





(C) 2012 C. Selby Herrin. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
A new species of Haemogamasus mites is described from Chubut, Neuquén and Río Negro Provinces, Argentina. It was collected primarily from rodents of the genus Abrothrix (65 of 77 collections): Abrothrix longipilis (63), Abrothrix olivaceus olivaceus (1) and Abrothrix olivaceus xanthorhinus (1). Additional collections were made from Geoxus valdivianus (5) and Loxdomtomys micropus (5). Possibly incidental or contaminate collections were recorded from Oligoryzomys longicaudatus (1) and Dromiciops gliroides (1). Most collections came from Río Negro Province (36), with 22 from Neuquén and 19 from Chubut. An identification key is provided to distinguish ♀s of this new species from ♀s of species from the western hemisphere, based on morphological characters.
ectoparasites, mites, Laelapidae, Haemogamasus, Argentina
Mites were collected by the junior author (RDS) during natural history studies of small mammals conducted in the Chubut, Neuquén and Río Negro Provinces, Argentina. Among those collected was a new species belonging to the genus Haemogamasus Berlese, 1889. This new species, Haemogamasus alongipilis, was collected primarily from rodents of the genus Abrothrix (65 of 77 collections): Abrothrix longipilis (Waterhouse) (63), Abrothrix olivaceus olivaceus (1) and Abrothrix olivaceus xanthorhinus (1). Additional collections were made from Geoxus valdivianus (5) and Loxdomtomys micropus (5). Possibly incidental collections were made from Oligoryzomys longicaudatus (1) and Dromiciops gliroides (1). The most specimens were taken from Río Negro Province (36), with 22 from Neuquén and 19 from Chubut. Mammal synonymies follow those of
In his major taxonomic classification treatment of Haemogamasinae,
In his studies of North American Haemogamasus liponyssoides, Radovski (1960) found 4 species with a distinguishing character that differs from all other Haemogamasus species. He found that Haemogamasus liponyssoides, Haemogamasus harperi, Haemogamasus occidentalis, and Haemogamasus keegani all have slender, untoothed chelae, and concluded that “They appear to constitute a closely related group within the genus, and are referred to here as the Haemogamasus liponyssoides complex.”
In a more recent treatment of the Haemogamasus species of North America,
We have chosen to follow the classification proposed by
The structural morphology and body setae nomenclature followed herein is basically from
Mammals were trapped with Museum Special snap traps (Woodstream Corporation®) baited with oatmeal. Traps were set out in the late afternoon and examined in the morning. Each trapped animals was placed in a plastic bag for later processing for ectoparasites. Ectoparasites were collected by exposing each animal to a stream of carbon dioxide while vigorously brushing over a funnel supported by a ring stand. A 7.5 cm × 15 cm plastic zip–lock bag containing 2 ml of 80% ethanol was affixed with tape to the bottom of the funnel. Ectoparasites (fleas, lice, mites, and staphylinid beetles) fell into the slick–walled funnel and were captured and preserved in the zip–lock bag containing an RDS–field number for the respective mammal host. Sealed bags were subsequently stored for longer periods in a bottle containing 80% ethanol. The funnel was thoroughly cleaned after each animal to preclude cross contamination of ectoparasites from one animal to the next.
Mite specimens were cleared in a solution of KOH and then put through a series of: distilled water, 70%, 85%, 95% & 100% ethyl alcohol, then methyl salicylate and finally xylene. The specimens were then mounted on slides in Canada balsam, and the slides were cured in a warming oven for several weeks in preparation for examination and identification.
Digital images in TIFF format were prepared by photographing with an Olympus BX61 Compound Microscope, Olympus CV12 digital camera using an Olympus Microsuite™ B3SV program. This photograph imaging system was also used to perform all measurements for the detailed descriptions of both ♀s and deutonymphs. All measurements are given in micrmeters (mµ) and all “µ” in the description. In many cases the measurement range is given as “ca. x”. We found that there is about a 5–10% variation in measurements of the same structure from specimen to specimen.
The TIFF images were cropped, enhanced, sized appropriately and saved in TIFF as well as JPG format with Adobe Photoshop CS5 Extended version 11.1. Line drawings illustrating diagnostic characters were prepared by printing the TIFF digital images, then tracing them on a light box, scanning the traced images and making necessary adjustment, including setae labeling, with Adobe Photoshop CS5.
The following acronyms for repositories may be associated with material listed under “Type Material” and under “Other Material Examined”: BYU—M. L. Bean Life Science Museum, Brigham Young University, Provo, Utah; MACN—Museo Argentino Bernardino Rivadavia de Ciencias Naturales, Buenos Aires, Argentina; and MCNLP—Museo de Ciencias Naturales de La Plata, La Plata, Argentina.
Resultsurn:lsid:zoobank.org:act:64F6027D-FC35-4638-AE03-8ED1FD954305
http://species-id.net/wiki/Haemogamasus_alongipilis
Figs 1-22Holotype, ♀: Argentina: Neuquén Province, Los Lagos Departamento, 4.8 km W, 12.2 km N Villa La Angostura (Parque Nacional Nahuel Huapi), elev. 840 m, (39°02'12.48"S, 70°18'25.68"W), trapline 9, mature Coíhue forest with bamboo, ex. Abrothrix longipilis, ♀, 07 XI 2006, R. D. Sage (slide RDS–18117, MACN); paratype, ♀: (same data as holotype, slide RDS–18117, MACN); paratypes, deutonumphs (2): (same data as holotype, slide RDS–18117, MACN); paratypes, ♀s (3) & deutonymphs (3): Argentina: Neuquén Province, Los Lagos Departamento, 3.5 km W, 12.8 km N Villa La Angostura (Parque Nacional Nahuel Huapi), elev. 830 m, trapline 10, mixed Coíhue/Ñire forest with bamboo, Abrothrix longipilis, 1 ♂ & 1 ♀, 6 XI 2006, R. D. Sage (slides RDS–18104 & 18106, MCNLP); paratype ♀: Neuquén Province, Aluminé Departamento, 3.8 km W, 0.7 km N Quillén, (Parque Nacional Lanín) elev. 980 m, (39°03'07.26"S, 70°20'42.24"W), trapline 4, Roble Pellín/Ñire forest with bamboo, ex. Abrothrix longipilis (Waterhouse), ♀, 13 X 2006, R. D. Sage (slide RDS–18030, BYU); paratypes, ♀s (2) & deutonymphs (4): Neuquén Province, Aluminé Departamento, 3.9 km W, 1.1 km N Quillén (Parque Nacional Lanín), elev. 1010 m, (39°03'07.26"S, 70°20'42.24"W), trapline 3, Roble Pellín forest with bamboo, ex. Abrothrix longipilis (Waterhouse), 2 ♂s, 17 X 2006, R. D. Sage (slide RDS–18039 & 18040, BYU); paratype, deutonymph: Argentina, Neuquén Province, Los Lagos Departamento, 4.8 km W, 12.2 km N Villa La Angostura (Parque National Nahuel Huapi), elev. 840 m, (39°02'12.48"S, ), trapline 9, mature Coíhue forest with bamboo, ex. Geoxus valdivianus Philippi, 1 ♂, 07 XI 2006, R. D. Sage (slide RDS–18112, BYU); paratype, deutonymph (1): Argentina, Neuquén Province, Los Lagos Departamento, 4.8 km W, 12.2 km N Villa La Angostura (Parque National Nahuel Huapi), elev. 840 m, (39°02'12.48"S, 70°18'25.68"W), trapline 9, mature Coíhue forest with bamboo, ex. Loxdontomys micropus (Waterhouse), ♂, 07 XI 2006, R. D. Sage (slides RDS–18113, BYU); paratypes, ♀ (1) & deutonymphs (10): Argentina, Neuquén Province, Los Lagos Departamento, 4.8 km W, 12.2 km N Villa La Angostura (Parque National Nahuel Huapi), elev. 840 m, (39°02'12.48"S, 70°18'25.68"W), trapline 9, mature Coíhue forest with bamboo, ex. Abrothrix longipilis (Waterhouse), 3 ♂s & 2 ♀s, 07 XI 2006, R. D. Sage (slides RDS–18114, 18116, 18117, 18118 & 18119, BYU); paratype, deutonymph: Argentina, Neuquén Province, Los Lagos Departamento, 4.8 km W, 12.2 km N Villa La Angostura (Parque National Nahuel Huapi), 840 m elev. (39°02'12.48"S, 70°18'25.68"W), trapline 9, mature Coíhue forest with bamboo, ex. Geoxus valdivianus Philippi, 1 ♀, 8 XI 2006, R. D. Sage (slide RDS–18121, BYU).
Other Material Examined (ordered by locality).
