





(C) 2011 Desiree R. Robertson. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Systematics of the genus Itauara Müller, 1888 are reviewed. A generic diagnosis, illustrations, and descriptions are provided for males. The genus can be identified by several features of the male genitalia including an extremely reduced phallobase and a phallic apparatus that consists of a sclerotized dorsal sheath covering a very membranous ventral portion. A total 18 species are described as new: Itauara alexanderi sp. n.(Brazil), Itaura bidentata sp. n. (Guyana), Itaura blahniki sp. n. (Brazil) Itaura charlotta sp. n. (Brazil), Itaura emilia sp. n. (Brazil), Itaura flinti sp. n. (Brazil), Itaura guyanensis sp. n. (Guyana), Itaura jamesii sp. n. (Brazil), Itaura julia sp. n. (Brazil), Itaura lucinda sp. n. (Brazil), Itaura ovis sp. n. (Guyana, Venezuela), Itaura peruensis sp. n. (Peru), Itaura rodmani sp. n. (Brazil), Itaura simplex sp. n. (Brazil), Itaura spiralis sp. n. (Guyana), Itaura stella sp. n. (Brazil), Itaura tusci sp. n. (Brazil), and Itaura unidentata sp. n. (Guyana). These additions bring the total fauna of Itauara to 22 species.
Trichoptera, Glossosomatidae, Protoptilinae, systematics, taxonomy, Itauara, new species, Neotropical, male genitalia
The genus Itauara Müller, 1888, belongs to the saddle-, or tortoise-case making caddisfly family Glossosomatidae. The name Itauara
comes from the Tupi-Guarani language and roughly translates to “born
from rock, ” likely referring to glossosomatid larval cases, often
found conspicuously on the surface of submerged rocks. Itauara
larvae construct rather loose, easily deformable cases of large and
small grains of sand that are vaulted dorsally, and almost flat
ventrally (
Like other members of Protoptilinae, Itauara adults are minute, usually less than 3 mm in size. Their wings are brownish and may have a conspicuous white spot at the arculus or transverse line along the anastomosis (Fig. 1). Itauara are rather rare in occurrence; indeed, several of the new species described here are known from only a single specimen. The 4 known species of Itauara occur only in South America, with 3 of these [Itaura brasiliana (Mosely, 1939), Itaura guarani (Angrisano, 1993), and Itaura plaumanni (Flint, 1974)] endemic to southeastern Brazil and surrounding regions of Argentina and Uruguay. A fourth species, Itaura amazonica (Flint, 1971), is known from Amazonas state, Brazil.

Adult, lateral view, Itauara julia sp. n.
A recent survey of the Trichoptera of southeastern Brazil by researchers at the University of Minnesota yielded numerous new species of Itauara and impetus for this study. Dr. Oliver S. Flint, National Museum of Natural History, Smithsonian Institution, generously provided several additional undescribed species from various locales in South America for inclusion in this study. In this paper, we determine the homologies and establish standardized terminology of the male genitalic structures among species. Additionally, we re-describe and illustrate the 4 known Itauara species and describe 18 new species from southeastern Brazil, Peru, Guyana, and Venezuela, bringing the total number of species to 22. These species are here assigned to 2 species groups and listed in Table 1. Finally, we provide a key to the males of Itauara.
Itauara species groups (Trichoptera: Glossosomatidae: Protoptilinae).
| Itauara amazonica species group |
|---|
| Itauara alexanderi sp. n. |
| Itauara amazonica (Flint, 1971) |
| Itauara bidentata sp. n. |
| Itauara emilia sp. n. |
| Itauara guyanensis sp. n. |
| Itauara jamesii sp. n. |
| Itauara lucinda sp. n. |
| Itauara ovis sp. n. |
| Itauara peruensis sp. n. |
| Itauara spiralis sp. n. |
| Itauara stella sp. n. |
| Itauara unidentata sp. n. |
| Itauara brasiliana species group |
| Itauara blahniki sp. n. |
| Itauara brasiliana (Mosely, 1954) |
| Itauara charlotta sp. n. |
| Itauara flinti sp. n. |
| Itauara guarani (Angrisano, 1993) |
| Itauara julia sp. n. |
| Itauara plaumanni (Flint, 1974) |
| Itauara rodmani sp. n. |
| Itauara simplex sp. n. |
| Itauara tusci sp. n. |
To observe certain structural features of the male
genitalia, soft tissues were cleared using a lactic acid method
outlined in detail by
Pencil sketches of the male genitalia were completed using either an optical grid on a dissecting microscope, or camera lucida
(drawing tube) mounted on a compound microscope. Pencil sketches were
scanned digitally, and then placed as a template layer in Adobe
Illustrator® for final rendering. Wing preparations were digitally
photographed using a Leica EC3 digital camera mounted on an Olympus
SZX12 dissecting microscope. Digital images were then placed as a
template layer in Adobe Illustrator® for final rendering. Descriptions
of species and generation of the identification key were facilitated by
using the software package DELTA (DEscriptive Language for Taxonomy) (
Females, with similar size and coloration as males that were collected at the same time and locality, are listed as paratypes under the material examined for some species. Previous experience has shown that having presumptively associated female specimens may be useful for future associative studies. However, since there is some uncertainty of association, we have deferred descriptions of females.
Morphological terminologyMorphological terminology for male genitalia was adapted from
Types and additional material examined for this study
are deposited at the British Museum of Natural History, London, UK
(BMNH); the National Museum of Natural History, Washington, DC, USA
(NMNH), the Museo Argentino de Ciencias Naturales Bernardino Rivadavia,
Buenas Aires, Argentina (MACN), the Museu de Zoologia, Universidade
de São Paulo, São Paulo, Brazil (MZUSP), and the University of
Minnesota Insect Collection, Saint Paul, USA (UMSP). All specimens or
lot of alcohol specimens examined in this study were affixed with a
barcode label with a unique 9 digit alphanumeric code starting with the
prefix UMSP. This prefix indicates that the specimen has been databased
at UMSP, but it is not meant to imply possession by UMSP.
Specimen-level taxonomic, locality, and other information, are stored
in the University of Minnesota Insect Collection Biota Trichoptera Database using the software program Biota (
Until recently, the generic status of Itauara was uncertain.
In a recent phylogenetic analysis of the entire protoptiline subfamily, Itauara was recovered as a monophyletic group with strong support (
http://species-id.net/wiki/Itauara
The genus Itauara can be identified by features of the male genitalia. The phallic apparatus consists of a sclerotized dorsal sheath covering a very membranous ventral portion, an apparent posterior extension of the phallobase or phallicata. Rarely, the phallicata is tubular or separated from the phallobase by a membranous portion. In some species, this sclerotized dorsal sheath seems to detach from the ventral membrane apically to reveal a single dorsomesal process or spine (e.g., Itaura amazonica). Mortoniella has a similar dorsomesal process or spine, but in Mortoniella it arises internally from the phallobase, whereas in Itauara it arises dorsobasally, as an extension of the phallicata. In several species the sheath produces a dorsolateral flange-like process, although this character is not diagnostic for the genus. Another genitalic feature characteristic of Itauara is an extremely reduced phallobase. In most species, the phallobase is barely visible, consisting of a small, very lightly sclerotized or an entirely membranous structure. The genera Mastigoptila and Canoptila display similar reductions or absences of the phallobase, but can easily be separated from Itauara by other genitalic characters: Mastigoptila has an elongate, whip-like process arising from the membranes of the phallocrypt; Canoptila has highly membranous digitate parameres. When present (they have been lost in many species), the inferior appendages are rather distinct for Itauara, consisting of a single or apically bifid process produced mesally and fused to the phallobase ventrobasally. This inferior appendage process articulates with the base of the phallobase and in doing so, is capable of pivoting downward (Fig. 13A and inset). All species, except in Itaura brasiliana, have rather elongate, sclerotized, rod-like parameres, whose shape varies greatly among species. In many species these parameres arise ventrobasally from the phallobase, with which they appear to articulate. As the inferior appendage process is absent in those species, it is possible that the parameres have taken on a clasper-like function.
The forewing venation of Itauara is most similar to that of Cariboptila and Canoptila, with apical forks I–III and a lack of 3A (Fig. 2A, B). A single species also possesses apical fork IV (Fig. 2C). Canoptila can be differentiated from Itauara by having stout setae occurring below Cu2 whereas in Itauara the setae occur along the vein. Cariboptila can be differentiated from Itauara by the presence of a short discoidal cell, that of Itauara being long. The lengths of the apical forks vary among species. The hind wing venation of Itauara is variable, with either apical forks II, III, and V (Fig. 3C); II and V (Fig. 3A); III only, or II only (Fig. 3B).
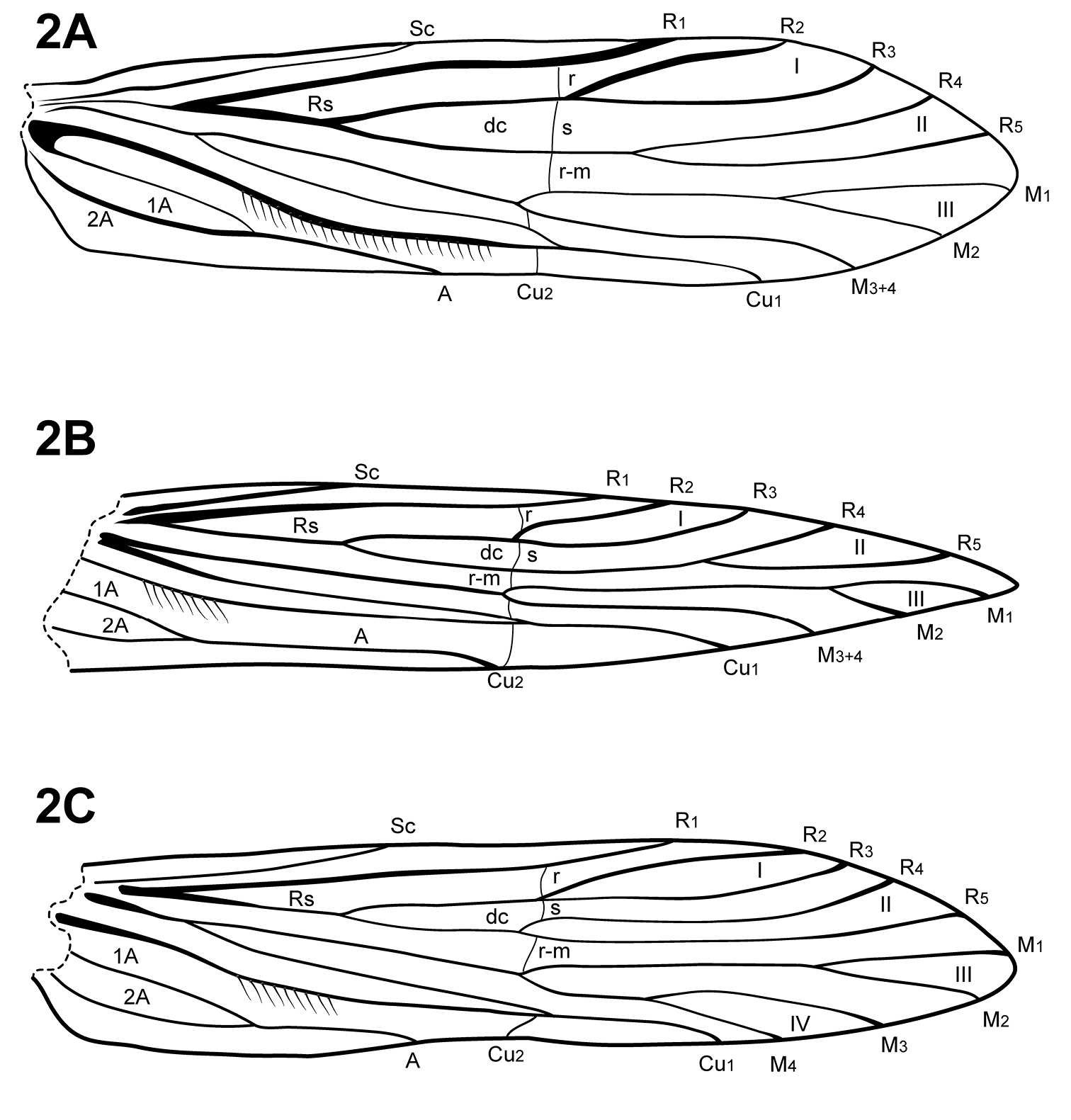
Forewings. (A) Itauara brasiliana (Mosely). (B) Itauara guyanensis sp. n. (C) Itauara unidentata sp. n. Wings between taxa not to scale.
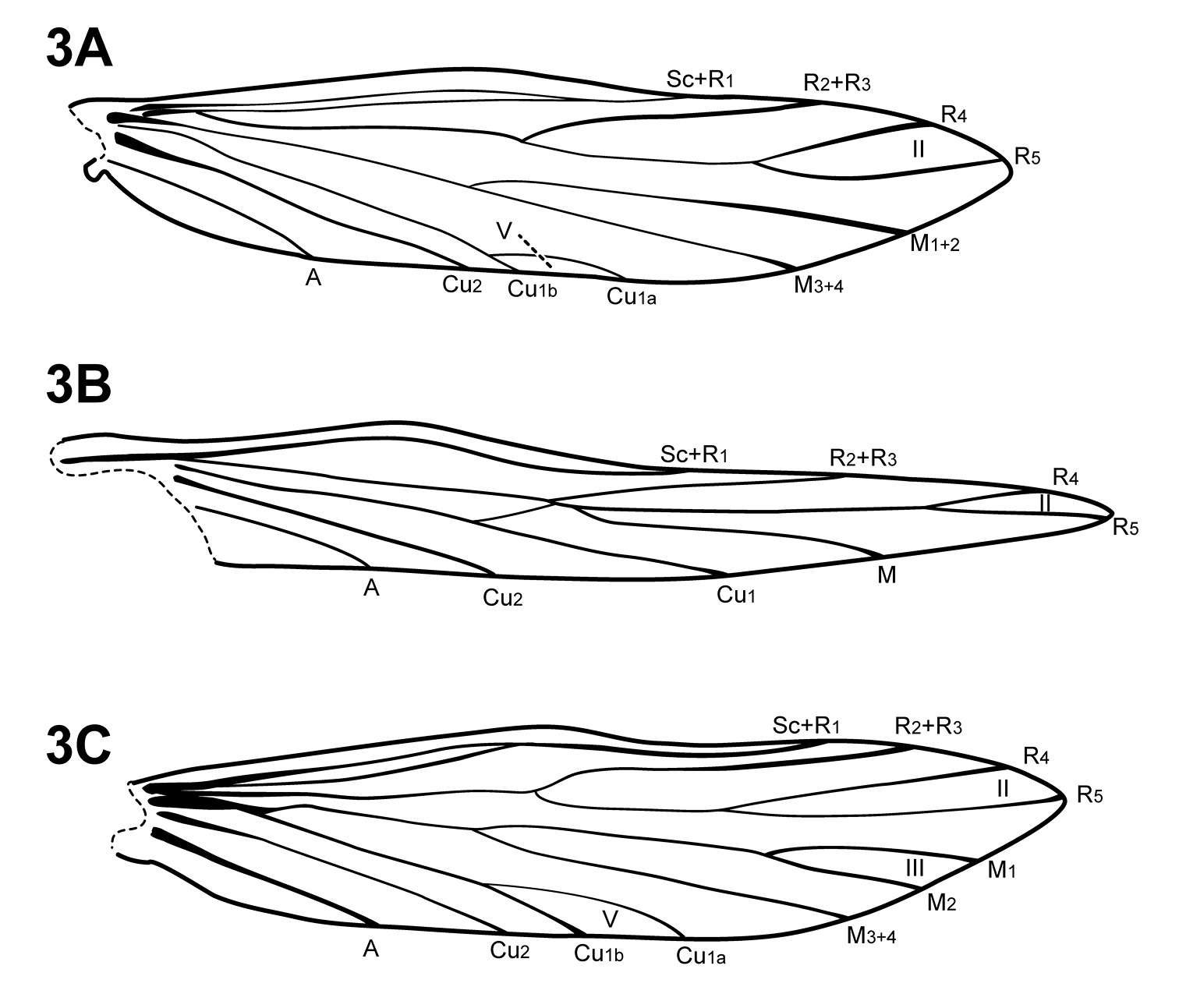
Hind wings. (A) Itauara brasiliana (Mosely). (B) Itauara guyanensis sp. n. (C) Itauara julia sp. n. Wings between taxa not to scale.
Adult. Body, wings, and appendages pale or tawny brown, often intermingled with rufous or golden hairs, tibia and tarsi yellowish brown (Fig. 1). Wings often with partial white transverse line along anastomosis not reaching costal margin, or often with conspicuous white spot at the arculus (Fig. 1). Head broader than long, vertex rounded, with pair of small anteromesal setal warts or with large anteromesal setal wart, either 1 distinct pair or 1 divided pair of suboval anterior setal warts, small or large suboval posterior warts, suboval or triangular and bulging posterolateral setal warts. Ocelli present. Antennal scape less than or equal to 2 times the length of pedicel. Maxillary palps 5 segmented, 1st and 2nd segments short; 2nd segment bulbous; last 3 segments each nearly same length as 1st and 2nd segments combined. Prothorax with 2 large subtriangular or suboval pronotal setal warts. Mesothorax wider than long, without apparent tegular glands; mesoscutum with pair of suboval anteromesal setal warts, suboval posterolateral warts; mesoscutellum sparsely setose, without distinct setal warts. Forewing (Fig. 2) usually relatively narrow, with margins nearly parallel, occasionally narrowed past anastomosis or much reduced, apex acute, subacute, or rounded. Male occasionally with callosity present in apical costal region of forewing. Forewing venation incomplete, with apical forks I, II, and III present, or rarely I–IV present; Sc and R1 distinct along their entire lengths; fork I sessile or only slightly petiolate with extremely short stem; fork II petiolate or sessile, when petiolate, stem length variable; fork III petiolate, stem variable in length; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present along Cu2; A3 absent; crossveins forming a relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing (Fig. 3) margins nearly parallel, tapering only slightly past anastomosis, or narrowed, scalloped past anastomosis, or much reduced; venation variable, either with apical forks II, III, and V present, II and V present, III present, or II present; Sc and R1 fused basally or converging near wing margin; A2 absent. Tibial spurs 1, 4, 4, rarely 1, 3, 4, foretibial spur extremely reduced and hairlike. Sixth sternal process present, short and digitate or thumb-like and prominent, apex rounded or attenuate and pointed, usually associated with oblique apodeme posteriorly.
Male genitalia. Segment IX usually rather broad, anterior margin rounded, posterolateral margin without lateral process or lobes in lateral view; tergum IX usually not well developed, simple, and without processes; sternum IX without modification, except in Itaura brasiliana, which bears 2 pairs of elongate, seta-like processes. Tergum X incompletely fused to tergum IX ventrolaterally or rarely (Itaura amazonica) completely fused and indistinct from tergum IX, shape extremely variable; dorsomesal margin may be simple without processes, bifid apicomesally, with a single broad, plate-like process, or irregular with several small processes; dorsolateral margin either a simple structure without processes, or more commonly with small paired lobes, elongate, down-turned, finger-like process, or irregular setose processes; ventrolateral margin with paired elongate or broad flange-like processes directed ventrally and sometimes anteriorly, or with one or more irregular, paired, setose, digitate lobes directed posteriorly. Inferior appendages either present or absent; when present, consisting of single or apically bifid process produced mesally, broadest at base and fused to phallobase ventrobasally. Parameres present except in Itaura brasiliana, arising either ventrobasally from phallobase or laterally from endotheca, sclerotized, shape variable. Phallobase extremely reduced and difficult to discern. Phallicata a sclerotized dorsal sheath covering membranous ventral portion, sometimes receding to a single dorsomesal process arising dorsobasally from phallobase, phallicata occasionally with dorsolateral flange, or occasionally with dorsomesal spine arising posteriorly to phallobase. Endophallus highly membranous, enlarged and convoluted when evaginated, occasionally bearing apical spine-like sclerites and processes.
Female genitalia. (Females unknown for many species.) Truncate posteriorly, not extensible. Abdominal segment VIII short, synscleritous, posterolateral margin slightly incised. Segments IX and X closely associated, with pair of small digitate cerci dorsolaterally.
The 22 species of Itauara fall into 2 broad species groups (Table 1). Members of the amazonica species group are recognized by the presence of a fused inferior appendage process. The group is also characterized by the position of the parameres, arising laterally from the endotheca in this group. Species included in the brasiliana species group have completely lost the inferior appendages. When parameres are present (they are vestigial in Itaura brasiliana) they arise ventrobasally from the phallobase, to which they are often fused. Several members of the brasiliana group also have lateral flange-like processes on the phallicata.
Species descriptionsurn:lsid:zoobank.org:act:55610B1E-3E37-4B64-9380-5C2A86CD3EA1
http://species-id.net/wiki/Itauara_alexanderi
Fig. 4A–CThis species is similar to Itauara emilia sp. n., Itaura lucinda sp. n., and Itaura stella sp. n., as discussed under each of those species. Each of these species possesses an inferior appendage process, a dorsomesal process on tergum X, and rather sinuous parameres. Of these species, Itaura alexanderi is most similar to Itaura stella. Both of these species have similarly shaped dorsomesal and ventrolateral processes of tergum X and both have apically bifid inferior appendage processes. Itauara alexanderi can be distinguished from Itaura stella by the length of the parameres, those of Itaura alexanderi being much shorter. Additionally, the inferior appendage process of Itaura alexanderi is broader than that of Itaura stella. Itauara lucinda differs from Itaura alexanderi in having a forked paramere and an inferior appendage process that is not bifid. Itauara alexanderi can be distinguished from Itaura emilia based on differences in the shape of the dorsomesal process of tergum X.
Adult. Body, wings, and appendages fuscous, intermingled with rufous or golden hairs, tibia and tarsi tawny brown. Wings with white transverse line along anastomosis. Forewing slightly broader past anastomosis, but with margins nearly parallel, apex rounded. Forewing venation incomplete, with apical forks I, II, and III present; Sc and R1 distinct along their entire lengths; fork I sessile; fork II petiolate, stem about the same length as fork; fork III petiolate, stem longer than fork; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present along Cu2; A3 absent; crossveins forming a relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing margins nearly parallel, tapering only slightly past anastomosis; apical forks II and V present; Sc and R1 fused basally; A2 absent. Tibial spurs 1, 4, 4, foretibial spur extremely reduced and hairlike. Sixth sternal process thumb-like, apex rounded, associated with oblique apodeme posteriorly.
