ZooKeys 111: 59–65, doi: 10.3897/zookeys.111.813
A new species of Amphoropsyche (Trichoptera, Leptoceridae) from Ecuador, with a key to the species in the genus
Ralph W. Holzenthal1,†, Luis Ernesto Rázuri-Gonzales2,‡
1 University of Minnesota, Department of Entomology, 1980 Folwell Ave., 219 Hodson Hall, St. Paul, Minnesota 55108, U.S.A.
2 Departamento de Entomología, Museo de Historia Natural, Universidad Nacional Mayor de San Marcos, Av. Arenales 1256, Apartado Postal 14–0434, Lima, Perú
Corresponding author: Ralph W. Holzenthal (holze001@umn.edu).
Academic editor: Christy Geraci
Abstract
A new species of Amphoropsyche
Holzenthal is described from Ecuador. It is similar to a group of
species with dorsomesal processes on the preanal appendages (i.e., Amphoropsyche woodruffi Flint & Sykora, Amphoropsyche refugia Holzenthal, and Amphoropsyche aragua
Holzenthal), but can be distinguished from these and other members of
the genus by the short, digitate dorsomesal processes on the preanal
appendages and the broad lateral processes of tergum X of the male
genitalia. A key to males of the 14 species now known in the genus is
presented based on characters of the genitalia.
KeywordsTrichoptera, Leptoceridae, Amphoropsyche, caddisfly, new species, Neotropics, Ecuador, key to species
Introduction
Flint (1968) described the male, female, and larva of a new species of longhorn caddisfly (Leptoceridae, Leptocerinae) from Dominica, Lesser Antilles, and tentatively placed it in the Chilean genus Brachysetodes,
based mainly on wing venation. He also noted that the structure of the
male genitalia seemed quite different between the Dominican species, Brachysetodes insularis Flint, and the type species, Brachysetodes trifidus
Schmid. He suggested that the discovery of the larva of the Chilean
species might show that the 2 species are not congeneric. Based on
character differences in the male genitalia, Holzenthal (1985) removed Brachysetodes insularis from Brachysetodes and established the genus Amphoropsyche for Flint’s species along with 9 new species from Ecuador, Colombia and Venezuela. In a separate paper, Holzenthal (1986a) described the immature stages of Brachysetodes, confirming Flint’s earlier supposition that Brachysetodes and Amphoropsyche are distinct. Holzenthal (1986b)
also described 1 additional species from Bolivia, extending the known
geographical range of the genus considerably southwards, and
redescribed the larva of the Dominican species. Since the mid 1980s,
only 3 additional nominal species were described, all from the Lesser
Antilles and Tobago: Amphoropsyche janstockiana Botosaneanu, 1990 from Saint Vincent, Amphoropsyche multispinosa Botosaneanu, 1993 (in Botosaneanu and Alkins-Koo 1993) from Trinidad, and Amphoropsyche woodruffi Flint & Sykora, 1993 from Grenada (subsequently recorded from northern Venezuela by Flint 1996). Flint (1996) considered Amphoropsyche multispinosa to be a geographical variant of Amphoropsyche woodruffi and changed its status to a subspecies of the latter. In the same paper, Flint (1996)
noted the presence of a single female specimen from Tobago with
distinctive genitalia probably representing yet another new species;
males have still not been collected and the species remains undescribed.
This genus seems to be especially species-rich in
mid-elevation streams (e.g., 1500–2500 m) in the northern Andes and
more species are expected to be collected and described (Holzenthal 1986b; Flint et al. 1999). Amphoropsyche
is characterized by the presence of large glands inside the preanal
appendages (probably producing pheromones), and the presence of a tuft
of strong hairs near the apex of the inferior appendage, probably
involved in dispersing these pheromones (Botosaneanu 1990). In this paper, we describe a new species of Amphoropsyche, the 14th in the genus, and provide a key to the males of the species.
Materials and methods
This study is based on a single pinned specimen collected
in Ecuador by Dr. Oliver S. Flint, Jr., National Museum of Natural
History, Smithsonian Institution, and kindly loaned to the first
author. Techniques and procedures used in the preparation and
examination of the specimen are those outlined by Blahnik and Holzenthal (2004) and Blahnik et al. (2007).
The illustration of the genitalia was prepared from a pencil sketch
made using a camera lucida mounted on an Olympus BX41 compound
microscope. The pencil sketch was then scanned and placed into an Adobe
Illustrator (version CS5, Adobe Systems, Inc.) document, to serve as a
template, and then traced to create a vector graphic illustration. A
graphic tablet and pen (BAMBOOTM, Wacom Technology Co.) facilitated
careful tracing of the original image.
