





(C) 2012 Marek Wanat. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Perapion connexum (Schilsky) is recorded for the first time from Hungary and Kyrgyzstan, and new distribution data from Ukraine and Russia are provided. Preliminary placements of this weevil in faunal checklists for Poland and Slovakia are here documented with detailed data. Its occurrence in Austria based on older evidence, is discussed. The neophytic and invasive in Central Europe sorrel Rumex confertus Willd. is confirmed to be its unique host plant in Poland. Morphology of the newcoming weevil is described and illustrated, and the key to all Central European species of Perapion is presented.
Weevils, Curculionoidea, Perapion, Poland, Hungary, Slovakia, Kyrgyzstan, new records, morphology, taxonomy, key, biology, Rumex confertus
The title weevil species was originally described from Aulie-Ata (currently Taraz) in SE Kazakhstan and long considered as confined to Asian fauna (
The study was based on 118 specimens collected by the authors in 2000-2008, obtained from other collectors or borrowed from several institutional collections.
Measurements were taken using a calibrated stereomicroscopic grid eyepiece. Body length excludes rostrum, but includes head; it was measured in lateral view from the anterior eye margin to the apex of the elytra. Width of head was measured across middle of eyes. Tarsal width was measured at the level of the 3rd segment.
Photos of specimens were taken with a Leica M205C stereomicroscope and attached JVC KYF75 digital camera, and processed using the AutoMontage Pro and Adobe Photoshop CS2 software programmes.
The abbreviations used are as follows: MW, AP, KS – authors’ respective acronyms, HG – G. Hegyessy, KMS – Kazinczy Museum, Sátoraljaújhely, nr. – near. Unless elsewhere stated, voucher specimens are in the collector’s collections.
Taxonomic treatmenthttp://species-id.net/wiki/Perapion_connexum
Perapion connexum is of the same size and at first glance very similar to the common in Europe Perapion curtirostre, from which it differs in a black tone of body integument (evidently grey in curtirostre), almost cylindrical and distinctly curved rostrum (thickened in basal half and nearly straight in curtirostre, as in Figs 5, 6, 9, 10), narrower subconical head, puncturation of vertex rugose and indefinite, smaller and not elongate scutellum (scutellar shield), slenderer tarsi and in male metatarsi devoid of ventral spines. It strongly resembles Aizobius sedi in the colour of integument, but the latter species has different frons sculpture, with well defined punctures and long median fovea, pronotum distinctly rounded at sides, and a ventrally spined basal segment on all male tarsi. See the key to species of Perapion occurring in Central Europe given below.

1–4 Perapion connexum (Schilsky), female 1 total view 2 antenna 3 tarsal claws 4 head with rostrum, lateral view 5 Perapion curtirostre (Gyllenhal), female head with rostrum, lateral view. 2–4 SEM photos.

6, 7 female anterior half of body, dorsal view 6 Perapion curtirostre (Gyllenhal) 7 Perapion connexum (Schilsky) 8, 9 male head with rostrum, dorsal view 8 Perapion connexum 9 Perapion curtirostre 10, 11 same in lateral view 10 Perapion curtirostre, 11 Perapion connexum.
Body length 2.0–2.3 mm.
Integument and vestiture. clearly black with slight “oily” glint (Fig. 1). Body covered with sparse and extremely fine white-semitransparent hair-like scales, on pronotum as long as diameter of the largest punctures, on elytral disc not longer than half interval’s width and unordered on intervals, not aggregated in any part of elytra, slightly denser on mesothoracic epimera and anepisterna, along metanepisterna condensed to form a thin white line. Entire body surface with dense microreticulation, scale-like and rough on head and the basal half of rostrum.
Rostrum in dorsal view subcylindrical with obtuse widening at antennal insertion, obscuredly punctured throughout, except distal third completely mat.
Head narrow, subconical, nearly as long as wide, about 1.5× narrower than pronotum (Figs 7, 8); eyes gently convex; frons slightly depressed in middle, with a few indictinct strigae partly obscured by dense microsculpture; puncturation on vertex lacking or indefinite, rarely with few punctures much smaller than on pronotal disc; head ventrally between eyes evenly scale-like microsculptured, without irregular asperities.