Male genitalia. Preanal appendages absent. Segment IX ventrally narrow, broad medially; anterior margin rounded; posterolateral margin membranous or very lightly sclerotized; sternum IX without modification. Tergum X incompletely fused to tergum IX with membrane or lightly sclerotized region ventrolaterally; dorsomesal margin with single, downturned, elongate process; dorsolateral margin without processes; ventrolateral margin with paired, broad flange-like setose process consisting of upper subtriangular lobe and lower subquadrate lobe. Inferior appendages present as apically bifid, setose process produced mesally, broadest at base and fused to phallobase ventrobasally, with 2 pairs of small digitate lobes ventrolaterally, each bearing a seta. Parameres present, paired, inserted in membranous lobe, arising laterally from endotheca, sclerotized and rod-like, relatively short, sinuous, directed ventrolaterally, apex pointed. Phallobase reduced, lightly sclerotized. Phallicata forming a long sclerotized dorsal sheath extending from phallobase, narrow and straight mesally, distal portion broad, curving dorsally. Endophallus membranous, enlarged and convoluted when invaginated, with 1 upper and 1 lower lobe.

Itauara alexanderi sp. n. (composite of UMSP000114626 & UMSP000052590). Male genitalia: (A) lateral; (B) dorsal; (C) ventral. Abbreviations: dm. pr. = dorsomesal process; enph. = endophallus; inf. ap. = inferior appendage process; phb = phallobase; phc. = phallicata; pmr . = paramere; t. X = tergum X; vl. pr. = ventrolateral process.
Holotype male: BRAZIL: Nova Friburgo, 22°16'00"S, 042°31'59"W, 950 m, 20.iv.1977 (C. & O. Flint) (UMSP000052592) (NMNH)
Paratypes: BRAZIL: Rio de Janeiro, Teresopolis, 18 km S, Km 17 (road), 1180 m, 18–19.iv.1977 (C. & O. Flint) — 2 males (NMNH).
We are delighted to name this species for the senior author’s husband, Alexander Bishop Thompson, in gratitude of his patience, support, and encouragement as she worked to finish her dissertation.
http://species-id.net/wiki/Itauara_amazonica
Fig. 5A–DThis species is distinct in having a very elongate inferior appendage process, a rather simple tergum X, and sharply bent apical spines in the endophallus. Itauara amazonica is most similar to 3 species from Guyana, Itaura bidentata sp. n., Itaura spiralis sp. n., and Itaura unidentata sp. n. These species, including Itaura amazonica, all have a dorsomesal spine arising from the phallicata. However, in Itaura amazonica, this spine appears as a short, posterior extension of the phallicata, whereas in the other species, the spine arises basally, as a separate spine. Itaura amazonica can further be distinguished from these species based on differences in the shape of tergum X and parameres.
Adult. Body, wings, and appendages pale or tawny brown in alcohol. Forewing relatively narrow, with margins nearly parallel, apex subacute. Forewing venation incomplete, with apical forks I, II, and III present; Sc and R1 distinct along their entire lengths; fork I sessile; fork II petiolate, stem shorter than fork; fork III petiolate, stem longer than fork; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present along Cu2; A3 absent; crossveins forming a relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing narrow and slightly scalloped past anastomosis; apical fork III present; Sc and R1 fused basally; A2 absent. Tibial spurs 1, 4, 4, foretibial spur extremely reduced and hairlike. Sixth sternal process short and digitate, apex attenuate and pointed, associated with weak oblique apodeme posteriorly.
Male genitalia. Preanal appendages absent. Segment IX relatively broad; anterior margin rounded; posterolateral margin membranous or very lightly sclerotized; sternum IX without modification. Tergum X completely fused to tergum IX, divided or bifid apicomesally, each half terminating in pointed process directed posteriorly; dorsolateral margin without processes; ventrolateral margin without processes. Inferior appendages present as single, elongate setose process produced mesally, broadest at base and fused to phallobase ventrobasally. Parameres present, paired, inserted in membranous lobe, arising laterally from endotheca, sclerotized and rod-like, slender and elongate, upturned, directed dorsally, apex pointed. Phallobase reduced, lightly sclerotized. Phallicata forming a long sclerotized dorsal sheath extending from phallobase, bent sharply upward at middle, divided apicomesally, terminating in 2 pointed processes. Endophallus membranous, enlarged and convoluted when invaginated, receding anterior to apex of phallicata, ventrally bearing a pair of broad, tooth-like downturned processes, apically with pair of sharply bent sclerotized spines, pointing anteriorly.
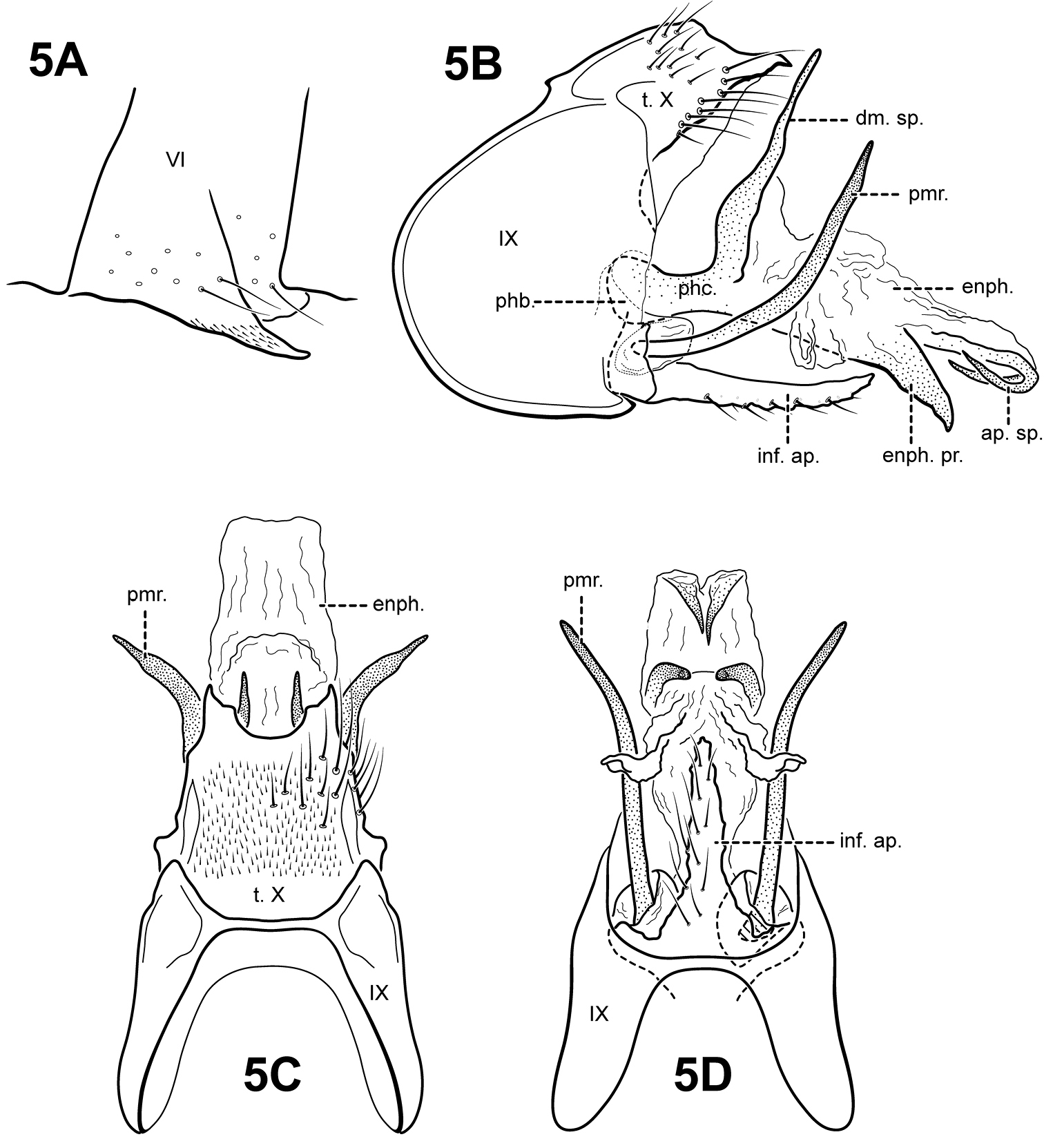
Itauara amazonica (Flint, 1971) (UMSP000027156). (A) Process of sternum VI. Male genitalia: (B) lateral; (C) dorsal; (D) ventral. Abbreviations: ap. sp. = apical spine; dm. sp. = dorsomesal spine; enph. = endophallus; enph. pr. = endophallic process; inf. ap. = inferior appendage process; phb = phallobase; phc. = phallicata; pmr . = paramere; t. X = tergum X.
Holotype male: BRAZIL: Amazonas: Rio Marauia, Endstation langer Cachoeira, Fluß tritt hier aus dem Gebirge mit starkem Gefälle, 00°23'00"N, 065°13'00"W, 28.i.1963 (E.J. Fittkau) (UMSP000027159) (NMNH).
Paratypes: BRAZIL: Amazonas: same data as holotype — 3 males, 2 females (NMNH).
urn:lsid:zoobank.org:act:76A03637-7788-4644-975B-4AC7F58E37EA
http://species-id.net/wiki/Itauara_bidentata
Fig. 6A–CItauara bidentata can be diagnosed by its large, bifid paramere process, and spade-like shaped inferior appendage. It is most similar to Itaura unidentata sp. n., which has a similarly shaped tergum X, dorsomesal spine, and apical sclerites. The 2 species can be separated by their paramere processes; in Itaura unidentata the paramere consists of a single large tooth-like spine, whereas in Itaura bidentata, the paramere process is bifid. Itauara spiralis sp. n., has a similarly shaped tergum X, but is easily distinguished from Itaura bidentata by differences in the shape of the inferior appendage process, parameres, and phallicata.
Adult. Body, wings, and appendages pale or tawny brown in alcohol. Forewing relatively narrow, with margins nearly parallel, apex subacute. Forewing venation incomplete, with apical forks I, II, and III present; fork I sessile; fork II petiolate, stem about the same length as fork; fork III petiolate, stem longer than fork; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present along Cu2; A3 absent; crossveins forming a relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing narrow and slightly scalloped past anastomosis; apical fork II present; Sc and R1 fused basally; A2 absent. Tibial spurs 1, 4, 4, foretibial spur extremely reduced and hairlike. Sixth sternal process short and digitate, apex attenuate and pointed, associated with strong oblique apodeme posteriorly.
Male genitalia. Preanal appendages absent. Segment IX ventrally narrow, broad medially; anterior margin rounded; posterolateral margin membranous or very lightly sclerotized; sternum IX without modification. Tergum X incompletely fused to tergum IX with membrane or lightly sclerotized region ventrolaterally; dorsomesal margin straight, without processes; dorsolateral margin with paired elongate, down-turned, finger-like process; ventrolateral margin with paired, broad flange-like setose process consisting of several small irregular lobes. Inferior appendages present as single, broad, irregular setose process, broadest basally, fused to phallobase ventrobasally, bearing a single pair of small digitate lobes ventrolaterally, each bearing a seta. Parameres present, paired, arising laterally from endotheca, strongly sclerotized, large bifid tooth-like process, curving ventrally and outward, apices pointed. Phallobase reduced, lightly sclerotized dorsally, laterally membranous, with 2 irregular and elongate sclerites arising basolaterally. Phallicata forming a short slerotized dorsal sheath with an elongate dorsomesal spine arising posteriorly to phallobase. Endophallus membranous, enlarged and convoluted when invaginated, apically bearing 3 small sclerotized spines.
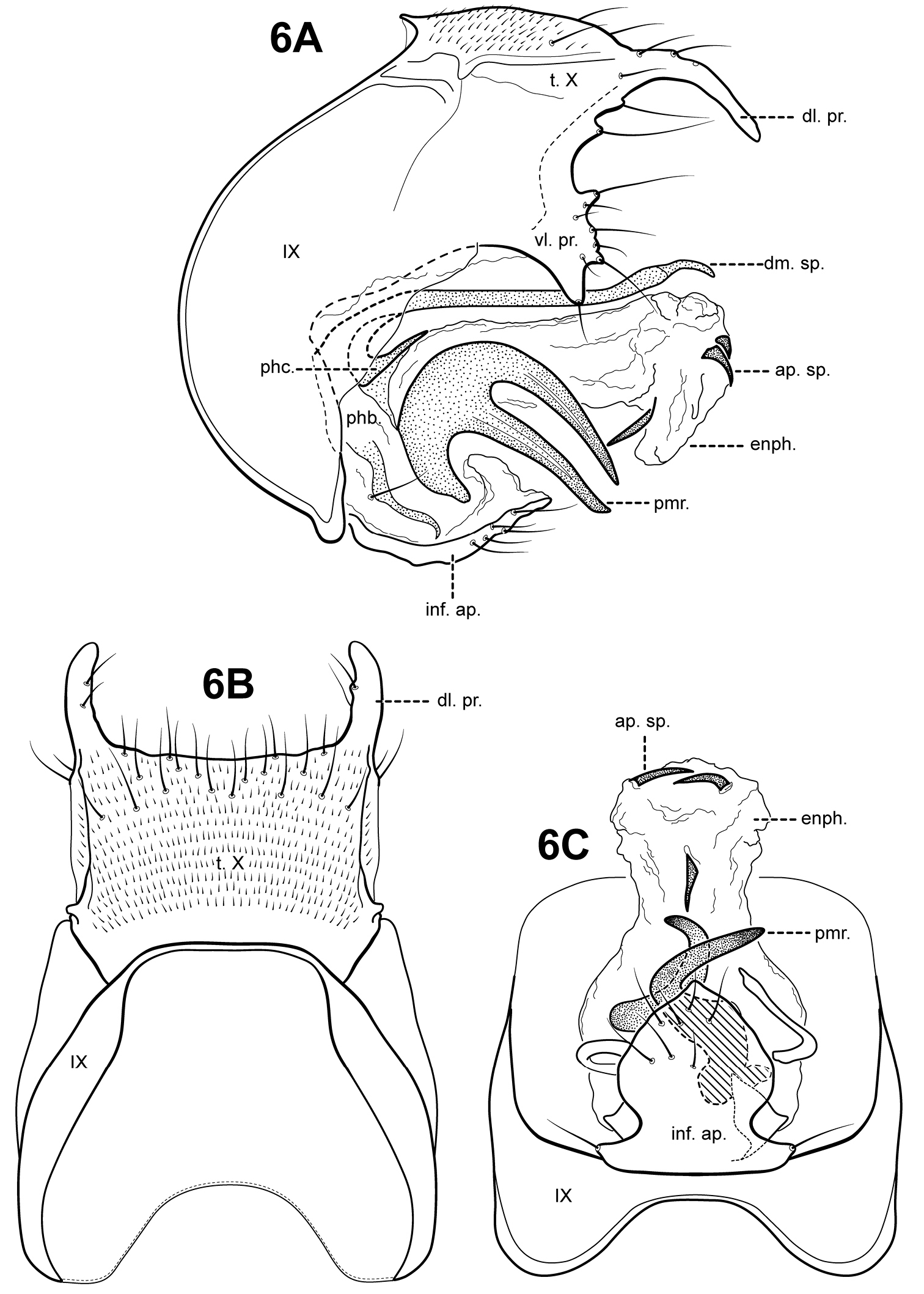
Itauara bidentata sp. n. (UMSP000210958). Male genitalia (A) lateral; (B) dorsal; (C) ventral. Abbreviations: ap. sp. = apical spine; dl. pr. = dorsolateral process; dm. sp. = dorsomesal spine; enph. = endophallus; inf. ap. = inferior appendage process; phb. = phallobase; phc. = phallicata; pmr . = paramere; t. X = tergum X; vl. pr. = ventrolateral process.
Holotype male: GUYANA: KUMU: 25 km. SE Lethem, 03°15'54"N, 059°43'36"W, 4–5.iv.1994 (O.S. Flint) (UMSP000127385) (NMNH)
Paratypes: GUYANA: Kumu: same data as holotype — 1 male, 3 females.
The name bidentata comes from the bidentate form of the paramere process.
urn:lsid:zoobank.org:act:2ED84A59-BC4E-4140-843D-DC11FFE4B8E6
http://species-id.net/wiki/Itauara_blahniki
Fig. 7A–CItauara blahniki can be recognized by the shape of the phallicata, which has a pair of very short spines dorsobasally, and a sclerotized lobe ventrobasally. The ventrolateral process of tergum X is also distinct, consisting of a an outwardly projecting flange-like setose process, and an inner, small digitate setose process. Itauara blahniki has elongate, tusk-like parameres. Itauara rodmani sp. n., and Itaura tusci sp. n., also have tusk-like parameres, but unlike Itaura blahniki, these species have flange-like lateral processes on the phallicata. The species also have differently shaped terga X.
Adult. Body, wings, and appendages pale or tawny brown, often intermingled with rufous or golden hairs, tibia and tarsi tawny brown. Wings with conspicuous white spot at the arculus. Forewing slightly broader past anastomosis, but with margins nearly parallel, apex rounded. Forewing venation incomplete, with apical forks I, II, and III present; Sc and R1 distinct along their entire lengths; fork I sessile; fork II sessile; fork III petiolate, stem about the same length as fork; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present along Cu2; A3 absent; crossveins forming a relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing margins nearly parallel, tapering only slightly past anastomosis; apical forks II, III, and V present; Sc and R1 fused basally; A2 absent. Tibial spurs 1, 4, 4, foretibial spur extremely reduced and hairlike. Sixth sternal process thumb-like, apex rounded, associated with oblique apodeme posteriorly.
Male genitalia. Preanal and inferior appendages absent. Segment IX dorsally and ventrally narrow, broad medially; anterior margin rounded; posterolateral margin membranous or very lightly sclerotized; sternum IX without modification. Tergum X incompletely fused to tergum IX with membrane or lightly sclerotized region ventrolaterally; dorsomesal margin with single, downturned, elongate process; dorsolateral margin irregular and setose; ventrolateral margin with paired, outwardly projecting flange-like setose process, and medially with paired digitate setose process. Parameres present, paired, arising ventrobasally from fused endotheca and phallobase, sclerotized and rod-like, slender and elongate, upturned, with distal portion slightly broader, directed dorsally, apex pointed, ventrobasally with small patch of setae. Phallobase reduced, mostly membranous, ventromesally bearing pair of small sclerotized spines. Phallicata forming a long sclerotized dorsal sheath, curving upward, dorsobasally with pair of short processes, ventrally, with lightly sclerotized lobe. Endophallus membranous, enlarged and convoluted when invaginated, with lightly sclerotized lobe ventrally.
Holotype male: BRAZIL: Sao Paulo: Estação Biológica Boraceia, Rio Guaratuba, 23°40'02"S, 045°53'46"W, 775 m, 17.ix.2002 (Blahnik, Prather, Melo, Froehlich, Silva) (UMSP000087057) (MZUSP).
Paratypes: BRAZIL: Sao Paulo: same data as holotype except 17.iv.1998 (Holzenthal, Melo, Froehlich) — 1 male (UMSP); same data as holotype — 1 female (UMSP).
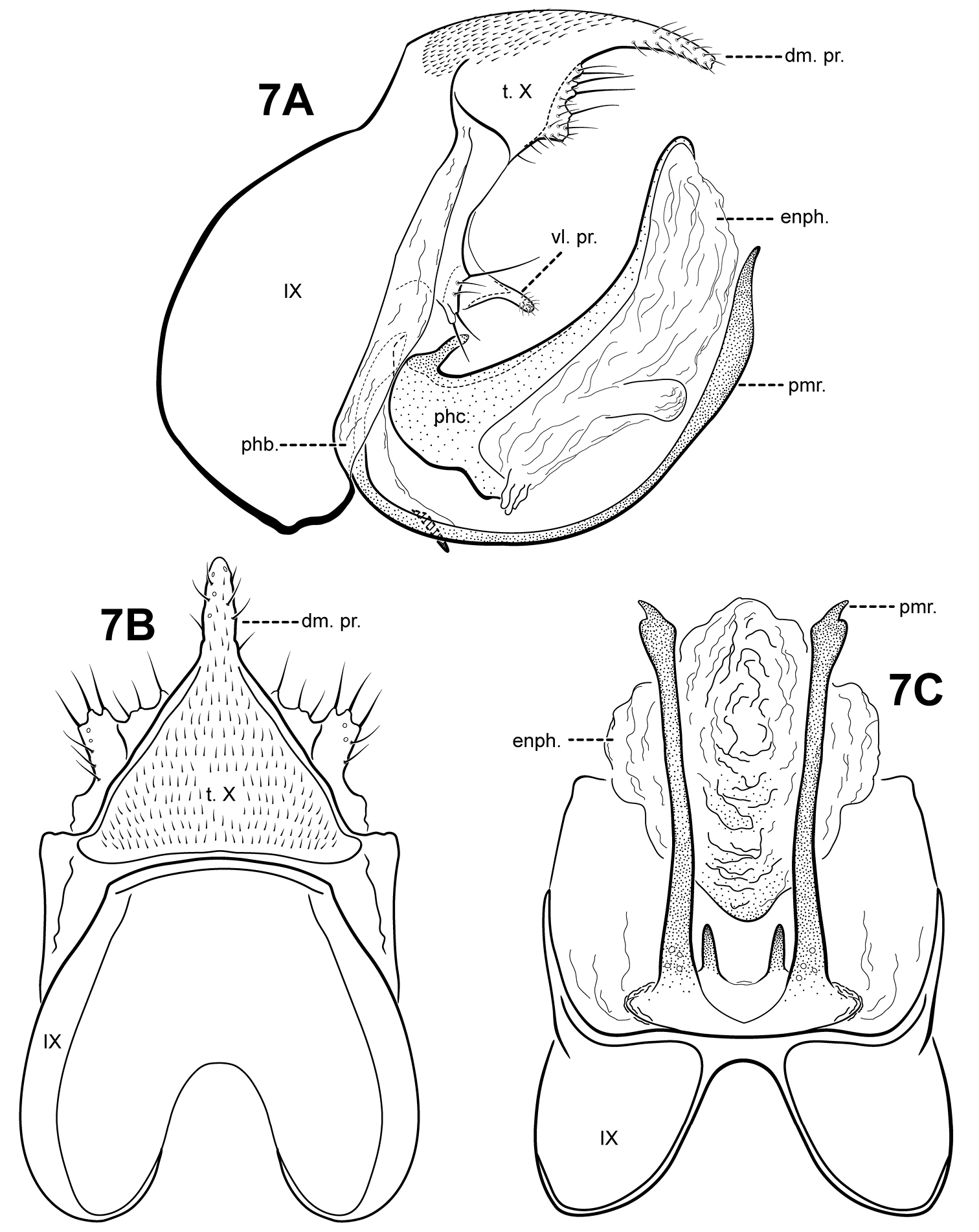
Itauara blahniki sp. n. (UMSP000087057) Male genitalia: (A) lateral; (B) dorsal; (C) ventral. Abbreviations: dm. pr. = dorsomesal process; enph. = endophallus; phb = phallobase; phc. = phallicata; pmr . = paramere; t. X = tergum X; vl. pr. = ventrolateral process.
