





A new genus and species of pseudosigmurethrous orthurethran pulmonate of the family Cerastidae, Darwininitium shiwalikianum gen. n. and sp. n. is described from the Lesser Himalaya of Nepal. It represents the first record of an orthurethran with a fully developed pseudosigmurethrous pallial system, having a completely closed secondary ureteric system. Biogeographically this new taxon provides a significant range extension for the family north of the previously known distribution range.
Pseudosigmurethrous, Orthurethran, Pulmonata, Darwininitium, Nepal
There are currently some 14 genera recognised in the orthurethran land snail family Cerastidae. Their greatest diversity is found in the Afro-tropical zone, but the family extends eastwards into the Indian subcontinent and beyond, and one genus, Amimopina, occurs in Australia and New Guinea (
The family Cerastidae Wenz, 1923 was previously included in the Enidae sensu lato (Enoidea sensu
The principal distinguishing anatomical feature of cerastids is the excretory system which, whilst generally being of the normal orthurethran form comprising an elongate kidney with a straight primary ureter running directly towards the pneumostome, has developed what
Here we present an account of a new genus and species of cerastid from the Lesser Himalaya of Central Nepal. Along its E-W axis, Nepal is divided into five east-west parallel physiographic elevational regions i.e. from south (lower) to north (higher): 1) Tarai (southern flat belt from 67 m – 300 m), 2) Shiwalik (from 300 m – 1000 m), 3) Mid Hill (1000 m – 3000 m), 4) Mid Mountain (3000 m – 5000 m) and 5) High Himalaya (> 5000 m). Our surveys were conducted in these five zones in Central Nepal but the new genus was reported only from the Shiwalik range (Figure 1). The type material has been deposited at the museum of the Central Department of Zoology, Tribhuvan University (CDZTU), Kathmandu, Nepal.
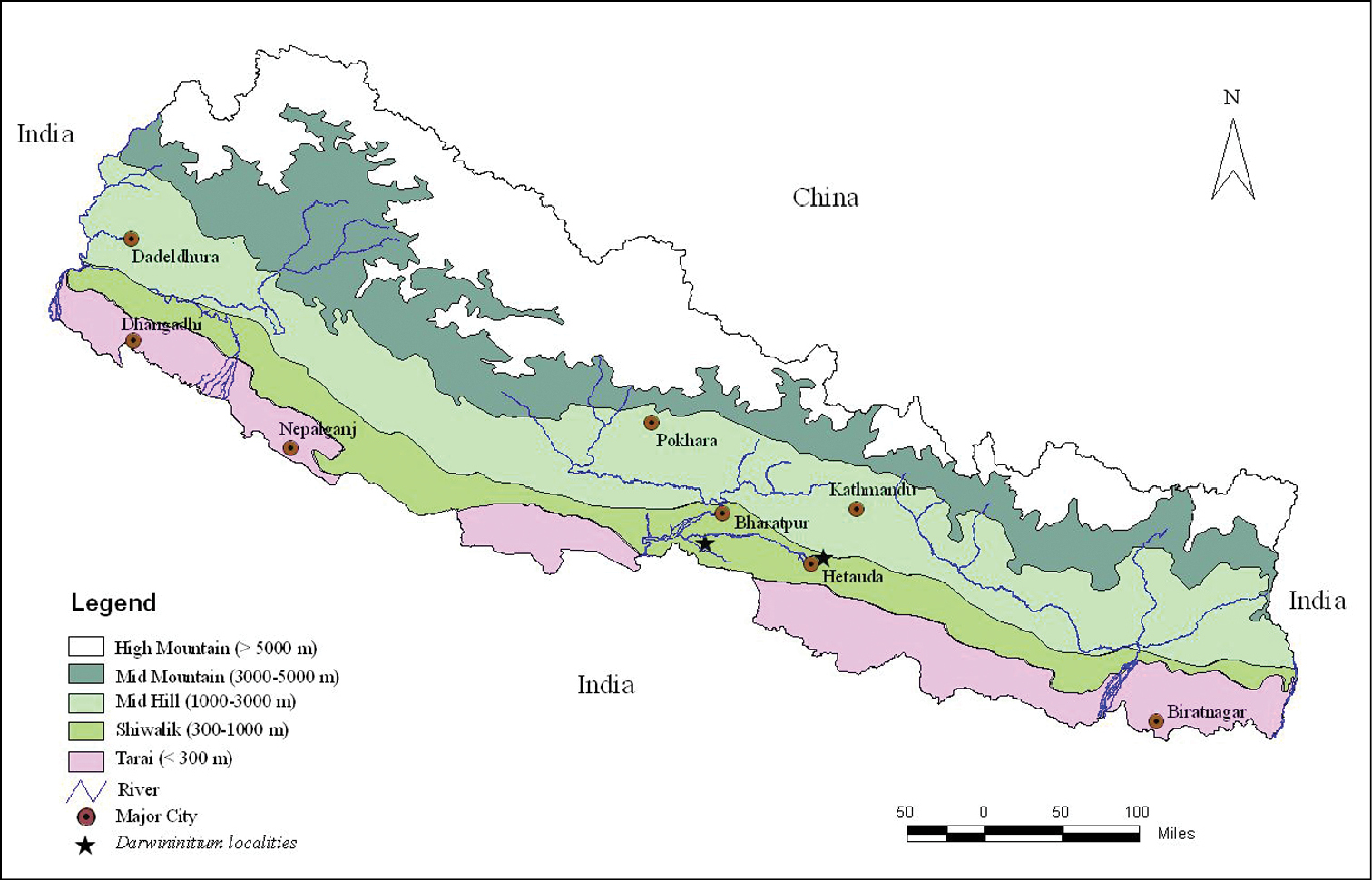
Map of Nepal showing the collection localities of Darwininitium shiwalikianum sp. n.
