





(C) 2010 Olavi Kurina. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
The females of the four continental Greenomyia Brunetti species in Europe are associated with the males, diagnosed and keyed, providing the first association and description of the females of Greenomyia baikalica Zaitzev, 1994 and Greenomyia stackelbergi Zaitzev, 1982. Colour photographs of their habitus and line drawings of their female terminalia are provided. Greenomyia mongolica Laštovka & Matile, 1974 is found to be a senior synonym of Greenomyia theresae Matile, 2002. syn. n. The diagnostic characters used to distinguish between Greenomyia and Neoclastobasis Ostroverkhova in keys did not hold up to a closer scrutiny and leave the status of Neoclastobasis as separate genus questionable.
Mycetophilidae, Greenomyia, Neoclastobasis, fungus gnats, systematics, identification key, synonymy, Europe
The genus Greenomyia Brunetti was erected to distinguish a single oriental species, Greenomyia nigricoxa Brunetti, 1912. Since then several species (mainly Holarctic) have been described and new combinations proposed.
Species of Greenomyia are not frequently encountered in Europe and apart from the widespread Greenomyia mongolica Laštovka & Matile, 1974 (e.g.
While the males of most Greenomyia species are adequately illustrated and keyed (
Material and collections from a wide range of Palaearctic
sources were studied. The collecting methods, if known, are referred
to in case by each specimen in the studied material section below. The
following codens obtained from
Coll. Hedmark Private collection of Kjell Hedmark, Orsa, Sweden.
Coll. Selin Private collection of Allan Selin, Tallinn, Estonia.
EIHU Hokkaido University Museum, Sapporo, Japan.
IZBE Institute of Agricultural and Environmental Sciences, Estonian University of Life Sciences (former Institute of Zoology and Botany), Tartu, Estonia.
MNHN Museum National d’Histoire Naturelle, Paris, France.
MZLU Museum of Zoology, Lund University, Lund, Sweden.
ZIN Zoological Institute of Russian Academy of Sciences, St. Petersburg, Russia.
ZMHB Museum für Naturkunde Humbolt-Universität zu Berlin, Germany.
ZSM Zoologische Staatssammlung in München, Germany
Three of four species were photographed and figured based on material collected in Vuollerim (Sweden), while illustrations of Greenomyia mongolica
were based on Greek material. The terminalia were detached and cleared
in a solution of KOH, followed by neutralization in acetic acid and
washing in distilled water or in alcohol. The remaining chitinous parts
were inserted into glycerine for detailed study, including
illustration, and thereafter preserved as glycerine preparations in
polyethylene micro vials. Habitus and wing photos were taken of
specimens in alcohol, using a Canon 7D camera fitted with a Canon
MP-E65 (F2.8 1–5 ×) lens. Illustrations of the terminalia were prepared
using a U-DA drawing tube attached to a Olympus CX31 compound
microscope. Terminalia are figured in three different positions:
laterally, dorsally and ventrally. Sternite VIII was detached and
figured separately to better expose the shape of hypoproct and
gonapophysis IX. The preservation method of the studied specimens is
indicated in the material section for each species. We used a 70–80 %
solution of ethanol for alcohol preservation and the chemical method
described by
Compiled from (
| 1 | Wing hyaline (Fig. 12). Coxae yellow, femora and tibiae yellow, only hind femur brown in apical fourth. Mesonotum yellow with three fused longitudinal brown stripes. Cercus one segmented. Sternite VIII with three apical incisions (Fig. 28) | Greenomyia stackelbergi Zaitzev, 1982 |
| – | Wing with apical or preapical dark band. Mesonotum dark brown to black. Coxae yellow or blackish. Cercus one- or two-segmented. Sternite VIII with single central incision apically (Figs 25–27) | 2 |
