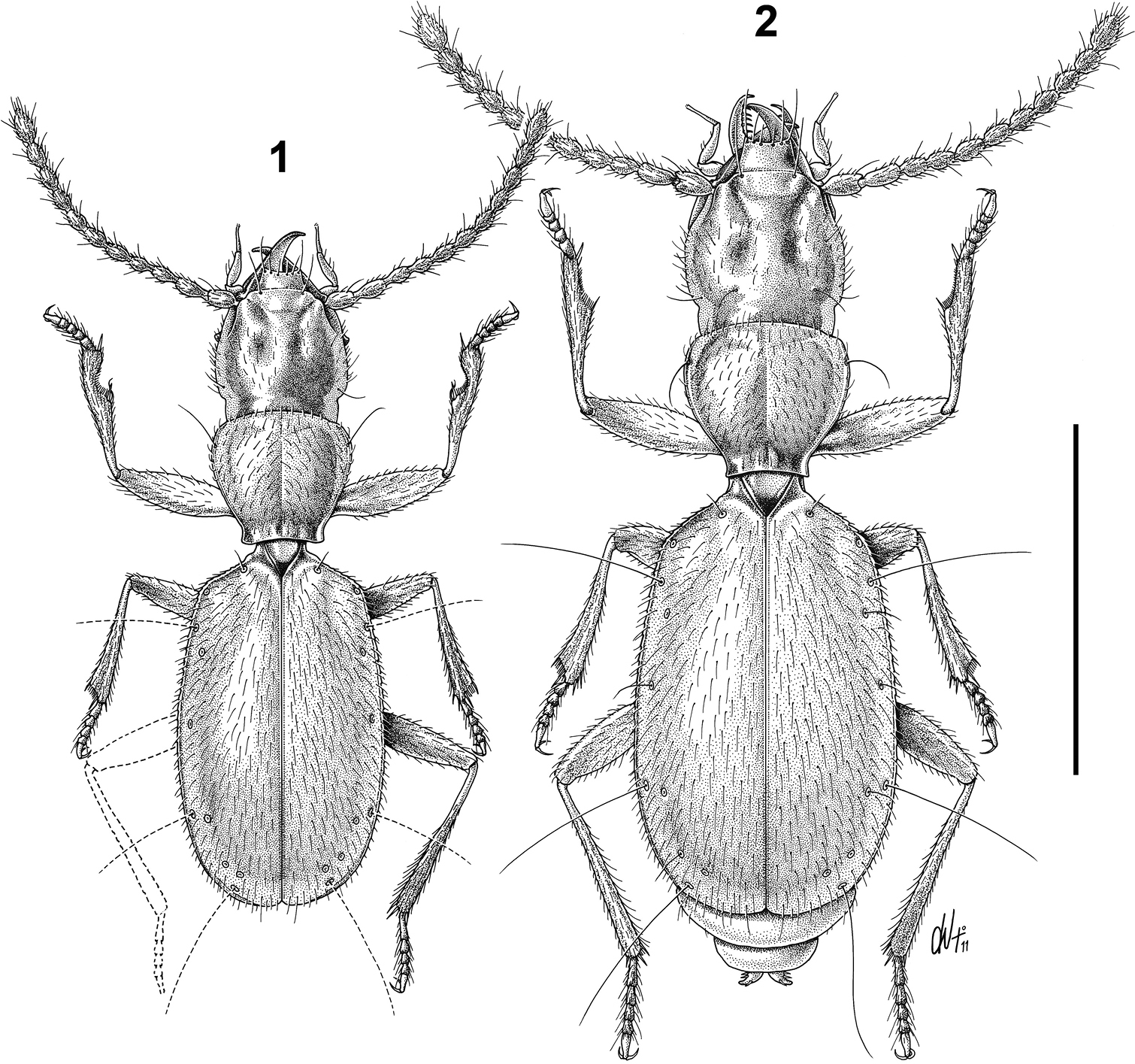
Habitusof Paralovricia gen. n. beroni sp. n. 1 HT ♂ 2 PT ♀. Scale bar: 1 mm.