We are delighted to name this species for Dr. Roger Blahnik, in honor of his many contributions to the systematics of Protoptilinae.
http://species-id.net/wiki/Itauara_brasiliana
Fig. 2A, 3A, 8A–C This species is easily diagnosed by the presence of 2 pairs of extremely elongate, seta-like processes on sternum IX. Itauara brasiliana
is also distinct in having vestigial parameres, consisting only of
very small, setose lobes. Another distinguishing characteristic is the
shape of segment IX, which is rather narrow, and receded ventrally.
The rather elongate profile of tergum X, as well as the shape of the
dorsomesal and lateral margins of tergum X, slightly resembles that of Itaura plaumanni (
Adult. Body, wings, and appendages pale or tawny brown in alcohol. Forewing slightly broader past anastomosis, but with margins nearly parallel, apex rounded. Forewing venation incomplete, with apical forks I, II, and III present; Sc and R1 distinct along their entire lengths; fork I sessile; fork II petiolate, stem shorter than fork; fork III petiolate, stem about the same length as fork; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present along Cu2; A3 absent; crossveins forming a relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing margins nearly parallel, tapering only slightly past anastomosis; apical forks II and V present; Sc and R1 converging near wing margin; A2 absent. Tibial spurs 1, 4, 4, foretibial spur extremely reduced and hairlike. Sixth sternal process thumb-like, apex rounded, associated with oblique apodeme posteriorly.
Male genitalia. Preanal and inferior appendages absent. Segment IX ventrally narrow, broad medially; anterior margin relatively straight from dorsum to medial area, ventral portion rounded; posterolateral margin highly membranous, receding ventrally; sternum IX bearing 2 pairs of extremely elongate, seta-like processes. Tergum X incompletely fused to tergum IX with membrane or lightly sclerotized region ventrolaterally; dorsomesal margin subtriangular, slightly upturned; dorsolateral margin slightly irregular, without processes; ventrolateral margin with 2 pairs of processes, the upper an elongate finger-like process slightly downturned, the lower a smaller lobe-like setose process. Parameres vestigial, consisting of a pair of small, digitate setose lobes arising ventrolaterally from endotheca. Phallobase apparently absent or entirely membranous. Phallicata forming a long, lightly sclerotized dorsal sheath, sinuous, broadest medially, narrowed distally. Endophallus membranous, enlarged and convoluted when invaginated.
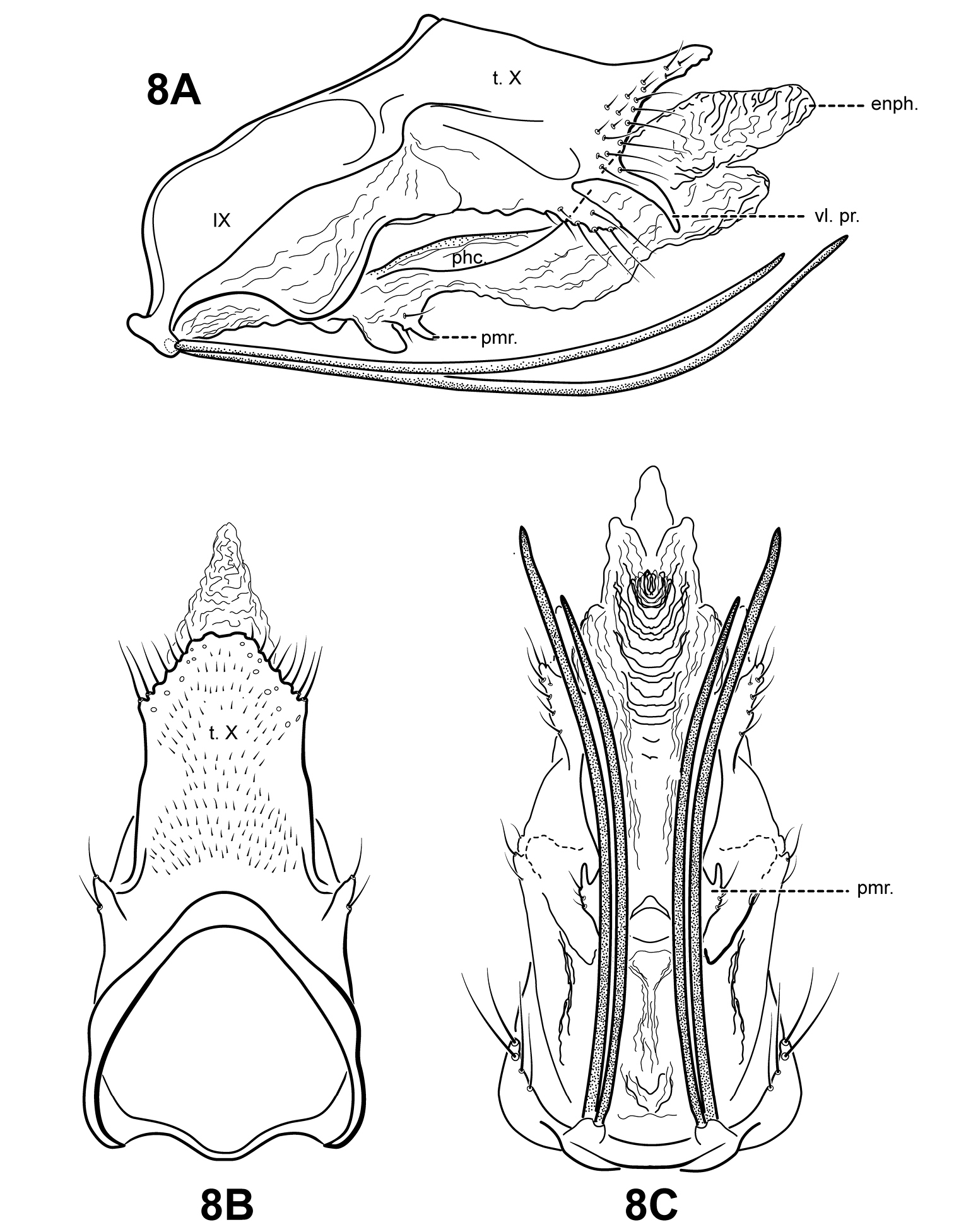
Itauara brasiliana (Mosely, 1939). Male genitalia. (A) lateral; (B) dorsal; (C) ventral. Abbreviations: enph. = endophallus; phc. = phallicata; pmr . = paramere; t. X = tergum X; vl. pr. = ventrolateral process.
Holotype male: Brazil: Santa Catarina: Nova Teutonia, ii. 1937 (F. Plaumann) (BMNH)
ARGENTINA: Misiones: Arroyo Piray Mini, W., Dos Hermanas, 23.11.1973 (O.S. Flint) — 5 males, 14 females (NMNH); BRAZIL: Santa Catarina: Seara (Nova Teutônia), 27°11'00"S, 052°23'00"W, 300–500 m, 10.1964 (F. Plaumann) — 1 male (NMNH).
urn:lsid:zoobank.org:act:EE8E2255-6A12-407B-9D1F-921D251401B7
http://species-id.net/wiki/Itauara_charlotta
Fig. 9A–CThis species is only known from the male holotype. Itauara charlotta is diagnosed by the unique combination of several male genitalic characters. The dorsomesal margin of tergum X has several small, irregular, setose processes. Itauara tusci sp. n., has a similarly shaped dorsomesal margin, but differs in the shape of the parameres and phallicata. The parameres of Itaura charlotta are similar to those of Itaura flinti sp. n.; both arise ventrobasally from the phallobase, and are curved and downturned. Itauara flinti is easily separated from Itaura charlotta based on differences in the shape of tergum X and phallicata processes.
Adult. Body, wings, and appendages pale or tawny brown in alcohol. Wings with conspicuous white spot at the arculus and faint transverse line along anastomosis. Forewing slightly broader past anastomosis, but with margins nearly parallel, apex rounded. Forewing venation incomplete, with apical forks I, II, and III present; Sc and R1 distinct along their entire lengths; fork I sessile; fork II petiolate, stem shorter than fork; fork III petiolate, stem longer than fork; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present along Cu2; A3 absent; crossveins forming a relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing margins nearly parallel, tapering only slightly past anastomosis; apical forks II, III, and V present; Sc and R1 fused basally; A2 absent. Tibial spurs 1, 4, 4, foretibial spur extremely reduced and hairlike. Sixth sternal process thumb-like, apex rounded, often associated with weak oblique apodeme posteriorly.
Male genitalia. Preanal and inferior appendages absent. Segment IX ventrally narrow, broad medially; anterior margin rounded; posterolateral margin membranous or very lightly sclerotized; sternum IX without modification. Tergum X incompletely fused to tergum IX with membrane or lightly sclerotized region ventrolaterally; dorsomesal margin slightly produced with several small irregular setose processes; dorsolateral margin with pair of large irregular, setose process and several smaller processes; ventrolateral margin without processes. Parameres present, paired, arising ventrobasally from fused endotheca and phallobase, sclerotized and rod-like, slender and elongate, slightly downturned, curved basally, straight medially and distally, directed posteriorly, apex pointed. Phallobase reduced, lightly sclerotized. Phallicata forming a long, lightly sclerotized dorsal sheath, slightly sinuous, medially with pair of slender lateral flanges projecting posteroventrally, apices pointed. Endophallus membranous, enlarged and convoluted when evaginated, with pointed apical sclerite.
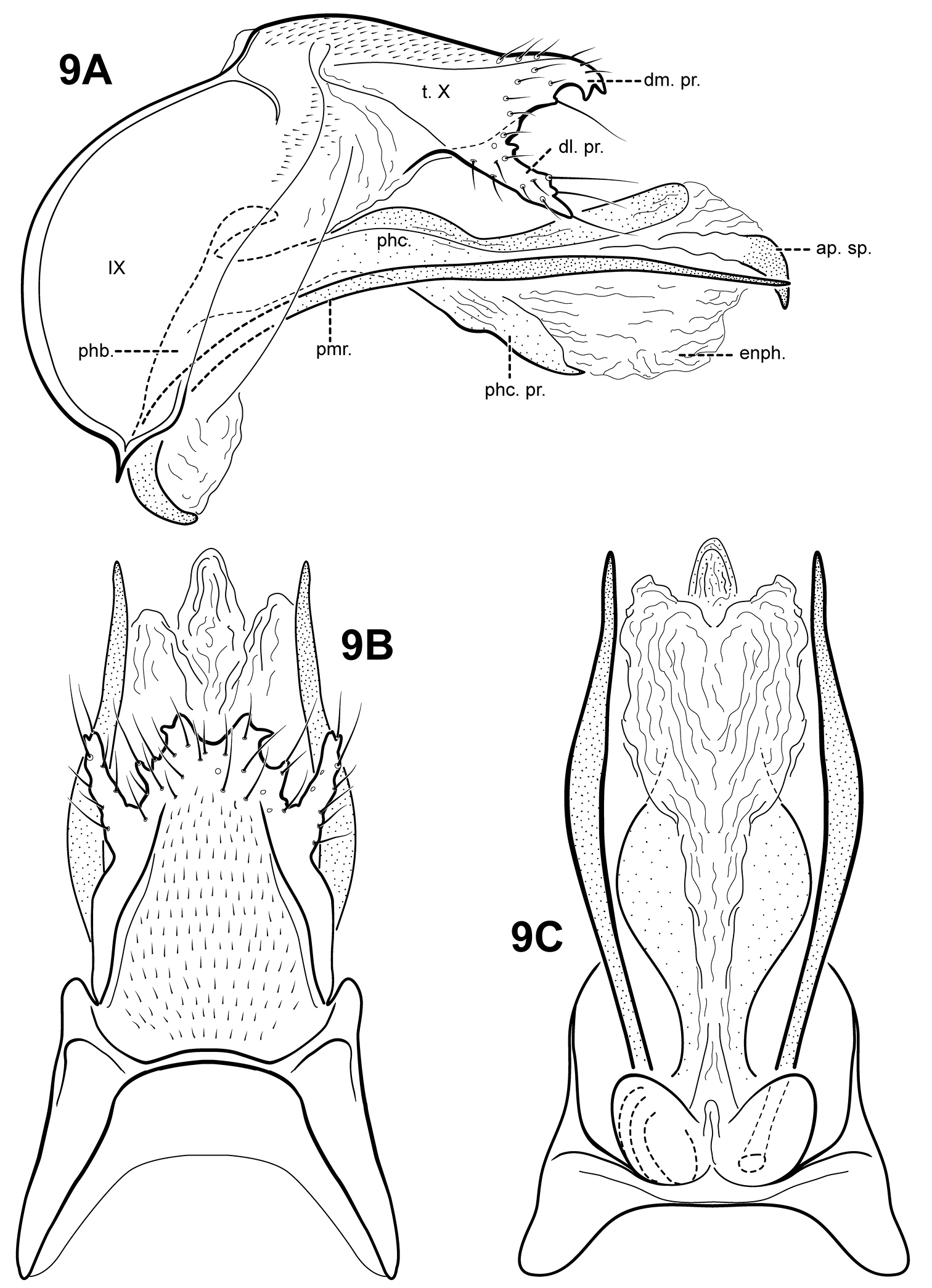
Itauara charlotta, sp. n. (UMSP000086390). Male genitalia: (A) lateral; (B) dorsal; (C) ventral. Abbreviations: ap. sp. = apical spine; dl. pr. = dorsolateral process; dm. pr. = dorsomesal process; enph. = endophallus; phb = phallobase; phc. = phallicata; phc. pr. = phallicata process; pmr . = paramere; t. X = tergum X.
Holotype male: BRAZIL: Minas Gerais: Serra do Cipó, Cardeal Mota, Cachoeira Veu da Noiva, 19°18'55"S, 043°36'16"W, 800 m, 12.11.2001 (Holzenthal, Amar., Blahnik, Paprocki) (UMSP000086390) (MZUSP).
We are delighted to name this species for the senior author’s mother, Charlotte Ruth Robertson.
urn:lsid:zoobank.org:act:EDA5DA9E-6661-4F15-96A2-B6166861CA49
http://species-id.net/wiki/Itauara_emilia
Fig. 10A–CThis species is known only from the male holotype. Itauara emilia can be recognized by the distinct, rather blunt shape of the dorsomesal process of tergum X. The species is similar to Itaura alexanderi sp. n., Itaura lucinda sp. n., and Itaura stella sp. n., as discussed under each of those species. Each of these species possess an inferior appendage process, a dorsomesal process on tergum X, and rather sinuous parameres. Itauara emilia is most similar to Itaura alexanderi and Itaura stella in having similarly shaped parameres and an apically bifid inferior appendage process. Itauara emilia differs from these 2 species in having a much more elongate inferior appendage process and a blunt dorsomesal process on tergum X.
Adult. Body, wings, and appendages tawny brown (specimen missing hairs). Forewing slightly broader past anastomosis, but with margins nearly parallel, apex rounded. Forewing venation incomplete, with apical forks I, II, and III present; Sc and R1 distinct along their entire lengths; fork I sessile; fork II petiolate, stem about the same length as fork; fork III petiolate, stem longer than fork; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present along Cu2; A3 absent; crossveins forming a relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing margins nearly parallel, tapering only slightly past anastomosis; apical forks II and V present; Sc and R1 fused basally; A2 absent. Tibial spurs 1, 4, 4, foretibial spur extremely reduced and hairlike. Sixth sternal process thumb-like, apex rounded, associated with oblique apodeme posteriorly.
Male genitalia. Preanal appendages absent. Segment IX ventrally narrow, broad medially; anterior margin rounded; posterolateral margin membranous or very lightly sclerotized; sternum IX without modification. Tergum X incompletely fused to tergum IX with membrane or lightly sclerotized region ventrolaterally; dorsomesal margin with large, blunt, dorsomesal process, in dorsal view, elongate, in lateral view, subtriangular; dorsolateral margin without processes; ventrolateral margin with paired, broad flange-like setose process consisting of small upper lobe and larger subtriangular lower lobe. Inferior appendages present as apically bifid, setose process produced mesally, broadest at base and fused to phallobase ventrobasally, with 2 pairs of small digitate lobes ventrolaterally, each bearing a seta. Parameres present, paired, inserted in membranous lobe, arising laterally from endotheca, sclerotized and rod-like, long, sinuous, directed inward and posteriorly, apex pointed. Phallobase reduced, lightly sclerotized. Phallicata forming a long sclerotized dorsal sheath extending from phallobase, straight, broadest basally, distal portion narrow. Endophallus membranous, enlarged and convoluted when invaginated, with 1 upper and 1 lower lobe.
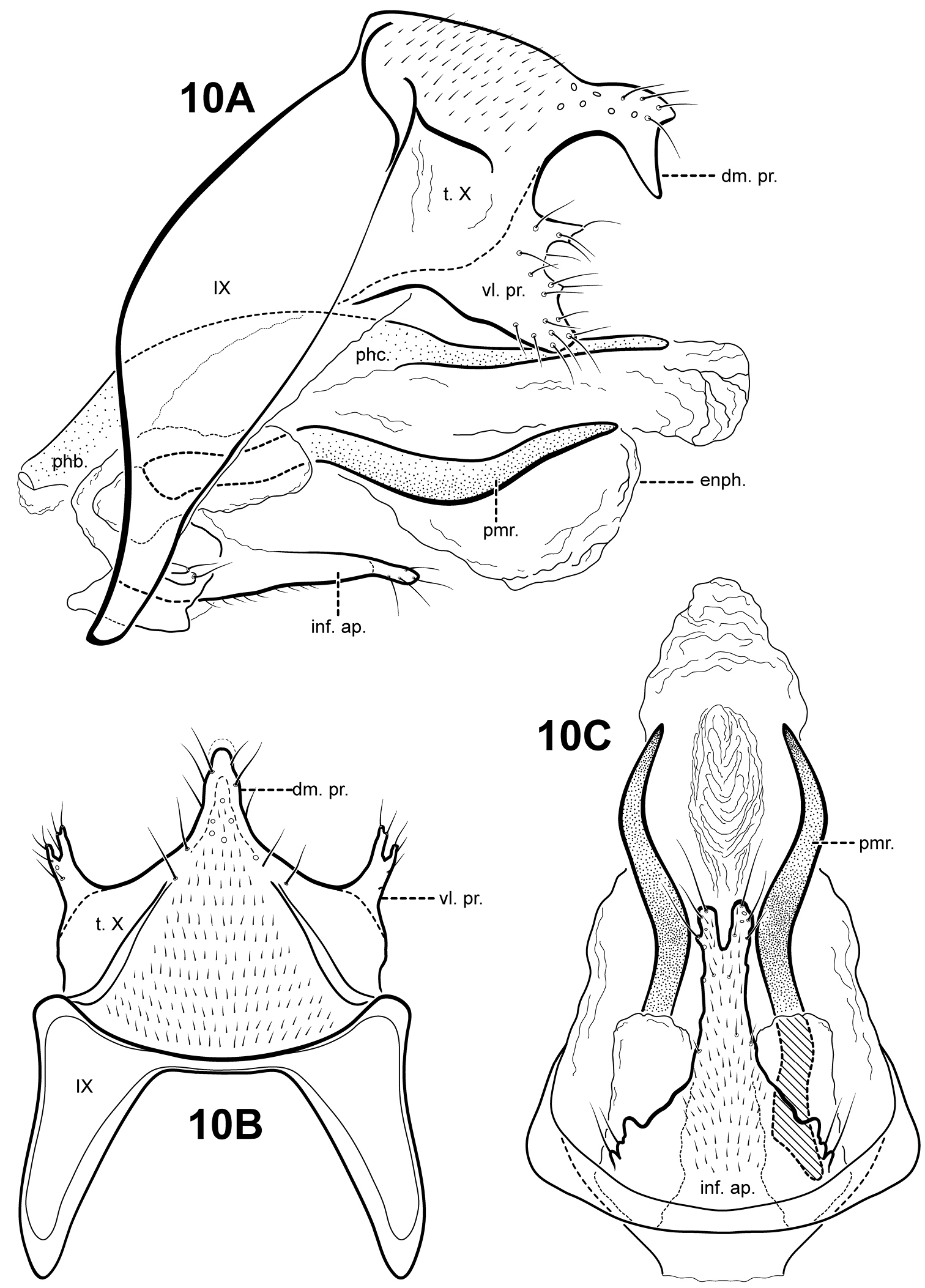
Itauara emilia (UMSP000029788). Male genitalia (A) lateral; (B) dorsal; (C) ventral. Abbreviations: dm. pr. = dorsomesal process; enph. = endophallus; inf. ap. = inferior appendage process; phb. = phallobase; phc. = phallicata; pmr . = paramere; t. X = tergum X; vl. pr. = ventrolateral process.
Holotype male: BRAZIL: Sao Paulo: Estação Biológica Boraceia, Rio Coruja, 23°40'06"S, 045°53'57"W, 850 m, 18.iv.1998 (Holzenthal, Melo, Froehlich) (UMSP000029788) (MZUSP).