| 2 | Wing tip darkened on about apical third (Fig. 11) | 3 |
| – | Wing with preapical dark band leaving tip hyaline (Figs 9–10) | 4 |
| 3 | Mid and hind coxae brown to black (Fig. 3). Cercus clearly two-segmented | Greenomyia mongolica Laštovka & Matile, 1974 |
| – | All coxae yellow or slightly darkened basally | Greenomyia lucida (Becker, 1908) [Endemic to the Canary Islands, not seen] |
| 4 | Last palpal segment elongated (Fig. 6). C terminating distinctly before apex of wing, making R5 straight to slightly sinuate (Fig. 10). Cercus two-segmented but segments partly fused (Figs 14, 18) | Greenomyia borealis (Winnertz, 1863) |
| – | Last palpal segment not elongated (Fig. 5). C terminating almost at apex of wing, making R5 distinctly arched (Fig. 9). Cercus one-segmented (Figs 13, 17) | Greenomyia baikalica Zaitzev, 1994 |
Figures 1, 5, 9, 13, 17, 21, 25
SWEDEN. 1♀, Lu. Jokkmokk, Vuollerim, Bomyrberget, in forest 135 m.a.s.l., Malaise trap, 16.–18.VIII.2006 (K. Hedmark leg.) [IZBE, mounted from alcohol]; 1♀, Lu. Jokkmokk, Vuollerim, Bomyrberget, 135 m.a.s.l., Malaise trap, 18.–24.VIII.2007 (K. Hedmark leg.) [MZLU, in alcohol]. FINLAND. 1♂, Kn. Sotkamo, Urpovaara, window trap N2, 26.VIII.–11.IX.1997 (M. Kuussaari leg.) [IZBE, on pin].
Female. Thorax blackish, abdomen brown. Legs yellow, cx3 with small dark markings basally. All trochanters yellow, with small black apicoventral spots. Tibiae with dense brown setae. Scape and pedicel dark yellow, first flagellomere basally yellowish, rest of flagellum brown. Mouthparts pale yellow. Apical palpal segment 1.4–1.6 (n=2) times as long as penultimate segment. Wing with narrow preapical brownish band, gradually tapering towards hind margin. C terminating almost at apex of wing, R5 distinctly arched (Fig. 9). Medial and cubital veins both reach wing margin, CuA1 basally obsolete, brownish shade along posterior margin of CuA2. Terminalia brown, cercus one-segmented, yellow apically. Tergite IX larger than tergite VIII. Gonapophysis IX visible in lateral view, with wide pear-shaped medial incision apically. Sternite VIII deeply incised apicomedially and moderately emarginated basally.
Male. Coloration and other non-terminal characters including palpi similar to female. Apical palpal segment is 1.4 (n=1) times as long as penultimate segment.

Habitus photos of European Greenomyia females. 1 Greenomyia baikalica 2 Greenomyia borealis 3 Greenomyia mongolica 4 Greenomyia stackelbergi.

Head and palpi of European Greenomyia females, lateral view. 5 Greenomyia baikalica 6 Greenomyia borealis 7 Greenomyia mongolica 8 Greenomyia stackelbergi. Last palpal segment is indicated by a red arrow.
Figures 2, 6, 10, 14, 18, 22, 26
SWEDEN. 2♂♂, SK, Lund and Lund, Abusa, undated (J. W. Zetterstedt leg.) [MZLU, on pins]; 1♀, ÖG, Valdemarsvik, Snäckevarp (Snäckehvarps gästgifvaregård 1 km NE Gryt), 11 Aug 1825 (C. Stenhammar leg.) [MZLU, on pin]; 1♀, Lu. Jokkmokk, Vuollerim, in a garden, 105 m.a.s.l., Malaise trap, 8.–15.VIII.2008 (K. Hedmark and M. Karström leg.) [IZBE, mounted from alcohol]; ESTONIA. 1♂, Nigula NR, Haavapeaksi, sweeping, 12.VII.1998 (O. Kurina leg.) [IZBE, on pin]; 1♀, Tartu Marja 14, on window 21.VIII.2008 (O. Kurina leg.) [IZBE, on pin]. GREECE. 1♂ 2♀♀, Central Macedonia, Kerkini lakes area, village Vironia, Ramna site, 41°17'42.5"N, 023°11'33.1"E, 750 m.a.s.l., Malaise trap, 7.–13.VII.2008 (G. Ramel leg.) [IZBE, mounted from alcohol]; 1♂, Central Macedonia, Kerkini lakes area, village Vironia, Beabies site, 41°19'15.4"N, 023°13'39.6"E, 1150 m.a.s.l., Malaise trap, 21.–27.VII.2008 (G. Ramel leg.) [IZBE, mounted from alcohol]; 1♂ Central Macedonia, Kerkini lakes area, village Neo Petritsi, Midway site, 41°18'49.8"N, 023°16'35.6"E, 750 m.a.s.l., Malaise trap, 23.–29.VI.2008 (G. Ramel leg.) [IZBE, mounted from alcohol]. KAZAKHSTAN. 1♀, Alma-Ata, 13.–16.VI. 1824 (Kuzin leg.) [ZIN, on pin].