Antennae short and thin, with large club nearly as long as six distal funicular segments combined, 2.10–2.25× as long as wide, having fused segments with their circular rims incomplete (Fig. 2); pedicel 1.4–1.6× longer than wide, twice as long as next segment, segments 2, 3 minute and weakly elongate, segments 4, 5 isodiametric, 6 sligthtly, and 7 markedly transverse.
Pronotum small, slightly shorter than wide, with weakly rounded sides, at base 1.1–1.2× as wide as at apex, coarsely punctured, the punctures usually of 3–4 combined ommatidia size, with flat, heavily and somewhat roughly microreticulate interspaces; prescutellar fovea not wider than single puncture, as long as 3–4 neighbouring punctures combined.
Scutellar shield small, isodiametric (Fig. 7).
Elytra widest clearly behind mid-length, 1.6–1.7× longer than wide, 3.4–3.8× as long as pronotum, with deeply impressed catenulate-punctate striae, on elytral disc half as wide as intervals; intervals flat, barely punctate; specialised setae single on 7th and 9th interval.
Wing without radial window.
Ventrites. Metaventrite and abdominal ventrites I, II microreticulate and evently punctate, shiny, the punctures much smaller than on pronotal disc, well over a diameter apart from each other; abdominal ventrites III–V with strong, scale-like microsculpture.
Legs slender; profemur 0.80–0.85× as thick as rostrum; protibia widening from base to apex, with obsolescent apical tuft of setae; tarsi slender, protarsus 3.15–3.40× as long as wide; claws untoothed, thickened basally (Fig. 3).
Male. Rostrum slightly shorter than pronotum, 2.20–2.35× longer than wide, in profile almost straight and somewhat wedge-like, distinctly narrowing apicad in distal half (Fig. 11). Antennal insertion at basal 0.38–0.42 of rostrum. Abdominal ventrite V very broadly rounded apically. Metatarsus unarmed. Pygidium half exposed, with very broad complete transverse sulcus. Terminalia only slightly different from those of Perapion curtirostre, mainly in more elongate tegminal plate and aedeagus. Sternite VIII broad, with very short and indistinct lobes. Sternite IX with slightly asymmetrical fork half as long as apodeme. Tegmen with phallobase as long as apodeme; tegminal plate fused, short, devoid of macrochaetae, with broadly and very deeply emarginate prostegium. Aedeagus short and flattened, with pedon about 4.5× as long as wide, membranous tectum and free apophyses less than 0.2× as long as pedon; endophallus finely and more or less evenly microspinose.
Female. Rostrum 1.00–1.15× as long as pronotum, 2.60–2.75× longer than wide, in profile distinctly curved and equally high along its length (Fig. 4). Antennal insertion at basal 0.35–0.39 of rostrum. Abdominal ventrite V narrowly rounded apically. Tergite VIII broad and strongly transverse, uniformly slerotized. Sternite VIII with large and broad basal arms. Gonocoxites less than 2.5× longer than wide, without median string of sclerotisation; styli slightly elongate, shortly setose apically.
Poland (E): Stare Stulno (51.3714°N, 23.6628°E), 1 VIII 2000, 4 exs, 2 VIII 2000, 10 exs, 5 VIII 2000, 10 exs, 7 VI 2001, 1 ex., 31 VII 2001, 16 exs; Rudka nr. Wola Uhruska (51.2761°N, 23.6694°E), 15 VII 2002, 1 ex.; Wołczyny (51.4392°N, 23.6656°E), 6 VII 2002, 2 exs; Orchówek-Obłonie (51.5291°N, 23.5950°E), 7 VII 2002, 2 exs; Sobibór (51.4680°N, 23.6599°E), 6 VII 2002, 13 exs; Kosyń (51.3903°N, 23.5750°E), 12 VII 2002, 7 exs; Hniszów (51.2646°N, 23.7119°E), 15 VII 2002, 2 exs – all leg. et coll. MW.