Terminology used in describing male genitalia follows that of (Holzenthal (1985, 1986b). The taxonomic key was based on published illustrations and descriptions of the male genitalia (Holzenthal 1985, 1986b; Botosaneanu 1990; Flint and Sykora 1993, Botosaneanu and Alkins-Koo 1993; Flint 1996
[these papers can be downloaded from the Trichoptera Literature
Database at www.trichoplit.umn.edu to facilitate comparisons]) and was
constructed using the DELTA system which facilitated taxonomic data
coding via the Delta editor v. 1.04 (Dallwitz 1980; Dallwitz et al. 1999 onwards).
The type is deposited in the United States National
Museum of Natural History, Smithsonian Institution, Washington, D.C.
(NMHN).
Species description
Description.
This species is characterized by the short,
digitate dorsomesal processes of the inferior appendages, the long
basoventral projection of the 1st article of the inferior appendages,
and the broad, lateral processes of segment X. It is most similar to
that group of species also possessing dorsomesal processes on the
preanal appendages(i.e., Amphoropsyche woodruffi, Amphoropsyche refugia, Amphoropsyche aragua), but differs in having much shorter processes that are unsclerotized.
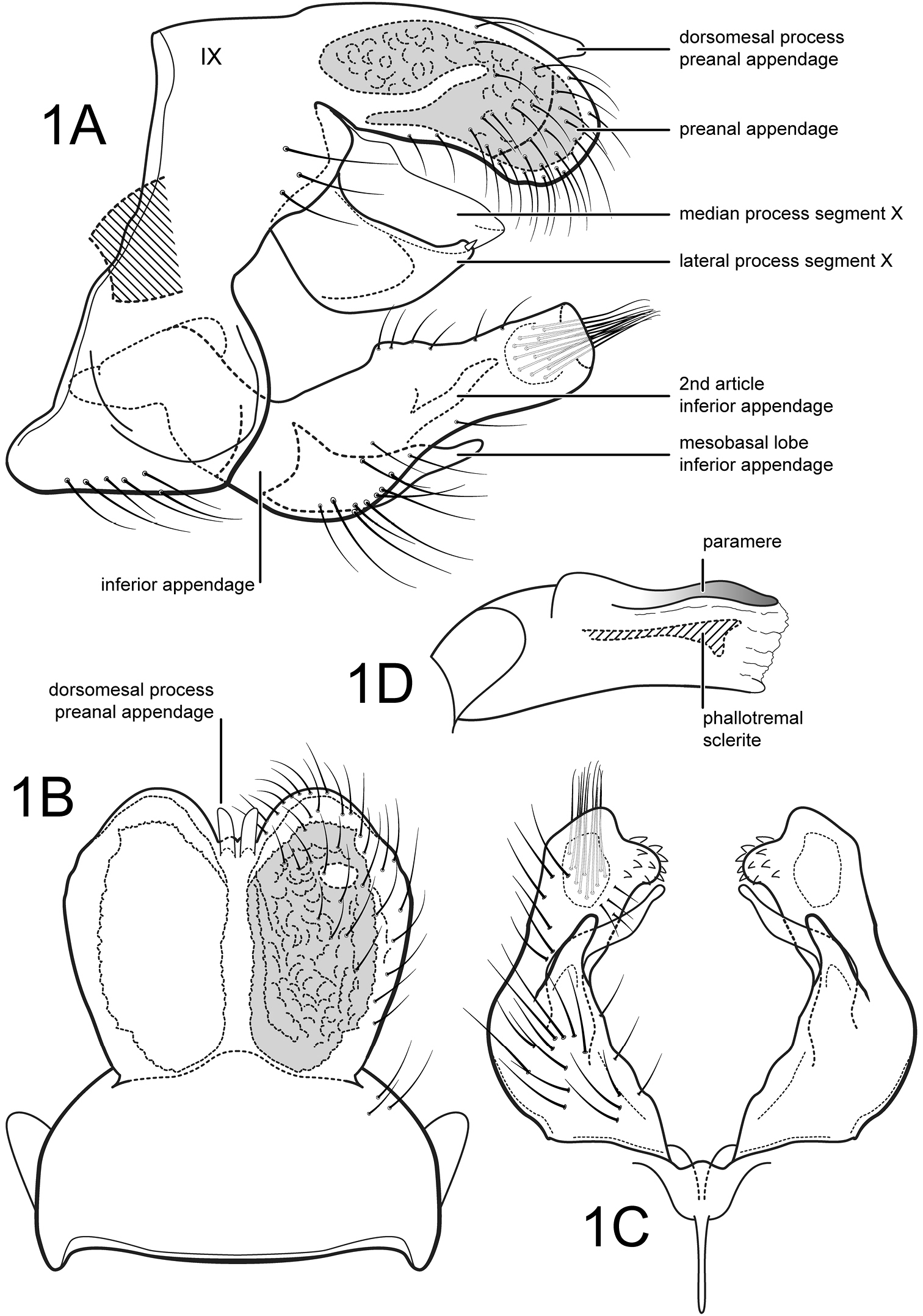
Male. Forewing length 4.8 mm. Wings and body color brown. Genitalia as in Fig. 1 A–D.