Female. Thorax brown to blackish. Abdomen entirely brown or first two segments slightly lighter. Legs pale to yellow, except cx2 and cx3 with dark markings basally and apically, all trochanters brown and f3 brown, with lateral parts lighter to yellow. Tibiae with dense brown setae. Scape and pedicel dark yellow, flagellomeres brown. Mouthparts yellow. Apical palpal segment 5.0–5.4 (n=4) times as long as penultimate segment. Wing with broad preapical brownish band, reaching hind margin but gradually paler. C terminating distinctly before apex of wing, R5 straight to slightly sinuate (Fig 10). M2 and CuA1 not reaching wing margin, CuA1 basally obsolete or very weak. Terminalia brown; cercus yellow, two-segmented, apical segment small and partly fused with basal segment. Gonapophysis IX well sclerotized apically, visible in lateral view and with well developed narrow apical incision. Tergite VIII equal in size but slightly wider than tergite IX. Sternite VIII with medial incision apically and well emarginated basally.
Male. Coloration and other non-terminal characters including palpi similar to female. Apical palpal segment is 4.7–5.7 (n=4) times as long as penultimate segment.

Wings of European Greenomyia females. 9 Greenomyia baikalica 10 Greenomyia borealis 11 Greenomyia mongolica 12 Greenomyia stackelbergi.