This species is named in loving memory of the senior author’s paternal grandmother, Grace Emily Gardner Robertson.
urn:lsid:zoobank.org:act:08207511-83D1-46A4-8168-BAE2935AAC0A
http://species-id.net/wiki/Itauara_flinti
Fig. 11A–C This species is known only from the male
holotype. It is diagnosed by a unique combination of male genitalic
characters and can be recognized by the shape of the phallicata process.
Itauara flinti is most similar to Itaura charlotta
sp. n., by having similarly shaped parameres, but these species are
easily separated based on differences in the shape of tergum X and the
phallicata process. The elongate and downturned dorsomesal process of
tergum X is similar to that of Itaura guarani (
Adult. Body, wings, and appendages pale or tawny brown in alcohol. Forewing relatively narrow, with margins nearly parallel, apex subacute. Forewing venation incomplete, with apical forks I, II, and III present; Sc and R1 distinct along their entire lengths; fork I petiolate, but with extremely short stem; fork II petiolate, stem about the same length as fork; fork III petiolate, stem longer than fork; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present along Cu2; A3 absent; crossveins forming a relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing narrow and slightly scalloped past anastomosis; apical forks II and V present; Sc and R1 fused basally; A2 absent. Tibial spurs 1, 4, 4, foretibial spur extremely reduced and hairlike. Sixth sternal process thumb-like, apex rounded, associated with weak oblique apodeme posteriorly.
Male genitalia. Preanal and inferior appendages absent. Segment IX dorsally narrow, broad ventrally; anterior margin rounded; posterolateral margin lightly sclerotized; sternum IX without modification. Tergum X incompletely fused to tergum IX with membrane or lightly sclerotized region ventrolaterally; dorsomesal margin with single, downturned, elongate process; dorsolateral margin irregular and setose; ventrolateral margin with 2 pairs of processes, the upper a small lobe-like setose process, the lower an elongate finger-like process bearing a few elongate setae. Parameres present, paired, arising ventrobasally from fused endotheca and phallobase, sclerotized and rod-like, slender and elongate, downturned, curved basally, straight medially and distally, directed ventrally and inward, apex pointed. Phallobase reduced, lightly sclerotized. Phallicata forming a long, lightly sclerotized dorsal sheath, slightly sinuous, medially with pair of lightly sclerotized rounded lateral flanges projecting posteroventrally, ventrally with several sclerotized points, thorn-like apices directed inward. Endophallus membranous, enlarged and convoluted when invaginated, with 1 large upper lobe and 1 smaller lower lobe.
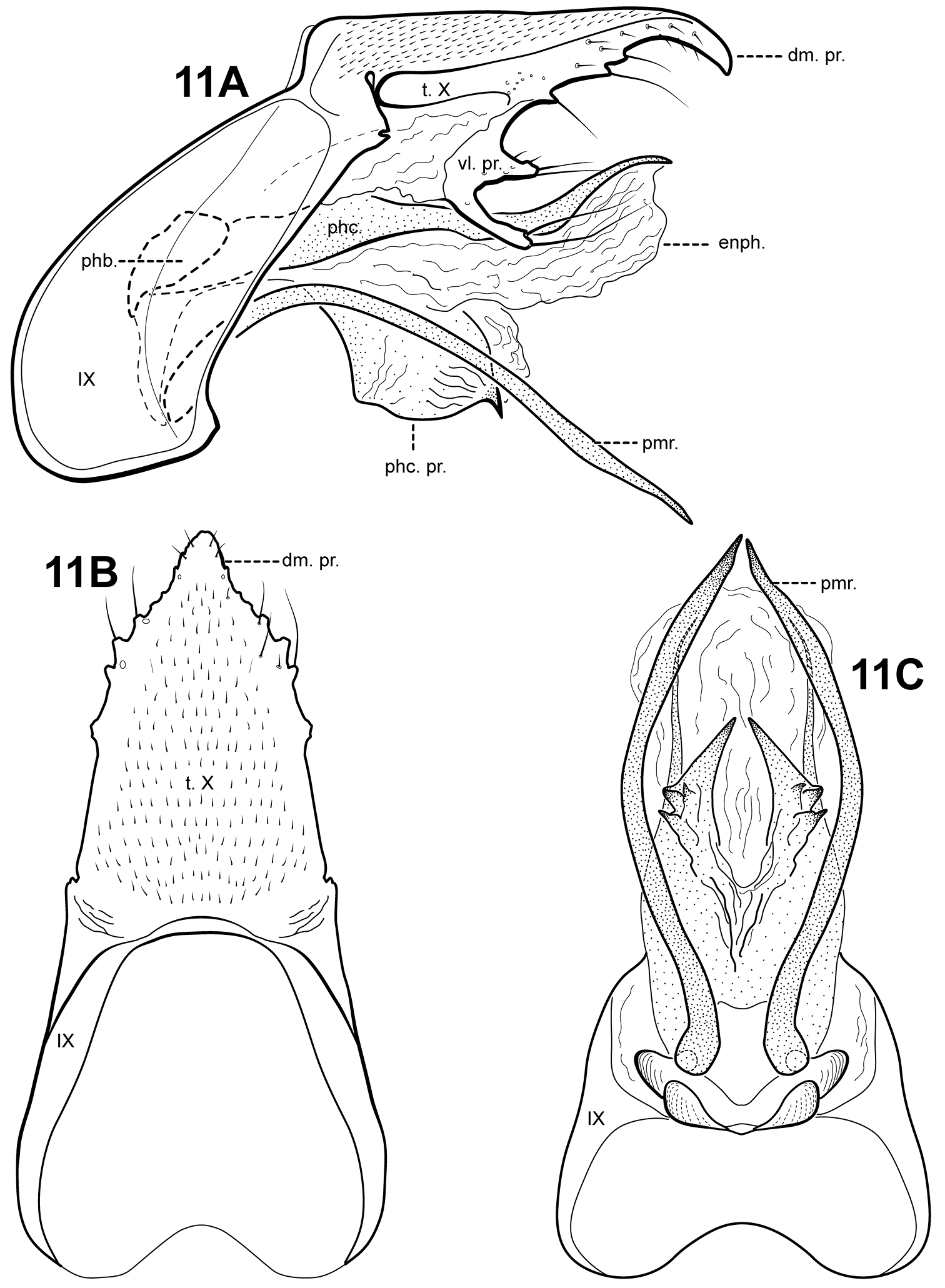
Itauara flinti sp. n. (UMSP000086388) Male genitalia: (A) lateral; (B) dorsal; (C) ventral. Abbreviations: dm. pr. = dorsomesal process; enph. = endophallus; phb = phallobase; phc. = phallicata; phc. pr. = phallicata process; pmr . = paramere; t. X = tergum X; vl. pr. = ventrolateral process.
Holotype male: BRAZIL: Sao Paulo: Parque Estadual de Campos do Jordão, Rio Galharada, 22°41'40"S, 045°27'47"W, 1530 m, 13–15.ix.2002 (Blahnik, Prather, Melo, Huamantinco) (UMSP000086388) (MZUSP).
We are delighted to name this species for Dr. Oliver Flint, Jr. in honor of his numerous important contributions to Neotropical caddisfly taxonomy.
http://species-id.net/wiki/Itauara_guarani
Fig. 12A–DThis species can be recognized by the very broad, lateral flanges on the phallicata. The phallicata of Itaura guarani is slightly sinuous, with a lightly sclerotized basal portion and a rugous or almost membranous distal portion. Itauara simplex sp. n., also has a very lightly sclerotized phallicata, but the 2 species differ in the shape of the parameres and Itaura simplex lacks the processes of phallicata. The parameres of Itaura guarani arise ventrobasally from the phallobase and are sinuous, like those in Itaura plaumanni. However, the phallicata in Itaura plaumanni is much more sclerotized and the 2 species also differ in the shape of tergum X and the phallicata processes.
Adult. Body, wings, and appendages pale or tawny brown in alcohol. Forewing slightly broader past anastomosis, but with margins nearly parallel, apex rounded. Forewing venation incomplete, with apical forks I, II, and III present; Sc and R1 distinct along their entire lengths; fork I petiolate, but with extremely short stem; fork II petiolate, stem shorter than fork; fork III petiolate, stem longer than fork; Cu1 incomplete, not reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present along Cu2; A3 absent; crossveins forming a relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing margins nearly parallel, tapering only slightly past anastomosis; apical forks II and III present; Sc and R1 fused basally; A2 absent. Tibial spurs 1, 4, 4, foretibial spur extremely reduced and hairlike. Sixth sternal process short and digitate, apex rounded, associated with weak oblique apodeme posteriorly.
Male genitalia. Preanal and inferior appendages absent. Segment IX dorsally narrow, broad ventrally; anterior margin rounded; posterolateral margin membranous or very lightly sclerotized; sternum IX without modification. Tergum X incompletely fused to tergum IX with membrane or lightly sclerotized region ventrolaterally; dorsomesal margin with single, downturned, elongate process; dorsolateral margin irregular and setose; ventrolateral margin with paired subtriangular setose process directed posteriorly. Parameres present, paired, arising ventrobasally and fused to phallobase, sclerotized and rod-like, slender and elongate, sinuous, strongly downturned basally, distal portion slightly upturned and broader, directed posteriorly, apex pointed. Phallobase reduced, lightly sclerotized. Phallicata sinuous, with lightly sclerotized base, distal portion membranous, with pair of broad, sclerotized wing-like lateral flanges. Endophallus membranous, enlarged and convoluted when invaginated, with 3 upper lobes and 1 large lower lobe.
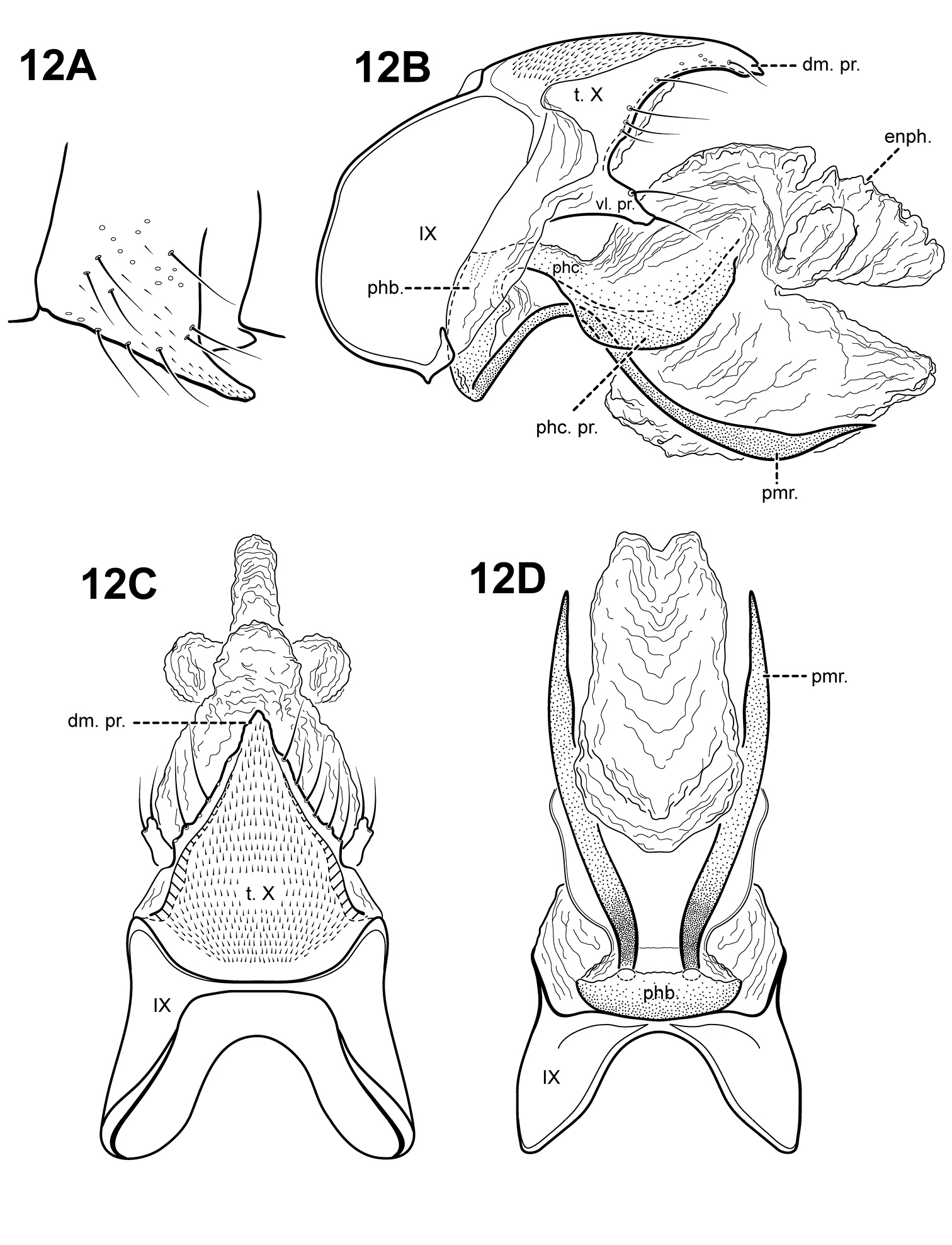
Itauara guarani (Angrisano, 1993) (UMSP000086361). Male genitalia: (A) Sternum VI process; ttt (B) lateral; (C)dorsal; (D) ventral. Abbreviations: dm. pr. = dorsomesal process; enph. = endophallus; phb = phallobase; phc. = phallicata; phc. pr. = phallicata process; pmr . = paramere; t. X = tergum X; vl. pr. = ventrolateral process.
Holotype male: ARGENTINA: Misiones: Departmento Belgrano, Rio Urugua-í (Releveamiento Faunístico Urugua-í [segundo campaña]) (UMSP000211316) (MACN).
Allotype female: ARGENTINA: Misiones: same data as holotype (MACN).
Paratypes: ARGENTINA: Misiones: same data as holotype – 3 males, 2 females (MACN).
The genitalia of the holotype and allotype were reported missing (E. Angrisano, personal communication).
urn:lsid:zoobank.org:act:3CD78294-B5E5-4191-A04E-23082102925F
http://species-id.net/wiki/Itauara_guyanensis
Fig. 2B, 3B, 13A–CItauara guyanensis has distinct extremely sinuous, almost corkscrew-shaped, parameres. The phallicata is short, sclerotized, and upturned apically and with 2 pointed lateral processes. The species can also be recognized by the thumb-like shape of the inferior appendage process and the presence of a bifid apical process in the endophallus. Itaura guyanensis is somewhat similar to Itaura jamesii sp. n., and resembles that species in the shape of the inferior appendage process and sinuous parameres. The 2 species can be easily separated by differences in the shape of tergum X and the phallicata. The species Itaura alexanderi sp. n., Itaura emilia sp. n., and Itaura stella sp. n., also have rather sinuous parameres, but differ in the shape of the phallicata, tergum X, and several other characters.
Adult. Body, wings, and appendages pale or tawny brown in alcohol. Forewing narrow past anastomosis, apex acute. Forewing venation incomplete, with apical forks I, II, and III present; Sc and R1 distinct along their entire lengths; fork I sessile; fork II petiolate, stem about the same length as fork; fork III petiolate, stem longer than fork; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present along Cu2; A3 absent; crossveins forming a relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing narrow and slightly scalloped past anastomosis; apical fork II present; Sc and R1 fused basally; A2 absent. Tibial spurs 1, 3, 4, foretibial spur extremely reduced and hairlike. Sixth sternal process short and digitate, apex attenuate and pointed, associated with strong oblique apodeme posteriorly.
Male genitalia. Preanal appendages absent. Segment IX dorsally narrow, broad medially and ventrally; anterior margin rounded; posterolateral margin membranous or very lightly sclerotized; sternum IX without modification. Tergum X incompletely fused to tergum IX with membrane or lightly sclerotized region ventrolaterally; dorsomesal margin with single, downturned, elongate process; dorsolateral margin with paired small, down-turned, finger-like process; ventrolateral margin with an outer pair of subquadrate setose processes directed posteriorly, and an inner pair of subtriangular processes directed posteroventrally. Inferior appendages present as single thumb-like setose process, broadest at base and fused to phallobase ventrobasally. Parameres present, paired, inserted in membranous lobe, arising laterally from endotheca, sclerotized and rod-like, extremely sinuous, corkscrew-shaped, apex pointed. Phallobase reduced, lightly sclerotized with phallic shield. Phallicata forming a rather short sclerotized dorsal sheath extending from phallobase, straight basally and medially, bent sharply upward, medially with pair of lightly slerotized lateral flanges with pointed apices projecting posteriorly. Endophallus membranous, apically with sclerotized bifid process.
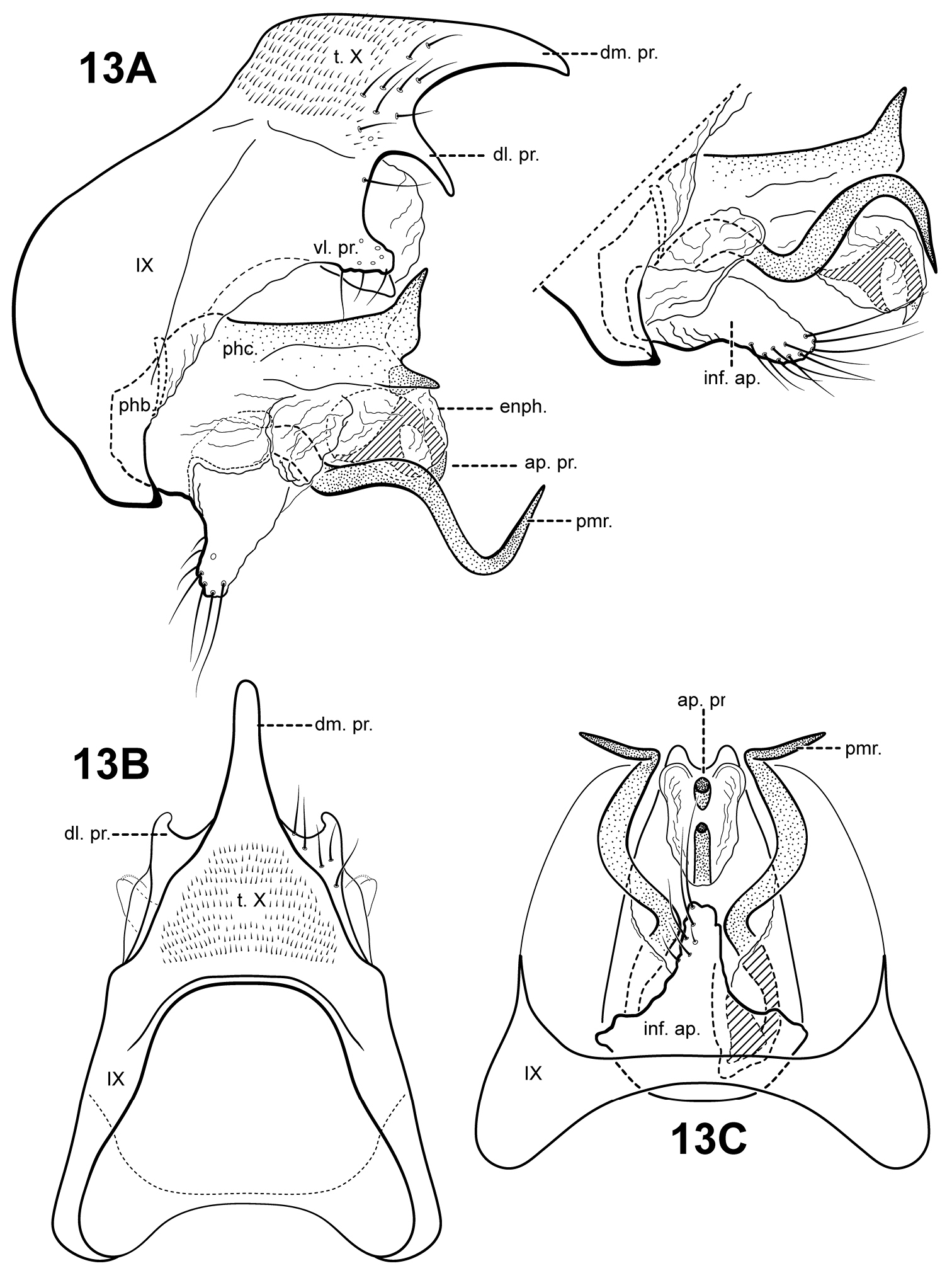
Itauara guyanensis sp. n. Male genitalia, everted (UMSP000210959) and non-everted (UMSP000210962). (A) lateral: left, everted; inset, non-everted (B) dorsal; (C) ventral. Abbreviations: ap. pr. = apical process; dl. pr. = dorsolateral process; dm. pr. = dorsomesal process; enph. = endophallus; inf. ap. = inferior appendage process; phb. = phallobase; phc. = phallicata; pmr . = paramere; t. X = tergum X; vl. pr. = ventrolateral process.
Holotype male: GUYANA: Dubulay Ranch: Warniabo Cr., 05°39'48"N, 057°53'24"W, 14–19.iv.1995 (O.S. Flint) (UMSP000210959) (NMNH).
Paratypes: GUYANA: Dubulay Ranch: Aramatani Cr., 05°39'24"N, 057°55'30"W, 15–18.iv.1995 (O.S. Flint) — 2 males, 2 females (NMNH).
This species is named for the country of Guyana, where the specimens were collected.
urn:lsid:zoobank.org:act:9EAE24A5-16C2-4B9F-8BDD-42976CE0E569
http://species-id.net/wiki/Itauara_jamesii
Fig. 14A–CItauara jamesii is known only from the male holotype, and its relationship to other species in not immediately evident. The parameres have a rather asymmetrical aspect, but it is possible that this particular specimen is distorted. The species has an inferior appendage process like several other species, but has a distinct subtriangular shape. The dorsomesal margin of tergum X is bifid, each half a small setose protuberance. Itauara peruensis sp. n., also has a bifid dorsomesal margin, but in that species, it appears as a prominent process with pointed apices.