Segment IX annular, sternum with anterior part slightly extended
anteriorly. Segment X composed of a single mesal process and pair of
lateral processes; mesal process lightly sclerotized, apex broadly
acute; lateral process broad, bearing apical spine-like setae. Preanal
appendages large, oval, almost completely fused along their
midlengths, with pair of short, digitate, membranous dorsomesal
processes; preanal appendage with large reticulate internal gland with
small ventral opening. Inferior appendage elongate, with long
basoventral projection; inferior appendage angulate basally in lateral
view, bent inwards in ventral view, bearing very short spine-like
setae on slightly protruding apicomesal corner; 2nd article of inferior
appendage elongate, thin, sinuate, slightly curved inwards, apex
narrow, rounded. Phallic apparatus with phallobase well developed; pair
of dorsal parameres present; phallotremal sclerite well developed,
elongate, widest apically in lateral view.
Female and larva: Unknown.
Figure 1.
Amphoropsyche tandayapa, sp. n. Male genitalia A lateral B segments IX–X, dorsal C inferior appendages, ventral D phallus, lateral.
Holotype male.
ECUADOR: Pichincha: 2.3 km S Tandayapa, 1800 m, 6.x.1990, O.S. Flint, Jr. (NMNH).
Etymology.
Named after the town of Tandayapa, located near where the holotype was collected.
| 1 |
Preanal appendages completely (Holzenthal 1985, Figs 8B, 10B; Flint and Sykora 1993, Fig. 20) or almost completely fused mesally (if the latter, apical emargination shallow, obtuse) (Fig. 1B; Holzenthal 1985, Fig. 3B)
|
2 |
| – |
Preanal appendages not fused mesally, divided to 1/3 to 2/3 of their length (apical emargination acute) (Holzenthal 1985, Figs 5B, 6B)
|
6 |
| 2(1) |
Preanal appendages with dorsomesal process (Holzenthal 1985, Figs 8B, 10B)
|
3 |
| – |
Preanal appendages without dorsomesal process (Holzenthal 1985, Figs 3A–D)
|
Amphoropsyche insularis
|
| 3(2) |
Dorsomesal process of preanal appendages
short, digitate, not exceeding length of preanal appendage; dorsomesal
processes of preanal appendages not sclerotized (Figs 1 A–D)
|
Amphoropsyche tandayapa sp. n.
|
| – |
Dorsomesal process of preanal appendages long, exceeding length of preanal appendage (Holzenthal 1985, Figs 8A, 10A; Flint and Sykora 1993, Fig. 18); dorsomesal processes of preanal appendages sclerotized
|
4 |
| 4(3) |
Second article of inferior appendages elongate, narrow (Holzenthal 1985, Fig. 8A)
|
5 |
| – |
Second article of inferior appendages short (Flint and Sykora 1993, Figs 18–20; Botosaneanu and Alkins-Koo 1993, Figs 97–101)
|
Amphoropsyche woodruffi
|
| 5(4) |
Dorsomesal process of preanal appendages bifid in dorsal view; ventral subterminal portion of phallobase serrate (Holzenthal 1985, Figs 8A–D)
|
Amphoropsyche refugia
|
| – |
Dorsomesal process of the preanal appendages entire in dorsal view; ventral subterminal portion of phallobase entire (Holzenthal 1985, Figs 10A–D)
|
Amphoropsyche aragua
|
| 6(1) |
Second article of inferior appendages present (Holzenthal 1985, Fig. 5A)
|
7 |
| – |
Second article of inferior appendages absent (Holzenthal 1985, Fig. 16C)
|
13 |
| 7(6) |
Tergum X with median process and paired lateral processes (Holzenthal 1985, Figs 5A, 14A)
|
8 |
| – |
Tergum X without median process, lateral processes with apical and subapical spinelike projections (Botosaneanu 1990, Figs 1–3)
|
Amphoropsyche janstockiana
|
| 8(7) |
Second article of inferior appendages short (Holzenthal 1985, Fig. 14C) or long, but broad (Holzenthal 1985, Fig. 6C)
|
9 |
| – |
Second article of inferior appendages elongate and narrow (Holzenthal 1985, Fig. 7C)
|
11 |
| 9(8) |
Phallus without parameres (Holzenthal 1985, Fig. 6D)
|
10 |
| – |
Phallus with parameres (Holzenthal 1985, Figs 14A–D)
|
Amphoropsyche quebrada
|
| 10(9) |
Second article of inferior appendages
short, with apical spine-like seta; lateral process of tergum X with
subapical spine-like seta; phallicata with pair of bifid, spiniferous,