Shell measurements are expressed in mm as follows: shell height × max. shell width × min. shell width. Abbreviation for internal parts used are; AG- albumen gland, ATR- atrium, AU- auricle, E- epiphallus, EC- epiphallar caecum, GS- gametolytic sac, HD- hermaphrodite duct, KI- kidney, P- penis, PI- pilaster, PNE- pnerumostome, PR- penis retractor, REU- renal branch of secondary ureter, RT- rectum, RTU- rectal branch of secondary ureter, SO- spermoviduct, UO- ureteric orifice, VD- vas deferens and VE- ventricle.
Taxonomic treatmenturn:lsid:zoobank.org:pub:D670F9B7-1061-4686-B54B-54B4A1DBD4D5
Darwininitium shiwalikianum Budha & Mordan, 2012, sp. n.
urn:lsid:zoobank.org:act:4C8FA14B-A9CA-49D6-A91A-B532B2EDBAEE
Holotype CDZTU0114: Kasara near Tamor Lake, Chitwan National Park, Central Nepal, ca 210 m. 27°61'10"N, 85°20'05"E, sal (Shorea robusta) forest, leg P.B. Budha 5 May 2008. Paratype from the type locality, CDZTU0114a/1 shell. Other paratypes CDZTU0115/2: Taubas along the road side of Tribhuvan Highway, Bhainse, left bank of Rapti river, Makwanpur [=district], Central Nepal, ca 520 m. 27°30'13"N, 85°02'59"E, 2 live. Mixed riverine forest, leg P.B. Budha 7 May 2008.
Cerastidae with a fully pseudosigmurethrous pallial system including fully enclosed renal and rectal branches of ureter; penial appendix lacking, which is present in all known cerastid genera. Shell carinated, white flecks on a dark-brown background.
Shell (Figure 2a, 2b): Dextral, globosely turbinate, weakly carinated, rimately perforate, whorls 5, apex blunt, suture shallow, chestnut with wide and irregular shaped radial white patches both in upper- and under- surface, first two whorls smooth but under magnification (60×) shallow dotted wrinkles visible, later whorls with weak radial striae, aperture ovate, peristome thin and reflected slightly descended toward the aperture, columellar margin reflected covering nearly half the umbilicus.

Shell of Darwininitium shiwalikianum sp. n. 2a holotype CDZTU0114 2b paratype CDZTU0115
(mm). Holotype: 13.6 × 15.2 × 12.6, aperture 7.9 × 8.0, whorls 5, paratype from the type locality 14.1 × 13.8 × 12.5, whorls 5, aperture 7.8 × 7.9, paratypes from Taubas 11.7 × 12.5 × 10.6, aperture 6.9 × 7.5, whorls 4.5, 12.3 × 15.7 × 12.9, aperture 8.1 × 10.5, wh. 4.5.
