Research Article |
|
Corresponding author: Iulia V. Miu ( iulia.miu@drd.unibuc.ro ) Academic editor: Jesus Maldonado
© 2018 Iulia V. Miu, Chisamera Gabriel B., Viorel D. Popescu, Ruben Iosif, Andreea Nita, Steluta Manolache, Viorel D. Gavril, Ioana Cobzaru, Laurentiu Rozylowicz.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Miu IV, Chisamera GB, Popescu VD, Iosif R, Nita A, Manolache S, Gavril VD, Cobzaru I, Rozylowicz L (2018) Conservation priorities for terrestrial mammals in Dobrogea Region, Romania. ZooKeys 792: 133-158. https://doi.org/10.3897/zookeys.792.25314
|
Abstract
Based on species occurrence records of museum collections, published literature, and unpublished records shared by mammalian experts, we compiled a distribution database for 59 terrestrial mammals populating the extensively protected Dobrogea Region of Romania. The spatial patterns of mammal distribution and diversity was evaluated and systematic conservation planning applied to identify priority areas for their conservation. The spatial analyses revealed that intensive sampling was not directly correlated to mammal diversity but rather to accessibility for inventory. The spatial prioritisation analysis indicated a relatively aggregated pattern of areas with a high or low conservation value with virtually no connecting corridors between them. The significant overlap between Natura 2000 sites and national protected areas induced an over-optimistic vision of the effectiveness and representativeness of existing Natura 2000 network for species found in Annexes II and IV of the Habitats Directive. These results represent a key step in identifying core areas for the protection of mammal diversity and dispersal corridors for improved connectivity, and to guide future conservation efforts in increasing the effectiveness of the existing protected areas in the context of environmental changes.
Keywords
Dobrogea, Natura 2000, species distribution, species richness, systematic conservation planning, terrestrial mammals
Introduction
Terrestrial mammals are well-studied taxa, yet their distribution and conservation status are not fully understood (
A common conservation strategy to prevent the loss of biodiversity is the creation of protected areas (
The Natura 2000 network of protected areas of European importance represents one of the most extensive networks of conservation areas worldwide (Nita et al. 2017). Scientists and policymakers often question the effectiveness of this network due to the Member States allocating fewer funds than needed to implement conservation programs (Nita et al. 2017,
One of Romania’s legal obligations since joining the European Union in 2007 was to designate Natura 2000 sites in a short time (
Due to the diverse landforms, climatic influences, and habitats, Dobrogea harbours a large number of mammal species (
One tool supporting management decisions and for investigating species population coverage within protected areas is spatial conservation prioritisation (
Materials and methods
Mammal species occurrences
To map the distribution of mammals in Dobrogea, we extracted species occurrence records from three primary sources: museum collections, published data, and unpublished field data. Occurrences that could not be georeferenced to a location (e.g., assigned to a large watershed or geographical province), or associated with unspecified taxa within genera, were not included in this geodatabase. The species taxonomy considered in this paper is based on
The dataset used to map the species distribution includes 6724 occurrence records for 59 mammal species. For spatial pattern analyses, we excluded species found exclusively in fenced areas (the European mouflon Ovis aries musimon), the vagrant species (elk Alces alces), and synanthropic species (the rats Rattus rattus, Rattus norvegicus and the house mouse Mus musculus (Table
Spatial bias in species occurrence
Potential bias at the scale of the study area was assessed using the overall spatial autocorrelation in mammal records per 5 × 5 km grid cell. We used Global Moran’s I test (
Estimating species richness, rarity, and dissimilarity
To emphasise regional patterns of richness, rarity, and dissimilarity of mammals of Dobrogea, we aggregated the occurrence records at 5 × 5 km and 10 × 10 km and imported them into BIODIVERSE software (v. 1.1) (
Richness index was measured as the number of species in each grid cell. Species rarity was assessed by dividing the corrected weighted rarity (CWE) by the total number of species in the respective cell, where CWE is (Equation 1).
CWE = WE / Richness (1)
Weighted rarity (WE) of a species represents the occurrence records of sample counts of the respective species divided by the number of occurrence records of all species in the dataset (Equation 2).