| 1 | Trapline 3, Roble Pellín forest with bamboo, 3.9 km W, 1.1 km N Quillén (Parque Nacional Lanín), elev. 1010 m, Aluminé Dpto., Neuquén Prov., Argentina, Richard D. Sage | 17 X 2006: RDS–18040 Abrothrix longipilis (♂) –1 deutonymph. 15 X 2007: RDS–18569, 18572 & 18573 Abrothrix longipilis (2 ♀s & 2 ♂s) – 4 deutonymphs. |
| 2 | Trapline 4, Roble Pellín/Ñire forest with bamboo, 3.8 km W, 0.7 km N Quillén (Parque Nacional Lanín), elev. 980 m, Aluminé Dpto., Neuquén Prov., Argentina, Richard D. Sage | 14 X 2007: RDS–18565 Abrothrix longipilis (♀) – 1 deutonymph. 16 X 2007: RDS–18565 Geoxus valdivianus (♀) – 1 deutonymph. |
| 3 | Trapline 9, Coíhue forest with bamboo, 4.8 km W, 12.2 km N Villa La Angostura (Parque Nacional Nahuel Huapi), elev. 840 m, Los Lagos Dpto., Neuquén Prov., Argentina, Richard D. Sage | 7 XI 2006: RDS–18112 Geoxus valdivianus (♂) – 1 deutonymph; RDS–18113 Loxdontomys micropus (♂) – 1 deutonymph. 8 XI 2006: RDS–18121 Geoxus valdivianus (♀) – 1 deutonymph. |
| 4 | Trapline 10, mixed Coíhue/Ñire forest with bamboo, 3.5 km W, 12.8 km N Villa La Angostura (Parque Nacional Nahuel Huapi), elev. 830 m, Los Lagos Dpto., Neuquén Prov., Argentina, Richard D. Sage | 6 XI 2006: RDS–18102 Abrothrix longipilis (♂) – 1 deutonymph; RDS–18103 Abrothrix olivaceus olivaceus (♂) – 1 deutonymph. |
| 5 | Trapline 6, in green bamboo/Coíhue/Ñire forest on Península Llao Llao, 0.4 km N of Parkguard station between Lagos Perito Moreno and Escondido (Parque Municipal Llao Llao), elev. 831 m, Bariloche Dpto., Río Negro Prov., Argentina, Richard D. Sage | 26 XI 2006: RDS–18144, 18145, 18148 & 18153 Abrothrix longipilis (3 ♂s & 1 ♀) – 4 deutonymphs. 27 V 2007: RDS–18433 & 18437 Abrothrix longipilis (1 ♂ & 1 ♀) – 2 ♀s, 1 deutonymph. 29 V 2007: RDS–18471 & 18475 Abrothrix longipilis (2 ♀s) – 4 deutonymphs. 23 X 2007: RDS–18581 Loxdontomys micropus (♀) – 1 deutonymph; RDS–18585, 18586, 18588 & 18598, Abrothrix longipilis (2 ♀s & 2 ♂s) – 5 deutonymphs; RDS–18593 Oligoryzomys longicaudatus (♀) – 2 deutonymphs. |
| 6 | Trapline 7, in green bamboo/Coíhue/Ñire forest on Peninsula Llao Llao, 0.3 km W of Parkguard station between Lagos Perito Moreno and Escondido (Parque Municipal Llao Llao), elev. 800 m, Bariloche Dpto., Río Negro Prov., Argentina, Richard D. Sage | 6 XI 2006: RDS–18127, 18128 Geoxus valdivianus (2 ♀s) – 2 ♀s & 1 deutonymph. 25 XI 2006: RDS–18130, 18131, 18132 & 18136 Abrothrix longipilis (1 ♂ & 3 ♀s) – 4 deutonymphs. 28 V 2007: RDS–18442 Loxodontomys micropus (♂) – 3 deutonymphs; RDS–18449, 18453, 18455, 18456 & 18461 Abrothrix longipilis (3 ♂s & 2 ♀s) – 9 ♀s & 7 deutonymphs. 1 XI 2007: RDS–18604 Dromiciops gliroides (♀) – 1 ♀; RDS–18608, 18609, 18610, 18612, 18615, 18616, 18620, 18621 & 18624 Abrothrix longipilis (5 ♂s & 3 ♀s) – 11 ♀s & 13 deutonymphs. |
| 7 | Trapline 1, mixed Ñire/Ciprés forest with bamboo, 5.5 km S, 2.8 km E Villa Futalaufquen (Parque Nacional Los Alerces), elev. 547 m, Futaleufú Dpto., Chubut Prov., Argentina Richard D. Sage | 26 X 2006: RDS–18071, 18072, 18078, 18079, 18082, 18083 & 18084 Abrothrix longipilis (4 ♂s, 3 ♀s) – 9 deutonymphs. 26–27 X 2006: RDS–18069 & 18078 Loxdontomys micropus (2 ♂s) – 3 deutonymphs. 13 XI 2007: RDS–18658 Abrothrix longipilis (♂) – 1 deutonymph. |
| 8 | Trapline 2, Ñire forest with bamboo, 1.5 km S, 0.5 km E Villa Futalaufquen (Parque Nacional Los Alerces), elev. 590 m, Futaleufú Dpto., Chubut Prov., Argentina Richard D. Sage | 25 X 2006: RDS–18059 Abrothrix olivaceus xanthorhinus (♂) –1 deutonymph; RDS–18060 Abrothrix longipilis (♂) – 1 deutonymph; RDS–18066 Abrothrix longipilis (♀) – 2 deutonymphs. 10 XI 2007: RDS–18628, 18630, 18633, 18635, 18638, 18640 & 18645 Abrothrix longipilis (6 ♂s & 2 ♀s) – 2 ♀s & 5 deutonymphs. |
♀s of Haemogamasus alongipilis possess a number of distinguishing characters: (1) Small body size (dorsal shield ca. 538 µ median long and ca.272 µ greatest width); (2) chelicera chelae (see Fig. 4) slender with untoothed (edentate) chelae; (3) sternal shield (see Fig. 4) approximately square in shape (length/width ratio 0.98), extending posteriorly to level of mid coxae III; (4) sterna shield with 6 pair plus 1 (13) accessory setae in addition to the usual 3 pair of setae; (5) no accessory setae on anterior margin of sterna shield (only st-1 setae on anterior margin); (6) gnathosomal, sternal shield, anal shield and leg setae smooth, none barbed; (7) posterior margin of sterna shield only slightly concave; (8) genitoventral shield (see Fig. 3) not unusually expanded posterior to coxae IV; (9) genitoventral shield with only 5 setae: The usual gv1 and gv2 pairs, plus 1 accessory seta centered at level posterior to gv2; (10) anal shield (see Fig. 2) with no accessory setae; and (10) the usual pair of paranal setae positioned well posterior to the anal orifice.
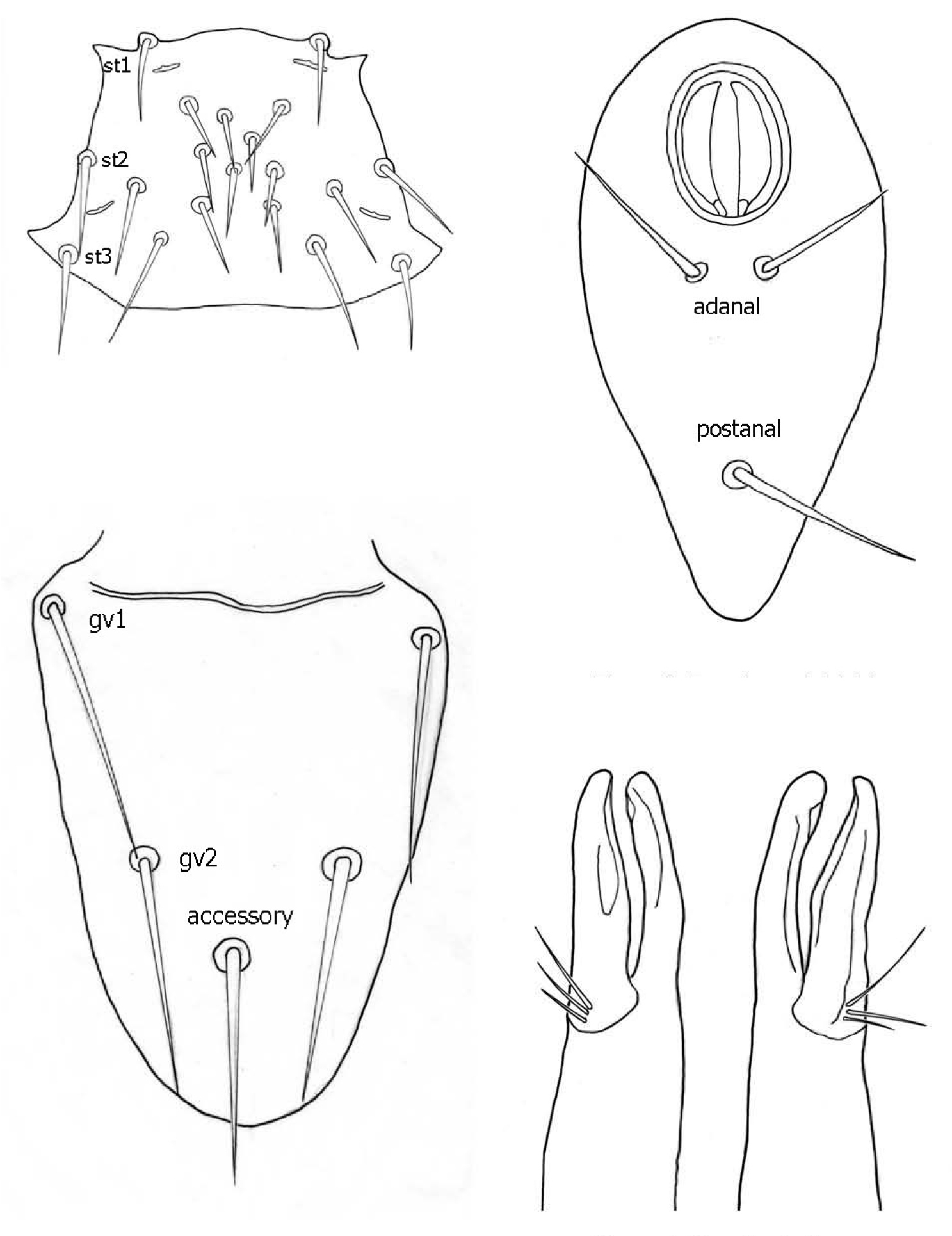
1 Female sternal shield 2 Female anal shield 3 Female genitoventral shield 4 Female chelicerae.
♀s of Haemogamasus alongipilis are separable from all Western Hemisphere Haemogamasus species by the following combination of characters: (1) Significantly smaller body size (dorsal shield ca. 538 µ median long and ca.272 µ greatest width); (2) sternal shield (see Fig. 1) approximately square in shape (length/width ratio 0.98); (3) No accessory setae on the anal shield (see Fig. 2); (4) paranal setae positioned at a level posterior to the anal orifice; and (5) genitoventral shield (see Fig. 3) shorter, not expanded posterior to gv1 or gv2 setae and with only 1 accessory seta centered at level posterior to gv2 seta. Haemogamasus alongipilis and Haemogamasus harperi share a single diagnostic character: Both have sternal shield accessory setae which distinguishes them from all other western hemisphere species.
Thus far, no ♂s of Haemogamasus alongipilis are known from the 77 collections in Argentina, probably because males are rarely collected; however many deutonymphs were collected and are described herein. It is interest to note that of the 77 collections, there were 99 deutonymphs collected whereas there were only 37 ♀s collected. This ratio of deutonumphs to females (3:1) is likely because the deutonymph stage is the migrating stage. This species is named after the predominant rodent host species, Abrothrix longipilis, from which 65 of the 77 collections were made.