The genus is named as a tribute to the Darwin Initiative for having supported land snail projects and in particular for supporting the senior authors’ participation in the project Developing land snail expertise in South and Southeast Asia from 2006-2011. The species name derives from the Lesser Himalaya Shiwalik range, from where it was reported.
(Figures 3a, 3b). The anterior of the uniformly pale cream body can extend significantly more than 2× of the shell length.

Live animal of Darwininitium shiwalikianum, paratype CDZTU0115 3a anterior portion extended 3b animal in situ.
(Figure 4a). Kidney of the typical orthurethran type, running approximately four-fifths the length of the pulmonary cavity; a thin-walled, completely closed tube runs from the renal pore along the full length of the inner margin of the kidney, folding at the top to run a short distance towards the rectum, and then down along the inner margin of the rectum, again as a fully closed tube, almost as far as the anus and close to the pneumostome, where it opens with a slight flare. Pallial venation is prominent and a mantle gland is lacking.
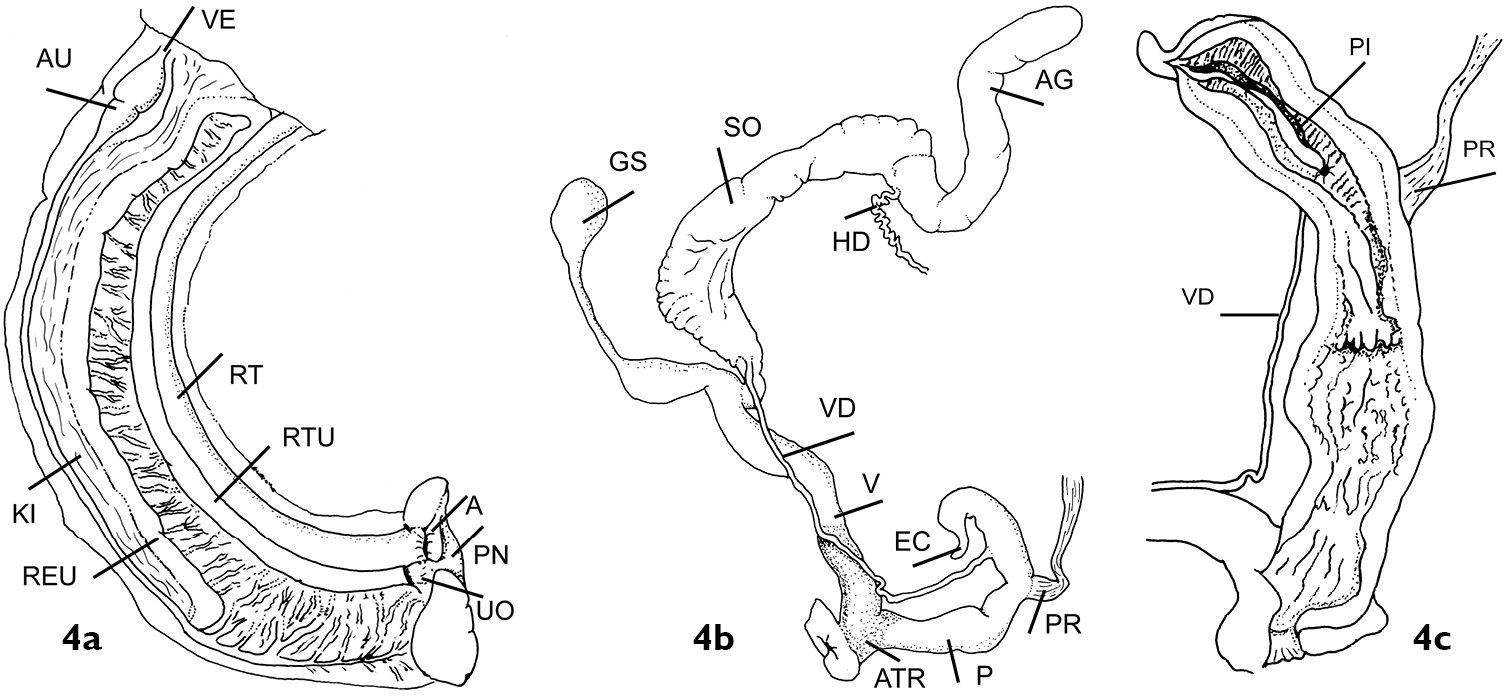
Genitalia in Darwininitium shiwalikianum sp. n. paratype CDZTU0115 4a Pallial cality 4b General view of genitalia 4c Interior of the penis
(Figures 4b, 4c). The female system has a well-developed gametolytic sac with a long peduncle with an expanded basal portion, and darkly pigmented spongy tissue in the atrium and at the base of the free-oviduct. The penis lacks an appendix, but has a prominent epiphallar caecum. The penial retractor inserts well below and opposite the point of insertion of the vas deferens. Internally the penis is separable into two regions, the lower having numerous longitudinal pilasters with a knobbly appearance. Above there is a slight constriction of the lumen by an undulating ring pilaster with a smooth, forked pilaster above, running downwards from the level of the opening of the vas deferens. The epiphallus has a thick pad of transverse, weakly ridged tissue running down its length more-or-less opposite the pore of the vas deferens, as well as a smooth longitudinal fold which runs down from within the caecum as far as the top of the vas deferens opening. There is no obvious penial sheath. The hermaphrodite duct lacks the clump of diverticulae characteristic of the Enidae sensu stricto.
The genus Darwininitium was collected in the Dun valley of the Lesser Himalaya in the Rapti river basin from an elevation of ca 210 m in Chitwan National Park and extending northward to Taubas, Bhainse Makwanpur, Central Nepal at an elevation of ca 520 m above sea level. The area has a humid, sub-tropical climate and comprises sandstones, siltstones and mudstones. The forest is dominated by tropical sal (Shorea robusta) mixed with Terminalia sp. in the national park, and mixed riverine forest with major tree species of Acacia catechu, Dalbergia sissoo, Bombax ceiba along the river belt at Bhainse (Figure 5).