Adult. Body, wings, and appendages pale or tawny brown, often intermingled with rufous or golden hairs, tibia and tarsi tawny brown. Wings with conspicuous white spot at the arculus and faint transverse line along anastomosis. Forewing slightly broader past anastomosis, but with margins nearly parallel, apex subacute. Forewing venation incomplete, with apical forks I, II, and III present; Sc and R1 distinct along their entire lengths; fork I sessile; fork II petiolate, stem shorter than fork; fork III petiolate, stem longer than fork; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present along Cu2; A3 absent; crossveins forming a relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing margins nearly parallel, tapering only slightly past anastomosis; apical forks II and V present; Sc and R1 fused basally; A2 absent. Tibial spurs 1, 4, 4, foretibial spur extremely reduced and hairlike. Sixth sternal process thumb-like, apex rounded, associated with weak oblique apodeme posteriorly.
Male genitalia. Preanal and inferior appendages absent. Segment IX dorsally and ventrally narrow, broad medially; anterior margin rounded; posterolateral margin membranous or very lightly sclerotized; sternum IX without modification. Tergum X incompletely fused to tergum IX with membrane or lightly sclerotized region ventrolaterally; dorsomesal margin bifid and slightly produced, each half small, setose, with a rounded apex; dorsolateral margin without processes; ventrolateral margin with paired, broad, semi-circular setose flange-like process. Inferior appendages present as single, broad, subtriangular setose process, fused to phallobase ventrobasally, bearing small digitate lobes ventrolaterally, each bearing a seta. Parameres present, paired, inserted in membranous lobe, arising laterally from endotheca, sclerotized and rod-like, extremely sinuous, seemingly asymmetrical, apex pointed. Phallobase reduced, lightly sclerotized with phallic shield. Phallicata forming a long sclerotized dorsal sheath extending from phallobase, broadest basally, bent upward at middle, with paired sclerotized concave discs arising basodorsally and forming a connection with posterior margin of segment IX. Endophallus membranous, enlarged and convoluted when invaginated.
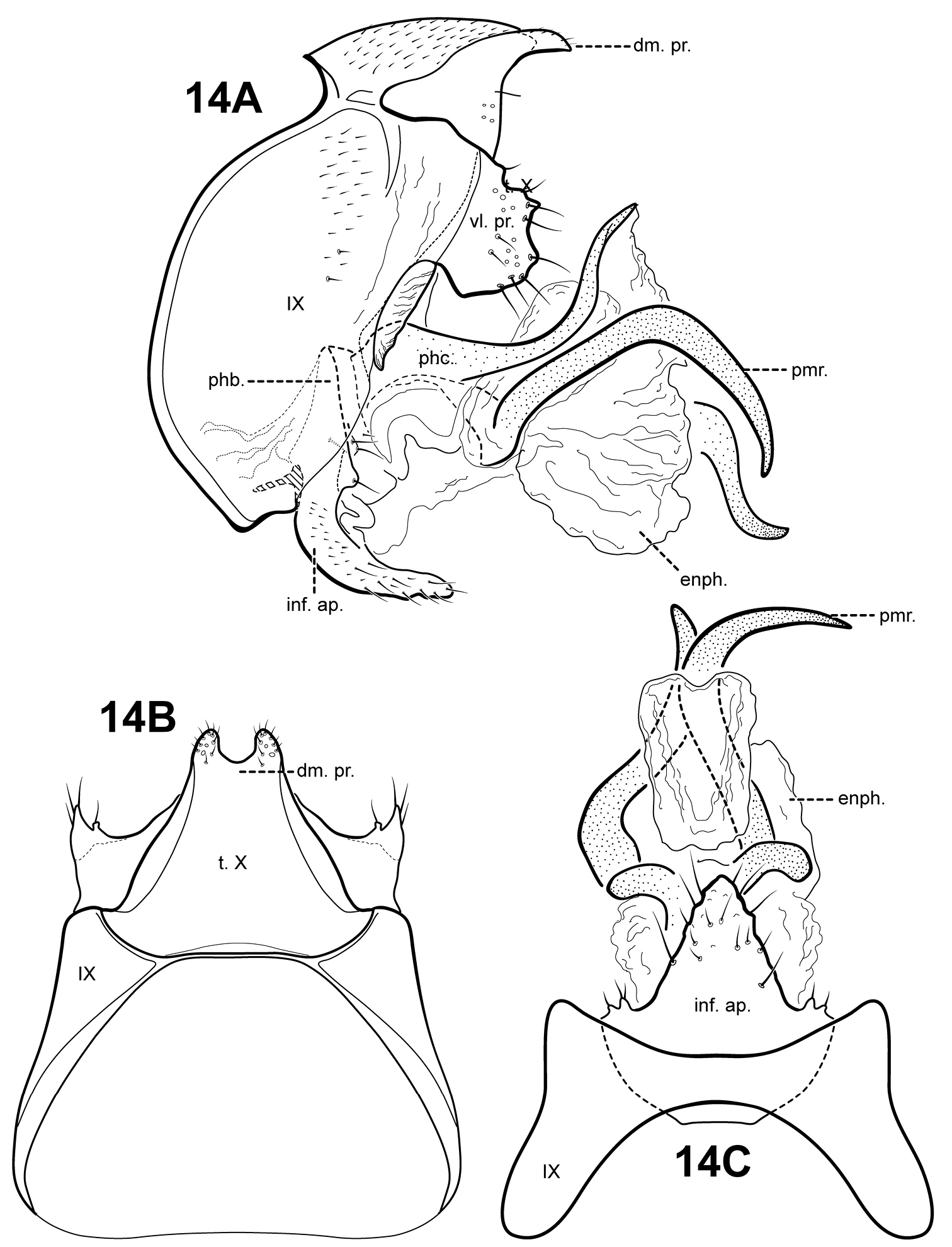
Itauara jamesii sp. n. (UMSP000087916). Male genitalia (A) lateral; (B) dorsal; (C) ventral. Abbreviations: dm. pr. = dorsomesal process; enph. = endophallus; enph. pr. = endophallic process; inf. ap. = inferior appendage process; phb. = phallobase; phc. = phallicata; pmr . = paramere; t. X = tergum X; vl. pr. = ventrolateral process.
Holotype male: BRAZIL: Minas Gerais: trib. to Rio do Salto, Ibitipoca, Fazenda Engenho, 21°44'06"S, 043°53'56"W, 875, 11–14.iii.2002 (Holzenthal, Blahnik, Paprocki, Prather) (UMSP000087916) (MZUSP).
We are delighted to name this species for the senior author’s father, James Gardner Robertson.
urn:lsid:zoobank.org:act:5DE975A9-37FA-4BB0-9A52-662621648658
http://species-id.net/wiki/Itauara_julia
Fig. 1, 3C, 15A–CThis species is distinct in having a curved, spatulate ventral process in the endophallus and having a phallicata that is not continuous with the phallobase. The parameres of Itaura julia are strongly bent dorsally at the base, and terminate in a sharp, downturned point. The parameres curve upward, are fused to and continuous with the phallobase, and arise ventrobasally. The parameres of Itaura blahniki sp. n., Itaura rodmani sp. n., and Itaura tusci sp. n., are similarly structured. However, in these species, the parameres are not as abruptly bent.
Adult. Body, wings, and appendages pale or tawny brown, often intermingled with rufous or golden hairs, tibia and tarsi tawny brown. Wings with conspicuous white spot at the arculus. Forewing slightly broader past anastomosis, but with margins nearly parallel, apex rounded. With apical forks I, II, and III present; Sc and R1 distinct along their entire lengths; fork I sessile; fork II sessile; fork III petiolate, stem shorter than fork; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present along Cu2; A3 absent; crossveins forming a relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing margins nearly parallel, tapering only slightly past anastomosis; apical forks II, III, and V present; Sc and R1 converging near wing margin; A2 absent. Tibial spurs 1, 4, 4, foretibial spur extremely reduced and hairlike. Sixth sternal process thumb-like, apex attenuate and pointed, associated with weak oblique apodeme posteriorly.
Male genitalia. Preanal and inferior appendages absent. Segment IX dorsally narrow, broad ventrally; anterior margin relatively straight from dorsum to medial area, ventral portion blunt; posterolateral margin membranous or very lightly sclerotized; sternum IX without modification. Tergum X incompletely fused to tergum IX with membrane or lightly sclerotized region ventrolaterally; dorsomesal margin straight, without processes; dorsolateral margin with several small irregular setose processes; ventrolateral margin with paired subquadrate setose process projecting ventrally. Parameres present, paired, arising ventrobasally and fused to phallobase, sclerotized and rod-like, slender and elongate, strongly bent upward basally, apex pointed and downturned, ventrobasally with a small patch of setae. Phallobase reduced, lightly sclerotized. Phallicata forming a long sclerotized dorsal sheath, mostly straight, broadest basally, apex with apicomesal point and 2 lateral downturned points. Endophallus membranous, enlarged and convoluted when invaginated, ventrally with a curved spatulate process.
Itauara julia sp. n. Male genitalia: Male genitalia (A) lateral; (B) dorsal; (C) ventral. Abbreviations: dl. pr. = dorsolateral process; enph. = endophallus; enph. pr. = endophallic process; phb. = phallobase; phc. = phallicata; pmr . = paramere; t. X = tergum X; vl. pr. = ventrolateral process.
Holotype male: BRAZIL: Rio de Janeiro: Parque Nacional do Itatiaia, Rio Campo Belo, trail to Veu da Noiva, 22°25'42"S, 044°37'10"W, 1310 m, 5.iii.2002 (Holzenthal, Blahnik, Paprocki, Prather) (UMSP000069560) (MZUSP).
Paratypes: BRAZIL: Rio de Janeiro: Parque Nacional do Itatiaia, same data as holotype — 9 males, 5 females (UMSP); same, 24.ix.2001 (Holzenthal, Blahnik, Neto, Paprocki) — 17 males, 8 females (UMSP); Rio Taquaral, 22°27'15"S, 044°36'34"W, 1300 m, 22–23.ix.2001 (Holzenthal & Blahnik) — 20 males; Rio Campo Belo, 22°27'02"S, 044°36'49"W, 1300 m, 23.ix.2001 (Holzenthal, Blahnik, Neto, Paprocki) — 28 males, 17 females (NMNH, UMSP); 7.iii.2002 (Holzenthal, Blahnik, Paprocki, Prather) —24 males, 46 females (MZUSP, UMSP).
We are delighted to name this species for Julie Martinez, who rendered the beautiful color plate of this species.
urn:lsid:zoobank.org:act:F8FA9F0B-D595-4FDC-92D8-7F64F183E91D
http://species-id.net/wiki/Itauara_lucinda
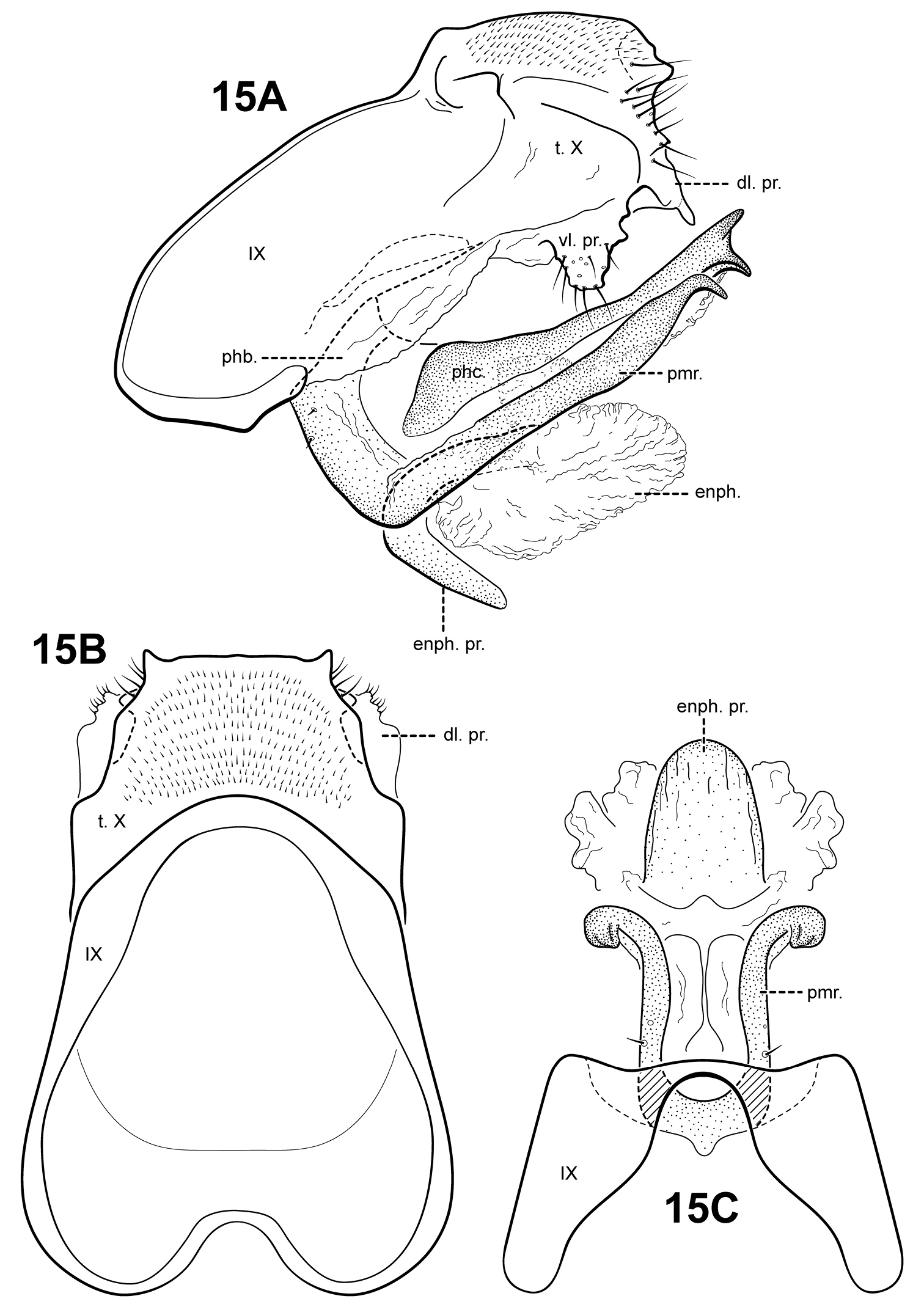
Fig. 16A–CThis species is similar to Itauara alexanderi sp. n., Itaura emilia sp. n., and Itaura stella sp. n., as discussed under each of those species. Each of these species possess an inferior appendage process, a dorsomesal process on tergum X, and rather sinuous parameres. Among these species, Itaura lucinda is distinct in having forked parameres and a non-bifid inferior appendage process.
Adult. Body, wings, and appendages fuscous, intermingled with rufous or golden hairs, tibia and tarsi tawny brown. Wings with white transverse line along anastomosis. Forewing slightly broader past anastomosis, but with margins nearly parallel, apex subacute. Forewing venation incomplete, with apical forks I, II, and III present; Sc and R1 distinct along their entire lengths; fork I sessile; fork II petiolate, stem about the same length as fork; fork III petiolate, stem longer than fork; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present along Cu2; A3 absent; discoidal cell longer than Rs vein. Hind wing margins nearly parallel, tapering only slightly past anastomosis; apical forks II and V present; Sc and R1 fused basally; A2 absent. Tibial spurs 1, 4, 4, foretibial spur extremely reduced and hairlike. Sixth sternal process thumb-like, apex attenuate and pointed, associated with oblique apodeme posteriorly.
Male genitalia. Preanal appendages absent. Segment IX dorsally narrow, broad medially and ventrally; anterior margin rounded; posterolateral margin membranous or very lightly sclerotized; sternum IX without modification. Tergum X incompletely fused to tergum IX with membrane or lightly sclerotized region ventrolaterally; dorsomesal margin with single, downturned, elongate process; dorsolateral margin without processes; ventrolateral margin with paired, broad flange-like setose process with small upper lobe and larger subquadrate lower lobe. Inferior appendages present as single, broad, subquadrate setose process, broadest basally, fused to phallobase ventrobasally, with 2 pairs of small digitate lobes ventrolaterally, each bearing a seta. Parameres present, paired, inserted in membranous lobe, arising laterally from endotheca, sclerotized and rod-like, bifid, with short lower process and longer, slightly medially bent upper process, directed posteriorly, apices pointed. Phallobase reduced, lightly sclerotized. Phallicata forming a long, straight sclerotized dorsal sheath extending from phallobase. Endophallus membranous, enlarged and convoluted when invaginated, with 1 upper and 1 lower lobe.
Itauara lucinda sp. n. (UMSP000052593). Male genitalia (A) lateral; (B) dorsal; (C) ventral. Abbreviations: dm. pr. = dorsomesal process; enph. = endophallus; inf. ap. = inferior appendage process; phb. = phallobase; phc. = phallicata; pmr . = paramere; t. X = tergum X; vl. pr. = ventrolateral process.
Holotype male: BRAZIL: Minas Gerais: Parque Nacional do Caparaó, small trib. to Rio Caparaó, Vale Verde, 20°25'02"S, 041°50'46"W, 1350, 12–14.iii.2002 (R.W. Holzenthal) (UMSP000052593) (MZUSP).
Paratype: BRAZIL: Minas Gerais: same data as holotype — 1 female (UMSP).
We are delighted to name this species for the senior author’s daughter, Lucinda Grace Thompson.
urn:lsid:zoobank.org:act:B534B28B-C523-4B04-9F33-FDE480C6D2BE
http://species-id.net/wiki/Itauara_ovis
Fig. 17A–CPerhaps the most notable feature of this species is the extremely curved, ram-like shaped parameres. Itauara spiralis sp. n., also has highly curved, spiral shaped parameres, but in Itaura spiralis, the paramere is curved along the entire length of the paramere, whereas in Itaura ovis, the paramere is curved basally, but straight distally. The 2 species differ in other respects, including the shape of the inferior appendage process. In Itaura ovis, the inferior appendage process is elongate, and rather inflated apically; in Itaura spiralis, the inferior appendage process is bifid. The 2 species also differ in the shape of tergum X. In Itaura ovis, tergum X is slightly notched apicomesally and has 2 pairs of rather small, subtriangular ventrolateral processes. In Itaura spiralis, tergum X is not notched, but has a pair of very long, finger-like dorsomesal processes and a pair of very broad, irregular ventrolateral processes. The phallicata of Itaura ovis is also quite distinct, being rather broad, and saddle-shaped, with a dorsobasal hump and upturned apex.
Adult. Body, wings, and appendages fuscous, intermingled with rufous or golden hairs, tibia and tarsi tawny brown. Wings often with a few pale cream-colored or white hairs at arculus. Forewing relatively narrow, with margins nearly parallel, apex subacute. Forewing venation incomplete, with apical forks I, II, and III present; fork I sessile; fork II petiolate, stem about the same length as fork; fork III petiolate, stem longer than fork; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present along Cu2; A3 absent; crossveins forming a relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing narrow and slightly scalloped past anastomosis; apical forks II and V present; Sc and R1 fused basally; A2 absent. Tibial spurs 1, 4, 4, foretibial spur extremely reduced and hairlike. Sixth sternal process short and digitate, apex attenuate and pointed, associated with strong oblique apodeme posteriorly.
Male genitalia. Preanal appendages absent. Segment IX ventrally narrow, broad medially; anterior margin rounded; posterolateral margin lightly sclerotized; sternum IX without modification. Tergum X incompletely fused to tergum IX with membrane or lightly sclerotized region ventrolaterally; dorsomesal margin subtriangular, slightly produced with small cleft; dorsolateral margin without processes; ventrolateral margin with an outer pair of subtriangular setose processes directed ventrally, and an inner pair of subtriangular processes directed posteroventrally. Inferior appendages present as single, rather elongate setose process produced mesally, apex broad and slightly irregular, fused to phallobase ventrobasally. Parameres present, paired, inserted in membranous lobe, arising laterally from endotheca, sclerotized and rod-like, ram-like, curving 360 degrees at base, distal portion straight, directed posteriorly, apex pointed. Phallobase extremely reduced and difficult to discern. Phallicata forming a saddle-shaped sclerotized dorsal sheath, with dorsal hump basally, distal portion curving upward. Endophallus membranous, enlarged and convoluted when evaginated, with pair of elongate lateral sclerites ventrally.

Itauara ovis sp. n. (UMSP000118534) Male genitalia. (A) lateral; (B) dorsal; (C) ventral. Abbreviations: enph. = endophallus; enph. scl. = endophallic sclerite; inf. ap. = inferior appendage process; phc. = phallicata; pmr . = paramere; t. X = tergum X; vl. pr. = ventrolateral process.
Holotype male: GUYANA: Kanuku Mountains: Kumu River & Falls, 03°15'54"N, 059°43'30"W, 28–30.iv.1995 (O.S. Flint) (UMSP000118534) (NMNH)
Paratypes: VENEZUELA: Bolivar: La Escalera, 108 km. S Rio Cuyuni, 11–12.ii.1976 (C. & O. Flint) — 5 males (NMNH).