Female terminalia of European Greenomyia species, lateral view. 13 Greenomyia baikalica 14 Greenomyia borealis 15 Greenomyia mongolica 16 Greenomyia stackelbergi. cerc = cercus; gp = gonapophysis; hyp = hypoproct; st = sternite; tg = tergite.

Female terminalia of European Greenomyia species, dorsal view. 17 Greenomyia baikalica 18 Greenomyia borealis 19 Greenomyia mongolica 20 Greenomyia stackelbergi.
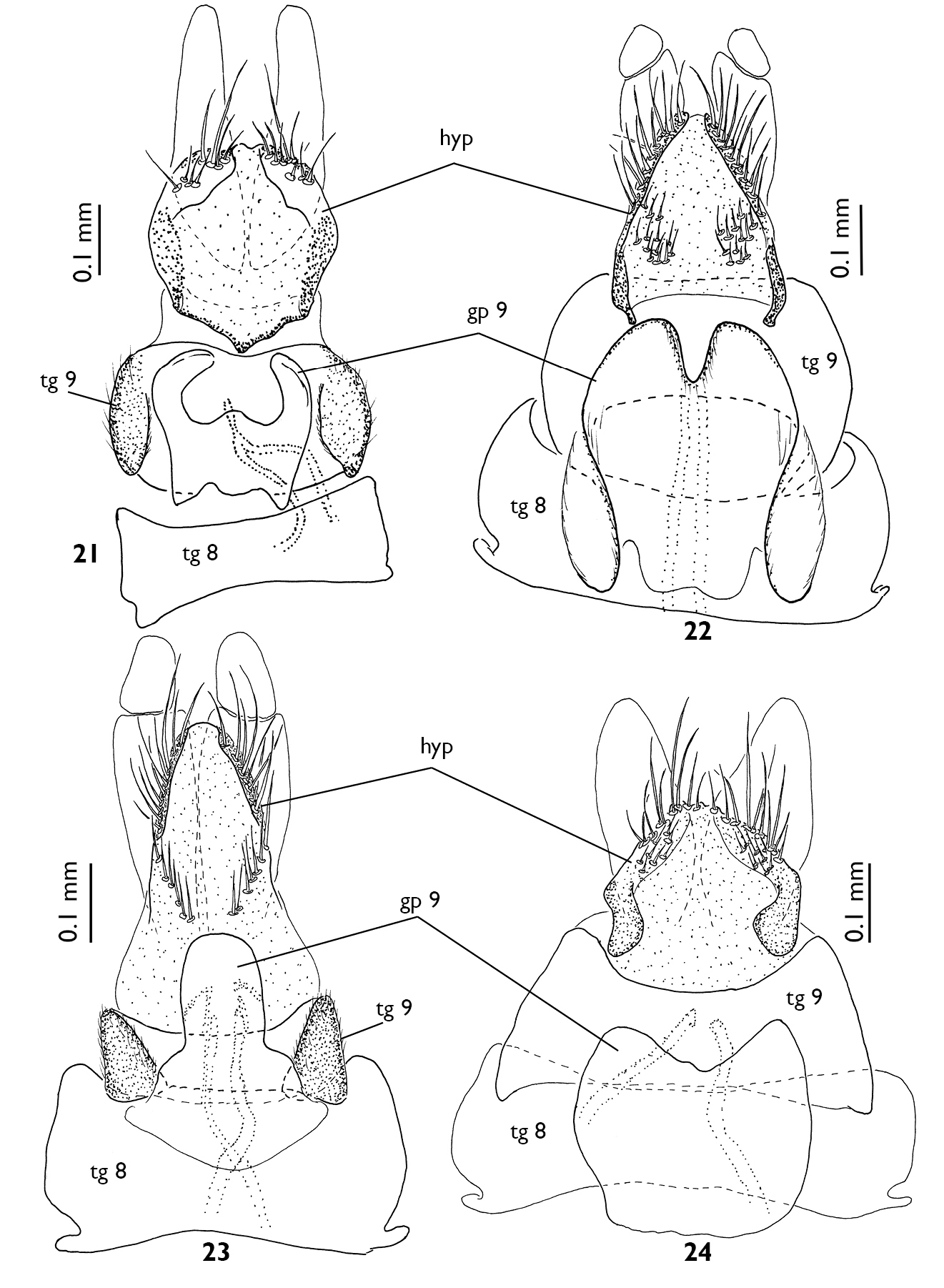
Female terminalia of European Greenomyia species, ventral view, sternite VIII detached. 21 Greenomyia baikalica 22 Greenomyia borealis 23 Greenomyia mongolica 24 Greenomyia stackelbergi.

Female terminalia of European Greenomyia species, ventral view of sternite VIII. 25 Greenomyia baikalica 26 Greenomyia borealis 27 Greenomyia mongolica 28 Greenomyia stackelbergi.
While studying the Swedish specimen from Vuollerim, it ran by the first attempt using the key by
Figures 3, 7, 11, 15, 19, 23, 27
Paratype ♂ of Greenomyia mongolica: MONGOLIA. Central aimak, Tosgoni ovoo, 5–10 km N von Ulan-Baator, 1500–1700 m a.s.l., Exp. Dr. Z. Kaszab 1967 nr. 926, 19–24 Jul 1967 (Z. Kaszab leg.) [MNHN, JKJ-SPM-011843, on pin]
Holotype ♂ of Greenomyia theresae: ITALY. Aosta, Champlong, Dessus, 1000 m a.s.l., “courant sur Feuille de Frêne - 2m -”, 26 Aug 1997 (L. Matile leg.) [MNHN, JKJ-SPM-011844, on pin].
