





(C) 2011 Ming-Luen Jeng. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
The female of Pyrocoelia prolongata Jeng & Lai, a diurnal lampyrid species from Taiwan, is described for the first time. A single individual was found in a small, shady, dry streambed at the edge of a mixed forest at 2700 m elevation. The individual glowed in darkness and would move its abdomen up and down when disturbed and as a deterring behavior. A key to the females of the species of Pyrocoelia Gorham in Taiwan is provided. The morphology of photogenic organs of females and the function of bioluminescence of day-active species of Pyrocoelia are discussed.
Pyrocoelia prolongata, female, Taiwan, diurnal, Lampyrinae, Asia
The genus Pyrocoelia Gorham is a group of Asian lampyrids including more than 60 known species (
The nomenclature of Pyrocoelia was clarified and the species of Taiwan were revised by
The only female specimen available for study was collected by the late Japanese coleopterist Dr. M. Satô while digging in a dry streambed for ground beetles at high elevations in southern Taiwan. The female was kept in a transparent plastic cup which was spread with wet tissue paper and moss to maintain suitable humidity and for observation.
Measurements were made by outlining the target structures under a Nikon SMZ-10 microscope with a camera lucida. Body length (BL) was measured as the distance between pronotal and abdominal apices when the body was fully stretched, and body width (BW) as the greatest distance across the abdomen. The abbreviations PL and PW denote pronotal length and width, respectively; while T#, P#, and S# refer to the tergite, pleurite, and sternite of the #th abdominal segment (true segmentation), respectively.
Systematicshttp://species-id.net/wiki/Pyrocoelia_prolongata
The female is characterized by the bell-shaped
pronotum which resembles that of the male and is one of the diagnostic
characters of the species (
Female (Figs 1–7): BL: 15.6 mm; BW: 4.6 mm. Body form elliptical and depressed. Head, antennae, and legs dark brown to black; pronotum translucent gray, with margins and central carina light brown and central disc pink; mesonotum pink, central disc and mesoscutellum dark brown, translucent gray at sides (elytral rudiments); metanotum bronze; abdominal T1 lighter than metanotum and mixed of pink; T2–7 pinkish white, bronze on posterior margins and central line, and translucent gray at lateral margins (Fig. 7A); T8 translucent gray; ventral side pink, brightly so on thorax and pale on abdomen; S8 with a pair of milky white photogenic organs; S8–9 translucent yellowish brown. Head spherical, about 1/3 as broad as pronotum; vertex concave along central line; frons about as broad as clypeus-labrum; antennal sockets widely separated from each other; antenna (Fig. 2) 11-segmented, weakly serrate and rod-like; antennomere 1 largest and 3 slightly smaller than it; clypeus-labrum (Fig. 3) not fused with frons, shell shape; mandibles partially sclerotized, thick in basal 2/3 and slender and somewhat curved at apex; maxillary palpus 4-segmented; labial palpus 3-segmented. Pronotum bell-shape, strongly convex on central disc and weakly so in windows, reflexed at margins; windows on apical 1/3 and divided by central carina. PL/PW = 0.85. Mesonotum as broad as central disc of pronotum, with elytral rudiments abbreviated and round at sides, not extending beyond pronotum; mesoscutellum transverse, broader than long by 2 times, with apex slightly notched. Metanotum broader than pronotum, transversely elliptical, with a concave central line. Metepisternum fused with metasternum (Fig. 5). Legs (Fig. 4) moderate in width and length, 5th tarsomere slightly longer than preceding tarsomere. Abdomen broadest at T2, T1 about as broad as T3 and diminishing in width accordingly toward apex; T1–8 each with short but clear hind angles; T1–7 with a paler central line; T8 (Fig. 6) transverse and subquadrate, sinuate at sides and central apex. P1–7 folded on inner margins, each bearing a spiracle at center. S8 deeply notched centrally, with a pair of photogenic organs at sides (= PO, Fig. 6, Fig. 7B).