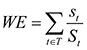 (2)
(2)
where t is a taxon in the set of taxa T across neighbourhood set 1, st is the sum of the sample counts for t across the elements in neighbouring sets 1 and 2, and St represents the total number of samples across the data set for t (
To calculate the differences in species composition across Dobrogea, we used the turnover index (S2), which refers to changes in species composition from one community to another along a gradient and across different sites (
where a is the total number of species found in both neighbour sets, b is the number of species unique to the neighbour set 1, and c is the number of species unique to the neighbour set 2 (
 (3)
(3)
Selecting the smallest values of b or c in the S2 equation denominator reduces the impact of imbalances of species richness on neighbour dissimilarity. The highest value that S2 can result is the value of one (1), which indicates the focal quadrat has no species in common with any neighbour and the lowest possible value is zero (0), indicating that all quadrats have an identical set of species (

Identifying high-priority areas for Natura 2000 mammal species conservation
To identify high-priority areas for mammal species conservation across Natura 2000 sites within Dobrogea Region, we used systematic conservation planning software ZONATION v4 (
For priority analysis, we used 5 × 5 km raster layers for presence/absence data for 14 mammal species listed in Annexes II and IV of the Habitats Directive and a hierarchic mask of the Natura 2000 Sites of Community Importance within Dobrogea Region (Figure
The outputs of the analysis are conservation priority ranking of the landscape, derived from the order of iterative cell ranking whereby each grid cell has a value between 0 and 1, indicating that ranking close to 0 are removed first (low priority), while ranking close to 1 are retained until the end of the iteration. The outputs show the most important areas for mammal species conservation across Natura 2000 sites and a set of curves describing the absolute performance levels of species conservation. We considered as high-priority areas for conservation, all grid cells falling in the top 20% of the predicted priority ranks, a proportion that maximises mammal species representation at the regional level (
The data underpinning the analysis reported in this paper are deposited at GBIF, the Global Biodiversity Information Facility, http://ipt.pensoft.net/resource?r=mammalsdobrogea.
Results
Mammal species occurrences in Dobrogea
We collected 4451 records from published museum collections data (66%), 1326 personal records shared by experts (20%), and 947 records from other papers reporting the results of fauna inventories (14%). Of all the accessible papers (published museum collections and fauna inventories) 67% were published before the year 1990 and 33% after 1990 (Suppl. material
The rate of accumulation of mammal occurrences increased in 1956 by 688 records, due to the rediscovery of the Southern birch mouse (Sicista nordmanni) at Valu lui Traian in 1955. That report attracted additional fieldwork by mammologists the following year, consequently, an increase in the number of records for other rodent species. After 1990, and up to 2017, the peak number of records per year took place in 2007 with 456 new records (Figure

| Order | Family | Species | Total number of records | New records (after 1990) | Total number of UTM 5 × 5 occupied cells | Habitats Directive Annexes | European Red List status |
|---|---|---|---|---|---|---|---|
| Rodentia | Sciuridae | Sciurus vulgaris (Linnaeus, 1758) | 6 | 3 | 4 | – | Least concern |
| Spermophilus citellus (Linnaeus, 1766) | 214 | 92 | 95 | II/IV | Vulnerable | ||
| Gliridae | Dryomys nitedula (Pallas, 1778) | 35 | 22 | 20 | IV | Least concern | |
| Muscardinus avellanarius (Linnaeus, 1758) | 1 | 1 | 1 | – | Least concern | ||
| Glis glis (Linnaeus, 1766) | 3 | 3 | 3 | – | Least concern | ||
| Castoridae | Castor fiber (Linnaeus, 1758) | 12 | 12 | 5 | II/IV | Least concern | |
| Dipodidae | Sicista nordmanni (Keyserling & Blasius, 1840) | 76 | 0 | 2 | II/IV | Vulnerable | |