Other material studied: SWEDEN. 1♂, SÖ, Stockholm, Skarpnäck, Skarpa by, 13.VII.-4.X.2003 (B. Viklund leg.) [MZLU, in alcohol]. GERMANY. 1♂, D. Oberpfals, NM Main-Donau-Kanal (Proj. Warncke), 12.IX.–5.X.1988 (S. Blank leg.) [No. 30132 in ZSM, in alcohol]. RUSSIA. 2♂♂ 3♀♀, Nikolsk-Ussur, 29.VII.1926 (Kuznetzov leg.) [ZIN, on pins]. ESTONIA. 1♀, Kääriku, 5.X.1985 (H. Remm leg.) [IZBE, on pin]; 1♀, Luunja, 20.X.1996, on the house wall (O. Kurina leg.) [IZBE, on pin]; 6♂♂ 3♀♀, Karilatsi near Tartu, bait traps, 19.–28.VIII.2005 and 04.–25.IX.2005 (T. Tammaru leg.) [IZBE, on pins]. HUNGARY. 38♂♂ 3♀♀, 10 km S Eger, 47°49'11"N, 020°21'37"E, 20 Aug 1989 (R. Danielsson leg.) [MZLU, on pins]. ITALY. 1♂, Aosta valley, Verrayes, Promellian, 1200 m.a.s.l., sweeping, 17.VI.2007 (V. Soon leg.) [IZBE, on pin]; 1♀, Siena, 6.V.2007 (A. Selin leg.) [Coll. Selin, on pin]; 3♂♂, Trentino-Alto Adige, Prov. Bolzano, Parco Nationale dello Stelvia, Sulden Valley near Schmelz southwest of Prad, 46°36'42.1"N, 010°34'35.6"E, 940 m.a.s.l., 5.IX.–14.X.2005 (J. Ziegler and C. Lange leg.) [1♂ in ZMHB, 2♂♂ in IZBE, mounted from alcohol]. GREECE. 1♂ 2♀♀, Central Macedonia, Kerkini lakes area, village Vironia, Beabies site, 41°19'15.4"N, 023°13'39.6"E, 1150 m.a.s.l., Malaise trap, 30.VI.–6.VII.2008 (G. Ramel leg.) [IZBE, mounted from alcohol]; 6♀♀, Central Macedonia, Kerkini lakes area, village Vironia, Ramna site, 41°17'42.5"N, 023°11'33.1"E, 750 m.a.s.l., Malaise trap, 23.–29.VI.2008 (G. Ramel leg.) [IZBE, mounted from alcohol].
Female. Thorax dark brown to blackish. Abdomen entirely blackish brown or first three segments slightly paler laterally. cx1 entirely yellow or darkened in basal half, cx2 and cx3 entirely dark brown to black. Fore trochanter yellow basally, brown apically. Mid and hind trochanters brown. f1 and f2 yellow, f3 yellow with brown apical fifth. Tibiae yellow, apically slightly darkened, with dense brown setae. Scape, pedicel and flagellomeres brown. Mouthparts pale yellow. Apical palpal segment 1.8–2.2 (n=5) times as long as penultimate segment. Wing tip shaded on about apical third, with darkened area along fore margin. All veins reach wing margin, M2 sometimes basally obsolete or very weak, A1 ending close to, sometimes fused into base of CuA2. Terminalia brown. Cercus distinctly two-segmented, apical segment small, ovate. Gonapophysis IX membranous, widely protruding apically, not visible in lateral view. Tergite VIII larger than tergite IX. Sternite VIII apically with shallow medial incision, moderately emarginated basally.
Male. Coloration and other non-terminal characters including palpi similar to female. Apical palpal segment is 1.7–2.1 (n=5) times as long as penultimate segment.
This species was originally described by
Figures 4, 8, 12, 16, 20, 24, 28
Holotype ♂, RUSSIA. Primorskiy Terr., Santaheza, 07.VII.1927 (A. Stackelberg leg.) [ZIN, on pin].