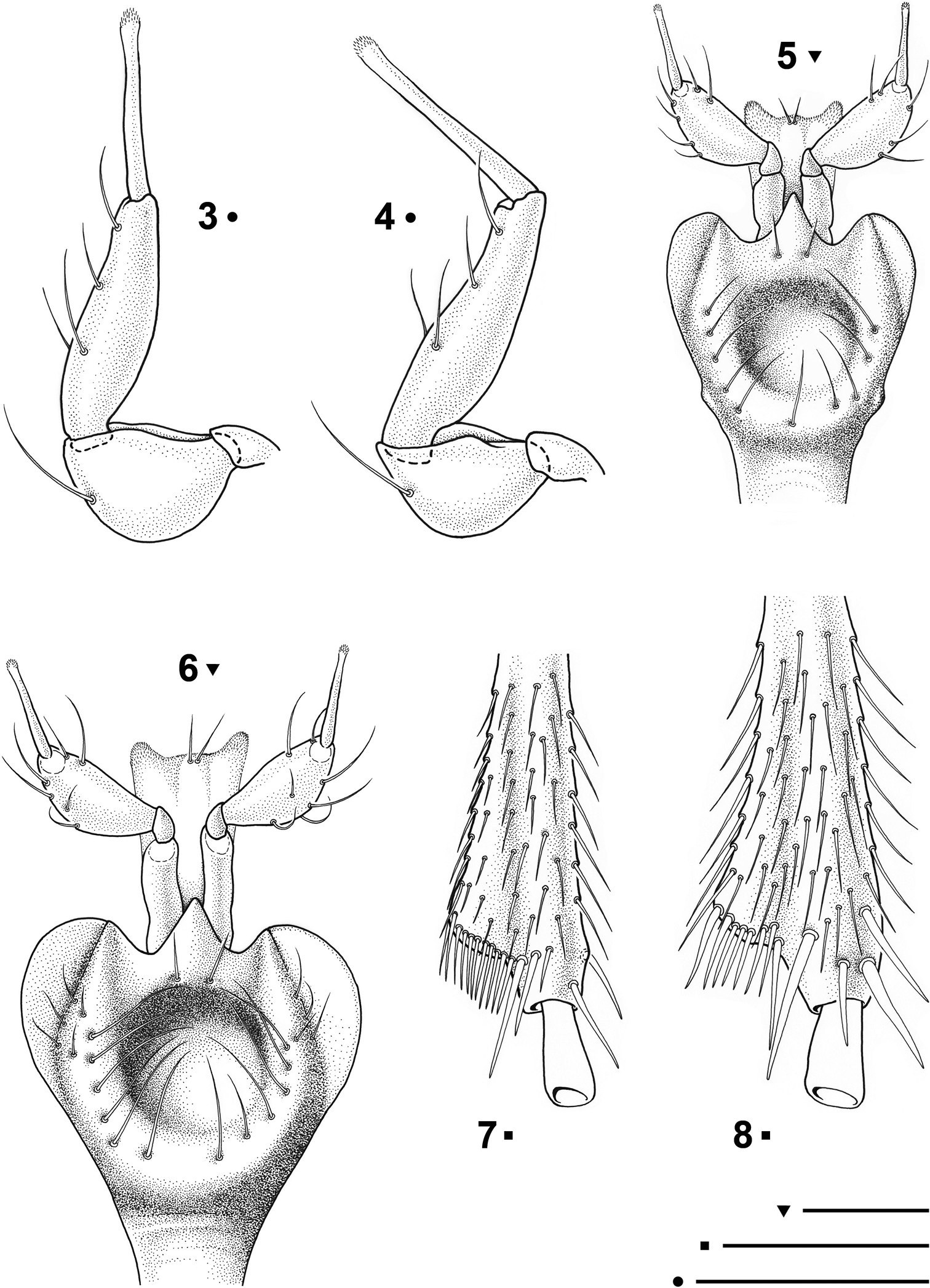
Paralovricia gen. n. beroni sp. n. 3 Maxillary palpomere, HT ♂ 4 Maxillary palpomere, PT ♀ 5 Mentum, labial palpomeres and ligula in ventral view, HT ♂ 6 Mentum, labial palpomeres and ligula in ventral view, PT ♀ 7 Apex of right mesotibia in ventral view, HT ♂ 8 Apex of right mesotibia in ventral view, PT ♀. Scale bars: 0.1 mm.