Ukraine (W): Podolia: Zvenihorod at Dniester riv., 48.5500°N, 26.2833°E, 25 VI 1996, 2 exs; Kamyanets Podilskiy, 48.6667°N, 26.5667°E, 26 VI 1996, 2 exs – all leg. et coll. MW;
Hungary: Borsod-Abaúj-Zemplén county: Füzér, Hosszú-rét (48.5644°N, 21.4324°E), 21.VII.2005, 3 ex., leg. Hegyessy G & S; Alsószuha, Hideg-kút-völgy (48.3586°N, 20.5144°E), 17 VI 2003, 5 exs, leg. HG – coll. AP (2 ex) and KMS (3 exs); Zalkod, Erkecse (48.1818°N, 21.4541°E), 10.VII.1993, 1 ex, leg. HG – coll. KMS; Szalonna, Köszvényeskút (48.4612°N, 20.7086°E), 10.V.2007, 1 ex, leg. HG – coll. KMS; Tornaszentandrás: Mile-völgy (48.5066°N, 20.7853°E), 10.V.2007, 1 ex, leg. HG – coll. KMS; Mád, Becsek (48.1826°N, 21.3056°E), 10.IV.2008, 1 ex, leg. HG & AP – coll. AP; Taktaszada: Ökör-mező (48.1122°N, 21.1504°E), 11.VI.2008, 1 ex, leg. HG – coll. KMS.
Slovakia (S, E): Železné env., Tornaľa - Starňa (48.4167°N, 20.4000°E), 26 V 2006, 1 ♂, leg. et coll. T. Kopecký; Zemplínské Kopčany (48.5833°N, 21.8833°E), 14 VI 2000, 1 ♂ 1 ♀, leg. P. Boža – coll. S. Benedikt, 20 V 2002, 7 ♂♂ 7 ♀♀, leg. M. Mantič – coll. M. Mantič & KS; Turňa nad Bodvou (48.6000°N, 20.8667°E), 9 VI 2001, 1 ♂, leg. R. Fornůsek – coll. S. Benedikt.
Russia: Kursk, 1 ex.; Orel [Oryol], 3 exs; Nikitskoe near Voronezh, 1 ex. – all coll. F. Schubert (in Naturhistorisches Museum Wien). W Siberia: Novosibirsk Area, Kochenevo distr., 43 km WNW of Kochenevo, Sektinskoye Lake, 27.05.1998, leg. R. Dudko & A. Legalov, det. A. Legalov, 6 exs – coll. KS (2 ♂♂ 2 ♀♀) and M. Koštál (1 ♂ 1 ♀). Rostov reg.: Krasny Sulin distr.: Donleskhoz env. (47.8627°N, 40.2405°E), 12 VI 2004, 1 ex., leg. D. Kasatkin – coll. MW.
Kyrgyzstan: Chüy province: Ala-Archa valley (42.6000°N, 74.4833°E), ca. 30 km S of Bishkek, above 1300 m alt., 4 VI 2003, 3 exs., leg. R. Królik – coll. MW.
Austria?, Hungary*, Kazakhstan, Kyrgyzstan*, Moldova, Poland (E), Russia (Central and South European Territory, Western Siberia), Slovakia (S and E), Ukraine, Uzbekistan (first records herein marked with asterisk).
The occurrence of Perapion connexum in Poland, as based on the abovementioned data, was earlier generally announced by
Rumex confertus is an invasive plant in Europe, and its natural range ends probably close to Southeastern Poland, in Southern Slovakia and Hungary (
Following current distribution of the host plant, further expansion of Perapion connexum in Central Europe from the sites showed in Fig. 12 seems very likely especially through the territory of Poland, and it could be monitored quite easily by summer sweeping of mature inflorescences of Rumex confertus. The same method should be applied on stabilized localities of Rumex confertus in Austria to record its occurrence and expansion.