The diagnostic key below for ♀s of the Western Hemisphere is adapted from
| 1 | Chlicerae slender with un-toothed chelae (edentate); anal shield elongate pyriform; no barbed setae on gnathosoma, sternal shield, anal shield and legs (liponyssoides group) | 2 |
| – | Chlicerae stout with strongly toothed chelae (dentate); anal shield broadly pyriform; some setae on gnathosoma, sternal shield, anal shield and legs barbed (reidi group) | 8 |
| 2 | Sternal shield with accessory setae | 3 |
| – | Sternal shield without accessory setae | 4 |
| 3. | Anal shield without accessory setae; genitoventral shield with only 1 accessory seta; dorsal shield length/width ca. 538 µ /ca.272 µ [Patagonia, Argentina] | Haemogamasus alongipilis sp. n. |
| – | Anal shield with accessory setae; genitoventral shield with 11-18 accessory setae; dorsal shield length/width ca. 966 µ /ca.570 µ [southeastern USA] | Haemogamasus harperi Keegan |
| 4 | Peritreme short (ca. 100 µ); genitoventral shield with 11-15 accessory setae on posterior one half of shield; tarsi abruptly narrowed distally [western USA] | Haemogamasus keegani (Jameson) |
| – | Peritreme longer (ca. 200 µ); genitoventral shield with more than 22 accessory setae on posterior one half of shield; tarsi not abruptly narrowed distally | 5 |
| 5 | Tectum with long distal and long lateral fimbriae; deutosternal groove narrow with one to eight teeth in anterior rows | 6 |
| – | Tectum with long distal and short lateral fimbriae; deutosternal groove broad with eight or more teeth in anterior rows | 7 |
| 6 | Sternal shield with posterior margin invaginated to level about midway between sternal setae II and III [eastern USA, southern Canada & Holarctic] | Haemogamasus liponyssoides liponyssoides Ewing |
| – | Sternal shield with posterior margin invaginated only to level of sternal setae III [California] | Haemogamasus liponyssoides hesperus Radovsky |
| 7 | Sternal shield with posterior margin nearly straight; genitoventral shield with 30 or more accessory setae; pilus dentilis and dorsal seta present [northwestern USA & southwestern Canada] | Haemogamasus occidentalis Keegan |
| – | Sternal shield with posterior margin invaginated nearly to level of sternal setae III; genitoventral shield with less than 20 accessory setae; pilus dentilis and dorsal seta absent [Alaska, USA] | Haemogamasus ghanii Williams et al. |
| 8 | Sternal shield with accessory setae; hypostomal setae I unbarbed; pilus dentilis setiform [Canada & USA and worldwide | Haemogamasus ambulans (Thorell) |
| – | Sternal shield without accessory setae; hypostomal setae I barbed or unbarbed; pilus dentilis variable | 9 |
| 9 | Sternal shield with posterior margin invaginated to between sternal setae I and II; hypostomal setae I unbarbed; setation of dorsal and genitoventral shields sparse; legs II-IV with relatively stout ventral setae [USA & Canada and worldwide] | Haemogamasus pontiger (Berlese) |
| – | Sternal shield with posterior margin never invaginated to or above sternal setae II; hypostomal setae I barbed; setation of dorsal and genitoventral shields dense; legs without stout ventral setae | 10 |
| 10 | Genitoventral shield very broad, flask-shaped, width about one-half length | 11 |
| – | Genitoventral shield shape variable, never extremely broad, width usually about one-third length | 12 |
| 11 | Genitoventral shield with accessory setae extending anteriorly to flap of shield; pilus dentilis very long, expanded laterally [eastern USA & Canada] | Haemogamasus longitartus (Banks) |
| – | Genitoventral shield with accessory setae only on posterior two-thirds of shield, none past genital setae; pilus dentilis long and basally swollen [California & Utah, USA] | Haemogamasus thomomysi Williams et al. |
| 12 | Pilus dentilis short, setiform; large corniculi (ca. 65 long); chelicerae with a conspicuous brush border at base; leg setae thickly barbed; most dorsal setae barbed [western USA] | Haemogamasus onychomydis (Ewing) |
| – | Pilus dentilis long, curved distally; smaller corniculi (ca. 30 long); no brush border at base of chelicerae; leg setae sparsely barbed; only posterior and marginal dorsal setae barbed [USA & Canada] | Haemogamasus reidi Ewing |
Figures 5-13
The following two line drawings show the general shape of the body as well as the most distinctive morphological characters. The dorsal view (Fig. 5) shows the outline of the dorsal shield with an anterior-lateral and a posterior-lateral cutout showing the relative size and number of setae. The ventral view (Fig. 6) shows the outline of the ventral shields with the relative size and number of setae associated with each.
Dorsum (Figs 7–8): Body elliptical with dorsal shield length/width ratio ca. 0.98; dorsal shield median length ca. 538 µ (515 –576 µ) and greatest width ca. 272 µ (250 –294 µ); anterior margin acutely rounded; lateral margins at level of anterior coxa I slightly concave; lateral margins at level between coxa I and coxa II moderately concave forming a rounded shoulder at level of coxa I; median lateral margins almost straight to slightly rounded; posterior lateral margins more strongly rounded; and posterior margin broadly rounded. Dorsal shield bearing numerous setae, far too many to count (see Figs 5–8). Marginal setae setaceous (ca. 24–26 µ long); medial setae setaceous (ca. 16–26 µ long); posterior terminal pair of setae (Z5) long and slender (ca. 54 µ); and J5 setae (just anterior to Z5) setaceous and longer (ca. 60 µ).
Gnathosoma (Figs 9): Greatest width at level of gnathosomal setae ca. 81 µ, and length from base to palpal trochanter ca. 114 µ. Gnathosomal setae setaceous and small, length ca. 19 µ; lateral hypostomal setae slightly longer (ca. 22 µ); medial hypostomal length ca. 39 µ; and distal hypostomal setae length ca. 18 µ. Distance between gnathosomal setae ca. 36 µ and distance from medial hypostomal setae to gnathosomal setae ca. 58 µ. Chelicera long and slender; moveable digit of chelae edentate; chelae length ca. 40 µ, with 2nd cheliceral segment ca. 111 µ long. Corniculi small (ca. 12-13 µ /3-5 µ) and somewhat horn shaped. Tectum of moderate length with 3 terminal fimbriae, middle much elongated. Deutosternal groove parallel sided with 8-10 transverse rows of tiny denticles. Pedipalps chaetotaxy 2–3–5– + terminal 10-14 tarsal setae; segment length/width as follows: Trochanter 41/20 µ, femur 28/27 µ, tibia 10/22 µ and tarsus 16/22 µ.
Venter (Figs 10–12): Tritosternum well developed with long, slender laciniae (ca. 108 µ) with base ca. 29 µ long and 17 µ wide. Presternal area reticulate, with no apparent minute spines.
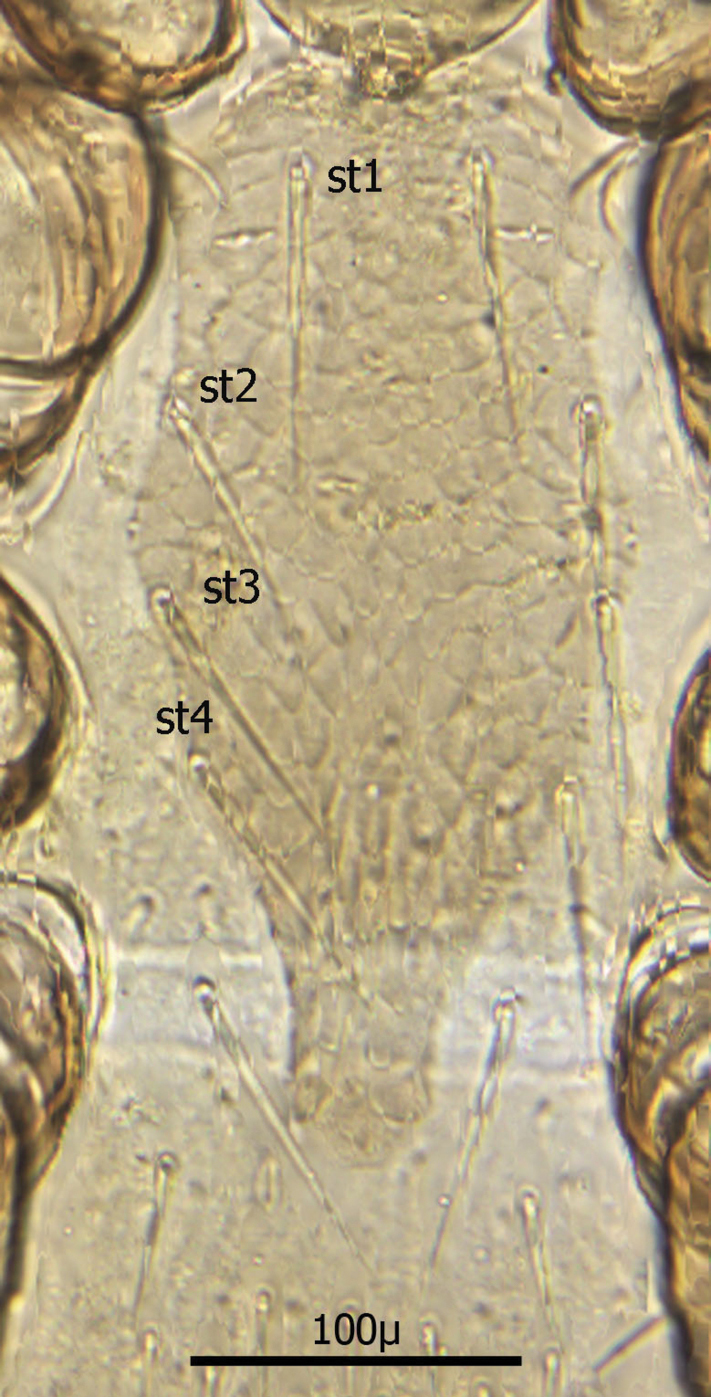
Sternal shield (Fig. 10) generally square in shape extending posteriorly to level of mid coxae III; length/width ratio 0.98; median length ca. 99 µ and width between coxae II ca. 101 µ. Anterior margin of sternal shield convex between st1 setae. Posterior margin slightly concave medially between st3 setae. Sternal shield bearing the usual 3 pairs of setae (st1, st2 and st3) plus 7 pairs of accessory setae and 1 single seta between and posterior to level of first pair of accessory setae. The usual 3 pairs of sternal setae are setaceous and subequal in length (ca. 36–42 µ). The accessory setae vary in length with anterior medial setae distinctly shorter than the posterior lateral setae.
Setae st1 on the anterior margin with concavities on both sides of each seta. Two pairs of distinct slit–like pores/ trichopores (lyriform in shape) associated with setae st1 and st2; the pair posterior to st1 generally parallel to anterior margin; the pair between st2 and st3 oriented at nearly 45 degree angle with medial end tipped up; third pair off shield on soft integument adjacent to st4.
Distance between st1 setae ca. 65 µ; distance between st2 setae ca. 116 µ; and distance between st3 setae ca. 129–235 µ. Distance between st1 and st2 setae ca. 46 µ and distance between st2 and st3 setae ca. 38 µ. Metasternal setae (st4) ca. 38 µ long; situated on small, oval metasternal shields at level of mid coxae III and just posterior to posterior–lateral margins of sterna shield.
Genitoventral shield (Fig. 11) tongue–shaped with 5 setae (2 pairs plus 1 single seta posterior to level of 2nd pair); gv1 setae positioned laterally adjacent to coxae IV; gv2 setae positioned a little more than half way between the gv2 setae and the posterior end of the genitoventral shield and not near margins; all 5 setae subequal in length (ca. 42–43 µ).
Distance between gv1 setae ca. 58 µ and distance between gv2 setae ca. 33 µ. Lateral margins of genitorventral shield gently converging with posterior end broadly rounded. Genitoventral shield length/width ratio 0.65; least width of the shield between coxae IV 70 µ; greatest width posterior to genitor ventral setae ca. 70 µ; greatest length (level between coxae IV to posterior end) ca. 100 µ; length from gv1 setae to posterior end ca. 82 µ. Parapodal shields posterior to coxae IV on unarmed venter oval to oblong (ca. 32 µ long and 17 µ wide). Unarmed venter thickly beset with setae; more medial setae short (16–23 µ long) and more lateral setae 60–66 µ long.