lateral extensions (Holzenthal 1986b, Figs 1A–D)
|
Amphoropsyche spinifera
|
| – |
Second article of inferior appendages
long, but broad, without apical spine-like seta; lateral process of
tergum X with several apical spine-like setae; phallicata without
lateral, bifid extensions, but phallobase with ventral spine-like
process (Holzenthal 1985, Figs 6A–D)
|
Amphoropsyche flinti
|
| 11(8) |
Phallus with parameres (Holzenthal 1985, Fig. 5D)
|
12 |
| – |
Phallus without parameres (Holzenthal 1985, Figs 11A–D)
|
Amphoropsyche choco
|
| 12(11) |
Lateral process of tergum X U-shaped, tip bifid, bearing small spine-like setae (Holzenthal 1985, Figs 5A–D)
|
Amphoropsyche napo
|
| – |
Lateral process of tergum X tapered to a sharp terminal point, without spine-like setae (Holzenthal 1985, Figs 7A–D)
|
Amphoropsyche stellata
|
| 13(6) |
Parameres small; inferior appendage with basoventral lobe (Holzenthal 1985, Figs 16A–D)
|
Amphoropsyche cauca
|
| – |
Parameres large; inferior appendage without basoventral lobe (Holzenthal 1985, Figs 12A–D)
|
Amphoropsyche ayura
|
Acknowledgements
We would like to thank Dr. Oliver S. Flint, Jr.,
National Museum of Natural History, Smithsonian Institution, for the
kind loan of the material on which this paper is based. The second
author would like to thank the Consejo Nacional de Ciencia, Tecnología e
Innovación Tecnológica (CONCYTEC) for supporting his visit to the
Department of Entomology, University of Minnesota, where the research
was accomplished. We thank Dr. Christy Geraci and 2 anonymous reviewers
for their helpful comments on the manuscript.
ReferencesBlahnik
RJ, Holzenthal RW (2004) Collection and curation of Trichoptera, with
an emphasis on pinned material. Nectopsyche, Neotropical Trichoptera
Newsletter 1:8-20.
Blahnik RJ, Holzenthal RW, Prather AL (2007)
The lactic acid method for clearing Trichoptera genitalia. In:
Bueno-Soria J, Barba-Álvarez R, Armitage BJ (Eds) Proceedings of the
12th International Symposium on Trichoptera. The Caddis Press,
Columbus, Ohio, 9–14.
Botosaneanu L (1990) A second Antillan species
of Amphoropsyche Holzenthal, 1985 (Trichoptera, Leptoceridae).
Bijdragen tot de Dierkunde 60:319-322.
Botosaneanu L, Alkins-Koo M (1993) The caddis
flies (Insecta: Trichoptera) of Trinidad and Tobago, West Indies.
Bulletin de l’Institut des Sciences Naturelles de Belgique, Entomologie
63:5-45.
Dallwitz MJ (1980) A general system for coding taxonomic descriptions. Taxon 29:41-46. doi:
10.2307/1219595
Dallwitz MJ, Paine TA, Zurcher EJ (1999 onwards) User’s guide to the DELTA editor. Available from:
http://delta-intkey.com/ [accessed 15 December 2010].
Flint OS, Jr. (1968)
Bredin-Archbold-Smithsonian Biological Survey of Dominica, 9. The
Trichoptera (caddisflies) of the Lesser Antilles. Proceedings of the
United States National Museum 125:1-86.
Flint OS, Jr. (1996) Studies of Neotropical
caddisflies LV: Trichoptera of Trinidad and Tobago. Transactions of the
American Entomological Society 122:67-113.
Flint OS, Jr., Holzenthal RW, Harris SC
(1999) Catalog of the Neotropical Caddisflies (Trichoptera). Special
Publication, Ohio Biological Survey, Columbus, Ohio, 239 pp.
Flint OS, Jr., Sykora JL (1993) New species
and records of caddisflies (Insecta: Trichoptera) from the Lesser
Antilles, with special reference to Grenada. Annals of Carnegie Museum
62:47-62.
Holzenthal RW (1985) Studies in Neotropical
Leptoceridae (Trichoptera) II: Amphoropsyche, a new genus and species
of Leptocerinae from northern South America. International Journal of
Entomology 27:255-269.
Holzenthal RW (1986a) Studies in Neotropical
Leptoceridae (Trichoptera), IV: a revision of Brachysetodes Schmid.
Transactions of the American Entomological Society 111:407-440.
Holzenthal RW (1986b) Studies in Neotropical
Leptoceridae (Trichoptera), a new species of Amphoropsyche, with a
redescription of the immature stages of A. insularis (Flint). Annals of
the Entomological Society of America 79:251-255.