| Spalacidae | Nannospalax leucodon (Nordmann, 1840) | 163 | 82 | 57 | – | Least concern | |
| Cricetidae | Cricetus cricetus (Linnaeus, 1758) | 2 | 0 | 1 | IV | Least concern | |
| Mesocricetus newtoni (Nehring, 1898) | 98 | 13 | 31 | II/IV | Near threatened | ||
| Ondatra zibethicus (Linnaeus, 1766) | 87 | 37 | 57 | – | Invasive | ||
| Arvicola amphibius (Linnaeus, 1758) | 29 | 12 | 22 | – | Least concern | ||
| Microtus agrestis (Linnaeus, 1761) | 28 | 11 | 18 | – | Least concern | ||
| Microtus arvalis (Pallas, 1779) | 187 | 40 | 44 | – | Least concern | ||
| Microtus levis (Miller, 1908) | 29 | 9 | 13 | – | Least concern | ||
| Microtus subterraneus (Selys-Longchamps, 1836) | 16 | 7 | 9 | – | Least concern | ||
| Myodes glareolus (Schreber, 1780) | 1 | 0 | 1 | – | Least concern | ||
| Muridae | Micromys minutus (Pallas, 1771) | 36 | 15 | 24 | – | Least concern | |
| Apodemus agrarius (Pallas, 1771) | 451 | 96 | 50 | – | Least concern | ||
| Apodemus flavicollis (Melchior, 1834) | 134 | 80 | 34 | – | Least concern | ||
| Apodemus sylvaticus (Linnaeus, 1758) | 1327 | 330 | 65 | – | Least concern | ||
| Apodemus uralensis (Pallas, 1811) | 16 | 6 | 8 | – | Least concern | ||
| Mus spicilegus (Petényi, 1882) | 20 | 20 | 19 | – | Least concern | ||
| Myocastoridae | Myocastor coypus (Molina, 1782) | 5 | 2 | 5 | – | Invasive | |
| Lagomorpha | Leporidae | Lepus europaeus (Pallas, 1778) | 262 | 255 | 102 | – | Least concern |
| Erinaceomorpha | Erinaceidae | Erinaceus roumanicus (Barrett-Hamilton, 1900) | 52 | 40 | 39 | – | Least concern |
| Soricomorpha | Soricidae | Crocidura leucodon (Hermann, 1780) | 85 | 14 | 18 | – | Least concern |
| Crocidura suaveolens (Pallas, 1811) | 131 | 40 | 36 | – | Least concern | ||
| Neomys anomalus (Cabrera, 1907) | 12 | 4 | 8 | – | Least concern | ||
| Neomys fodiens (Pennant, 1771) | 5 | 1 | 4 | – | Least concern | ||
| Sorex araneus (Linnaeus, 1758) | 63 | 14 | 25 | – | Least concern | ||
| Sorex minutus (Linnaeus, 1766) | 15 | 9 | 10 | – | Least concern | ||
| Talpidae | Talpa europaea (Linnaeus, 1758) | 65 | 53 | 55 | – | Least concern | |
| Carnivora | Felidae | Felis silvestris (Schreber, 1777) | 101 | 94 | 52 | IV | Least concern |
| Felidae | Lynx lynx (Linnaeus, 1758) | 2 | 1 | 2 | II/IV | Least concern | |
| Canidae | Canis aureus (Linnaeus, 1758) | 214 | 198 | 94 | – | Least concern | |
| Canis lupus (Linnaeus, 1758) | 27 | 22 | 14 | II/IV | Least concern | ||
| Nyctereutes procyonoides (Gray, 1834) | 87 | 35 | 41 | – | Invasive | ||
| Vulpes vulpes (Linnaeus, 1758) | 230 | 223 | 122 | – | Least concern | ||
| Mustelidae | Mustela erminea (Linnaeus, 1758) | 25 | 7 | 23 | IV | Vulnerable | |
| Mustela eversmanii (Lesson, 1827) | 31 | 24 | 25 | II/IV | Vulnerable | ||
| Mustela lutreola (Linnaeus, 1761) | 119 | 109 | 50 | II/IV | Endangered | ||
| Mustela nivalis (Linnaeus, 1766) | 67 | 54 | 50 | – | Least concern | ||
| Mustela putorius (Linnaeus, 1758) | 89 | 74 | 61 | – | Least concern | ||
| Vormela peregusna (Güldenstädt, 1770) | 70 | 16 | 39 | II/IV | Vulnerable | ||
| Martes foina (Erxleben, 1777) | 98 | 97 | 58 | – | Least concern | ||
| Martes martes (Linnaeus, 1758) | 36 | 36 | 20 | – | Least concern | ||
| Meles meles (Linnaeus, 1758) | 102 | 92 | 60 | – | Least concern | ||
| Neovison vison (Schreber, 1777) | 2 | 2 | 1 | – | Invasive | ||
| Lutra lutra (Linnaeus, 1758) | 55 | 49 | 35 | II/IV | Near threatened | ||
| Artiodactyla | Suidae | Sus scrofa (Linnaeus, 1758) | 221 | 204 | 105 | – | Least concern |
| Cervidae | Dama dama (Linnaeus, 1758) | 46 | 29 | 21 | – | Least concern | |
| Cervus elaphus (Linnaeus, 1758) | 38 | 32 | 25 | – | Least concern | ||
| Capreolus capreolus (Linnaeus, 1758) | 262 | 190 | 119 | – | Least concern |
Spatial patterns in mammal species occurrences in Dobrogea
Of 757 UTM 5 × 5 km grid cells encompassing the Dobrogea Region, only 335 grid cells (i.e., 44%) include reported mammal sightings (Figure

Mammals reported occurrences in Dobrogea Region, Romania at 5 × 5 km resolution. Grids with reported occurrences before 1990 were plotted as old records whereas those with reported occurrences after 1990 were considered new records (reports of synanthropic and vagrant mammals were excluded).