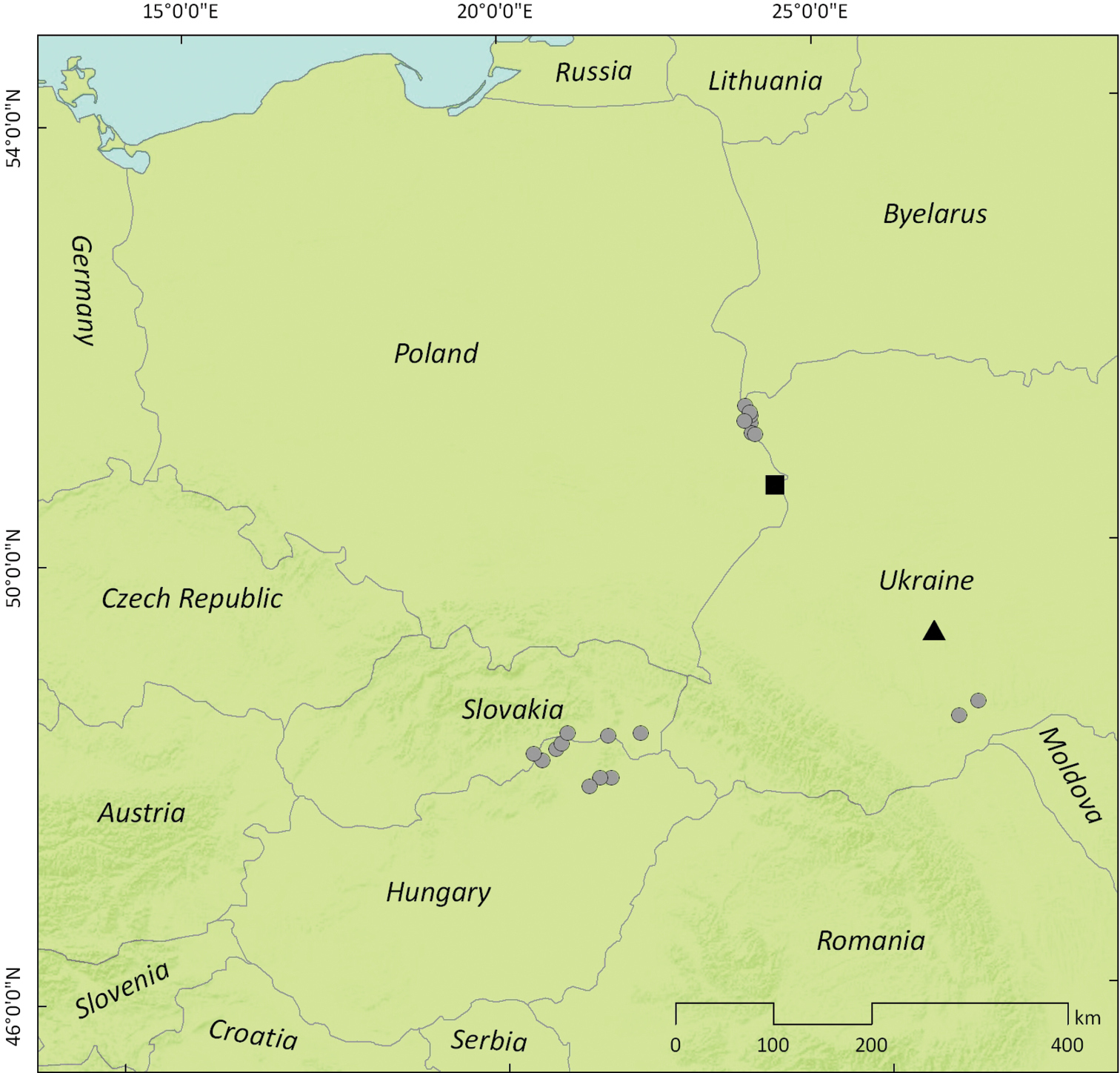
Westernmost recent localities of Perapion connexum (Schilsky) in Central Europe mentioned in the text (circles – new records; black square – record by
The key includes also Aizobius sedi (Germar, 1818), a related and morphologically similar species.