Anal shield (Fig. 12) not in juxtaposition with genitoventral shield; distance between genitoventral and anal shield ca. 86 µ. Anal shield length/width ratio 1.93; anterior margin strongly rounded with lateral margins gently rounded; length from anterior margin to postanal seta ca. 98 µ; greatest width at level of anal orifice ca. 50 µ; adanal setae situated at level posterior to anal orifice; distance between adanal setae ca. 13 µ; adanal setae setaceous ca. 22 µ long; distance from anterior margin to adanal setae ca. 45 µ; distance from adanal setae to postanal setae ca. 34 µ; postanal setae long setaceous ca. 33 µ long.
Shorter setae of unarmed venter length 16–23 µ; longer setae length 60–66 µ. Stigma of peritreme (Fig. 13) located lateral to coxae III and IV with only a short posterior peritremala extension; anterior extension of peritreme narrowing somewhat and reaching anteriorly to level slightly anterior to coxae II; total length of peritreme ca. 235 µ.
Legs The length and width of leg segments is given in Table 2 below. All measurements were made ventrally. Length measurements were made mid-ventrally and widths were made at the widest point. The measurements are approximate because there is about a 5–10% variation in measurements from specimen to specimen.
The leg segment chaetotaxy for females is shown in table 3 below. The chaetotaxy formula for each leg segment is as follows: al — d/v — pl (anterior laterals — dorsals/ventrals — posterior laterals). The total for each leg segment is in parenthesis bolded.
Leg I: Coxae I greatest width ca. 54 µ and mid–ventral length ca. 55 µ; distal setae setaceous and ca. 25 µ long; proximal setae setaceous and ca. 31 µ long. Proximal ventral seta of trochanter I slender setaceous ca. 34 µ long, and distal seta ca. 26 µ long. Distal dorsal setae of femur I setaceous but with one short (ca. 18 µ) and the other almost twice as long (ca. 30 µ). Proximal–most dorsal seta of genu I with one short (ca. 12 µ) and other longer (ca. 28 µ). Tarsus I with all setae setaceous.
Leg II: Coxae II greatest width ca. 91 µ and mid–ventral length ca. 27 µ; anterior seta setaceous and short (ca. 24 µ); posterior seta setaceous and much longer (ca. 39 µ). Ventral setae of trochanter setaceous and proximal seta much longer (ca. 31 µ) than distal (ca. 15 µ). Two distal dorsal setae of femur II slender setaceous and much longer (ca. 23–24 µ) than any others. Proximal dorsal setae of genu II setaceous and not unusually long. Tarsus II with all pre–apical seta setaceous.
Leg III: Coxae III greatest width ca. 79 µ and mid–ventral length ca. 25 µ. Anterior seta setaceous and slightly longer than posterior (ca. 25 µ); posterior seta setaceous and shorter (ca. 24 µ). Anterior distal setae of trochanter setaceous and ca. 27 µ long; posterior seta of trochanter slender setaceous (ca. 18 µ long). Setae of femur III all setaceous and of approximate equal length (18–20 µ). All setae of genu III setaceous and not unusually long (16–24 µ). Tarsus III with two pre–apical seta large and more spine–like and others setaceous.
Leg IV: Coxae IV greatest width ca. 65 µ and mid–ventral length ca. 29 µ; single seta setaceous and short ca. 17 µ long. Two anterior distal setae of trochanter setaceous (21–28 µ long); posterior proximal seta setaceous and longer (ca. 31 µ); posterior distal seta smaller (ca. 22 µ long). Distal setae of femur IV stout setaceous (ca. 21 µ long); posterior distal setae setaceous and longer (ca. 32 µ). Anterior distal setae of genu IV longer setaceous (ca. 24 µ), with all others setaceous and smaller (17–20 µ). Tarsus IV with pre–apical seta stout setaceous and others more slender setaceous.
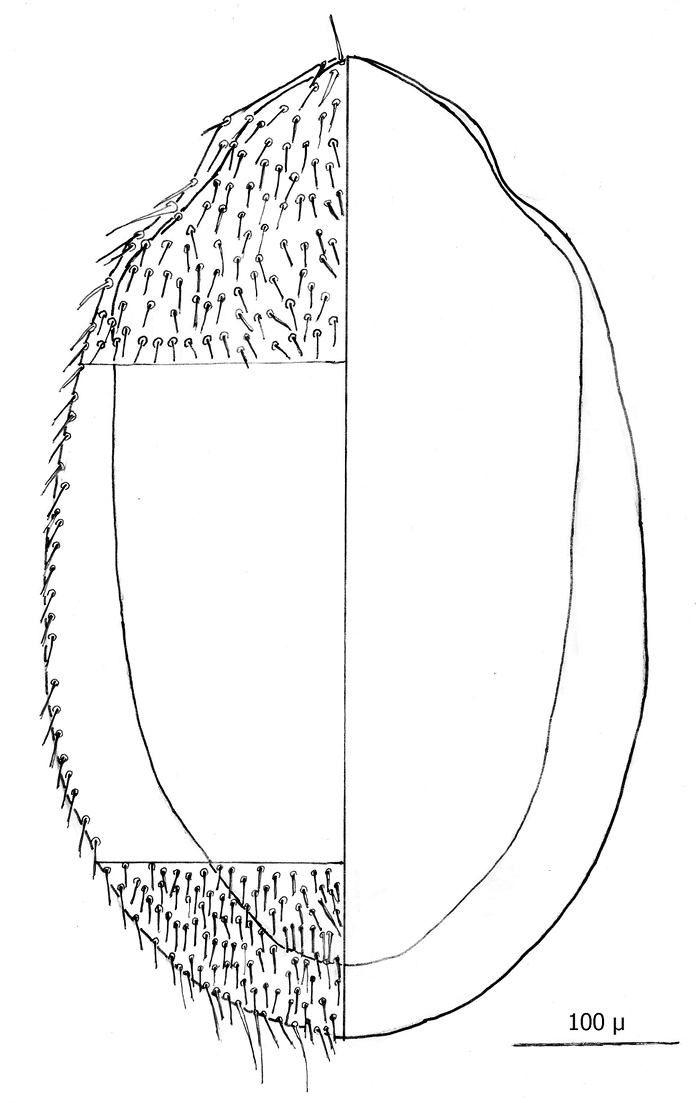
Line drawing of ♀ dorsum.
Line drawing of ♀ venter.

Anterior dorsum.
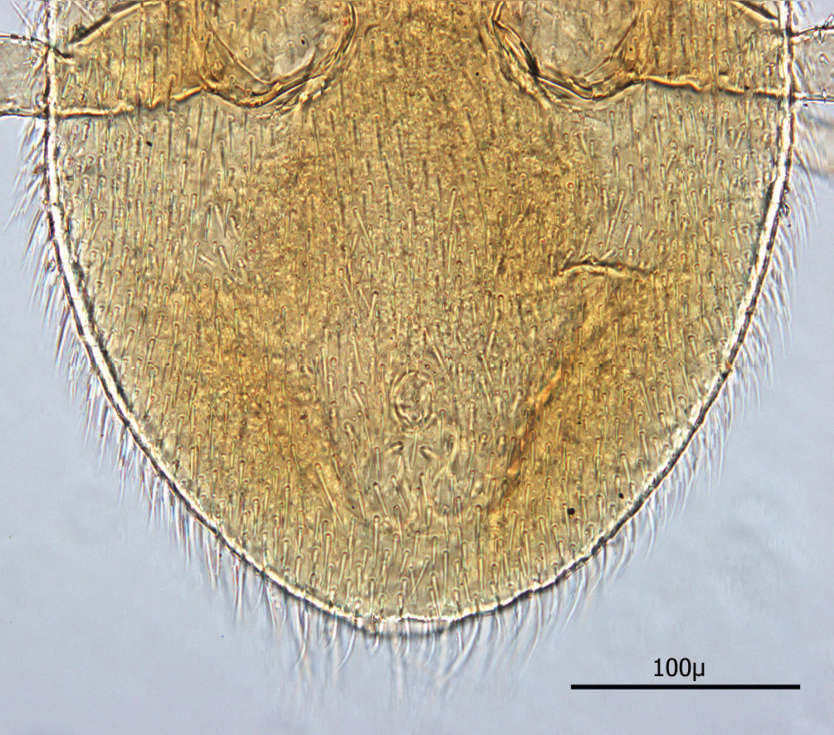
Posterior dorsum.
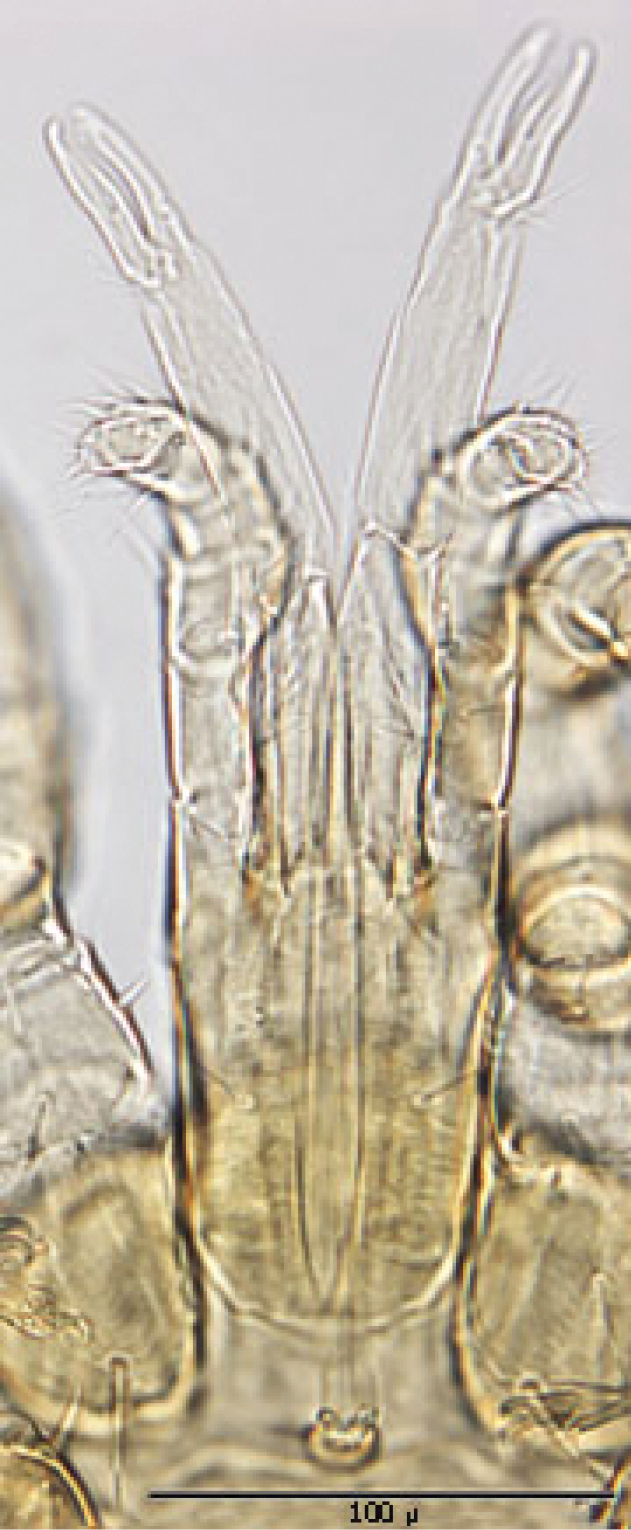
Ventral gnathosoma.