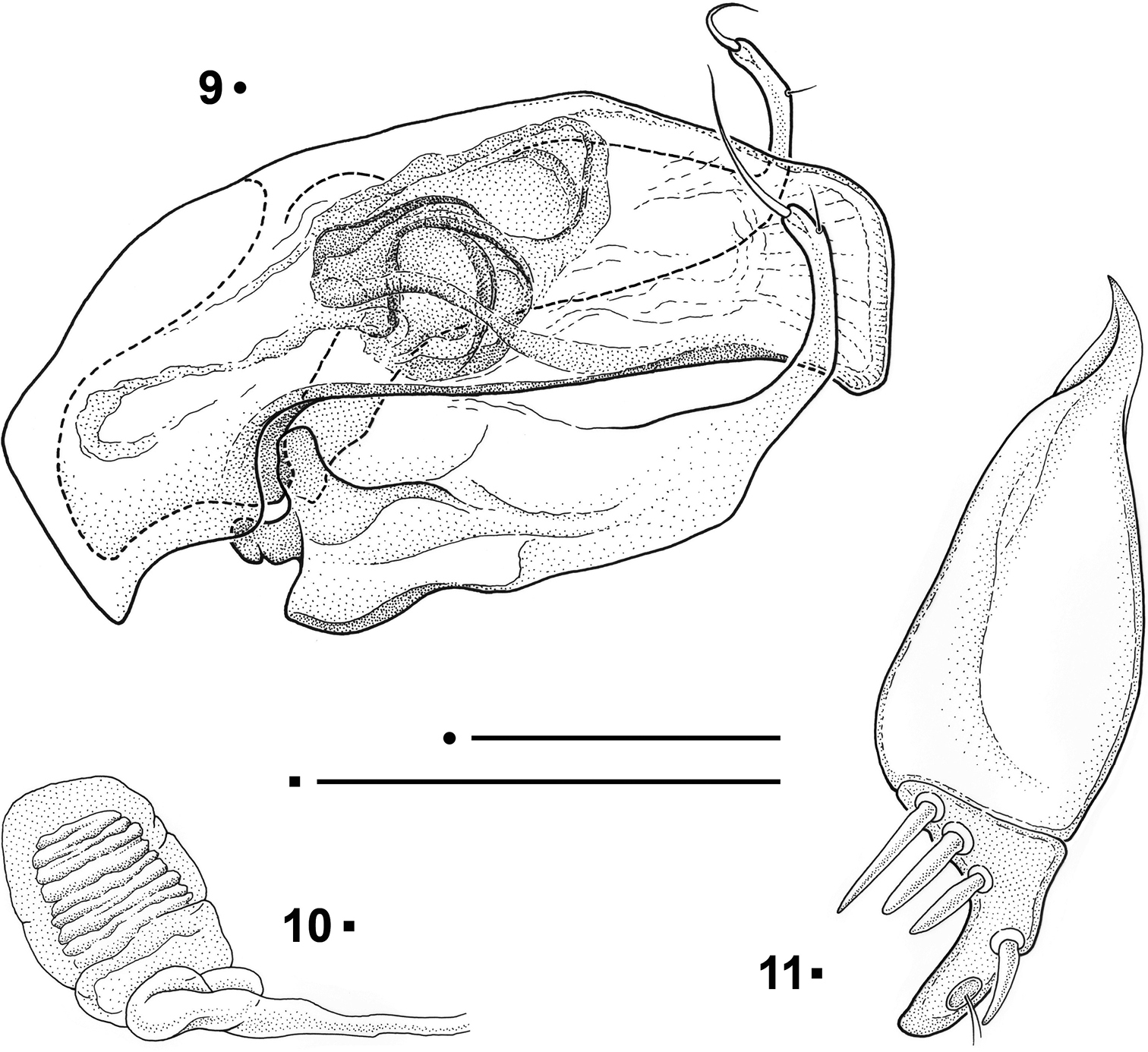
Paralovricia gen. n. beroni sp. n. 9 Aedeagus in lateral view, HT ♂ 10 Spermatheca, PT ♀ 11 Right gonocoxite in ventral view, PT ♀. Scale bars: 0.1 mm
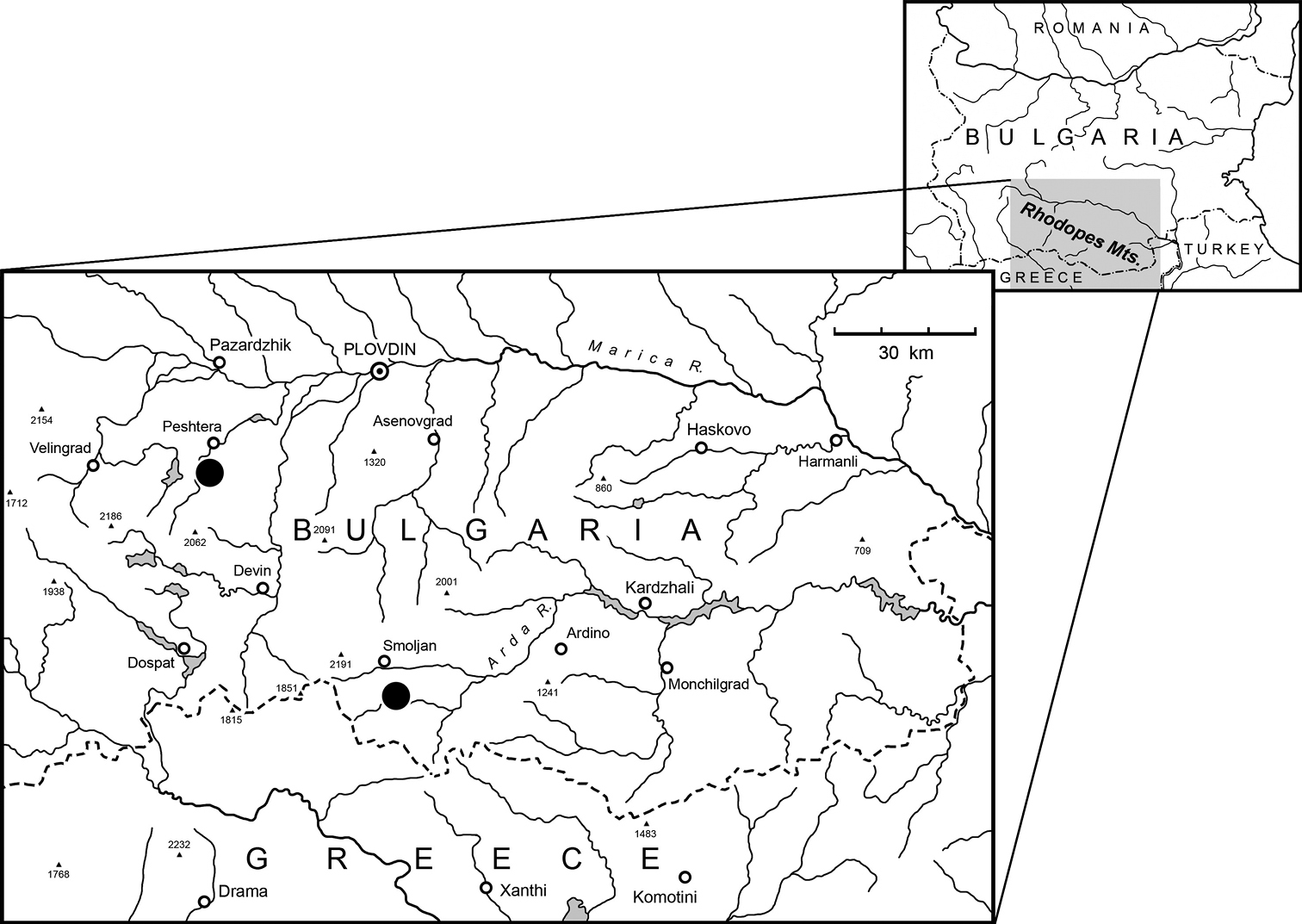
Distribution map of genus Paralovricia gen. n. (black circles indicate two known findings of Paralovricia beroni sp. n.).
Lovricia Pretner, 1979
A subtribe characterized by genera that present the following synapomorphic characters: last maxillary palpomeres very long and narrow, basal angles of the pronotum without seta, elytra without discal setiferous punctures, elytral umbilicate series of nine pores in which the main pores are the 2nd, 6th and 9th ones, apical recurrent striole absent and mesotibial apex expanded outward.