| 1 | Elytra with distinct metallic blue or green shine | 2 |
| – | Elytra black, concolorous with rest of body | 5 |
| 2 | Body longer than 2.5 mm. Scutellar shield elongate, subrectangular. Elytra 1.5–1.7× longer than wide. Male basal segment of metatarsus with a ventral spine | 3 |
| – | Body shorter than 2.5 mm. Scutellar shield isodiametric, rather triangular. Elytra nearly always 1.4-1.5× longer than wide. Tarsi without ventral spines | 4 |
| 3 | Abdominal ventrites 1-2 entire coarsely punctate, punctures nearly as large as those on pronotal disc. Pronotum 1.4-1.7× wider than head across eyes. Rostrum slightly curved, in male at least 2.5× longer than its basal width, in female longer than pronotum and more or less cylindrical | Perapion violaceum (Kirby) |
| – | Abdominal ventrites 1-2 sparsely and finely punctate, punctures several times smaller than those on pronotal disc. Pronotum at most 1.2–1.4× wider than head across eyes. Rostrum shorter, straight, in male tapering from base to apex and less than 2.5× longer than its basal width, in female as long as or shorter than pronotum, narrowing apicad from antennal insertion | Perapion hydrolapathi (Marsham) |
| 4 | Genae coarsely punctured and finely strigose. Punctures on head dorsum and pronotum mostly slightly elongate | Perapion affine (Kirby) |
| – | Genae largely impunctate, only transversely strigose. Punctures on head dorsum and pronotal disc round | Perapion marchicum (Herbst) |
| 5 | Frons with evident median sulcus, though not wider nor deeper than neighbouring punctures. Pronotum with markedly rouded sides and very thick walls, its disc distinctly convex. In male all tarsi ventrally spinose. On Crassulaceae (Sedum spp.) | [Aizobius sedi (Germar)] |
| – | Frons finely punctate or strigose. Pronotum with weakly rounded to nearly straight sides and thinner walls, its disc barely convex. At most male metatarsi with ventral spines. On Polygonaceae (Rumex spp., Polygonum spp.) | 6 |
| 6 | Body vestiture distinct, composed of cream-yellowish hair-like scales, on elytra forming a condensed patch on the outermost interval along metathoracic ventrite. Rostrum cylindrical, thinner than profemur. On Polygonum aviculare L. | Perapion lemoroi (Ch. Brisout) |
| – | Body vestiture less distinct, the scales finer, evenly confused on elytra. Rostrum at base as thick as or thicker than profemur | 7 |
| 7 | Head and basal half of rostrum with indistinct puncturation obscured by very dense rough scale-like microreticulation. Rostrum in female distinctly curved, in male somewhat wedge-like due to prominent septum between antennal insertions (Figs 4, 11). Scutellar shield isodiametric, flat (Fig. 7). Body vestiture very fine, beetle appears evidently black. Male tarsi unarmed. On Rumex confertus Willd | Perapion connexum (Schilsky) |
| – | Head and basal half of rostrum clearly punctured, the punctures dense and on vertex nearly as large as those on pronotal disc. Rostrum straight to slightly arched, in lateral view equally high throughout (Figs 5, 10). Scutellar shield elongate, furrowed (Fig. 6). Body vestiture more distinct, altogether with integument microsculpture giving beetle a greyish colouration. In male basal segment of metatarsus with small ventral spine | 8 |
| 8 | Body larger, 2.2-3.0 mm long. Rostrum longer, in male ca. 3×, in female ca. 3.5× longer than its width at antennal insertion, in female subequally wide at narrowest points before and behind antennal insertion. On Rumex acetosa L. | Perapion oblongum (Gyllenhal)‡ |
| – | Body smaller, 1.5-2.6 mm long. Rostrum in male ca. 2.5×, in female ca. 3.0× longer than its width at antennal insertion, in female at narrowest point basad of antennal insertion clearly wider than at narrowest point of apical half of rostrum (Fig. 6). On a wide range of Rumex spp., occasionally on Polygonum bistorta L. | Perapion curtirostre (Germar) |
† Generic definition after
‡ This species was synonymised with Perapion curtirostre by
We thank Roman Królik (Kluczbork) for delivery of the specimens from the expedition to Kyrgyzstan, and to Zygmunt Dajdok (Faculty of Biological Sciences, University of Wrocław) and Bruno Wallnöfer (Naturhistorisches Museum Wien) for their help and advice in collecting data on the distribution of Rumex confertus in Central Europe. Remigiusz Pielech is thanked for help in preparation of the distribution map, and Marek Halama for delivery of some botanical literature (both: Faculty of Biological Sciences, University of Wrocław).