The mammal occurrences at 5 × 5 km resolution ranged between 1 and 35 reported species per quadrat (Figure


Checklist of synanthropic and vagrant mammals of Dobrogea Region, Romania.
| Order | Family | Species | Total number of records | New records (after 1990) | Total number of UTM 5 × 5 occupied cells |
|---|---|---|---|---|---|
| Rodentia | Muridae | Rattus norvegicus (Berkenhout, 1769) | 114 | 49 | 64 |
| Rodentia | Muridae | Rattus rattus (Linnaeus, 1758) | 3 | 2 | 3 |
| Rodentia | Muridae | Mus musculus (Linnaeus, 1758) | 1001 | 139 | 78 |
| Artiodactyla | Cervidae | Alces alces (Linnaeus, 1758) | 4 | 0 | 3 |
| Artiodactyla | Bovidae | Ovis aries musimon (Pallas, 1881) | 9 | 5 | 4 |
Species richness, rarity, and dissimilarity
When aggregating species records at 10 × 10 km, the number of reported species ranged from 2 to 45 per cell grid, with the highest species diversity located in the northern part of Dobrogea Region overlapping the following Natura 2000 sites: North Dobrogea Plateau with a maximum richness of 45 species, western part of Danube Delta with 39 species and Agighiolului Hills with 38 species. Most of the grid cells with species richness are concentrated in the northern region reflecting an optimal sampling of mammal species (number of species from 29 to 37) (Figure

Corrected weighted rarity (CWE) varied across Dobrogea from 0.0087 for cell grids with widespread species to 0.62 grid cells with species of restricted distribution. The highest value of corrected weighted rarity can be found in the Danube Delta, specifically in the levee complex of Puiu – Roșu – Lumina, with a value up to 0.62 (Figure
The values of dissimilarity index S2 ranged from 0 to 1 with the highest turnover quadrats in the southern area of Dobrogea where there are low richness zones. The value of 1 implies that the quadrat has no species in common with any neighbour (Figure


High-priority areas for conservation within Natura 2000 sites
Based on the Zonation analysis results, the top spatial conservation priorities overlap Danube Delta, North Dobrogea Plateau, and the Măcin Mountains in the northern part of Dobrogea region, where a relatively aggregated pattern of top conservation value areas appear due to their extensive wetland area and forested habitats. Isolated hotspots are represented by Dumbrăveni-Urluia Valley-Vederoasa Lake in the southwest, Hagieni – Cotul Văii Forest in the southeast, and Cheia Jurassic Reefs in Central Dobrogea. Grid cells with the lowest ranking are located in the central and southern part of Dobrogea Region, where the majority of the regions’ agricultural lands are clustered (Figure


Zonation performance curves quantifying the proportion of remaining species occurrences covered by Natura 2000 sites in Dobrogea. When considering 20% of landscape within Natura 2000 sites as protected (e.g., conservation measures are enforced), 45% of Natura 2000 mammal occurrences are protected.
Discussion
By using an updated distribution of terrestrial mammals, we identified high priority areas for protecting mammal diversity to guide future conservation efforts in an extensively protected Romanian region. In the broader context of systematic conservation planning, the prioritisation analysis is a useful tool to identify key areas for biodiversity conservation, e.g., where species are more likely to survive (
The number of reported occurrences in Dobrogea varied among species. The largest number of records (20%) are for the wood mouse (Apodemus sylvaticus), mostly because they are widespread within the region, have a higher population abundance, and are evidently. The wood mouse may be easily misidentified as a yellow-necked mouse (Apodemus flavicollis) (
The lowest number of records in Dobrogea is recorded for the hazel dormouse (Muscardinus avellanarius) and the bank vole (Myodes glareolus), with only one record per each species. Two other widespread species, but with an uncharacteristically low number of reported presences are the red fox (Vulpes vulpes) and the roe deer (Capreolus capreolus), possibly because they are common species, with minor interest for biologists. The research effort for species sampling was focused on selected areas: Măcin Mountains National Park, Danube Delta Biosphere Reserve, and the North Dobrogea Plateau. Here, we recorded a higher than expected number of species occurrences per grid cell, mostly because the long-term protection status attracted faunistic inventory projects over time. Typically, the sampling bias is higher in protected areas because they attract more conservation funds leading to greater efforts for biodiversity research (e.g.,
We found that high species richness did not match all the hotspots of sampling efforts, such as in Valu lui Traian Biological Research Station and Fântânița-Murfatlar (location 1 in Figure