Other material studied: SWEDEN. 4 ♀♀, Lu. Jokkmokk, Vuollerim, in garden, 105 m.a.s.l., Malaise trap, 11.VIII.–19.IX.2003 (K. Hedmark and M. Karström leg.); 1♂, the same locality, Malaise trap 11.–19.VIII.2004; 25♂♂ 15♀♀, the same locality, Malaise trap 11.VIII.–7.X.2005; 11♂♂ 4♀♀, the same locality, Malaise trap 11.VIII.–22.IX.2006; 18♂♂ 14♀♀, the same locality, yellow pan-trap VII–08.X.2006; 17♂♂ 7♀♀, the same locality, yellow pan-trap 16.VI.–20.VII.2007; 14♂♂ 4♀♀, the same locality, Malaise trap 12.VIII.–28.IX.2007; 9♂♂ 3♀♀, the same locality, window trap VI–11.IX.2007; 4♂♂ 1♀, the same locality, Malaise trap 13.–27.VI.2008; 1♀, the same locality, yellow pan-trap 19.VI.2008; 1♂, the same locality, window trap 1.VI.–1.VII.2008. In total 153 specimens: 100♂♂ 53♀♀, [most in Coll. Hedmark, some in IZBE and MZLU, most of the material preserved in alcohol, while some specimens are mounted from alcohol to pins or slide mounted].
Female.Thorax bi-coloured; mesonotum yellow with variably developed black thoracic stripes; pronotum and propleuron yellow, other pleural parts brown to blackish. Abdominal sternites I-IV entirely yellow or slightly brownish; tergites of first four segments bi-coloured: basally yellow, apically brown (in a few occasions first four tergites entirely brown). Legs all yellow except dark brown band on apical fourth of hind femur. Tibiae densely covered with brown setulae. Scape, pedicel, and 3–5 flagellomeres yellow, rest of flagellum light brown. Mouthparts yellow. Apical palpal segment 4.1–4.4 (n=5) times as long as penultimate segment. Wing hyaline with slight yellowish tinge, all veins reach wing margin, M1 and CuA2 basally obsolete or very weak. Terminalia brown, cercus one-segmented, apically yellow. Gonapophysis IX membranous, subsquare with shallow incision apically, not visible in lateral view. Tergite VIII wider than tergite IX. Sternite VIII medially with deep and narrow incision, lateral incisions more shallow.