Sternal shield.
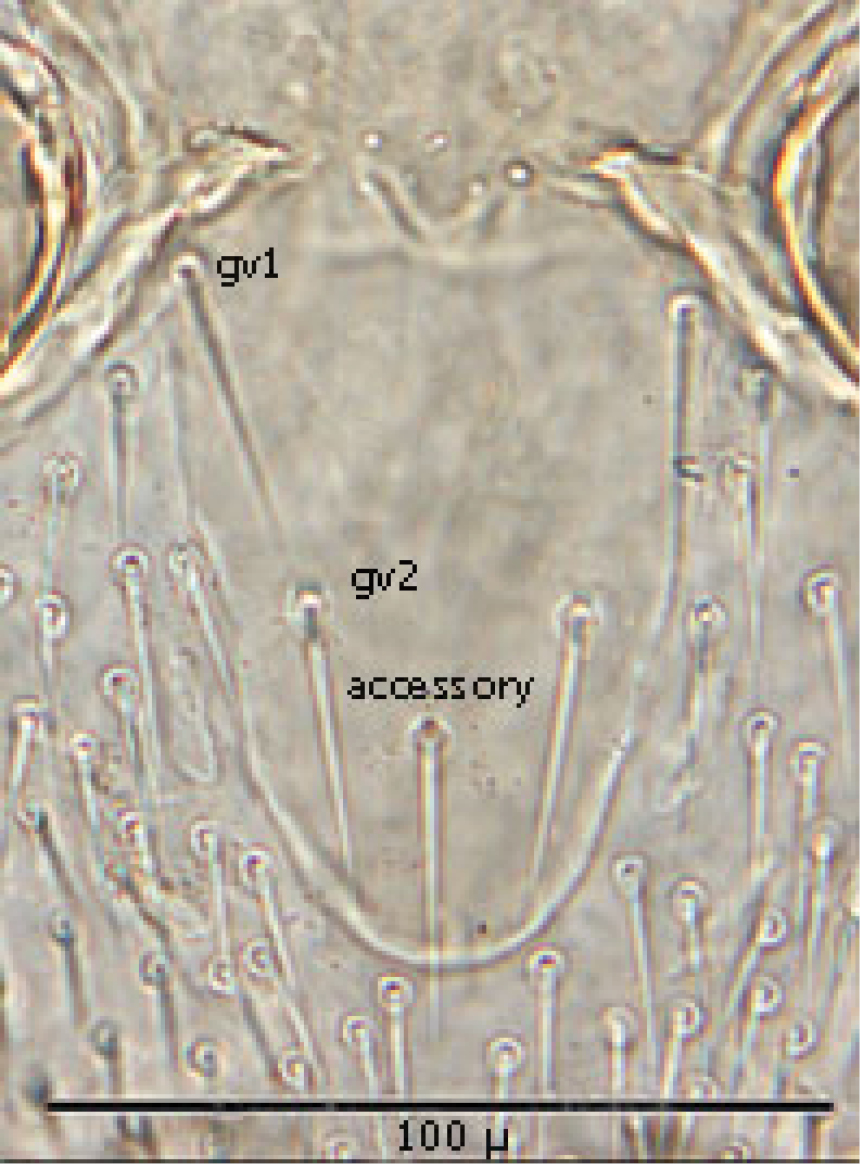
Genitoventral shield.
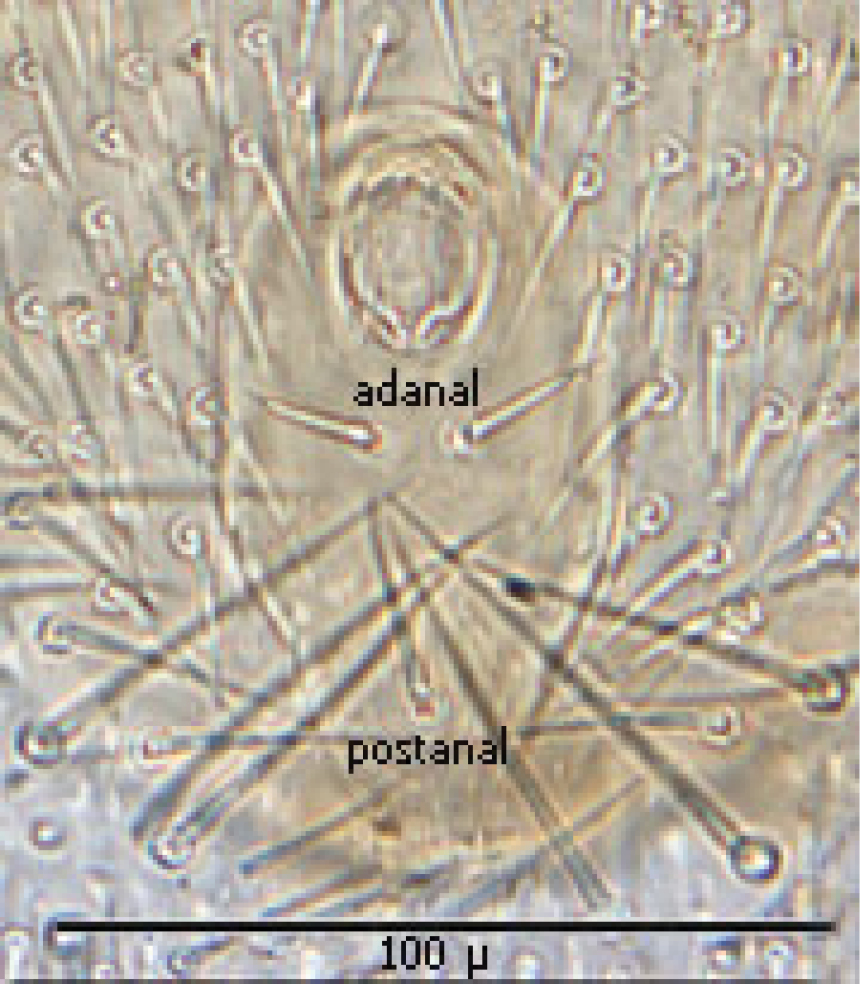
Anal shield.
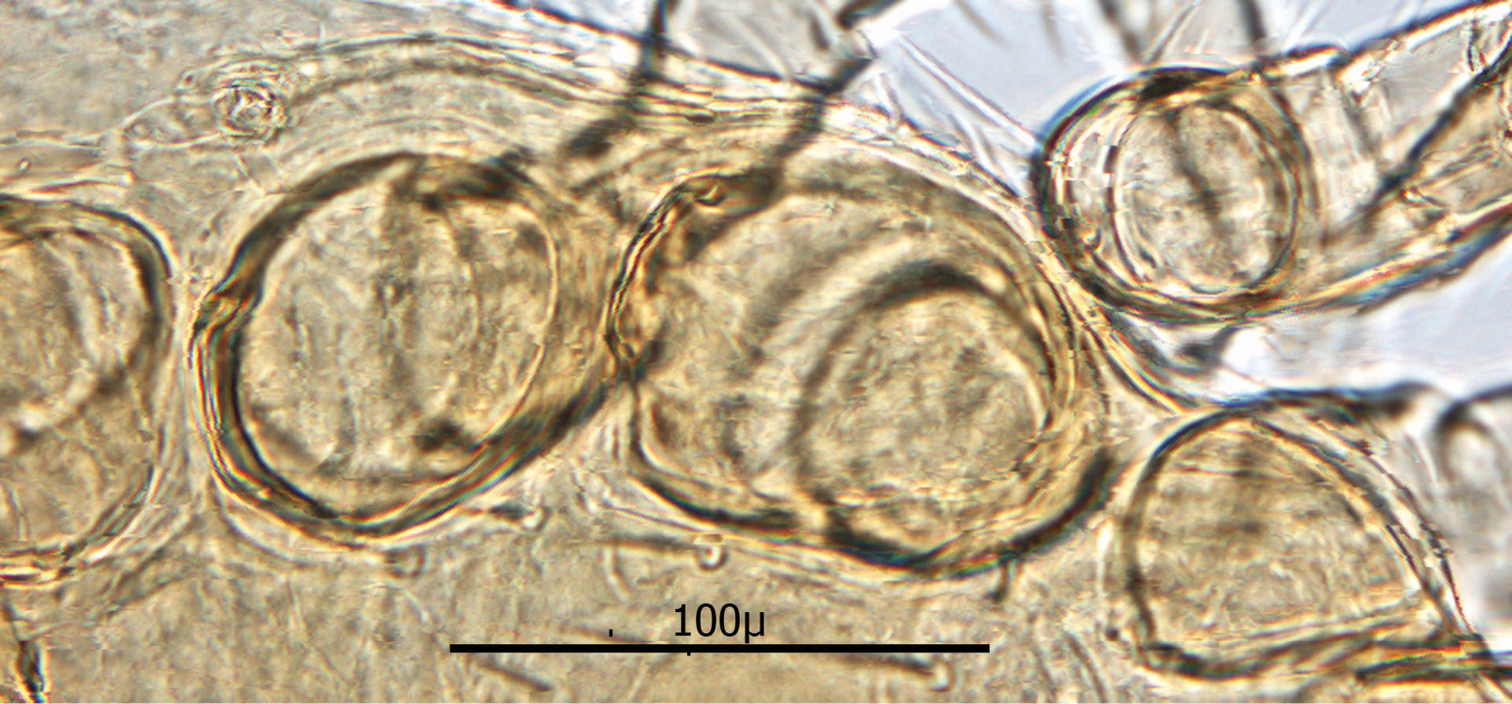
Peritreme.