The spatial turnover index (S2) suggests that areas with lower species richness are dissimilar compared to their neighbours. Notably, we observed some affinities of particular species towards low species richness areas (
Our results regarding the identification of high-priority areas in Dobrogea for mammal species listed in Annexes II and IV of the Habitats Directive highlight a relatively aggregated pattern of the grid cells with high conservation value in the northeastern and northern Dobrogea Region, where there are extensive wetlands and forests (i.e., Danube Delta, Măcin Mountains National Park, and North Dobrogea Plateau) (
The significant overlap between Natura 2000 sites and the other protected areas statutes leads to misunderstandings in law enforcement and an over-optimistic vision of their effectiveness (
The absence of buffer zones and corridors between Natura 2000 sites and small isolated protected areas (the area of the smallest Natura 2000 site in Dobrogea is 0.11 km2), are not beneficial in maintaining viable populations, causing the isolation of species with low mobility and specific habitat requirements (
Our study is limited by the lack of viable and current distribution data. Most records do not identify geographical coordinates, but localities or toponymies. This makes the niche modelling at a fine scale a challenge. Furthermore, elusive species such as Mesocricetus newtoni, Sicista nordmanni, and Vormela peregusna, are data deficient, and the lack of records (false absences) may influence the results of the analysis. Similarly, misidentification of sibling species may lead to over- or under- estimation of their range. Notably, a study analysing the distribution of amphibians in Dobrogea (
Future research may focus on identifying buffer zones around Natura 2000 sites to minimise potential negative impacts, particularly in Natura 2000 sites that are adjacent to agricultural areas. From this assessment, we envisage further mapping of corridor networks between small isolated protected areas in southern and central Dobrogea. New research should focus on systematic surveys of agricultural landscapes in central and southern Dobrogea, where vegetation patches remain as refugees for some species listed in Annexes II and IV of the Habitats Directive (Vormela peregusna, Mustela eversmanii, Spermophilus citellus, Mesocricetus newtoni, Cricetus cricetus, and Sicista nordmanni).
Acknowledgments
IVM, VDP, AN, SM, and LR were supported by a grant from the Romanian National Authority for Scientific Research, PN-III-P4-ID-PCE-2016-0483. Editing and proofreading by Edward F. Rozylowicz streamlined the flow of the manuscript. We thank Maria Patroescu, Tiberiu Sahlean, and an anonymous reviewer for their comments that improved the manuscript. We are grateful to Dumitru Murariu, Nastase Radulet, Bogdan Bejenariu, Catalin Stanciu, Dragos Mantoiu, Ionut Stefan Iorgu, Daniyar Memedemin, Razvan Popescu Mirceni, Silviu Petrovan, Marius Skolka, Razvan Zaharia, Alexandra Telea, Dragos Balasoiu, Mihaela Stanescu, and Elena Buhaciuc-Ionita for sharing their data on species occurrences.
References
- Araújo MB, Alagador D, Cabeza M, Nogués-Bravo D, Thuiller W (2011) Climate change threatens European conservation areas. Ecology Letters 14: 484–492. https://doi.org/10.1111/j.1461-0248.2011.01610.x
- Arponen A, Heikkinen RK, Thomas CD, Moilanen A (2005) The value of biodiversity in reserve selection: Representation, species weighting, and benefit functions. Conservation Biology 19: 2009–2014. https://doi.org/10.1111/j.1523-1739.2005.00218.x
- Arslan A, Kryštufek B, Matur F, Zima J (2016) Review of chromosome races in blind mole rats (Spalax and Nannospalax). Folia Zoologica 65: 249–301. https://doi.org/10.25225/fozo.v65.i4.a1.2016
- Ausländer D, Hellwing S (1957) Beiträge zur variabilität und biologie der streifenmaus (Sicista subtilis nordmanni Keys. et Blas, 1840). Travaux du Muséum d’Histoire naturelle Grigore Antipa 1: 255–274.
- Bartolommei P, Sozio G, Bencini C, Cinque C, Gasperini S, Manzo E, Petre S, Solano E, Cozzolino R, Mortelliti A (2016) Field identification of Apodemus flavicollis and Apodemus sylvaticus: a quantitative comparison of different biometric measurements. Mammalia 80: 541–547. https://doi.org/10.1515/mammalia-2014-0051
- Botts EA, Erasmus BFN, Alexander GJ (2011) Geographic sampling bias in the South African Frog Atlas Project: Implications for conservation planning. Biodiversity and Conservation 20: 119–139. https://doi.org/10.1007/s10531-010-9950-6
- Bunescu A (1959) Contribution à l’étude de la répartition geographique de quelques mammifères méditerranéene en Roumanie. Säugerierkundliche Mitteilungen 1: 1–4.
- Bunescu A (1961) Contribuţii la studiul răspândirii geografice a unor animale mediteraneene din R. P. Română, Nota II – Vertebrate. Probleme de Geografie 8: 123–144.