The name ovis, comes from the Latin for sheep, and is suggested by the shape of the parameres, which are reminiscent of a ram’s horn.
urn:lsid:zoobank.org:act:0F3E4131-B78D-470A-9DA5-83228D16BEC6
http://species-id.net/wiki/Itauara_peruensis
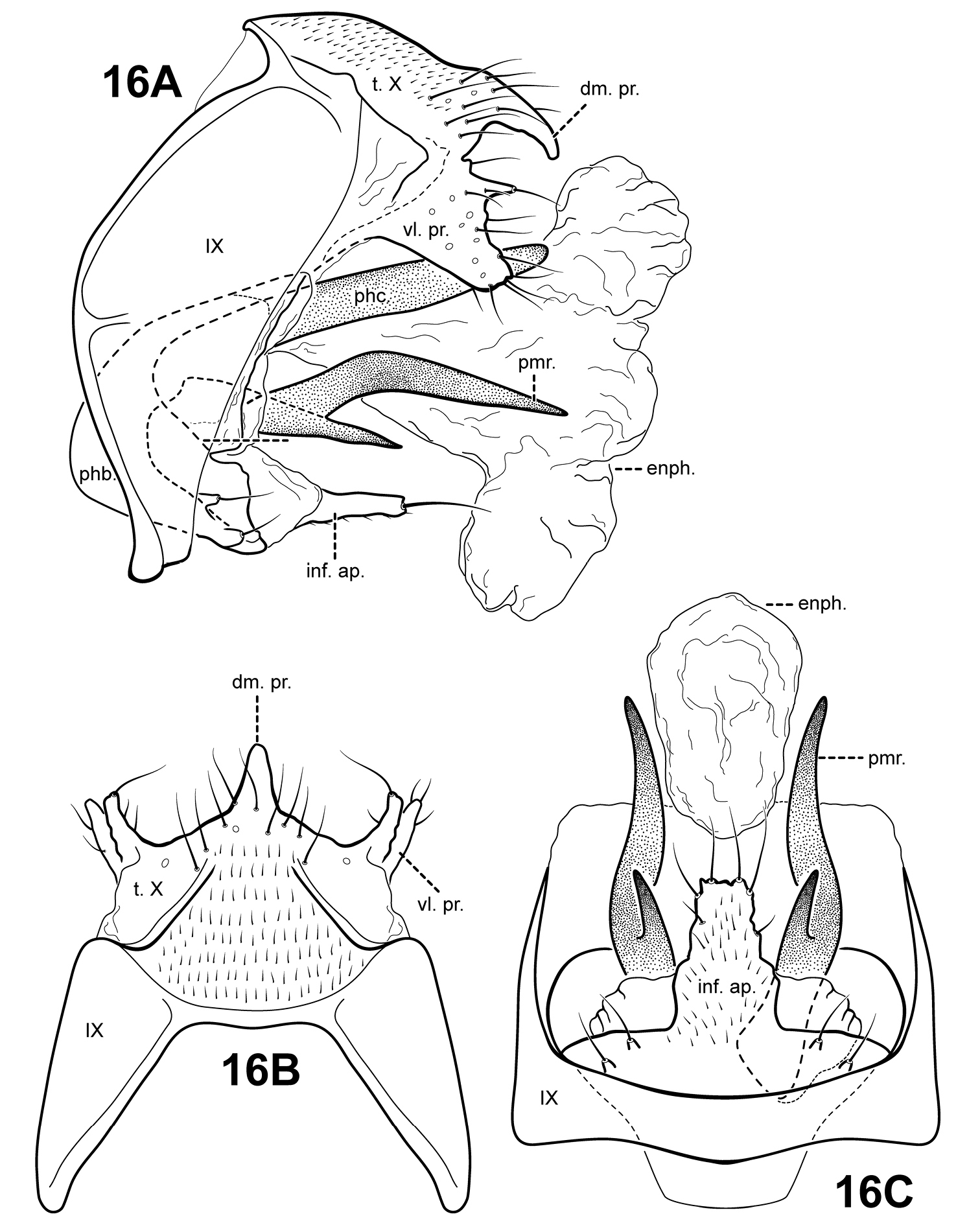
Fig. 18A–CThis species is distinct in having a lightly sclerotized endophallus. Ventrally, the endophallus is membranous, but in lateral view, it has the appearance of being entirely sclerotized. Another unique feature is the prominent bifid dorsomesal process of tergum X. Itauara peruensis has a rather elongate inferior appendage process like Itaura ovis, but it is not inflated apically like that species. The species also differ in the shape of the parameres; those of Itaura peruensis are nearly straight, while those of Itaura ovis are spiral-shaped.
Adult. Body, wings, and appendages pale or tawny brown in alcohol. Forewing narrow past anastomosis, apex acute. Forewing venation incomplete, with apical forks I, II, and III present; Sc and R1 distinct along their entire lengths; fork I sessile; fork II petiolate, stem about the same length as fork; fork III petiolate, stem longer than fork; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present along Cu2; A3 absent; crossveins forming a relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing narrow and slightly scalloped past anastomosis; apical fork II present; Sc and R1 fused basally; A2 absent. Tibial spurs 1, 4, 4, foretibial spur extremely reduced and hairlike. Sixth sternal process short and digitate, apex attenuate and pointed, associated with strong oblique apodeme posteriorly.
Male genitalia. Preanal appendages absent. Segment IX ventrally narrow, broad medially; anterior margin rounded; posterolateral margin membranous or very lightly sclerotized; sternum IX without modification. Tergum X incompletely fused to tergum IX with membrane or lightly sclerotized region ventrolaterally; dorsomesal margin with bifid process, each half with a pointed apex; dorsolateral margin slightly irregular, without processes; ventrolateral margin with paired subtriangular setose process directed ventrally. Inferior appendages present as single, narrow, rather short setose process produced mesally, fused to phallobase ventrobasally. Parameres present, paired, arising laterally from anterior portion of phallobase, sclerotized and rod-like, slender and elongate, straight, very slightly downturned, apex pointed. Phallobase reduced, lightly sclerotized. Phallicata forming a long sclerotized dorsal sheath, mostly straight, broadest basally, distal portion very slightly upturned. Endophallus lightly sclerotized tubular structure, ventrally with membranous folds, apically with small phallotremal sclerite.
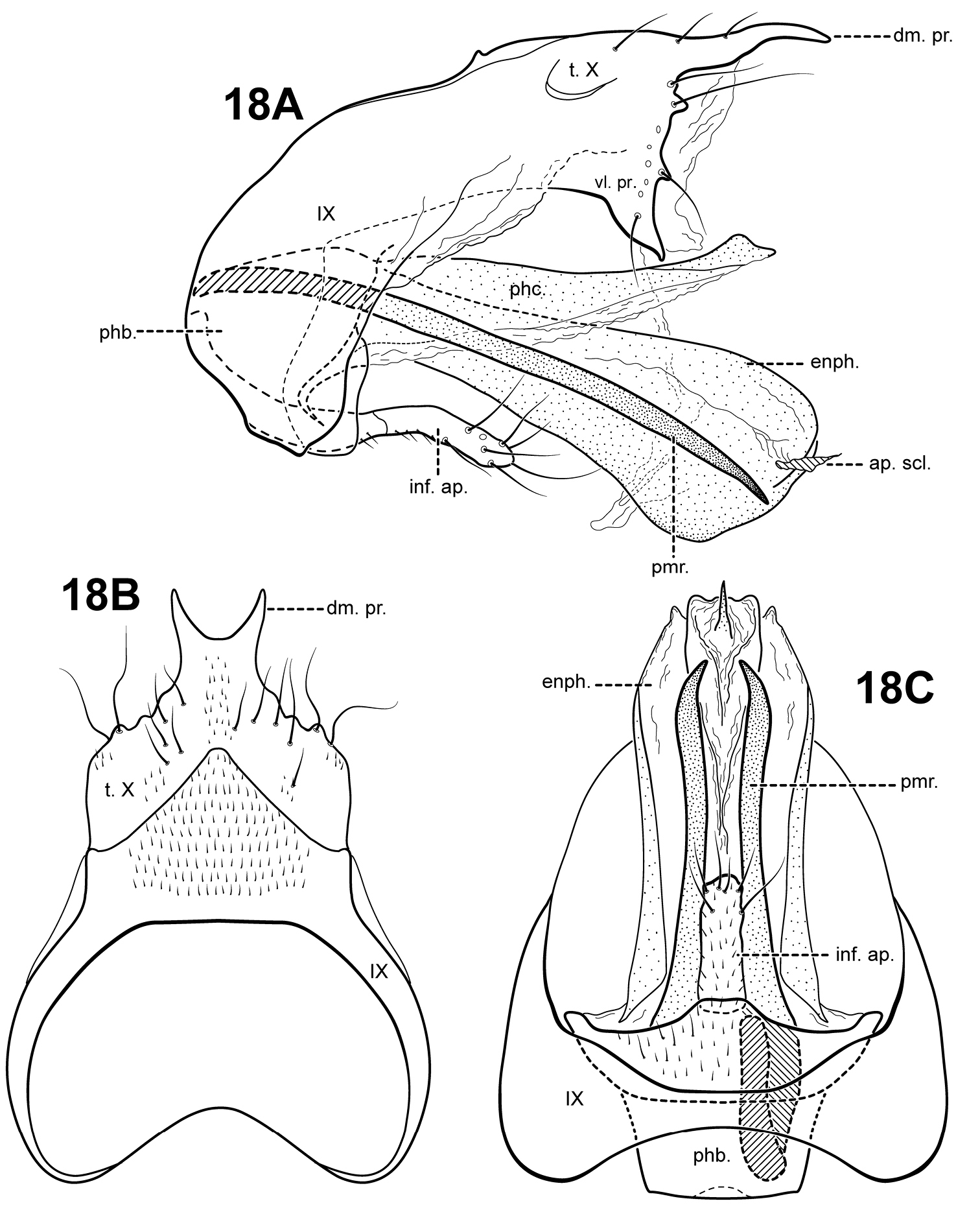
Itauara peruensis sp. n. (UMSP000210957). Male genitalia (A) lateral; (B) dorsal; (C) ventral. Abbreviations: ap. scl. = apical sclerite; dm. pr. = dorsomesal process; enph. = endophallus; inf. ap. = inferior appendage process; phb. = phallobase; phc. = phallicata; pmr . = paramere; t. X = tergum X; vl. pr. = ventrolateral process.
Holotype male: PERU: Madre de Dios: Manu Biosphere Reserve, Pakitza Biological Station, Trail 2, 1st stream, 12°07'00"S, 070°58'00"W, 250 m, 14–23.ix.1988 (Flint & Adams) (UMSP000210957) (NMNH)
Paratypes: PERU: Madre de Dios: same data as holotype — 7 males, 21 females; same, 17–20.ix.1988 (Flint & Adams) — 1 female (NMNH).
This species is named for the country of Peru, where the specimens were collected.
http://species-id.net/wiki/Itauara_plaumanni
Fig. 19A–DItauara plaumanni can be recognized by the irregular lobe-like shape of the lateral flanges on the phallicata. Additionally, this species has a rather elongate segment IX and tergum X. The profile of tergum X resembles that of Itaura brasiliana, but Itaura plaumanni has fewer lateral processes than that species. Itauara plaumanni has parameres similar in shape to those of Itaura guarani, yet these species are easily separated by differences in the shape of the lateral flanges of the phallicata, and tergum X.
Adult. Body, wings, and appendages pale or tawny brown in alcohol. Forewing slightly broader past anastomosis, but with margins nearly parallel, apex rounded. Forewing venation incomplete, with apical forks I, II, and III present; Sc and R1 distinct along their entire lengths; fork I sessile; fork II petiolate, stem shorter than fork; fork III petiolate, stem about the same length as fork; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present along Cu2; A3 absent; crossveins forming a relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing margins nearly parallel, tapering only slightly past anastomosis; apical forks II, III, and V present; Sc and R1 fused basally; A2 absent. Tibial spurs 1, 4, 4, foretibial spur extremely reduced and hairlike. Sixth sternal process short and digitate, apex attenuate and pointed.
Male genitalia. Preanal and inferior appendages absent. Segment IX relatively broad; anterior margin rounded; posterolateral margin membranous or very lightly sclerotized; sternum IX without modification. Tergum X incompletely fused to tergum IX with membrane or lightly sclerotized region ventrolaterally; dorsomesal margin slightly produced as small irregular point; dorsolateral margin with 2 pairs of processes, the upper an elongate subtriangular process slightly downturned, the lower a small lobe-like setose process; ventrolateral margin without processes. Parameres present, paired, arising ventrobasally and fused to phallobase, sclerotized and rod-like, slender and elongate, sinuous, downturned basally, distal portion slightly upturned, directed posteriorly and inward, apex pointed, ventrobasally with small patch of setae. Phallobase reduced, lightly sclerotized. Phallicata forming a long sclerotized dorsal sheath, slightly bent upward medially, with pair of irregular lobe-like lateral flanges projecting posteroventrally. Endophallus membranous, enlarged and convoluted when invaginated, with lightly sclerotized lobe ventrally and laterally, containing 2 small sclerites.
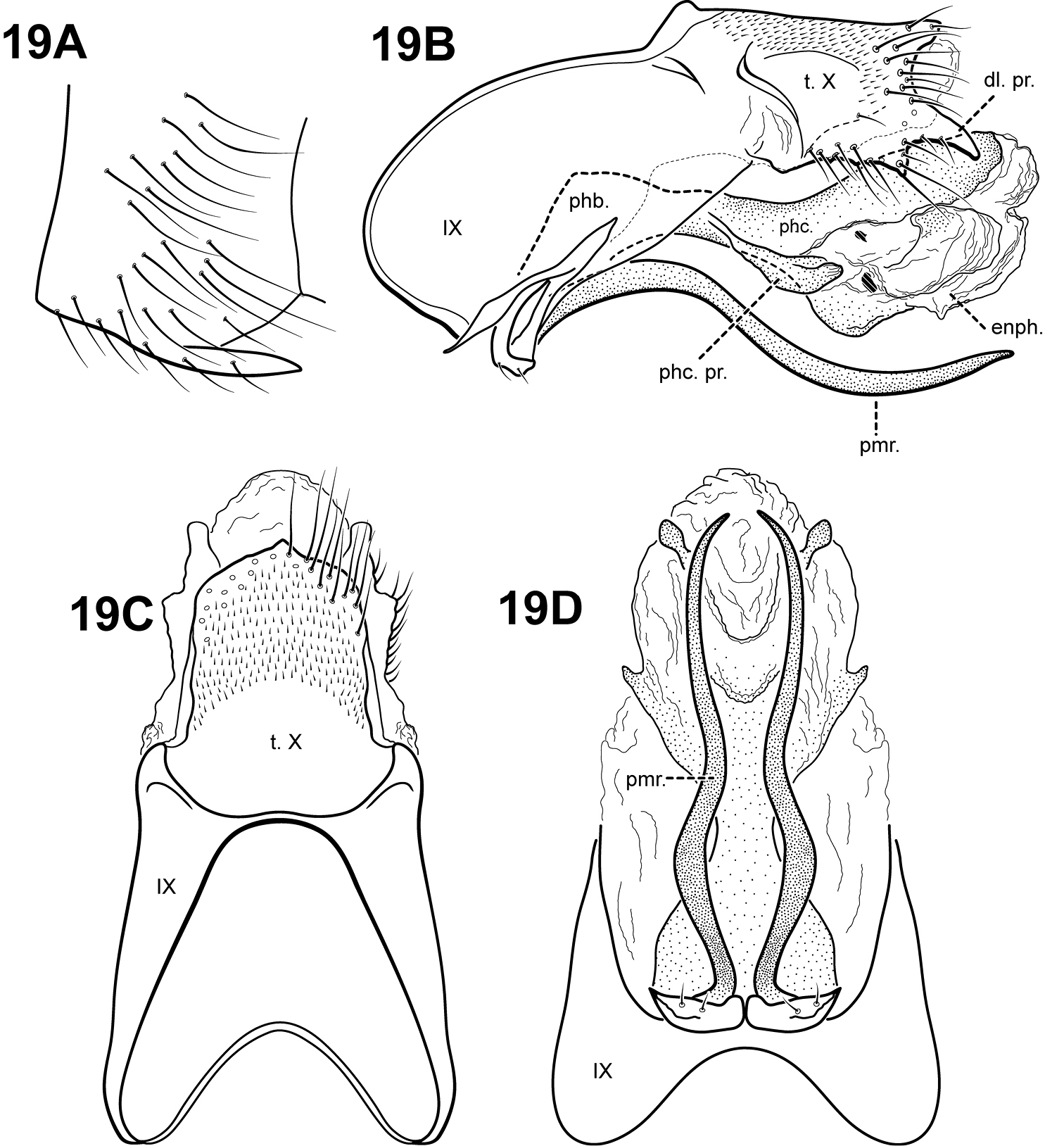
Itauara plaumanni (Flint, 1974) (UMSP000086359). Male genitalia: (A) Sternum VI process; ttt (B) lateral; (C) dorsal; (D) ventral. Abbreviations: dl. pr. = dorsolateral process; enph. = endophallus; phb = phallobase; phc. = phallicata; phc. pr. = phallicata process; pmr . = paramere; t. X = tergum X.
Holotype male: BRAZIL: Santa Catarina: Nova Teutonia, 27°03'00"S, 052°24'00"W, 1.ii.1964 (F. Plaumann) (UMSP000027160) (NMNH).
Paratypes: BRAZIL: Santa Catarina: same data as holotype, 1.ix.1963 (F. Plaumann) — 1 male (NMNH), same, 1.xi.1963 (F. Plaumann) — 7 males (NMNH); same, Nova Teutonia, 27°11'00"S, 052°23'00"W, 300–500 m, 1.i.1963 (F. Plaumann) — 1 male, (NMNH), same, 1.i.1964 (F. Plaumann) — 7 males (NMNH).
Additional material examined: ARGENTINA: Misiones: Cataratas del Iguazú, 14.x.1985 — 2 males (MACN); Salto: Salto Grande, cascada, 10.xi.1955 (C.S. Carbonell) — 46 males (MACN); URUGUAY: Artigas: San Gregorio, 30°33'00"S, 057°52'00"W (Carbonell, Mesa, & San Martin) — 1 male (MACN); Orillas Rio Uruguay (Carbonell, AM, PSM) — 1 male, 3 females (MACN); Paysandu: Sta. Rita, Orilla Rio Uruguay, 32°07'00"S, 058°09'00"W, 8.xii.1955 (C.S. Carbonell) — 10 males (MACN); 1.xii.1959 (C.S. Carbonell) — 1 male (MACN).
urn:lsid:zoobank.org:act:B5D07A37-7980-4300-B3E1-C856A34EA331
http://species-id.net/wiki/Itauara_rodmani
Fig. 20A–DThis species is very similar to Itaura tusci sp. n., which also has very long, upturned, tusk-like parameres and a strongly upturned phallicata. The 2 species can be separated based on the shape of the phallicata process, which is pointed and blade-like in Itaura tusci and rounded or blunt in Itaura rodmani. The phallicata is also more sclerotized in Itaura rodmani. Additionally, the dorsomesal margin of tergum X is irregular, with several small setose processes in Itaura tusci, whereas in Itaura rodmani, the dorsomesal margin is rather smooth and triangular. The ventrolateral processes of the 2 species also differ: Itaura tusci has a small upper and more elongate lower process; Itaura rodmani has a single, short, digitate process. Itaura blahniki also has upturned, tusk-like parameres, but is easily distinguished from Itaura rodmani based on differences in tergum X and the phallicata. Itauara blahniki and Itaura blahniki sp. n., also has tusk-like parameres, but they are not as long and curved as Itaura rodmani.
Adult. Body, wings, and appendages pale or tawny brown in alcohol. Wings with conspicuous white spot at the arculus. Forewing slightly broader past anastomosis, but with margins nearly parallel, apex subacute. Forewing venation incomplete, with apical forks I, II, and III present; Sc and R1 distinct along their entire lengths; fork I sessile; fork II petiolate, stem shorter than fork; fork III petiolate, stem about the same length as fork; Cu1 incomplete, not reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present along Cu2; A3 absent; crossveins forming a relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing margins nearly parallel, tapering only slightly past anastomosis; apical forks II, III, and V present; Sc and R1 fused basally; A2 absent. Tibial spurs 1, 4, 4, foretibial spur extremely reduced and hairlike. Sixth sternal process thumb-like, apex rounded, associated with weak oblique apodeme posteriorly.
Male genitalia. Preanal and inferior appendages absent. Segment IX relatively broad; anterior margin rounded; posterolateral margin membranous or very lightly sclerotized; sternum IX without modification. Tergum X incompletely fused to tergum IX with membrane or lightly sclerotized region ventrolaterally; dorsomesal margin subtriangular, very slightly downturned; dorsolateral margin with paired small, slightly down-turned, setose process; ventrolateral margin with an outer and inner pair of small setose processes directed posteriorly. Parameres present, paired, arising ventrobasally from fused endotheca and phallobase, sclerotized and rod-like, tusk-like, strongly curving upward, apex pointed. Phallobase reduced, lightly sclerotized with phallic shield. Phallicata forming a long sclerotized dorsal sheath extending from phallobase, strongly curving upward with apex directed dorsally, with pair of broad, sclerotized wing-like lateral flanges with rounded or subquadrate ventral margins. Endophallus membranous, enlarged and convoluted when invaginated, with 1 tubular upper lobe and 3 smaller lower lobes.
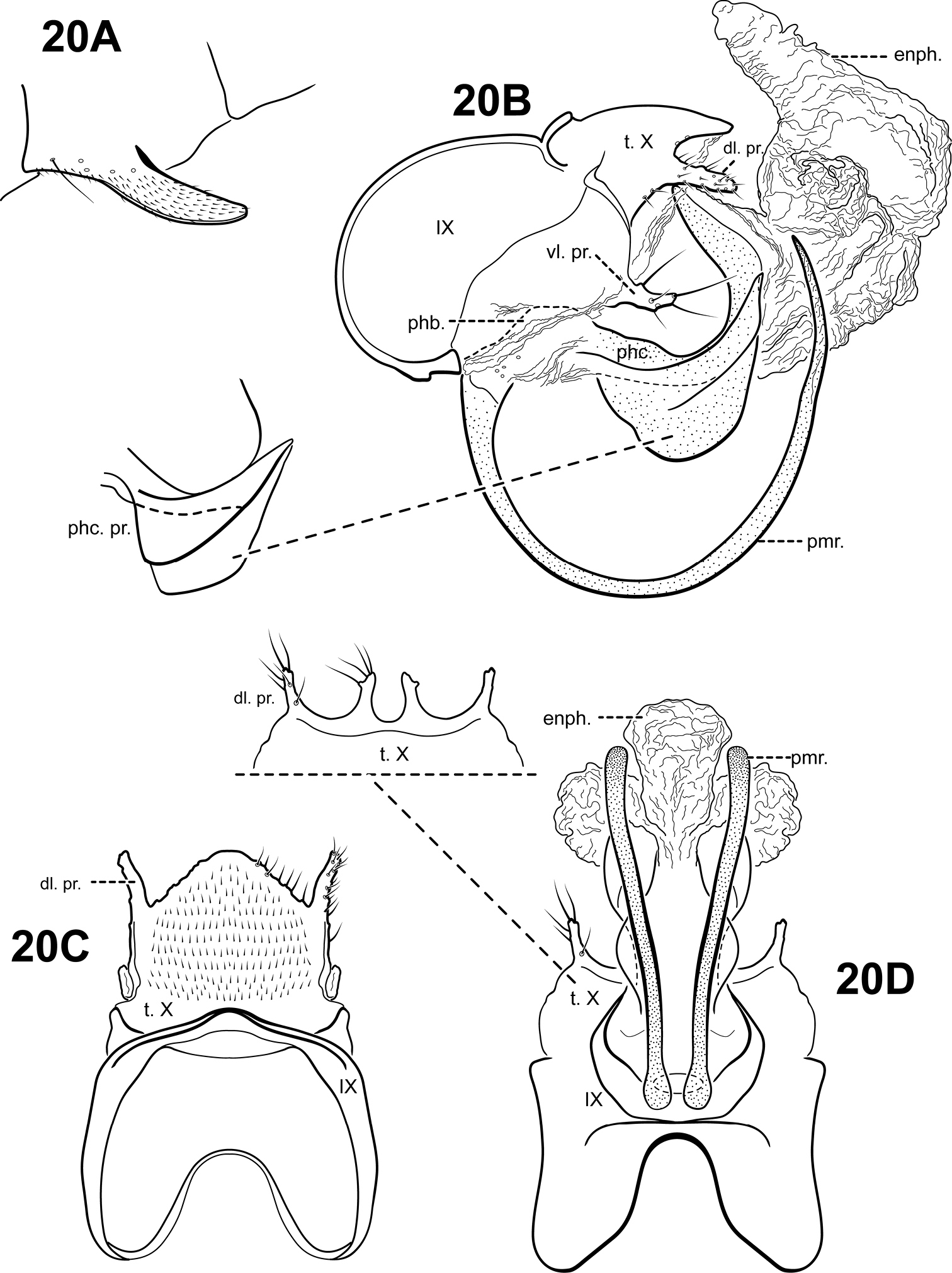
Itauara rodmani sp. n. (UMSP000081856). Male genitalia. (A) sternum VI process; (B) lateral; inset, variant (C) dorsal; (D) ventral; inset, tergum X. Abbreviations: dl. pr. = dorsolateral process; enph. = endophallus; phb = phallobase; phc. = phallicata; phc. pr. = phallicata process; pmr . = paramere; t. X = tergum X; vl. pr. = ventrolateral process.