Now the examination of several characters allows us to define better the position of Lovriciina within the subfamily Trechinae (sensu
| 1 | Pronotum cordate, wider than long (ratio pronotum width / pronotum length more than 1.05) | Paralovricia gen.n. |
| – | Pronotum sub-elongate, longer than wide (ratio pronotum width / pronotum length less than 0.95). | 2 |
| 2 | Smaller species (length of body less than 2.2 mm). Shoulders of elytra distinct, more or less angulate. | Neolovricia Lakota, Jalžic & Moravec, 2009 |
| - | Larger species (length of body more than 2.3 mm). Shoulders of elytra indistinct, oblique, without distinct angles. | Lovricia Pretner, 1979 |
Analysis, from a historical zoogeographical point of view, of the distribution of the phyletic lineage of Lovriciina
(Fig. 13) provides several interesting insights. First we must consider
that the currently known distribution, although widely disjoint,
ranging from the Dinarides to the Rhodopes and that, as widely
discussed for Anillina (
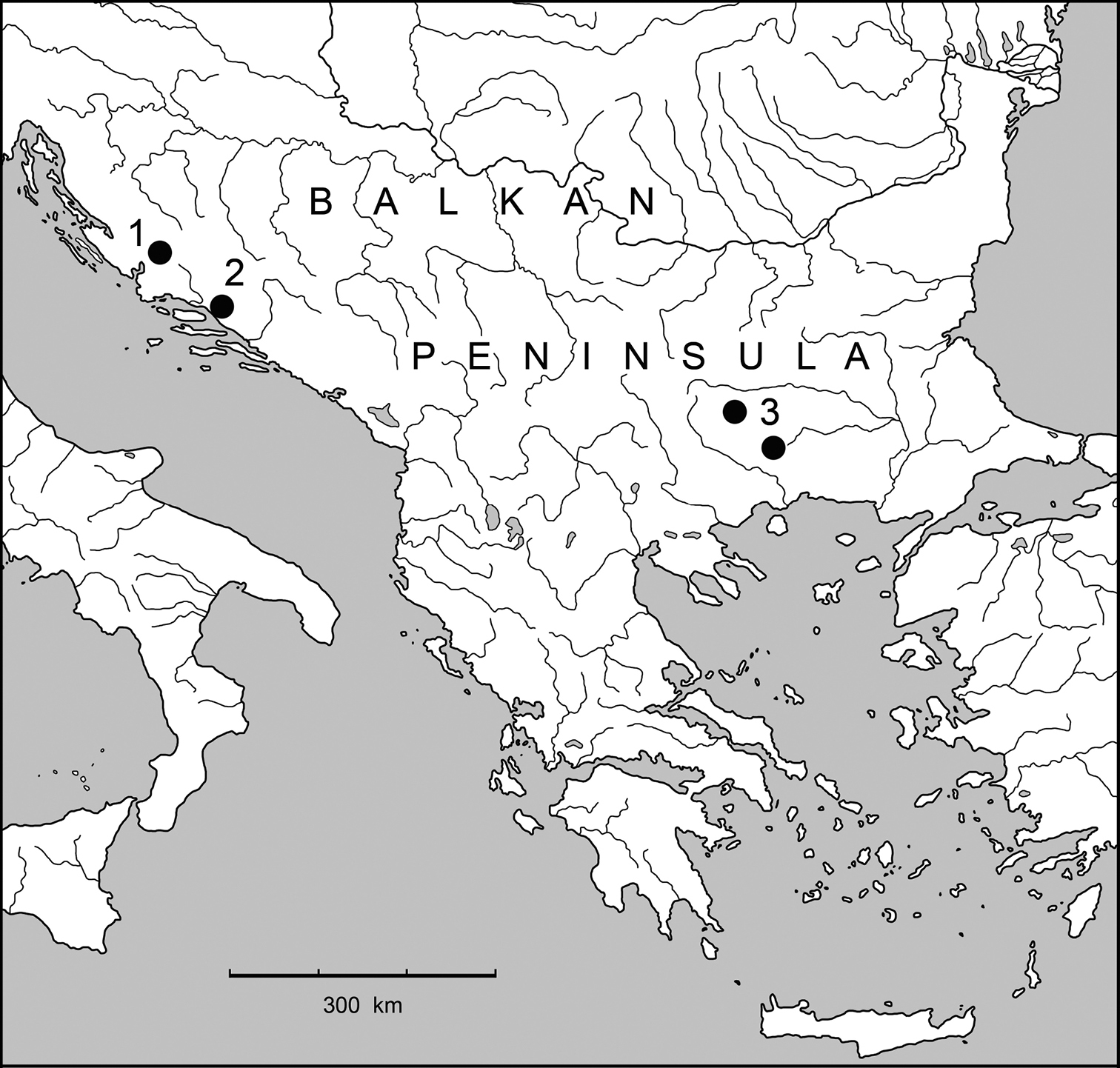
Distribution map of the species from subtribe Lovriciina (1 – Lovricia jalzici Pretner; 2 – Lovricia aenigmatica Lakota, Mlejnek & Jalžić and Neolovricia ozimeci Lakota, Jalžić & Moravec; 3 – Paralovricia beroni sp. n.).
We are very obliged to the following colleagues for giving us material for study: Petar Beron (NMNHS), Boyan Petrov (NMNHS), and Pavel Stoev (NMNHS). Special thanks to Boyan Petrov for providing us with proper information on the caves, and to Achille Casale (Dipartimento di Zoologia e Genetica Evoluzionistica, University of Sassari) for his critical remarks on the manuscript. Rostislav Bekchiev (NMNHS) helped us adjusting Fig. 12. Pensoft Publishers kindly provided support for the publishing this work.