- Christie MR, Knowles LL (2015) Habitat corridors facilitate genetic resilience irrespective of species dispersal abilities or population sizes. Evolutionary Applications 8: 454–463. https://doi.org/10.1111/eva.12255
- Cogalniceanu D, Rozylowicz L, Székely P, Samoilǎ C, Stanescu F, Tudor M, Székely D, Iosif R (2013) Diversity and distribution of reptiles in Romania. ZooKeys 341: 49–76. https://doi.org/10.3897/zookeys.341.5502
- Crooks KR, Burdett CL, Theobald DM, Rondinini C, Boitani L (2011) Global patterns of fragmentation and connectivity of mammalian carnivore habitat. Philosophical transactions of the Royal Society of London: Series B, Biological sciences 366: 2642–2651. https://doi.org/10.1098/rstb.2011.0120
- Cuzic M, Marinov M (2004) Date privind situația populației de nurcă europeană (Mustela lutreola L, 1761) (Mammalia, Carnivora, Mustelidae) în câteva zone din Rezervația Biosferei Delta Dunării. Muzeul Brukenthal. Studii și Comunicări Științele Naturii 29: 231–239.
- Eklund J, Arponen A, Visconti P, Cabeza M (2011) Governance factors in the identification of global conservation priorities for mammals. Philosophical transactions of the Royal Society of London: Series B, Biological sciences 366: 2661–2669. https://doi.org/10.1098/rstb.2011.0114
- Ferrier S, Wintle BA (2009) Quantitative approaches to spatial conservation prioritization: matching the solution to the need. In: Moilanen KA, Wilson HPP (Eds) Spatial Conservation Prioritization: Quantitative Methods & Computational Tools.Oxford University Press, Oxford, 1–13.
- Fortin M, Dale M (2005) Spatial Analysis – A Guide for Ecologists. Cambridge University Press, Cambridge, 365 pp.
- Gaston KJ, Davies RG, Orme CDL, Olson VA, Thomas GH, Ding TS, Rasmussen PC, Lennon JJ, Bennett PM, Owens IP, Blackburn TM (2007) Spatial turnover in the global avifauna. Proceedings of the Royal Society B: Biological Sciences 274: 1567–1574. https://doi.org/10.1098/rspb.2007.0236
- Grand J, Cummings MP, Rebelo TG, Ricketts TH, Neel MC (2007) Biased data reduce efficiency and effectiveness of conservation reserve networks. Ecology Letters 10: 364–374. https://doi.org/10.1111/j.1461-0248.2007.01025.x
- Hamar M, Schutowa M (1966) Neue daten über die geographische veränderlichkeit und die entwicklung der gattung Mesocricetus Nehring, 1898 (Glires, Mammalia). Zeitschrift-Saeugetierkunde 31: 237–251.
- Ioja CI, Patroescu M, Rozylowicz L, Popescu VD, Vergheleţ M, Zotta M, Felciuc M (2010) The efficacy of Romania’s protected areas network in conserving biodiversity. Biological Conservation 143: 2468–2476. https://doi.org/10.1016/j.biocon.2010.06.013
- Jaarola M, Martínková N, Gündüz İ, Brunhoff C, Zima J, Nadachowski A, Amori G, Bulatova NS, Chondropoulos B, Fraguedakis-Tsolis S, González-Esteban J, José López-Fuster M, Kandaurov AS, Kefelioğlu H, da Luz Mathias M, Villate I, Searle JB (2004) Molecular phylogeny of the speciose vole genus Microtus (Arvicolinae, Rodentia) inferred from mitochondrial DNA sequences. Molecular Phylogenetics and Evolution 33: 647–663. https://doi.org/10.1016/j.ympev.2004.07.015
- Kiss JB, Doroșencu A, Marinov ME, Alexe V, Bozagievici R (2012) Considerations regarding the occurrence of the Eurasian Beaver (Castor fiber Linnaeus 1758) in the Danube Delta (Romania). Scientific Annals of the Danube Delta Institute 18: 49–56. https://doi.org/10.7427/DDI.18.05
- Kiss JB, Marinov M, Alexe V, Doroșenco A (2014) Eurasian Beaver (Castor fiber L. 1758), Pine Marten (Martes martes L. 1758) and Stone Marten (Martes foina Erxleben, 1777) in the Danube Delta (Romania). Beitrage zur Jagd-und Wildforschung 39: 347–355.
- Kiss JB, Marinov M, Alexe V, Doroșenco A, Sandor A (2013) Pătrunderea jderului de copac (Martes martes, Linnaeus 1758) în Delta Dunării (România) și considerații privind urmările ecologice scontate al acestui fenomen. Revista Pădurilor 128(3): 38–47.
- Kiss JB, Marinov M, Alexe V, Sandor A (2012) First record of the Pine Marten (Martes martes) in the Danube Delta. North-Western Journal of Zoology 8: 195–197.