Holotype male: BRAZIL: Minas Gerais: Corrego das Aguas Pretas & tribs., ca. 15 km S Aiuruoca, 22°03'42"S, 044°38'14"W, 1386 m, 21.xi.2001 (Holzenthal, Blahnik, Neto, Paprocki) (UMSP000081857) (MZUSP).
Paratypes: BRAZIL: Minas Gerais: same data as holotype — 6 females, 3 males (UMSP).
We are delighted to name this species for Dr. James Rodman, the NSF program director who initiated the Partnership for Enhancing Expertise in Taxonomy (PEET) program. The PEET program provides funding for the training of taxonomists of little known organisms. The senior author is grateful for the wonderful experience she had while participating in the PEET program as a doctoral student and the opportunity to study Trichoptera taxonomy.
urn:lsid:zoobank.org:act:E7688425-1DE8-4A29-8902-DCDCFC75B036
http://species-id.net/wiki/Itauara_simplex
Fig. 21A–CThis species can be recognized by its rather simple genitalic capsule. Tergum X is produced dorsomesally into a broad, elongate plate and has just one small ventrolateral process. The parameres are relatively short, straight basally, but slightly bent downward distally. The phallicata is short and very lightly sclerotized, and the endophallus is large and membranous, with 2 lateral patches or elongate setae apically. Itauara guarani also has a very lightly sclerotized phallicata, but the 2 species differ in the shape of the parameres and Itaura guarani has lateral flange-like processes on the phallicata.
Adult. Body, wings, and appendages fuscous, intermingled with rufous or golden hairs. Wings with conspicuous white spot at the arculus. Forewing slightly broader past anastomosis, but with margins nearly parallel, apex subacute. Forewing venation incomplete, with apical forks I, II, and III present; Sc and R1 distinct along their entire lengths; fork I petiolate, but with extremely short stem; fork II petiolate, stem about the same length as fork; fork III petiolate; stem longer than fork; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present along Cu2; A3 absent; crossveins forming a relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing margins nearly parallel, tapering only slightly past anastomosis; apical fork II present; Sc and R1 fused basally; A2 absent. Tibial spurs 1, 4, 4, foretibial spur extremely reduced and hairlike. Sixth sternal process thumb-like, apex rounded, associated with weak oblique apodeme posteriorly.
Male genitalia. Preanal and inferior appendages absent. Segment IX dorsally narrow, broad medially and ventrally; anterior margin rounded; posterolateral margin lightly sclerotized; sternum IX without modification. Tergum X incompletely fused to tergum IX with membrane or lightly sclerotized region ventrolaterally; dorsomesal margin produced into a single broad, plate-like process; dorsolateral margin slightly irregular, without processes; ventrolateral margin with small, irregular, paired setose process. Parameres present, paired, arising ventrobasally from fused endotheca and phallobase, sclerotized and rod-like, slender and elongate, straight medially and basally, curving downward distally, directed posteroventrally, apex pointed. Phallobase reduced, lightly sclerotized with phallic shield. Phallicata short, with lightly sclerotized base, rugous medially, becoming membranous distally. Endophallus membranous, enlarged and convoluted when invaginated, with1 upper lobe and 2 lower lobes, with paired patch of elongate setae laterally.
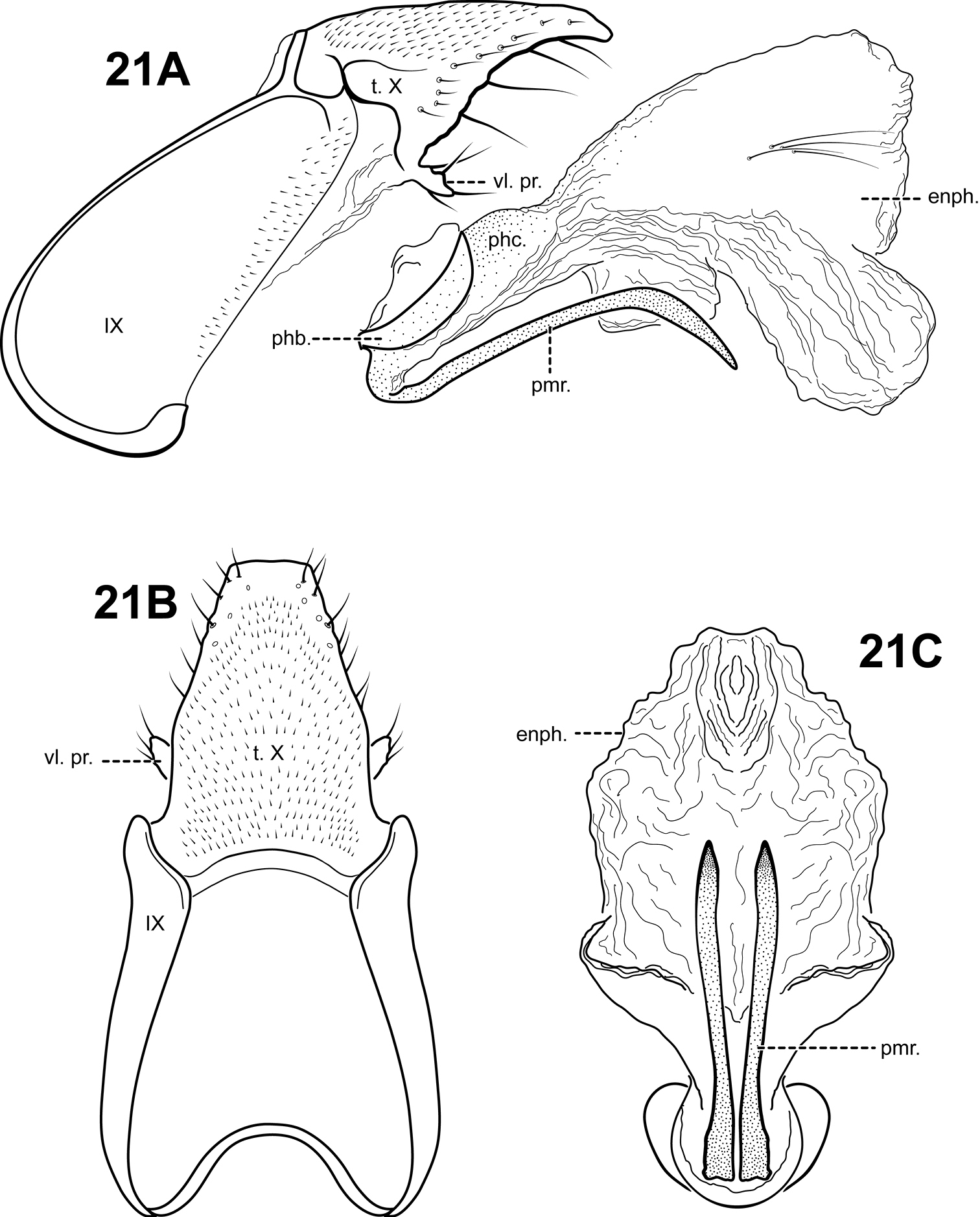
Itauara simplex sp. n. (UMSP000069700). Male genitalia: (A) lateral; (B) dorsal; (C) ventral. Abbreviations: enph. = endophallus; phb = phallobase; phc. = phallicata; pmr . = paramere; t. X = tergum X; vl. pr. = ventrolateral process.
Holotype male: BRAZIL: São Paulo: Parque Nacional da Serra da Bocaina, Cachoeira dos Posses, 22°46'26"S, 044°36'15"W, 1250, 3.iii.2002 (Holzenthal, Blahnik, Paprocki, Prather) (UMSP000069700) (MZUSP).
Paratype: BRAZIL: São Paulo: same data as holotype — 1 female (UMSP).
This species is so named for the rather simple structure of the phallic apparatus and genital capsule.
urn:lsid:zoobank.org:act:0568AD89-DD46-4FCB-87F4-0F9D884AEC04
http://species-id.net/wiki/Itauara_spiralis
Fig. 22A–CThis species is distinct in having a sclerotized, tubular phallicata, and an elongate, laterally compressed, dorsomesal spine. The phallicata in other species are less tubular, appearing as a dorsal sheath. This dorsal sheath was identified as a synapomorphy for the genus in a previous phylogenetic study of Protoptilinae (see Chapter 1, this work). Itauara spiralis was not included in that study, however, Itaura spiralis is placed in Itauara since it shares many other characteristics common to the genus such as an inferior appendage process, and a tergum X that is nearly identical to Itaura bidentata sp. n. and Itaura unidentata sp. n.
Itauara spiralis can be recognized by the extremely curved, spiral-shaped parameres. Itaura ovis sp. n., also has highly curved, spiral shaped parameres, but in Itaura spiralis, the paramere is curved along the entire length of the paramere, whereas in Itaura ovis, the paramere is curved basally, but straight distally. The 2 species differ in other respects, including the shape of the inferior appendage process, which is bifid in Itaura spiralis and inflated apically in Itaura ovis. The 2 species also differ in the shape of tergum X and the phallicata. Tergum X is very similar to those of Itaura bidentata and Itaura unidentata; all have elongate, finger-like dorsolateral processes and broad, irregular, setose ventrolateral processes. Itauara spiralis is distinguished from these other 2 species by having a bifid inferior appendage process, spiral-shaped parameres, and laterally compressed dorsomesal spine.
Adult. The only specimen of this species is in very poor condition. Therefore, head, thoracic, and wing characters could not be observed. However, the genitalia are intact.
Male genitalia. Preanal appendages absent. Segment IX ventrally narrow, broad medially; anterior margin rounded; posterolateral margin membranous or very lightly sclerotized; sternum IX without modification. Tergum X incompletely fused to tergum IX with membrane or lightly sclerotized region ventrolaterally; dorsomesal margin straight, shallowly excavate; dorsolateral margin with paired elongate, down-turned, finger-like process; ventrolateral margin with paired, very broad flange-like setose process consisting of several small irregular lobes. Inferior appendages present as apically bifid, setose process produced mesally, broadest at base and fused to phallobase ventrobasally. Parameres present, paired, arising laterally from endotheca, spiral-shaped, curving 360 degrees at base with curve continuing to apex, directed posteroventrally, apex pointed. Phallobase reduced, lightly sclerotized. Phallicata forming a short slerotized dorsal tube extending from phallobase, with a long, broad dorsomesal spine arising posteriorly to phallobase. Endophallus membranous, rather small, apically sharply bent downward, pointing anteroventrally.

Itauara spiralis sp. n. (UMSP000210960). Male genitalia (A) lateral; (B) dorsal; (C) ventral. Abbreviations: dl. pr. = dorsolateral process; dm. sp. = dorsomesal spine; enph. = endophallus; inf. ap. = inferior appendage process; phb. = phallobase; phc. = phallicata; pmr . = paramere; t. X = tergum X; vl. pr. = ventrolateral process.
Holotype male: GUYANA: Paramakatoi: 04°42'00"N, 059°42'48"W, 24–25.viii.1997 (W.N. Mathis) (UMSP0000210960) (NMNH).
The name spiralis refers to the spiral form of the parameres.
urn:lsid:zoobank.org:act:D8B2481D-60A2-42D3-A6A7-3D904B397203
http://species-id.net/wiki/Itauara_stella
Fig. 23A–CThis species is associated with Itauara alexanderi sp. n., Itaura emilia sp. n., and Itaura lucinda sp. n., as discussed under each of those species. Each of these species possess an inferior appendage process, a dorsomesal process on tergum X, and rather sinuous parameres. Of these species, Itaura stella is most similar to Itaura alexanderi. Both of these species have similarly shaped elongate dorsomesal processes and broad ventrolateral processes of tergum X. Both also have apically bifid inferior appendage processes. Itauara stella can be distinguished from by the length of the parameres; those of Itaura stella are longer and more strongly directed laterally than those of Itaura alexanderi. Additionally, the inferior appendage process of Itaura alexanderi is broader than that of Itaura stella. Itauara stella can be differentiated from Itaura emilia by the shape of the dorsomesal process and from Itaura lucinda by the shape of the parameres and inferior appendage process.
Adult. Body, wings, and appendages pale or tawny brown in alcohol. Wings with white transverse line along anastomosis. Forewing slightly broader past anastomosis, but with margins nearly parallel, apex rounded. Forewing venation incomplete, with apical forks I, II, and III present; Sc and R1 distinct along their entire lengths; fork I sessile; stem about the same length as fork; fork III petiolate, stem about the same length as fork; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present along Cu2; A3 absent; crossveins forming a relatively linear transverse cord. Hind wing margins nearly parallel, tapering only slightly past anastomosis; apical forks II and V present; Sc and R1 fused basally; A2 absent. Tibial spurs 1, 4, 4, foretibial spur extremely reduced and hairlike. Sixth sternal process thumb-like, apex attenuate and pointed, associated with weak oblique apodeme posteriorly.
Male genitalia. Preanal appendages absent. Segment IX ventrally narrow, broad medially; anterior margin rounded; posterolateral margin lightly sclerotized; sternum IX without modification. Tergum X incompletely fused to tergum IX with membrane or lightly sclerotized region ventrolaterally; dorsomesal margin with single, downturned, elongate process; dorsolateral margin irregular and setose; ventrolateral margin with paired, broad flange-like setose process consisting of small upper lobe and larger subtriangular lower lobe. Inferior appendages present as apically bifid, setose process produced mesally, broadest at base and fused to phallobase ventrobasally, with 2 pairs of small digitate lobes ventrolaterally, each bearing a seta. Parameres present, paired, inserted in membranous lobe, arising laterally from endotheca, sclerotized and rod-like, long, sinuous, directed outward and posteriorly, apex pointed. Phallobase reduced, lightly sclerotized. Phallicata forming a long sclerotized dorsal sheath extending from phallobase, broadest basally, narrowed slightly, distal portion curving dorsally. Endophallus membranous, enlarged and convoluted when invaginated, with 1 tubular upper lobe and 1 smaller lower lobe.
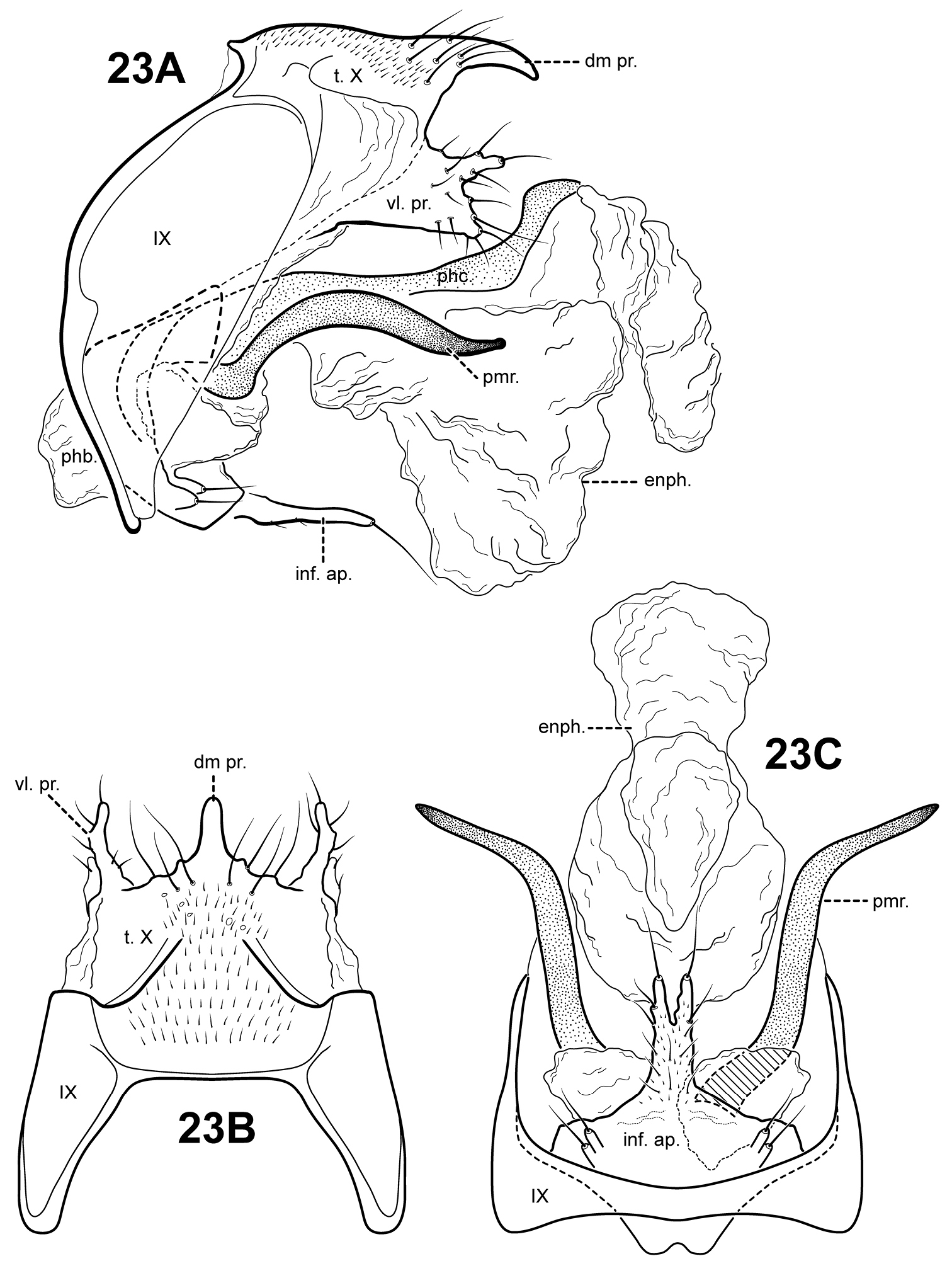
Itauara stella sp. n. (UMSP000052589). Male genitalia (A) lateral; (B) dorsal; (C) ventral. Abbreviations: dm. pr. = dorsomesal process; enph. = endophallus; inf. ap. = inferior appendage process; phb. = phallobase; phc. = phallicata; pmr . = paramere; t. X = tergum X; vl. pr. = ventrolateral process.
Holotype male: BRAZIL: Sao Paulo: Estação Biológica Boraceia: Rio Venerando & tribs, 23°39'11"S, 045°53'25"W, 850 m, 18–21.ix.2002 (Blahnik, Prather, Melo, Froehlich, Silva) (UMSP000052589) (MZUSP)
Paratypes: BRAZIL: Sao Paulo: same data as holotype — 9 males, 9 females (UMSP).
We are delighted to name this species for the senior author’s daughter, Stella Claire Thompson.
urn:lsid:zoobank.org:act:8080F8F1-4471-4A27-A919-65C247501BDD
http://species-id.net/wiki/Itauara_tusci
Fig. 24A–CThis species is very similar to Itaura rodmani sp. n., which also has very long, upturned, tusk-like parameres and a strongly upturned phallicata. The 2 species are separated based on the shape of the phallicata process, which is pointed and blade-like in Itaura tusci and rounded or blunt in Itaura rodmani. The phallicata of Itaura tusci is more lightly sclerotized than Itaura rodmani, especially at the distal portion. Additionally, in Itaura tusci, the dorsomesal margin of tergum X is irregular, with several small setose processes, whereas in Itaura rodmani, the dorsomesal margin is rather smooth and triangular. The ventrolateral processes of the 2 species also differ: Itaura tusci has a small upper and more elongate lower process; Itaura rodmani has a single, short, digitate process. Itauara blahniki also has upturned, tusk-like parameres, but is easily distinguished from Itaura tusci based on differences in tergum X and the phallicata.
Adult. Body, wings, and appendages pale or tawny brown, often intermingled with rufous or golden hairs, tibia and tarsi tawny brown. Wings with conspicuous white spot at the arculus. Forewing slightly broader past anastomosis, but with margins nearly parallel, apex rounded. Forewing venation incomplete, with apical forks I, II, and III present; Sc and R1 distinct along their entire lengths; fork I sessile; fork II petiolate, stem shorter than fork; fork III petiolate, stem about the same length as fork; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present along Cu2; A3 absent; crossveins forming a relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing margins nearly parallel, tapering only slightly past anastomosis; apical forks II, III, and V present; Sc and R1 fused basally; A2 absent. Tibial spurs 1, 4, 4, foretibial spur extremely reduced and hairlike. Sixth sternal process thumb-like, apex attenuate and pointed, associated with weak oblique apodeme posteriorly.