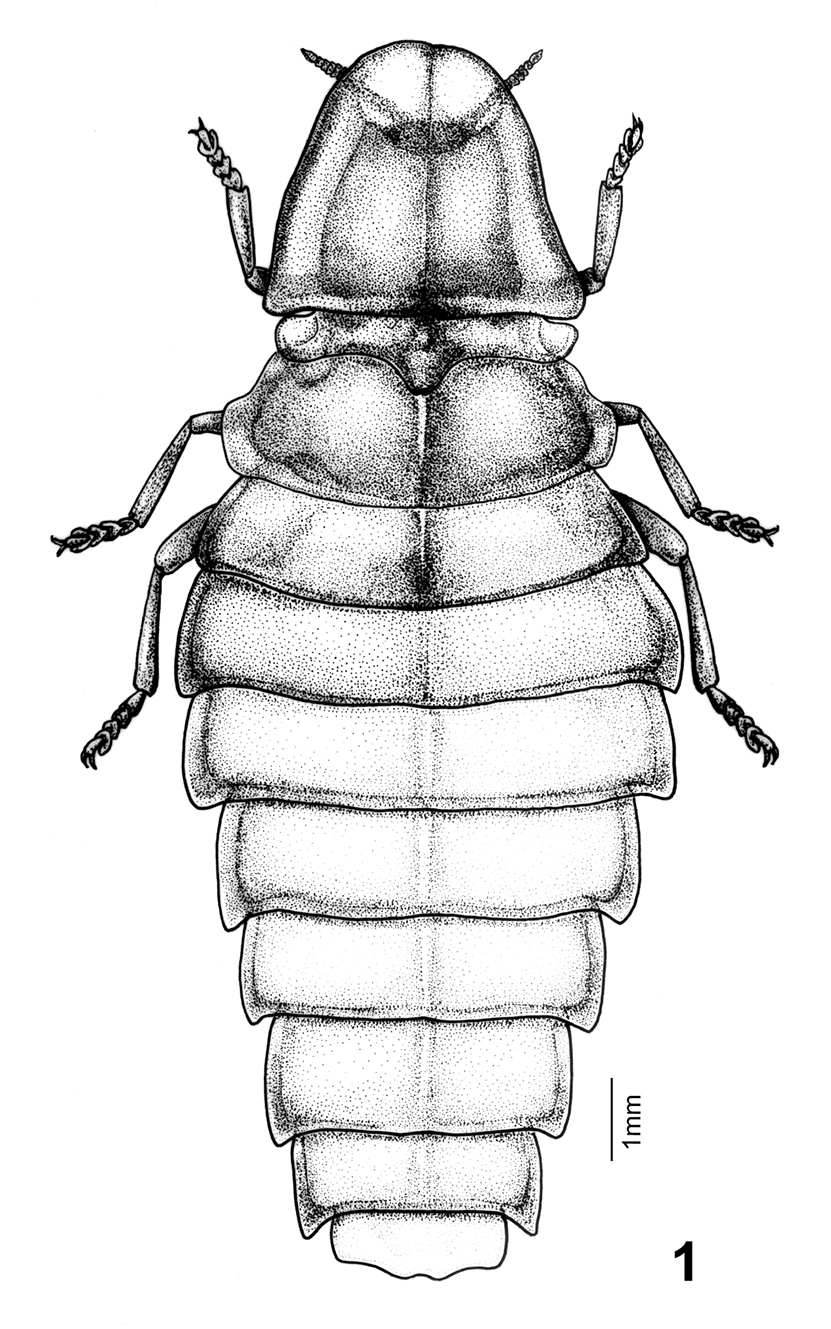
Female dorsal habitus of Pyrocoelia prolongata Jeng and Lai.