- Kukkala AS, Moilanen A (2013) Core concepts of spatial prioritisation in systematic conservation planning. Biological Reviews 88: 443–464. https://doi.org/10.1111/brv.12008
- Kukkala AS, Santangeli A, Butchart SHM, Maiorano L, Ramirez I, Burfield IJ, Moilanen A (2016) Coverage of vertebrate species distributions by Important Bird and Biodiversity Areas and Special Protection Areas in the European Union. Biological Conservation 202: 1–9. https://doi.org/10.1016/j.biocon.2016.08.010
- Laffan SW, Lubarsky E, Rosauer DF (2010) Biodiverse, a tool for the spatial analysis of biological and related diversity. Ecography 33: 643–647. https://doi.org/10.1111/j.1600-0587.2010.06237.x
- Lehtomäki J, Moilanen A (2013) Methods and workflow for spatial conservation prioritization using Zonation. Environmental Modelling and Software 47: 128–137. https://doi.org/10.1016/j.envsoft.2013.05.001
- Lennon JJ, Koleff P, Greenwood JJD, Gaston KJ (2001) The geographical structure of British bird distributions: Diversity, spatial turnover and scale. Journal of Animal Ecology 70: 966–979. https://doi.org/10.1046/j.0021-8790.2001.00563.x
- Lung T, Meller L, van Teeffelen AJA, Thuiller W, Cabeza M (2014) Biodiversity Funds and Conservation Needs in the EU Under Climate Change. Conservation Letters 7: 390–400. https://doi.org/10.1111/conl.12096
- Maiorano L, Amori G, Montemaggiori A, Rondinini C, Santini L, Saura S, Boitani L (2015) On how much biodiversity is covered in Europe by national protected areas and by the Natura 2000 network: Insights from terrestrial vertebrates. Conservation Biology 29: 986–995. https://doi.org/10.1111/cobi.12535
- Manolache S, Nita A, Ciocanea CM, Popescu VD, Rozylowicz L (2018) Power, influence and structure in Natura 2000 governance networks. A comparative analysis of two protected areas in Romania. Journal of Environmental Management 212: 54–64. https://doi.org/10.1016/j.jenvman.2018.01.076
- Margules CR, Pressey RL (2000) Systematic conservation planning. Nature 405: 243–253. https://doi.org/10.1038/35012251
- Memedemin D, Tudor M, Cogalniceanu D, Skolka M, Banica G, Rozylowicz L (2017) Update on the Geographic Distribution of Lutra lutra at the Romanian Black Sea Coast. Travaux du Muséum National d’Histoire Naturelle Grigore Antipa 60: 413–417. https://doi.org/10.1515/travmu-2017-0004
- Moilanen A, Veach V, Meller L, Arponen A, Kujala H (2014) Spatial conservation planning methods and software. Version 4. User Manual. University of Helsinki, Helsinki, Finland, 290 pp. https://doi.org/10.1017/CBO9781107415324.004
- Murariu D (1996) Mammals of the Danube Delta (Romania). Travaux du Museum National d’Histoire Naturelle 36: 361–371
- Murariu D (2006) Mammal ecology and distribution from North Dobrogea (Romania). Travaux du Muséum national d’Histoire naturelle 49: 387–399.
- Murariu D, Atanasova I, Raykov I, Chişamera G (2009) Results on mammal (Mammalia) survey from Bulgarian and Romanian Dobrogea. Travaux du Museum national d’Histoire naturelle 52: 371–386.
- Murariu D, Chişamera G, Petrescu A, Atanasova I, Raykov I (2010) Terrestrial vertebrates of Dobrogea – Romania and Bulgaria. Travaux du Muséum national d’Histoire naturelle Grigore Antipa 53: 357–375. https://doi.org/10.2478/v10191-010-0027-2
- Murariu D, Munteanu D (2005) Fauna României – Mammalia – Carnivora. Editura Academiei Române, Bucharest, 224 pp
- Nita A, Rozylowicz L, Manolache S, Ciocanea CM, Miu IV, Popescu VD (2016) Collaboration Networks in Applied Conservation Projects across Europe. PLoS ONE 11(10): e0164503. https://doi.org/10.1371/journal.pone.0164503
- Nita A, Ciocanea CM, Manolache S, Rozylowicz L (2018) A network approach for understanding opportunities and barriers to effective public participation in the management of protected areas. Social Network Analysis and Mining 8. https://doi.org/10.1007/s13278-018-0509-y
- Ord JK, Getis A (1995) Local spatial autocorrelation statistics: distributional issues and an application. Geographical Analysis 27: 286–306. https://doi.org/10.1111/j.1538-4632.1995.tb00912.x
- Popescu A, Murariu D (2001) Fauna României. Mammalia. Rodentia. Editura Academiei Române, Bucharest, 214 pp.