Male genitalia. Preanal and inferior appendages absent. Segment IX dorsally narrow, broad medially and ventrally; anterior margin rounded; posterolateral margin membranous or very lightly sclerotized; sternum IX without modification. Tergum X incompletely fused to tergum IX with membrane or lightly sclerotized region ventrolaterally; dorsomesal margin slightly produced with several small irregular setose processes; dorsolateral margin with paired small, slightly down-turned, setose process; ventrolateral margin with 2 pairs of processes, the upper a small lobe-like process, the lower an elongate finger-like process bearing a few setae. Parameres present, paired, arising ventrobasally from fused endotheca and phallobase, sclerotized and rod-like, tusk-like, strongly curving upward, apex pointed. Phallobase reduced, lightly sclerotized with phallic shield. Phallicata forming a long lightly sclerotized dorsal sheath extending from phallobase, rugous distally, strongly curving upward with apex directed anterodorsally, with pair of broad, sclerotized blade-like lateral flanges, apex pointed and directed posteriorly. Endophallus membranous, enlarged and convoluted when invaginated, with 1 upper and 1 lower lobe.
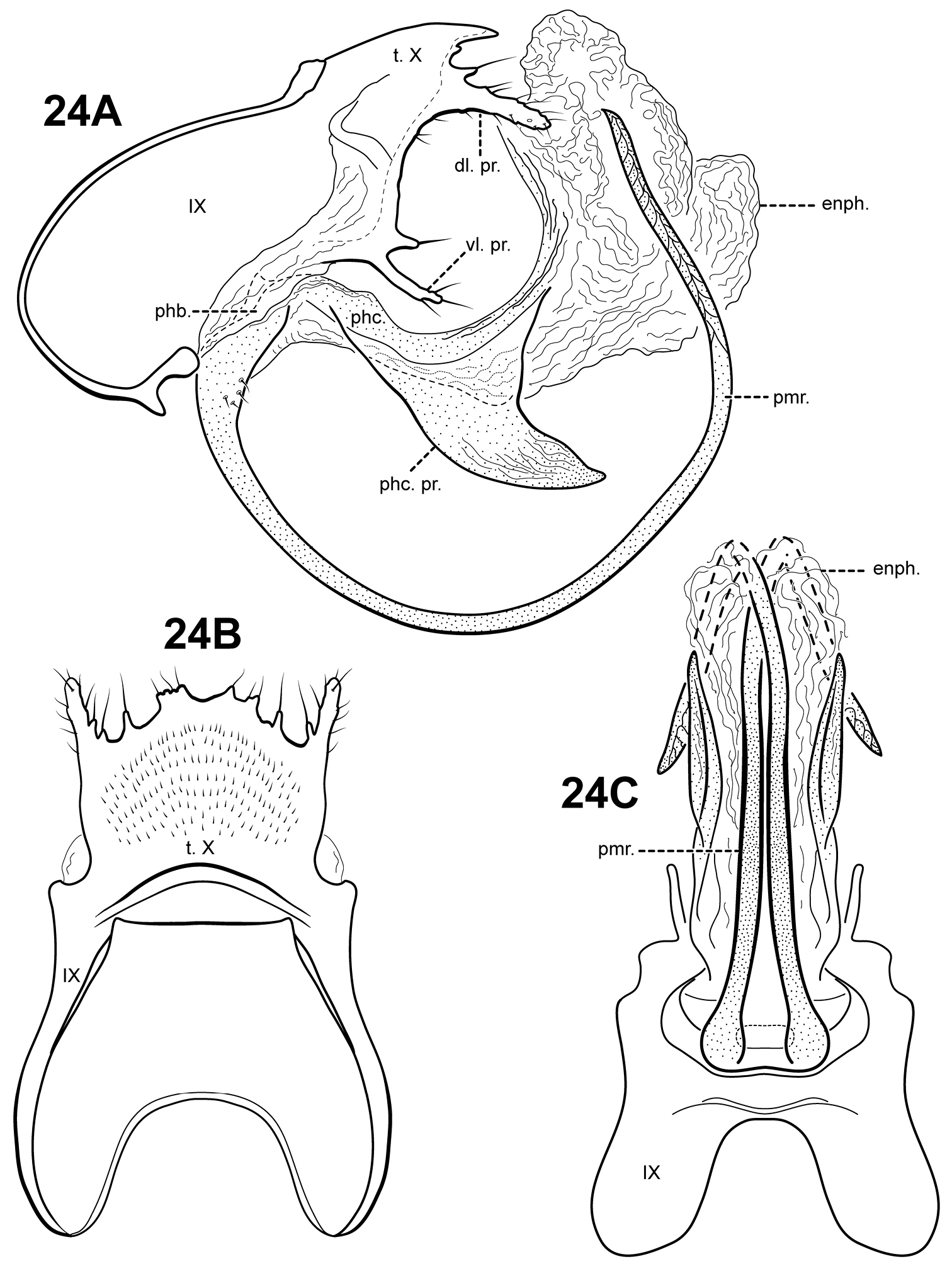
Itauara tusci sp. n. (UMSP000070932). Male genitalia: (A) lateral; (B) dorsal; (C) ventral. Abbreviations: dl. pr. = dorsolatera; process; enph. = endophallus; phb = phallobase; phc. = phallicata; phc. pr. phallicata process; pmr . = paramere; t. X = tergum X; vl. pr. = ventrolateral process.
Holotype male: BRAZIL: Rio de Janeiro: Rio das Flores, Macaé de Cima, 10 km SE Mury, 1000 m, 9.iii.2002 (Holzenthal, Blahnik, Paprocki, Prather) (UMSP000070932) (MZUSP).
Paratypes: BRAZIL: Rio de Janeiro: same data as holotype — 4 males, 26 females (UMSP); Rio Macaé, Macaé de Cima, 22°23'41"S, 042°30'08"W, 1000 m, 8.iii.2002 (Holzenthal, Blahnik, Paprocki, Prather) — 2 males, 10 females (UMSP); Encontro dos Rios (Macaé/Bonito), 6 km S Lumiar, 22°23'29"S, 042°18'42"W, 600 m, 10.iii.2002 (Holzenthal, Blahnik, Paprocki, Prather) — 64 males, 145 females (MZUSP, UMSP).
The name tusci is derived from the Old English word for tusk, and refers to the extremely long parameres of this species.
urn:lsid:zoobank.org:act:63403AB7-370C-4DA2-8628-C630D0C9C1E6
http://species-id.net/wiki/Itauara_unidentata
Fig. 2C, 25A–CThis species can be diagnosed by its large, tooth-like paramere process, and broad inferior appendage process. It is most similar to Itaura bidentata sp. n., which has a similarly shaped tergum X, dorsomesal spine, and apical sclerites. The 2 species can be separated by their paramere processes; in Itaura unidentata the paramere consists of a single large tooth-like process, whereas in Itaura bidentata, the paramere process is bifid. Itauara amazonica also has a dorsomesal spine, but can be distinguished from Itaura unidentata by the simple shape of tergum X and parameres. Itauara spiralis sp. n., has a similarly shaped tergum X, but is easily distinguished from Itaura unidentata by differences in the shape of the inferior appendage process, parameres, and phallicata.
Adult. Body, wings, and appendages pale or tawny brown in alcohol. Forewing slightly broader past anastomosis, but with margins nearly parallel, apex subacute. Forewing venation incomplete, with apical forks I, II, III, and IV present; Sc and R1 distinct along their entire lengths; fork I sessile; fork II sessile; fork III petiolate, stem longer than fork; fork IV petiolate, stem slightly shorter than fork; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present along Cu2; A3 absent; crossveins forming a relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing narrow and slightly scalloped past anastomosis; apical forks II and V present; Sc and R1 fused basally; A2 absent. Tibial spurs 1, 4, 4, foretibial spur extremely reduced and hairlike. Sixth sternal process short and digitate, apex attenuate and pointed, associated with strong oblique apodeme posteriorly.
Male genitalia. Preanal appendages absent. Segment IX dorsally and ventrally narrow, broad medially; anterior margin rounded; posterolateral margin membranous or very lightly sclerotized; sternum IX without modification. Tergum X incompletely fused to tergum IX with membrane or lightly sclerotized region ventrolaterally; dorsomesal margin straight, without processes; dorsolateral margin with paired elongate, down-turned, finger-like process; ventrolateral margin with paired, broad flange-like setose process consisting of several small irregular lobes. Inferior appendages present as single, broad, irregular setose process, broadest basally, fused to phallobase ventrobasally, bearing a single pair of small digitate lobes ventrolaterally, each bearing a seta. Parameres present, paired, arising laterally from endotheca, strongly sclerotized, large tooth-like process, curving ventrally and outward, apex pointed. Phallobase reduced, lightly sclerotized dorsally, laterally membranous, with 2 irregular and elongate sclerites arising basolaterally. Phallicata forming a short slerotized dorsal sheath with an elongate dorsomesal spine arising posteriorly to phallobase.
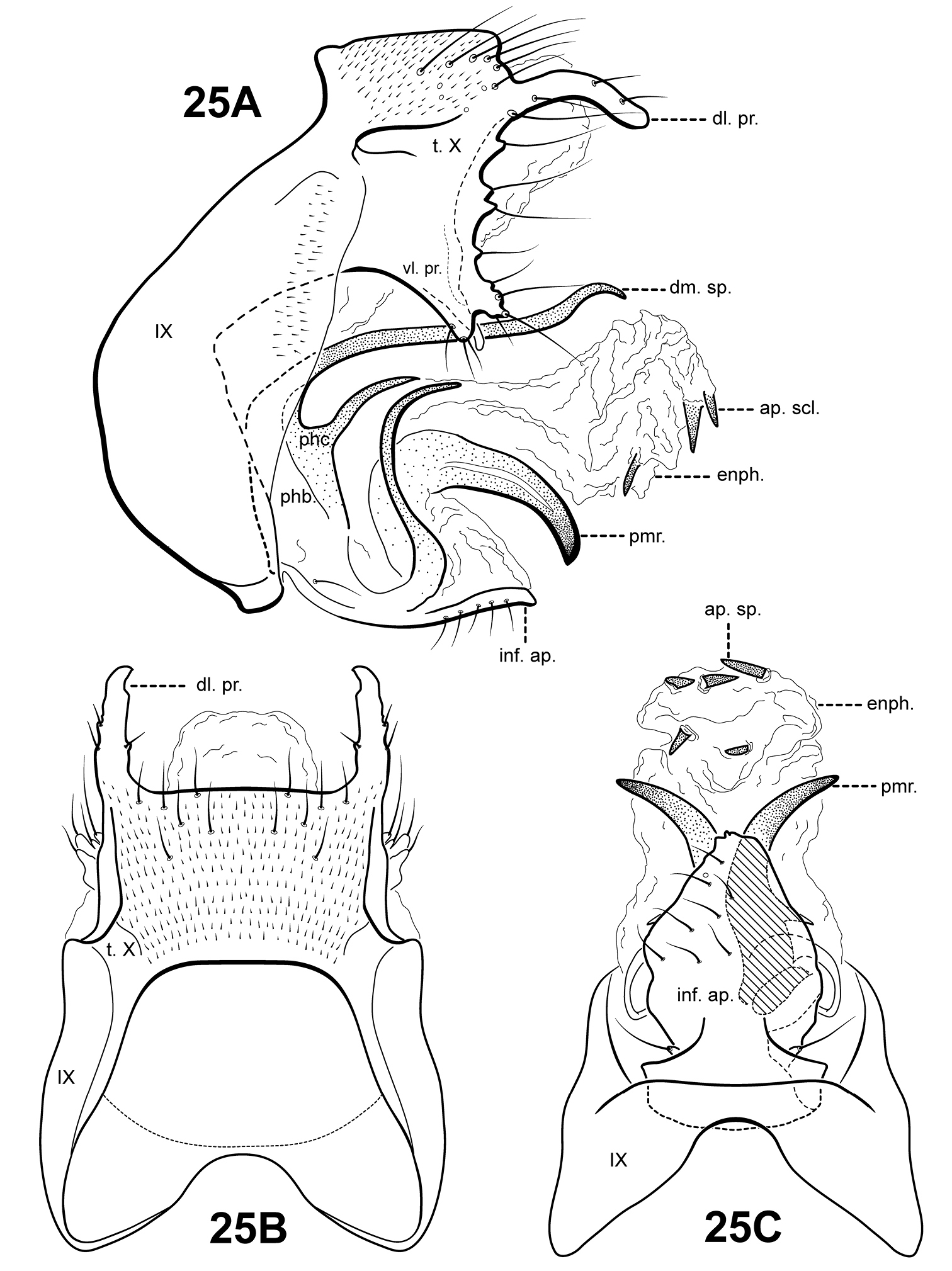
Itauara unidentata sp. n. (UMSP000118535). Male genitalia (A) lateral; (B) dorsal; (C) ventral. Abbreviations: ap. sp. = apical spine; dl. pr. = dorsolateral process; dm. sp. = dorsomesal spine; enph. = endophallus; inf. ap. = inferior appendage process; phb. = phallobase; phc. = phallicata; pmr . = paramere; t. X = tergum X; vl. pr. = ventrolateral process.
Holotype male: GUYANA: Kanuku Mountains: Kumu River & Falls, 03°15'54"N, 059°43'30"W, 28–30.iv.1995 (W.N. Mathis) (UMSP000118535) (NMNH).
Paratypes: GUYANA: Kanuku Mountains: same data as holotype, (W.N. Mathis) — 1 female (NMNH); same, (O.S. Flint) — 1 male (NMNH).
The name unidentata is suggested by the single tooth-like paramere process.
Key to males of Itauara
In most cases, it should be possible to identify most species by simple comparisons to illustrations and reference to the species diagnoses and descriptions. The following key is meant to help the user focus on male genitalic features most useful in identifying species and should be used in conjunction with the provided illustrations and descriptions.
| 1 | Inferior appendages present as single, sometimes apically bifid, setose process, fused to phallobase ventrobasally (Figs 4–6, 10, 13, 14, 16–18, 22, 23, 25) | 2 |
| – | Inferior appendages absent (Figs 7–9, 11, 12, 15, 19–21, 24) | 13 |
| 2(1) | Inferior appendage process relatively short and broad (Figs 4C, 6C, 13C, 14C, 16C, 22C, 25C) | 3 |
| – | Inferior appendage process relatively narrow and elongate (Figs 5D, 10C, 17C, 18C, 23C) | 9 |
| 3(2) | Tergum X dorsolateral margin with paired, very long, finger-like processes; phallicata with elongate dorsomesal spine (Figs 6, 22, 25) | 4 |
| – | Tergum X dorsolateral margin without paired long, finger-like processes; phallicata without dorsomesal spine (Fig. 4, 13, 14, 16) | 6 |
| 4(3) | Inferior appendage process bifid apically; endophallus without apical sclerites; parameres rather elongate, spiral-shaped, curving nearly 360 degrees (Fig. 22) | Itauara spiralis sp. n. |
| – | Inferior appendage process not bifid; endophallus with apical sclerites; parameres rather broad and tooth-like, not spiral-shaped (Figs 6, 25) | 5 |
| 5(4) | Parameres bifid (Fig. 6) | Itauara bidentata sp. n. |
| – | Parameres not bifid (Fig. 25) | Itauara unidentata sp. n. |
| 6(3) | Tergum X with elongate, attenuate, downturned dorsomesal process (Figs 4A–B, 13A–B, 16A–B) | 7 |
| – | Tergum X dorsomesal margin bifid (Fig. 14) | Itauara jamesii sp. n. |
| 7(6) | Inferior appendage process bifid apically (Fig. 4C) | Itauara alexanderi sp. n. |
| – | Inferior appendage process not bifid (Figs 13C, 16C) | 8 |
| 8(7) | Parameres extremely sinuous, not forked; endophallus with large, bifid apical processes; tergum X ventrolateral margin with small, subtriangular setose process (Fig. 13) | Itauara guyanensis sp. n. |
| – | Parameres forked, not sinuous; endophallus entirely membranous without apical processes or sclerites; tergum X ventrolateral margin with broad, irregular, flange-like setose process (Fig. 16) | Itauara lucinda sp. n. |
| 9(2) | Tergum X dorsomesal margin divided apicomesally, slightly notched, or with large, prominent bifid process; inferior appendage process not bifid (Figs 5, 17, 18) | 10 |
| – | Tergum X dorsomesal margin not divided apicomesally, bifid, slightly notched, or with large, prominent bifid process; inferior appendage process bifid apically (Figs 10, 23) | 12 |
| 10(9) | Parameres curving upward or tusk-like; phallicata with elongate, apically bifid, dorsomesal spine (Fig. 5) | Itauara amazonica (Flint 1971) |
| – | Parameres not curving upward or tusk-like; phallicata without dorsomesal spine (Figs 17, 18) | 11 |
| 11(10) | Parameres spiral-shaped or ram-like, curving nearly 360 degrees; endophallus largely membranous, without apical sclerite (Fig. 17) | Itauara ovis sp. n. |
| – | Parameres nearly straight; endophallus lightly sclerotized, with small apical sclerite (Fig. 18) | Itauara peruensis sp. n. |
| 12(9) | Tergum X with elongate, attenuate, downturned dorsomesal process (Fig. 23) | Itauara stella sp. n. |
| – | Tergum X with large, blunt, dorsomesal process, elongate in dorsal view, subtriangular in lateral view (Fig. 10) | Itauara emilia sp. n. |
| 13(1) | Phallicata with paired lateral flanges or processes (Figs 9A, 11A, 12B, 19B, 20B, 24A) | 14 |
| – | Phallicata without paired lateral flanges or processes (Figs 7A, 8A, 15A, 21A) | 19 |
| 14(13) | Tergum X dorsomesal margin irregular, with several small setose processes (Figs 9B, 24B) | 15 |
| – | Tergum X dorsomesal margin not irregular (Figs 11A–B, 12B–C) | 16 |
| 15(14) | Parameres curving upward, tusk-like; endophallus without apical processes or sclerites (Fig. 24) | Itauara tusci sp. n. |
| – | Parameres arcuate, curving downward; endophallus with tooth-like apical sclerite (Fig. 9) | Itauara charlotta sp. n. |
| 16(14) | Tergum X with elongate, attenuate, downturned dorsomesal process (Figs 11A–B, 12B–C) | 17 |
| – | Tergum X without elongate dorsomesal processes (Figs 19B–C, 20B–C) | 18 |
| 17(16) | Parameres sinuous; phallicata very lightly sclerotized basally, rugous or membranous distally (Fig. 12) | Itauara guarani (Angrisano 1993) |
| – | Parameres arcuate, curving downward; phallicata entirely sclerotized (Fig. 11) | Itauara flinti sp. n. |
| 18(16) | Parameres curving upward, tusk-like; phallicata strongly curved medially, directed anterodorsally (Fig. 20) | Itauara rodmani sp. n. |
| – | Parameres sinuous; phallicata nearly straight, distal portion slightly upturned (Fig. 19) | Itauara plaumanni (Flint 1974) |
| 19(13) | Sternum IX bearing 2 pairs of extremely elongate, seta-like processes; parameres vestigial, consisting only of a pair of small, digitate setose lobes arising ventrolaterally from endotheca (Fig. 8) | Itauara brasiliana (Mosely 1939) |
| – | Sternum IX without modification; parameres prominently present (Figs 7, 15, 21) | 20 |
| 20(19) | Tergum X with elongate, attenuate, slightly downturned dorsomesal process; parameres curving upward, tusk-like (Fig. 7) | Itauara blahniki sp. n. |
| – | Tergum X without elongate dorsomesal processes; parameres not curving upward or tusk-like (Figs 15, 21) | 21 |
| 21(20) | Tergum X dorsomesal margin blunt; parameres bent basally at nearly 90 degree angle, directed dorsally; phallicata sclerotized, not continuous from phallobase (Fig. 15) | Itauara julia sp. n. |
| – | Tergum X dorsomesal margin roof-like, strongly produced; parameres arcuate, curving downward; phallicata very lightly sclerotized basally, more membranous and rugous distally, continuous from phallobase (Fig. 21) | Itauara simplex sp. n. |
We are thankful to Len Ferrington, George Weiblen, and Susan Weller (University of Minnesota) for helpful comments and suggestions on earlier drafts of this manuscript. We thank Roger Blahnik (U. of MN) for his valuable insight regarding protoptiline morphology, systematics, and evolution. We express our appreciation to Ivalio Stoyanov, ZooKeys Editorial Secretary, C.J. Geraci, ZooKeys Guest Editor, and 2 anonymous reviewers for their helpful comments. We also thank Jolanda Huisman, John Luhman, Steffen Pauls, and Claire Serieyssol for translating various taxonomic works. Julie Martinez rendered the beautiful full habitus illustration. DRR thanks the following individuals for assistance throughout this project: Roger Blahnik, Phil Clausen, Lourdes Chamorro, John Luhman, Patina Mendez, Henrique Paprocki, Steffen Pauls, Greg Setliff, Robin Thomson, Anne Wasmund, and Jen Zaspel. DRR is grateful to her parents, Jim & Charlotte Robertson, for the financial assistance they provided which allowed her to complete this work. She also extends her deepest gratitude and affection to her husband, Alex Thompson, and their daughters, Stella & Lucinda.
We are grateful to the many curators, colleagues, and staff at several natural history collections around the world who loaned material: Elisa Angrisano (MACN), David Goodger (BMNH), and Oliver Flint (NMNH). We are particularly grateful for the assistance provided by Oliver Flint, National Museum of Natural History, Smithsonian Institution. Many of the new species described here were collected by Dr. Flint and other researchers at the NMNH.
DRR is grateful for the following funding sources received as a graduate student: Alexander and Lydia P. Anderson Fellowship (University of Minnesota, Graduate School), Doctoral Dissertation Fellowship (U. of MN, Graduate School), Dayton & Wilkie Natural History Fund (U. of MN, Bell Museum), Boesel-Sanderson Natural History Award (North American Benthological Society), Student Conservation Award (North American Benthological Society), NSF GK-12 Fellowship (U. of MN, Bell Museum), Morris & Elaine Soffer Rockstein Fellowship (Dept. of Entomology, U. of MN), MN Futures grant (to D. Andow), Walter Judd International Fellowship (U. of MN, Graduate School), and U. of MN, Graduate School Block travel grants. This material is based upon work supported by the National Science Foundation under Grant Nos. DEB 0117772 and 9971885 (to RWH).