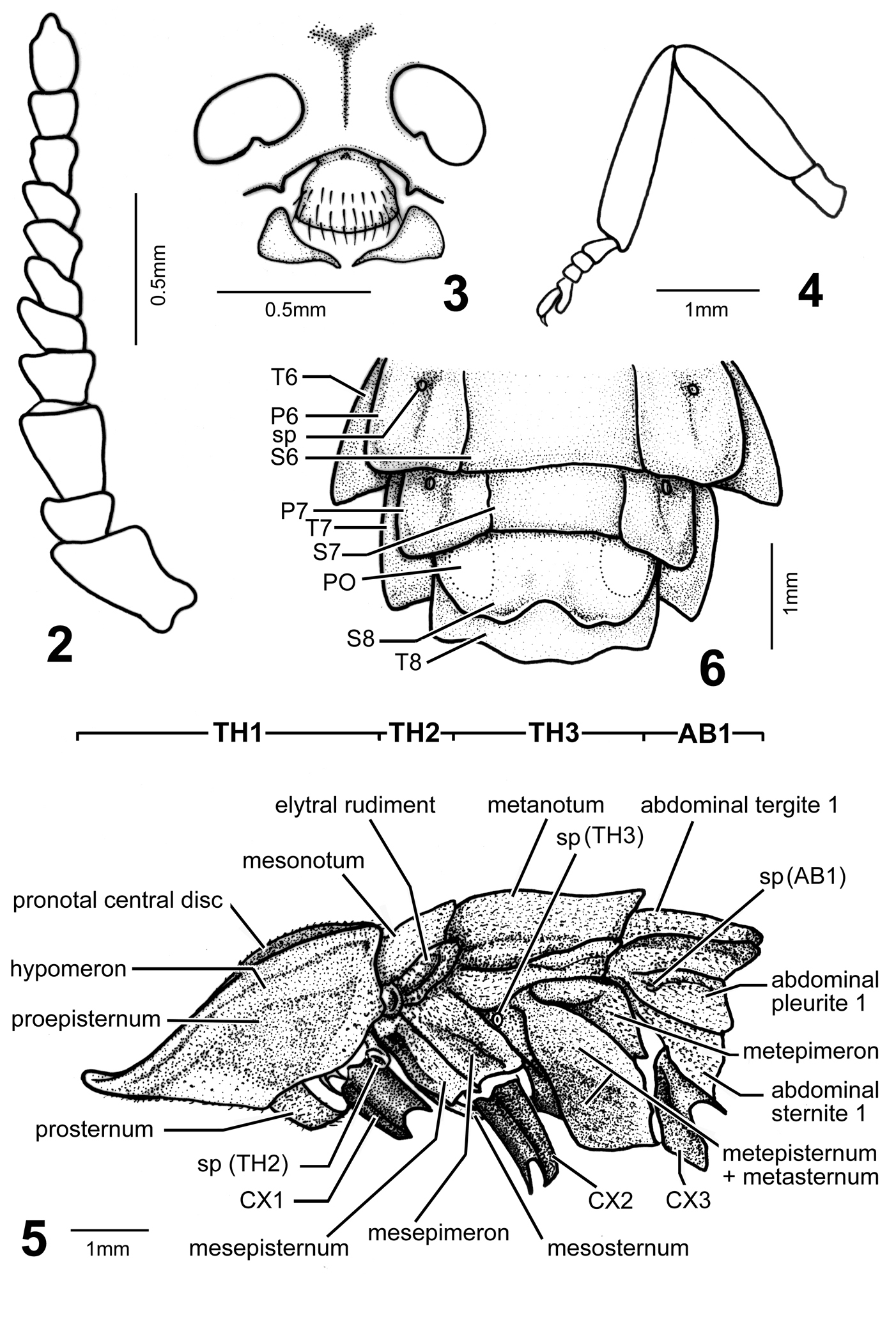
Pyrocoelia prolongata Jeng and Lai, female 2 Antenna 3 Partial head in frontal aspect (frons, antennal sockets, clypeus-labrum, and mandibles), antennae removed 4 Hind leg (metatrochanter–metatarsus) 5 Thoracic segments (TH1–3), coxae (CX1–3), and first abdominal segment (AB1), lateral aspect; sp (#) = spiracle on #th segment 6 Abdominal segments 6–8, ventral aspect, T#, P#, and S# = abdominal tergite, pleurite, and sternite of #th segment; PO = photogenic organs.
1♀, S. Taiwan: Kaohsiung County (now Kaohsiung City), Yako logging trail (abandoned), 120º57’E, 23º16’N, 2700m above sea level, 8.VII.2004 around midday, M. Satô leg.; specimen deposited in the collection of the senior author, National Museum of Natural Science, Taiwan.

Photos of female of Pyrocoelia prolongata Jeng & Lai A Dorsal aspect B Ventrolateral oblique aspect. Note the pink and bronze luster on the dorsum as well as the distinct photogenic organs (PO) on the venter.