Male. Coloration and other non-terminal characters similar to female. The apical palpal segment is 4–5 (n=5) times as long as penultimate segment.
Besides its peculiar distribution (see Fig. 29), Greenomyia stackelbergi
is unique among the four studied species in having vivid yellowish
colouration and hyaline wings. It was described from South Primorje in
the Russian Far East (
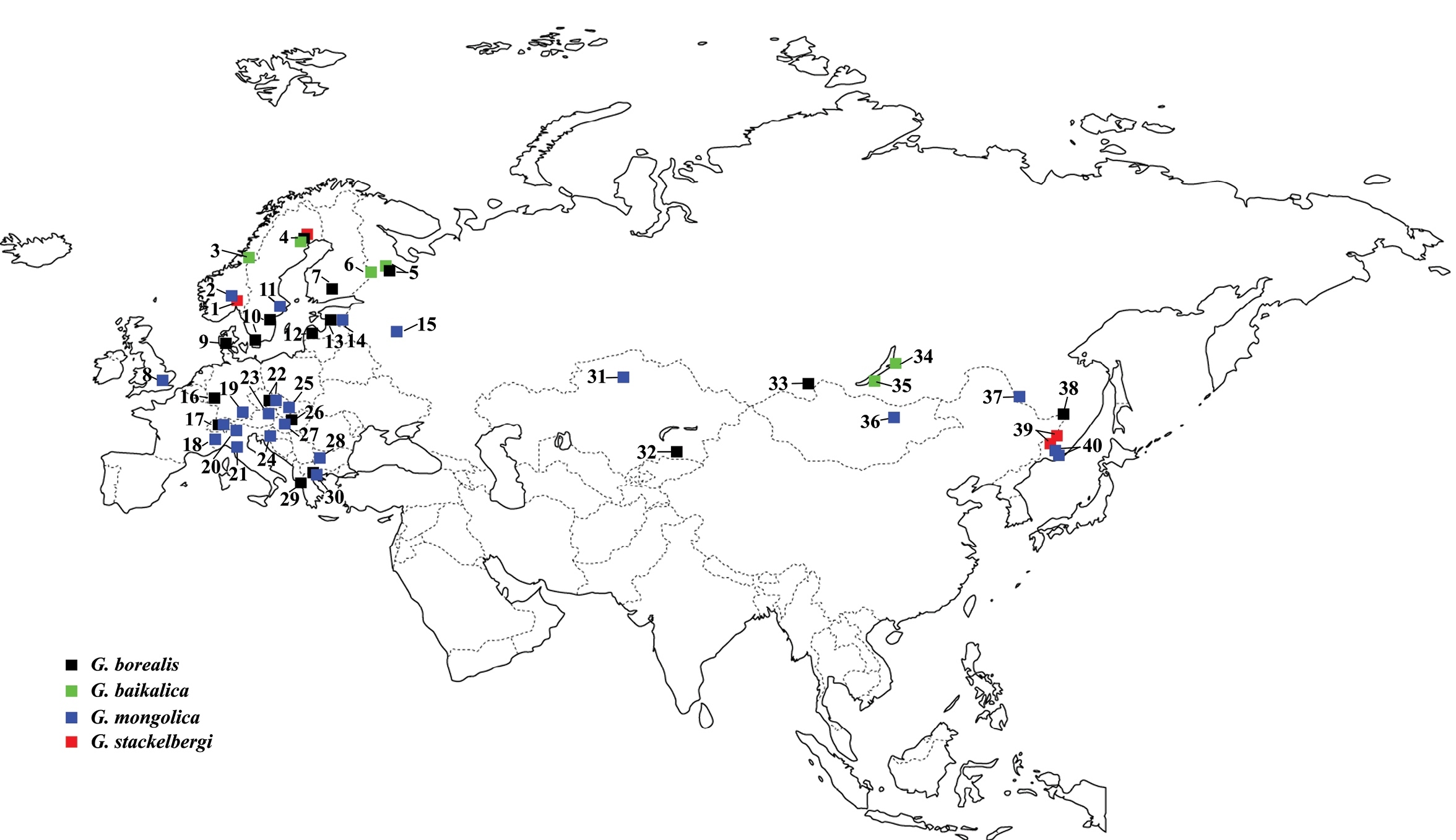
Known records of European Greenomyia species. Greenomyia borealis (black squares): 4 original data 5
Species descriptions of fungus gnats are largely based
and depending on characters in the male terminalia. Females are often
ignored in taxonomic reviews and only a few generic reviews cover all or
the majority of associated females (e.g.
Our study of Greenomyia revealed that the diagnostic characters used to distinguish Greenomyia and Neoclastobasis in keys (e.g.
OK was funded by grant 7558 of the Estonian Science Foundation, JK by the Swedish Taxonomy Initiative. We are grateful to the European Commission’s Research Infrastructure for funding the study visits to MNHN via the SYNTHESYS programme (FR-TAF-956 for JK and FR-TAF-5005 for OK). We are much obliged to the curators M. Baylac (MNHN), C. Daugeron (MNHN), E. Nartshuk (ZIN), W. Schacht (ZSM) and J. Ziegler (ZMHB) for the opportunity to work with the collections and for loans of material. A. Polevoi (Petrozavodsk, Russia), in addition to valuable comments and information, is thanked for assistance in determining the first specimens of Greenomyia stackelbergi and for providing a male of Greenomyia baikalica for study. A. Selin (Tallinn, Estonia), J. Jakovlev (Vantaa, Finland) and G. Ramel (Kerkini, Greece) helped with material for study. Lars Hedström (Uppsala, Sweden) is acknowledged for help with interpreting an old specimen label of Greenomyia borealis. Peter Kerr (Sacramento, USA) and an anonymous reviewer are thanked for their comments and suggestions on the manuscript.