Figures 14–22
Diagnostic characters for deutonymphs:
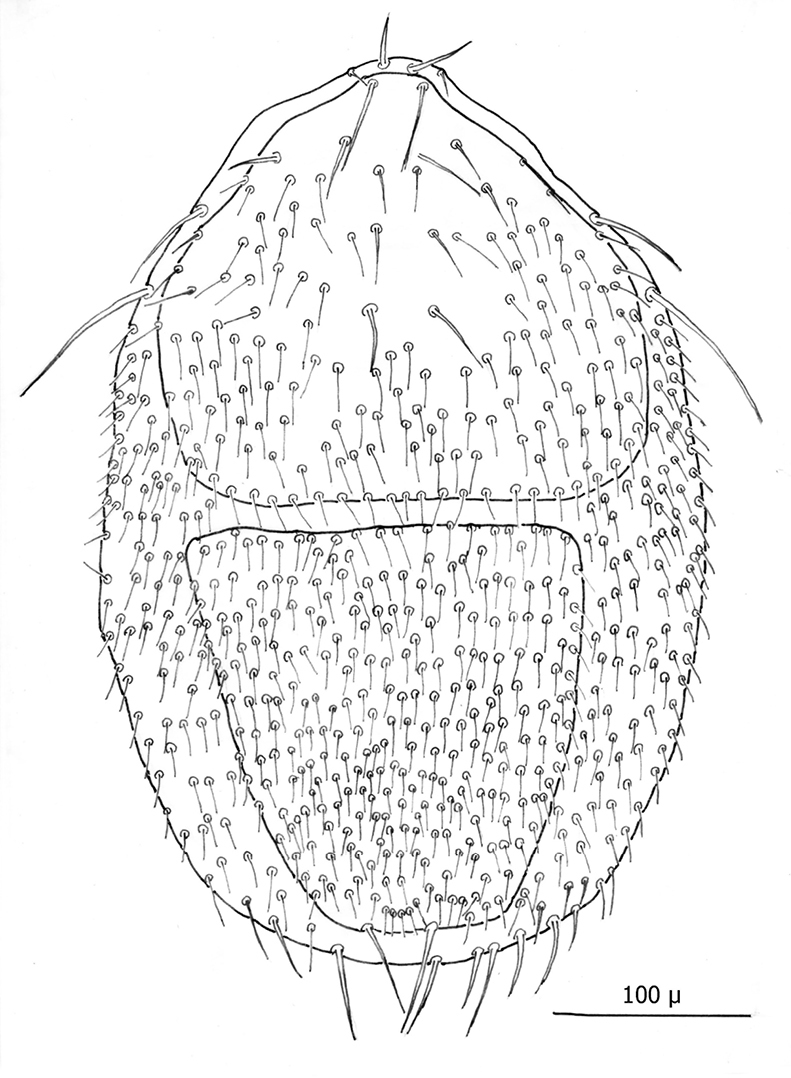
(1) Body elliptical and significantly larger than ♀s: length ca. 707 µ, and width at level of coxae III ca. 430 µ.
(2) Dorsal shield divided at level between coxae III and IV, with both dorsal shields thickly beset with setae.
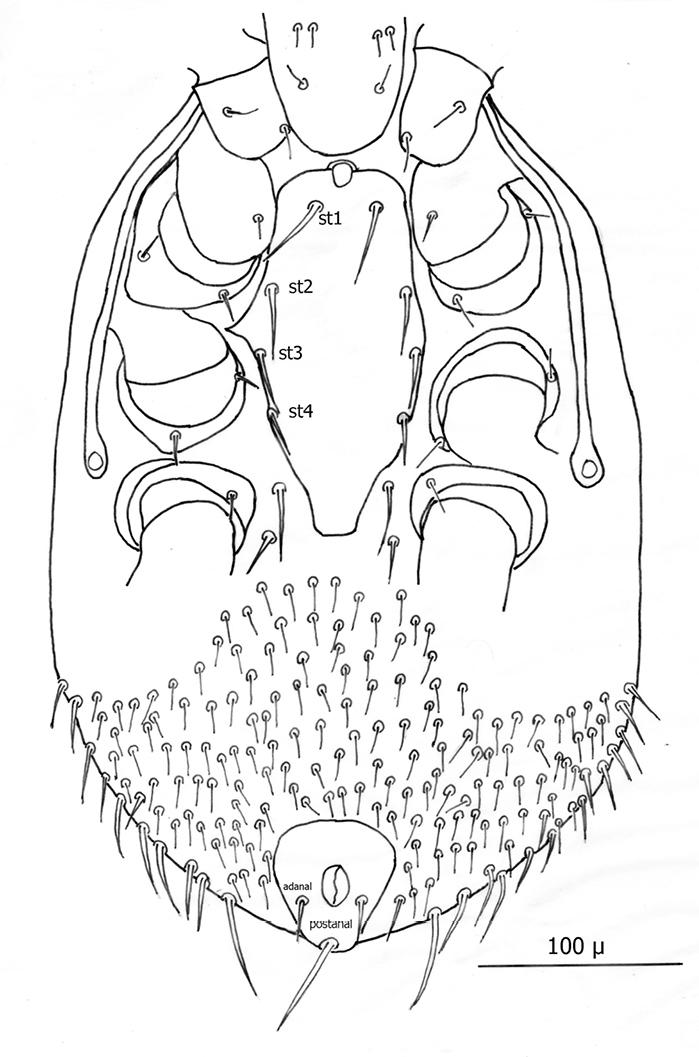
(3) Ventral shield (no separate sternal and genitoventral shields) elongate, extending from level of coxae I to posterior 1/3 of coxae IV and tapering strongly between coxae IV to a blunt point posteriorly.
(4) Ventral shield with 4 pairs of setae, which are the three pairs of usual sternal setae and the single pair of metasternal setae.
(5) Two pairs of ventral setae on soft integument between posterior extension of ventral shield and coxae IV; second pair posterior to ventral shield and adjacent to posterior of coxae IV.
(6) Anal shield somewhat diamond shaped with length and width about equal; adanal setae short not extending to base of postanal seta, which is very long (about 3½ times as long as paranal setae).
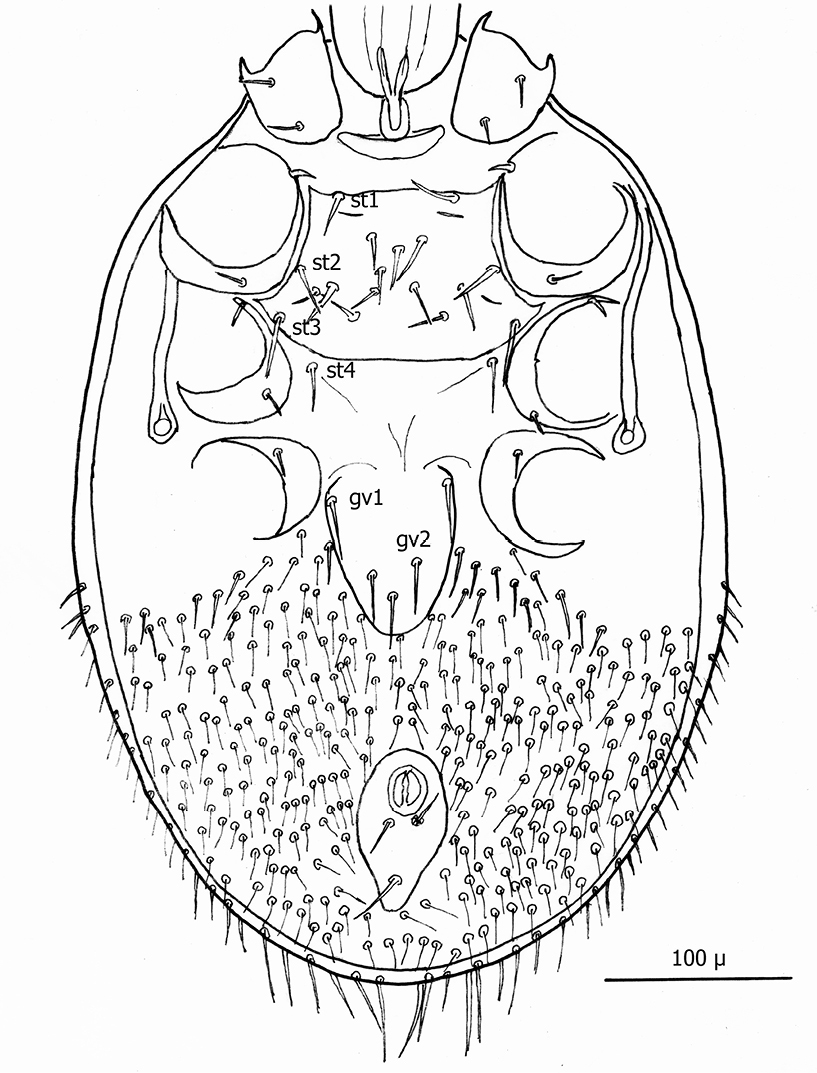
The following two line drawings show the general shape of the body as well as the most distinctive morphological characters. The dorsal view (Fig. 14) shows the outline of the dorsal shields showing the relative size and number of setae. The ventral view (Fig. 15) shows the outline of the ventral shields with the relative size and number of setae associated with each.
Dorsum (Figs 16–17): Body elliptical with two dorsal shields divided at level between coxae III and IV. Anterior and posterior dorsal shields combined length/width ratio ca. 1.75. Anterior dorsal shield length ca. 390 µ (371–442 µ) and greatest width ca. 447 µ (389–500 µ). Posterior dorsal shield length ca. 364 µ (323–402 µ) and greatest width ca. 341 µ (328–359 µ). Anterior tip of anterior dorsal shield pointed with only one “shoulder” at level of r4 setae. Anterior–lateral margins somewhat concave between anterior tip and “shoulder”; lateral margins posterior to “shoulder” broadly rounded. All dorsal setae setaceous and of varying lengths: Anterior marginal setae (r2, r3 & r4) shorter (35–43 µ long) than central setae; anterior central setae (i2, i3, i4, s1& s2) 73–91 µ long, and posterior central setae (j2, j3 & j4) shorter (38–42 µ long); Posterior marginal setae somewhat longer than anterior marginal (48–51 µ long); posterior terminal pair of setae (Z5) long and slender (81–82 µ long), and J5 setae (just anterior to Z5) short about same as j2–j4 (34–35 µ long).