- Popescu VD, Rozylowicz L, Cogalniceanu D, Niculae IM, Cucu AL (2013) Moving into protected areas? Setting conservation priorities for Romanian reptiles and amphibians at risk from climate change. PLoS ONE 8: e79330. https://doi.org/10.1371/journal.pone.0079330
- Possingham HP, Wilson KA, Andelman SJ, Vynme CH (2006) Protected areas: Goals , limitations , and design. In: Groom MJ, Meffe GK, Carroll CR (Eds) Principles of Conservation Biology.Sinauer Associates, Sunderland, 509–533.
- Pouzols F, Toivonen T, Di Minin E, Kukkala AS, Kullberg P, Kuusterä J, Lehtomäki J, Tenkanen H, Verburg PH, Moilanen A (2014) Global protected area expansion is compromised by projected land-use and parochialism. Nature 516: 383–386. https://doi.org/10.1038/nature14032
- Pressey RL, Cabeza M, Watts ME, Cowling RM, Wilson K (2007) Conservation planning in a changing world. Trends in Ecology & Evolution 22: 583–92. https://doi.org/10.1016/j.tree.2007.10.001
- Rey V, Ianos I, Groza O, Patroescu M (2007) Atlas de la Roumanie Nouvelle edition. Reclus, Montpellier, 208 pp.
- Santos BPA, Bitencourt C, Rapini A (2017) Distribution patterns of Kielmeyera (Calophyllaceae): the Rio Doce basin emerges as a confluent area between the northern and southern Atlantic Forest. Neotropical Biodiversity 3: 1–9. https://doi.org/10.1080/23766808.2016.1266730
- Rodrigues ASL, Andelman SJ, Bakarr MI, Boitani L, Brooks TM, Cowling RM, Fishpool LDC, Da Fonseca GAB, Gaston KJ, Hoffmann M, Long JS, Marquet PA, Pilgrim JD, Pressey RL, Schipper J, Sechrest W, Stuart SN, Underhill LG, Waller RW, Watts MEJ, Yan X (2004) Effectiveness of the global protected area network in representing species diversity. Nature 428: 640–3. https://doi.org/10.1038/nature02422
- Rodrigues ASL, Grenyer R, Baillie JEM, Bininda-Emonds ORP, Gittlemann JL, Hoffmann M, Safi K, Schipper J, Stuart SN, Brooks T (2011) Complete, accurate, mammalian phylogenies aid conservation planning, but not much. Philosophical Transactions of the Royal Society B: Biological Sciences 366: 2652–2660. https://doi.org/10.1098/rstb.2011.0104
- Rozylowicz L, Nita A, Manolache S, Ciocanea CM, Popescu VD (2017) Recipe for success: A network perspective of partnership in nature conservation. Journal for Nature Conservation 38: 21–29. https://doi.org/10.1016/j.jnc.2017.05.005
- Sánchez-Fernández D, Abellán P, Aragón P, Varela S, Cabeza M (2017) Matches and mismatches between conservation investments and biodiversity values in the European Union. Conservation Biology 32: 109–115. https://doi.org/10.1111/cobi.12977
- Székely P, Iosif R, Székely D, Stanescu F, Cogalniceanu D (2013) Range extension for the Eastern spadefoot toad Pelobates syriacus (Boettger, 1889) (Anura: Pelobatidae). Herpetology Notes 6: 481–484.
- Székely P, Plaiaşu R, Tudor M, Cogalniceanu D (2009) The distribution and conservation status of amphibians in Dobrudja (Romania). Turkish Journal of Zoology 33: 147–156. https://doi.org/10.3906/zoo-0805-12
- Temple HJ, Terry A (2009) European mammals: Red List status, trends, and conservation priorities. Folia zoologica 58: 248
- Whittaker RH (1972) Evolution and measurement of species diversity. Taxon 21: 213–251. https://doi.org/10.2307/1218190
- Williams PH, Margules CR, Hilbert DW (2002) Data requirements and data sources for biodiversity priority area selection. Journal of Biosciences 27: 327–338. https://doi.org/10.1007/BF02704963
- Wilson DE, Reeder DM (2005) Mammal Species of the World. A Taxonomic and Geographic Reference (3rd edn). Johns Hopkins University Press, 2142 pp.
- Wilson KA, Cabeza M, Klein CJ (2009) Fundamental concepts of spatial conservation prioritization. In: Moilanen A, Wilson KA, Possingham HP (Eds) Spatial Conservation Prioritization: Quantitative Methods and Computational Tools.Oxford University Press, New York, 16–27.
Supplementary materials
Publications used to compile distribution of terrestrial mammal species from Dobrogea Region, Romania (field reports and data from museum collections)
Occurrences maps for 59 mammal species
Methodology used to identify high-priority Natura 2000 sites with Zonation v4