Females differ from males by (generic level differences) 1, having shorter and less serrate antennae, much smaller compound eyes, and smaller palpi; 2, the mesoscutellum as broad as long; 3, the highly abbreviated elytra and absence of hind wings; 4, the fusion of metepisternum and metasternum; 5, having an intact abdominal S1; 6, having the hind angles of abdominal T1–7 not very acute; 7, S8 being deeply emarginate (cf.,
The female was found in a small, shady, and
damp dry streambed at the edge of a mixed forest dominant with Taiwan
Red False Cypress [Chamaecyparis formosensis Matsumura (Cupresaceae)] (Fig. 8A). It stayed on the substrate mixed of gravel and sand under a slate stone (Fig. 8B). An Alishan salamander (Hynobius arisanensis Maki) was also found in the microhabitat. Only a few species of Lampyrinae such as Diaphanes nubilus Jeng & Lai, Pyrocoelia formosana Olivier, and Pyrocoelia prolongata have ever been recorded from localities at such high elevations in Taiwan (

Habitat of Pyrocoelia prolongata Jeng & Lai female A General locality in Yako logging trail, a dry streambed at 2700 m above sea level, southern Taiwan B Close-up, indicating where the female was discovered (arrow).
A male of Pyrocoelia formosana collected from the same locality was put together with the female in a transparent plastic container to observe their interaction. At first the male did not show interest in the female. Later it attempted to copulate with the female but failed due to the female’s resistance. They eventually copulated after one day of captivity. The male perished soon after copulation, while the female survived the following week and laid three or four eggs before she died. All eggs failed to hatch and decomposed. The female glowed in darkness via a pair of photogenic organs on abdominal sternite 8. When wagging a finger around the female’s head in a dusky environment, the female responded by powerfully raising her abdomen vertically to the body axis then laid down. It was found to be a one-to-one response after several repetitions. Apparently the female can detect the nearby moving subject and the action could be an intimidation to predators.
Key to the females of Pyrocoelia species of Taiwan
| 1 | Elytral rudiments clearly extending beyond pronotum in width | 2 |
| – | Elytral rudiments short, not or slightly extending beyond pronotum in width | 4 |
| 2 | Elytral rudiments broad, lobe-like, extending posterolaterally and covering part of metanotum | 3 |
| – | Elytral rudiments narrow, sword-like, extending laterally and not covering any part of metanotum | Pyrocoelia sanguiniventer Olivier |
| 3 | Elytral rudiments longer than broad by 1.5 times; mesoscutellum about as broad as long; pronotal windows ambiguous | Pyrocoelia analis (F.) |
| – | Elytral rudiments longer than broad by 3 times; mesoscutellum transverse; pronotal windows clear | Pyrocoelia praetexta Olivier |
| 4 | Abdomen without morphologically recognizable photogenic organs; metanotum milky white; pronotum semicircular, translucent gray in explanate area | Pyrocoelia formosana Olivier |
| – | Abdomen with a pair of photogenic organs on sternite 8; metanotum bronze; pronotum more-or-less projecting forward, dark in margins and central carina between windows | Pyrocoelia prolongata Jeng & Lai |
The photogenic organs of females Pyrocoelia vary in number and position. Among the known taxa from Japan and Taiwan, Pyrocoelia analis, Pyrocoelia praetexta, Pyrocoelia rufa Olivier, Pyrocoelia miyako Nakane, and Pyrocoelia atripennis Lewis each have two pairs of morphologically recognizable lanterns on sternites 6 and 7 (
Both sexes of Pyrocoelia prolongata
can emit green light from a pair of lanterns on sternite 8. The males
are active during the daytime to locate mates by raising their heads and
elevating the antennae in a V-shaped position, used to detect female
pheromones, as observed for many other diurnal species (
We are grateful to the late Dr. M. Satô (1937–2006) for providing us the rare female specimen of Pyrocoelia prolongata available for this study. Thanks are also due to C.-L. Li (Taiwan University, Taipei) and C.-F. Lee (Taiwan Agricultural Research Institute, Taichung) for their assistance during the collecting trip; and to H.-Y. Lee for photographs of the female of Pyrocoelia prolongata. This study was supported financially by the National Science Council of the Republic of China, grant Nr. NSC99-2621-M-002-027.