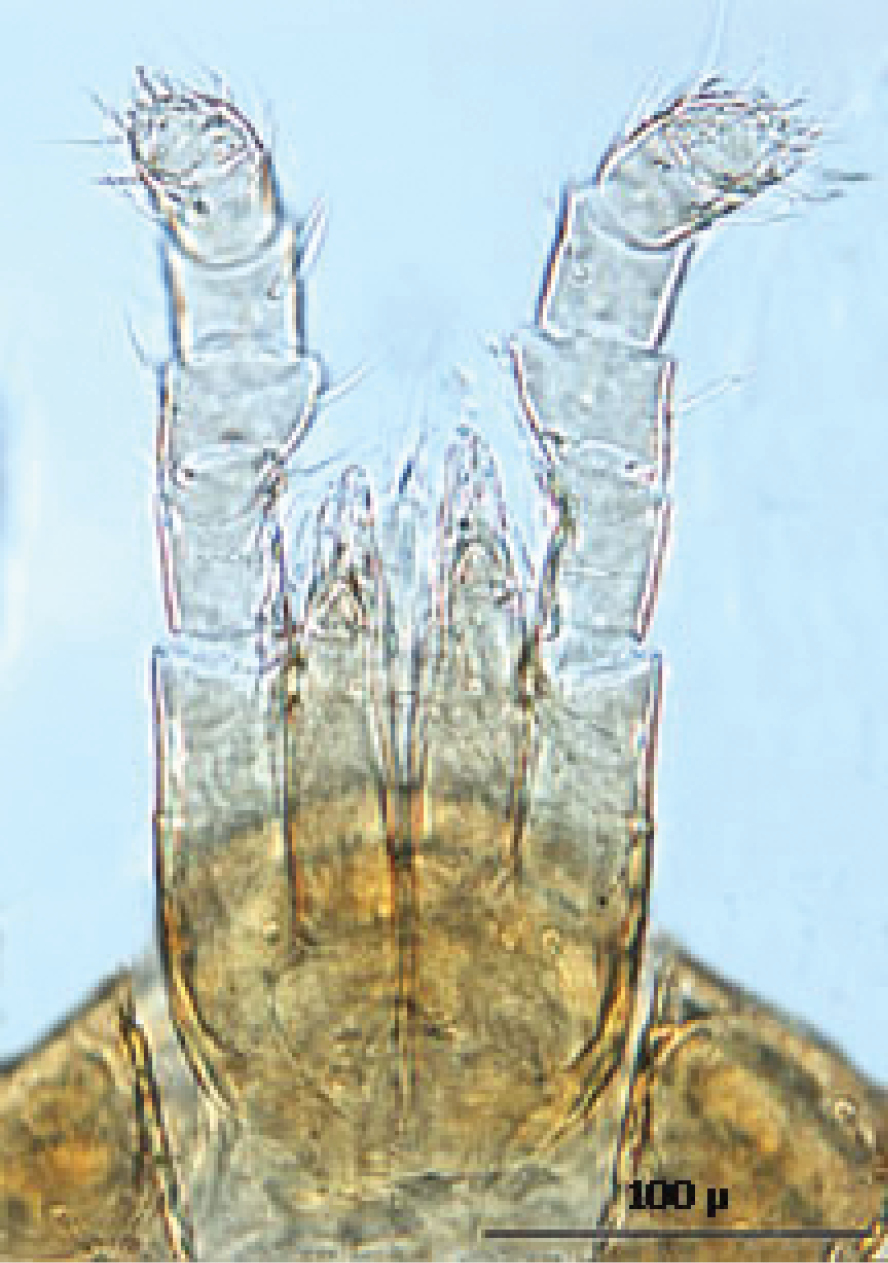
Gnathosoma (Figs 18—19): Greatest width at level of gnathosomal setae ca. 124 µ, and length from base to palpal trochanter ca. 119 µ. Gnathosomal setae setaceous ca. 56 µ long. Hypostomal setae all setaceous; lateral hyposomal setae ca. 24 µ long; medial hyposomal 48–58 µ long; and distal hyposomal setae 20–23 µ long. Distance between gnathosomal setae ca. 72 µ, and distance from medial hypostomal setae to gnathosomal setae ca. 63 µ. Moveable digit of chelae minutely toothed; chelae ca. 37 µ long, with 2nd cheliceral segment ca. 116 µ long. Corniculi distinctly horn shaped (ca. 22/5 µ). Tectum of moderate length with 3 terminal fimbriae, middle much elongated. Deutosternal groove parallel sided with 14-16 transverse rows of possibly minute denticles. Pedipalps chaetotaxy 2–5–6–9 + terminal 12-14 tarsal setae; segments length/width as follows: Trochanter 29/32 µ, femur 43/39 µ, genu 23/37 µ, tibia 17/30 µ and tarsus 30/30 µ.
Venter (Figs 20–22): Tritosternum well developed with long, slender laciniae (ca. 129 µ) with base ca. 11 µ long and 18 µ wide. Deutonymphs do not have separate sternal and genitoventral shields, but rather a single ventral shield that is elongated and extending from level between coxae I and II to level midway between coxae IV. This ventral shield (Fig. 20) has 4 pairs of setae, which represent the three pairs of usual sternal setae and the single pair of metasternal setae of. Ventral shield median length ca. 306 µ; width between coxae II ca. 147 µ; width between coxae III ca. 123 µ; width between coxae IV ca. 98 µ. Anterior margin of ventral shield irregular with concavity at level of tritosternum base; posterior end of ventral shield narrowly rounded posterior to a pair of ventral setae that flank the shield on the soft integument. two pair of ventral setae are posterior to ventral shield on the soft integument and adjacent to posterior 1/3 of coxae IV. All ventral shield setae setaceous and rather long: Gv1 setae 96–100 µ; gv2 setae ca. 99 µ; gv3 setae ca. 100 µ; and gv4 setae 93–94 µ; setae adjacent and posterior to ventral shield ca. 93–94µ long. One pair of slit–like pores/ trichopores (lyriform in shape) posterior to setae gv1; pores not evident adjacent to other ventral shield setae. Distance between gv1 setae ca. 51 µ; distance between gv2 setae ca. 122 µ; distance between gv3 setae ca. 113 µ; and distance between gv4 setae ca. 112 µ. Distance between gv1 and gv2 setae ca. 78 µ; distance between gv2 and gv3 setae ca. 57 µ; and distance between gv3 and gv4 setae ca. 50 µ.
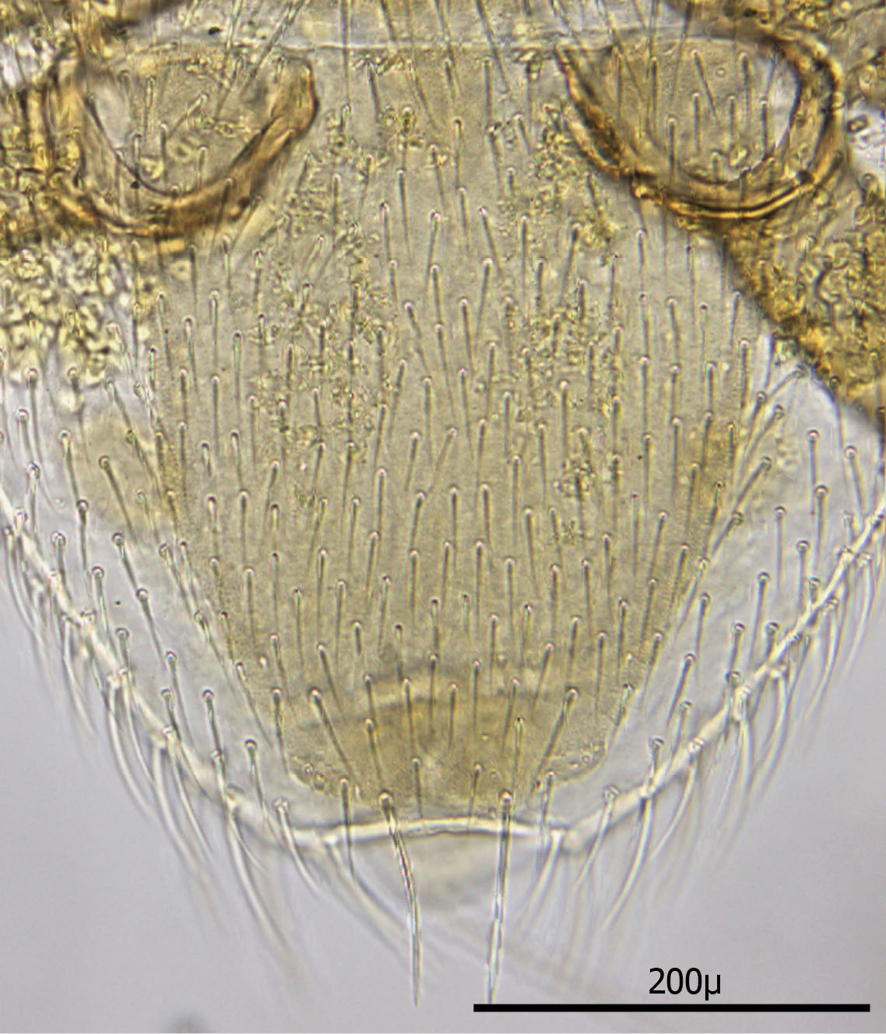
Anal shield (Fig. 21) not in juxtaposition with ventral shield; distance between ventral shield and anal shield ca. 317 µ. Anal shield somewhat diamond shaped and slightly wider than long with length/width ca. 0.81/0.86 µ; anterior margin acutely rounded and lateral margins more broadly rounded; length from anterior margin to postanal seta 99–100 µ; greatest width at level of anal orifice 122–139 µ; adanal setae situated at level of posterior 1/3 of anal orifice; distance between adanal setae 49–54 µ; adanal setae setaceous and 33–40 µ long; distance from anterior margin to adanal setae ca. 64 µ; distance from adanal setae to postanal seta 37–55 µ; postanal seta very long, 124–137 µ. Shorter setae of unarmed venter 38–52 µ long with longer setae 56–66 µ long.
Stigma of peritreme (Fig. 22) located lateral to level between coxae III and IV with no peritremala extension posteriorly; anterior extension of peritreme narrowing somewhat and reaching to level of mid coxae I; total length of peritreme ca. 384 µ.
Legs: The length and width of leg segments is given in table 4. All measurements were made ventrally. Length measurements were made mid-ventrally and widths were made at the widest point. The measurements are approximate because there is about a 5–10% variation in measurements from specimen to specimen.
The leg chaetotaxy for deutonymphs is shown in table 5. The chaetotaxy formula for each leg segment is as follows:
al — d/v — pl (anterior laterals — dorsals/ventrals — posterior laterals). The total for each leg segment is in parenthesis bolded.
Leg I: Coxae I greatest width ca. 79 µ and mid–ventral length ca. 100 µ. Coxae I setae setaceous; proximal setae longer (ca. 54 µ) and distal setae shorter (ca. 49 µ). Proximal ventral seta of trochanter slender setaceous ca. 40 µ long and distal seta ca. 48 µ long. Distal dorsal setae of femur setaceous but with one short (ca. 20 µ long) and the other somewhat longer (ca. 32 µ). Proximal–most dorsal seta of genu ca. 30 µ long and central setae ca. 42 µ long. Tarsus with setae all setaceous.
Leg II: Coxae II greatest width ca. 135 µ and mid–ventral length ca. 45 µ. Anterior seta setaceous and ca. 39 µ long; posterior seta setaceous and longer, ca. 54 µ. Ventral setae of trochanter setaceous but with proximal seta much longer (ca. 50 µ) than distal (ca. 35 µ). Distal dorsal setae of femur stout, setaceous and short (ca. 43 long µ). Proximal anterior dorsal seta of genu short setaceous (ca. 30 µ long), whereas posterior seta longer setaceous (ca. 52 µ long). Tarsus with pre–apical seta all setaceous.
Leg III: Coxae III greatest width ca. 121 µ and mid–ventral length ca. 61 µ. Both coxal setae setaceous; anterior seta shorter (ca. 35 µ long) than posterior seta (ca. 59 µ long). Posterior proximal setae of trochanter setaceous and long (ca. 81 µ long); posterior distal seta shorter, slender setaceous (ca. 27 µ long). Setae of femur III stout, long setaceous (ca. 81 µ long). Proximal 2 setae of genu setaceous and subequal in length (51–60 µ). Tarsus with pre–apical seta setaceous.