Habitat of Darwininitium shiwalikianum sp. n.
Darwininitium is of particular interest on two counts: firstly its distribution represents a considerable extension of the known range of the Cerastidae, and secondly it exhibits the most advanced development of the pseudosigmurethrous condition yet found in the family, or indeed in any orthurethran species.
A very short partial renal fold is found in some genera in a few families of Orthurethra, for example Acanthinula (family Acanthinulidae), but a secondary ureteric structure is by far the most clearly and widely developed in the Cerastidae. Pseudosigmurethry was first described in the Australian cerastid Amimopina macleayi (Brazier) by
The position of Darwininitium within the Cerastidae is confirmed by the well-developed pseudosigmurethry, as well as by the highly characteristic darkly pigmented spongy tissue lining the atrium, not known in any other orthurethran family, and the prominent pallial venation (
In addition to its unique pallial anatomy, Darwininitium is immediately separable from other cerastid genera for which there is anatomical information, by the absence of a penial appendix: an elongate, tubular structure, highly differentiated into several distinct regions along its length, inserting well below the point of insertion of the vas deferens, and with its own retractor muscle system. The appendix is found in all other cerastid genera, and occurs commonly throughout the Orthurethra. Indeed, it is difficult to associate Darwininitium phylogenetically with any of the existing cerastid genera.
Biogeographically this new record provides a significant range extension to the family Cerastidae, Nepal lying significantly further north of the previous known distribution.
The detailed anatomy of the Australian species, Amimopina macleayi, was described by
Prem Budha is extremely grateful to the Darwin Initiative Programme “Developing land snail expertise in South and Southeast Asia” run by the Natural History Museum London, for financial support and arranging museum visits to study reference collections. Prem Budha would also like to acknowledge the Malacological Society of London and University Grants Commission, Nepal for a research grant and the Department of National Parks and Wildlife Conservation, Kathmandu and its Chief warden Mr. Megh Bahadur Pandey for providing permission to collect snail specimens from the park. Thierry Backeljau was supported by BELSPO project MO/36/017. Shell images of paratype were produced by Harold Taylor, Natural History Museum, London.