Leg IV: Coxae IV greatest width ca. 101 µ and mid–ventral length ca. 61 µ. Single seta setaceous and 40–46 µ long. Dorsal proximal setae of trochanter setaceous (ca. 56 µ long) and distal setae shorter (ca. 38 µ) long. Dorsal distal setae of femur long setaceous (ca. 103 µ); setae of genu IV setaceous and subequal in length (43–48 µ); tarsus IV with pre–apical setae setaceous.
Length/width (in µ) of leg segments for females.
| Segment | Leg I | Leg II | Leg III | Leg IV |
|---|---|---|---|---|
| Trochanter | 47/42 | 46/63 | 56/45 | 65/46 |
| Femur | 59/43 | 63/59 | 54/34 | 69/40 |
| Genu | 56/38 | 50/52 | 31/35 | 51/34 |
| Tibia | 60/35 | 38/45 | 32/41 | 47/35 |
| Tarsus | 100/30 | 85/31 | 98/26 | 228/39 |
Leg segmental chaetotaxy for females.
| Segment | Leg I | Leg II | Leg III | Leg IV |
|---|---|---|---|---|
| Trochanter | 1 — 1/3 — 1 (6) | 1 — 1/2 — 1 (5) | 1 — 1/3 — 1 (6) | 1 — 1/3 — 1 (6) |
| Femur | 2 — 4/4 — 1 (11) | 2 — 3/3 — 2 (10) | 2 — 2/2 — 1 (7) | 2 — 2/1 — 1 (6) |
| Genu | 2 — 6/3 — 2 (13) | 2 — 4/2 — 2 (10) | 2 — 4/1 — 1 (8) | 2 — 4/2 — 1 (9) |
| Tibia | 2 — 6/3 — 2 (13) | 1 — 4/3 — 2 (10) | 2 — 4/1 — 1 (8) | 2 — 4/3 — 1 (10) |
| Tarsus | 3 — 6/6 — 3 (18) | 3 — 4/4 — 3 (14) | 3 — 4/5 — 3 (15) | 3 — 4/5 — 3 (15) |
Length/width (in µ) of leg segments for Deutonymphs.
| Segment | Leg I | Leg II | Leg III | Leg IV |
| Trochanter | 67/58 | 67/87 | 70/90 | 103/94 |
| Femur | 117/61 | 108/98 | 92/72 | 147/67 |
| Genu | 111/57 | 54/88 | 56/68 | 92/63 |
| Tibia | 141/56 | 73/88 | 73/64 | 110/58 |
| Tarsus | 191/47 | 166/48 | 183/45 | 254/43 |
Leg segmental Chaetotaxy for Deutonymphs.
| Segment | Leg I | Leg II | Leg III | Leg IV |
| Trochanter | 1 — 1/3 — 1 (6) | 1 — 1/2 — 1 (5) | 1 — 1/2 — 2 (6) | 1 — 1/2 — 1 (5) |
| Femur | 1 — 3/4 — 1 (9) | 3 — 6/4 — 2 (15) | 1 — 3/1 — 1 (6) | 3 — 3/1 — 1 (8) |
| Genu | 2 — 6/3 — 2 (13) | 2 — 5/3 — 2 (12) | 2 — 4/2 — 1 (9) | 2 — 5/2 — 1 (10) |
| Tibia | 2 — 6/4 — 2 (14) | 1 — 4/3 — 2 (10) | 2 — 3/2 — 1 (8) | 2 — 4/2 — 2 (10) |
| Tarsus | 4 — 8/5 — 4 (21) | 3 — 5/4 — 3 (15) | 3 — 5/5 — 3 (16) | 3 — 3/5 — 3 (14) |
Line drawing of dny dorsum.
Line drawing of dny venter.

Anterior dorsum.
Posterior dorsum.
Gnathosoma.

Chelicerae.
Ventral shield.
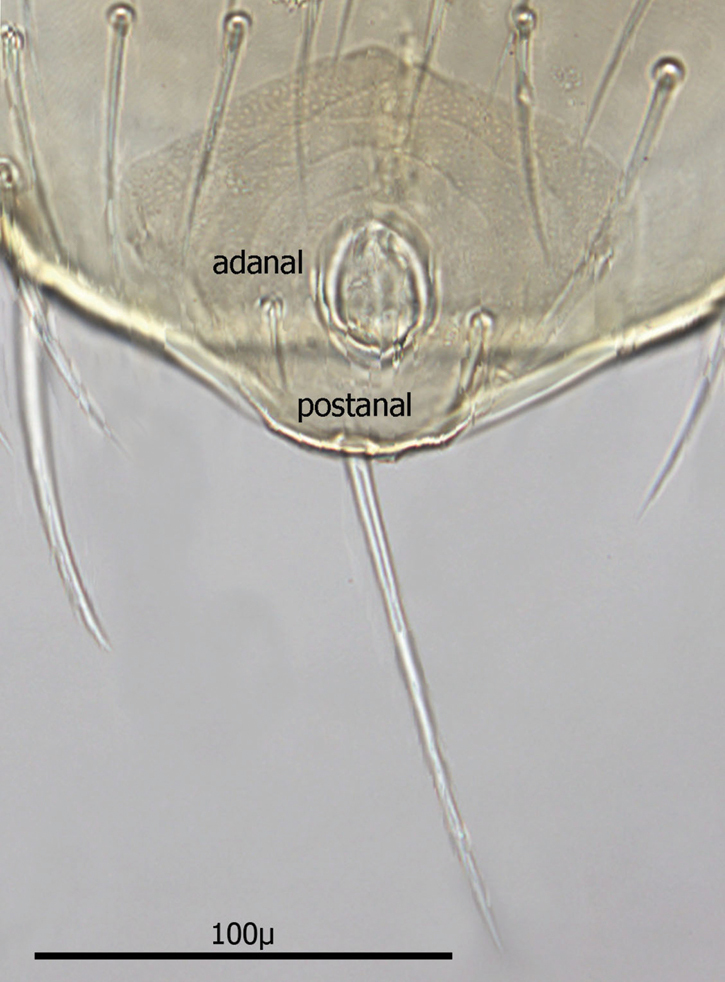
Anal shield.

Peritreme.
Haemogamasus alongipilis was frequently collected along with another mite species: Androlaelaps (Eubrachylaelaps) abrothrix
There were a total of 128 Haemogamasus alongipilis specimens mounted and identified. Of these, 33 (25.8%) were ♀s and 95 (74.2%) were deutonymphs. There were no male specimens found in the collections, likely because males are rare compared to females and deutonymphs. For the 3 Abrothrix host species there were 110 Haemogamasus alongipilis specimens mounted and identified. Of these 30 (27.3%) were ♀s and 80 (72.7%) were deutonymphs. Because Haemogamasus mites are primarily nest parasites and the deutonymphs are the migrating stage, the ratio (about 1:3) of females to deutonymphs in the collection is not surprising.
Table 6 below lists the Haemogamasus alongipilis collection records for each of the 7 host species collected in the 3 Argentina provinces. Most of the collections of Haemogamasus alongipilis were collected from Abrothrix rodents (84.4%), almost exclusively from Abrothrix longipilis (81.8%), with only 1 collection each from Abrothrix olivaceus and Abrothrix xanthorhinus. The 4 non–Abrothrix host species account for only 12 collections (15.6%). The 2 non–Abrothrix host species with 5 collections of Haemogamasus alongipilis each (Geoxus valdivianus & Loxdontomys micropus) are definitely valid records. The other 2 non–Abrothrix host species (Oligoryzomys longicaudatus & Dromiciops gliroides), with only 1 collection of Haemogamasus alongipilis each, are most likely incidental or accidental.
Table 7 lists the number and incident frequency of Haemogamasus alongipilis specimens collected from each of the 7 host species. For Abrothrix longipilis there were 135 host animals sampled with a total of 63 (46.7%) specimens of Haemogamasus alongipilis identified. Of the remaining 6 host species only 14 specimens of Haemogamasus alongipilis were identified. From the total of 224 host animals collected, 34.4% was found to host the 77 collections of Haemogamasus alongipilis.
Our study shows that the range of Haemogamasus alongipilis extends over more than three degrees of latitude (39°2' to 42°5'S) and a distance of 400 kilometers. All of the localities where the mice had this mite are located within the Valdivian rain forest phytogeographical region (
A majority of the described species of Haemogamasus have been reported only from Europe and Asia. Of the twelve species reported from the western hemisphere, only 2 (Haemogamasus ambulans & Haemogamasus pontiger) are known to occur worldwide, and in North America from most of Canada and the United States. The other 10 species have been reported exclusively from North America.
The new species described herein, Haemogamasus alongipilis, is the first Haemogamasus species described from South America. However, there is one literature report of Haemogamasus pontiger (Berlese) having been collected in South America. This report is from Argentina by Silvia
Collection records by host and province.
|
Province →Host species |
Chubut | Neuquén | Río Negro | Total |
| Abrothrix longipilis | 17 | 16 | 30 | 63 |
| Abrothrix olivaceus olivaceus | 0 | 1 | 0 | 1 |
| Abrothrix olivaceus xanthorhinus | 0 | 1 | 0 | 1 |
| Geoxus valdivianus | 0 | 3 | 2 | 5 |
| Loxdontomys micropus | 2 | 1 | 2 | 5 |
| Oligoryzomys longicaudatus | 0 | 0 | 1 | 1 |
| Dromiciops gliroides | 0 | 0 | 1 | 1 |
| Total | 19 | 22 | 36 | 77 |
Incident frequency of Haemogamasus alongipilis for each host species.
| Host species | Total Hosts Sampled | Haemogamasus alongipilis Collected | Frequency Rate |
| Abrothrix longipilis | 135 | 63 | 46.7% |
| Abrothrix olivaceus olivaceeus | 6 | 1 | 16.7% |
| Abrothrix olivaceus xanthorhinus | 16 | 1 | 6.3% |
| Geoxus valdivianus | 12 | 5 | 41.7% |
| Loxdontomys micropus | 18 | 5 | 27.8% |
| Oligoryzomys longicaudatus | 33 | 1 | 3.0% |
| Dromiciops gliroides | 4 | 1 | 25.0% |
| Total | 224 | 77 | 34.4% |
We thank Claudio Chehébar, Gustavo Iglesias, and Eduardo Ramilo of the Delegación Regional Patagonia of the Argentina National Park Service, for approving our projects (APN No. 696 and 889) and providing transport permits. Additional acknowledgments are extended to Julie Woodruff, Museum of Vertebrate Zoology, University of California, Berkeley, CA and Michael Mares and Janet Braun, Oklahoma Museum of Natural History, University of Oklahoma, Norman, OK for transporting and shipping specimens. The samples were exported to Brigham Young University by permit of the Dirección de Fauna Silvestre, Buenos Aires.