





(C) 2010 Oscar A. Alcober. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Herrerasauridae comprises a basal clade of dinosaurs best known from the Upper Triassic of Argentina and Brazil, which have yielded remains of Herrerasaurus ischigualastensis and Staurikosaurus pricei, respectively. Systematic opinion regarding the position of Herrerasauridae at the base of Dinosauria has varied. Here we describe a new herrerasaurid, Sanjuansaurus gordilloi gen. n., sp. n., based on a partial skeleton from Carnian-age strata of the the Upper Triassic Ischigualasto Formation of northwestern Argentina. The new taxon is diagnosed by numerous features, including long, band-shaped and posterolaterally oriented transverse process on the posterior cervical vertebrae; neural spines of the sixth to eighth dorsal vertebrae, at least, bearing acute anterior and posterior processes; scapula and coracoid with everted lateral margins of the glenoid; and short pubis (63% of the femoral length). Phylogenetic analysis placed Sanjuansaurus within a monophyletic Herrerasauridae, at the base of Theropoda and including Herrerasaurus and Staurikosaurus. The presence of Sanjuansaurus at the base of the Ischigualasto Formation, along with other dinosaurs such as Herrerasaurus, Eoraptor, Panphagia, and Chromogisaurus suggests that saurischian dinosaurs in southwestern Pangea were already widely diversified by the late Carnian rather than increasing in diversity across the Carnian-Norian boundary.
Dinosauria, Saurischia, Herrerasauridae, Carnian, Ischigualasto
Herrerasauridae (
The close relationship between its two best known genera, Herrerasaurus and Staurikosaurus is well supported by a suite of synapomorphies (
Here we describe a new herrerasaurid from strata near the basal contact of the Ischigualasto Formation. The new taxon is based on an associated, partially articulated skeleton recovered from Ischigualasto Provincial Park in 1994.
Geological and palaeontological settingsThe holotype of the new taxon (PVSJ 605) was found in
1994 during fieldwork carried out by the Instituto y Museo de Ciencias
Naturales of the Universidad Nacional de San Juan. The Ischigualasto
Formation crops out in northwestern Argentina and forms part of the
Ischigualasto-Villa Unión Basin (Figure 1).
It reaches up to 700 m in thickness and comprises fluvial channel
sandstones with sandstones and mudstones deposited on a well-drained
floodplain. Interlayered volcanic ashes 20 m above the base of the
formation provide chronostratigraphic control and have yielded an age of
231.4 Ma, placing them in the Carnian stage (
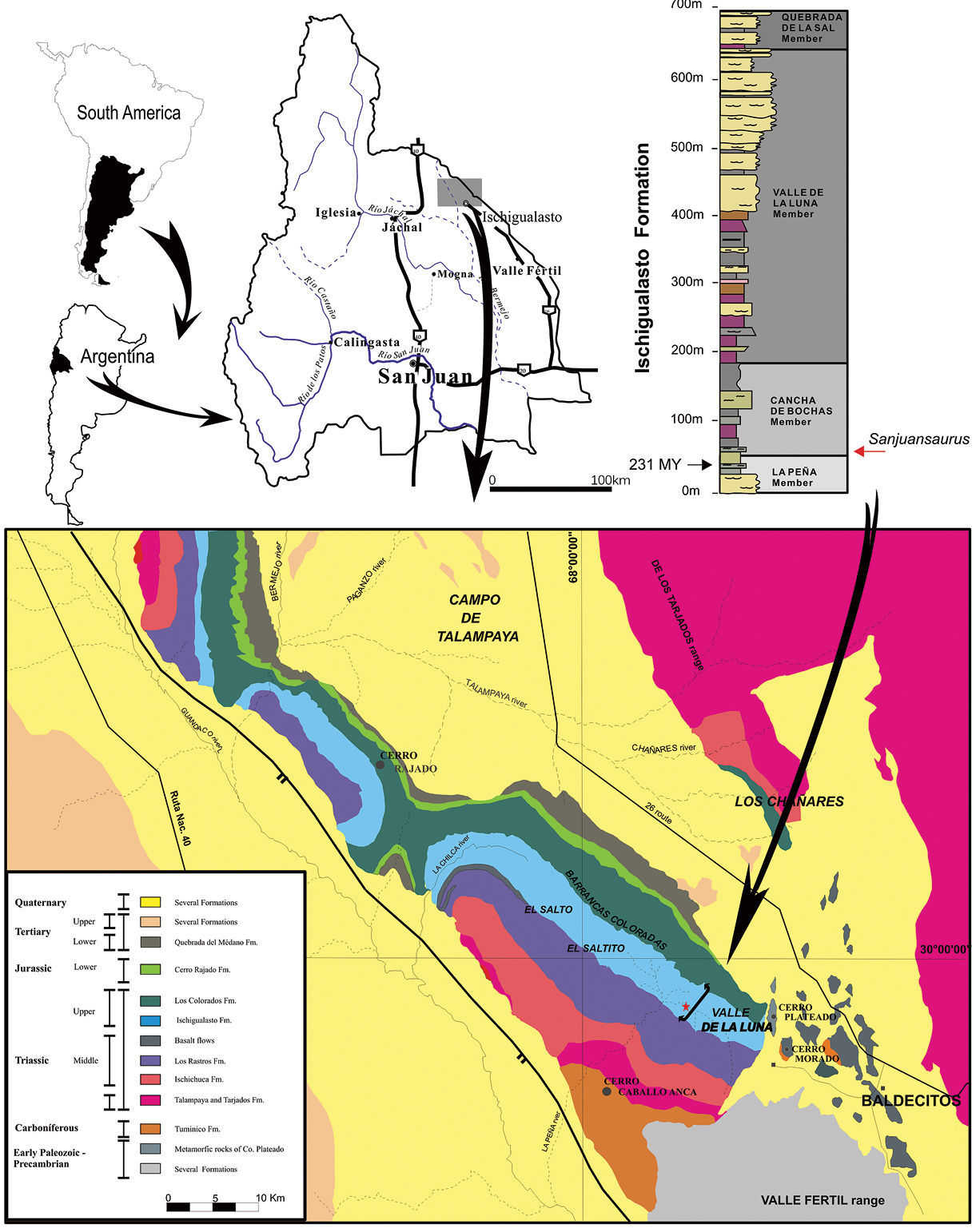
Geological map of the Ischigualasto – Villa Unión Basin (northwestern Argentina) and section of the Ischigualasto Formation at the type locality. The red star indicates the site of the holotype of Sanjuansaurus gordilloi (PVSJ 605), near the base of the Ischigualasto Formation.
The Ischigualasto Formation is divided into four members (
The new fossil was excavated at the La Gallinita
locality, which is located in the lowest levels of the Cancha de Bochas
Member. Itwas found 40 m above the base of the formation. Dinosaurs,
including several specimens of Herrerasaurus ischigualastensis and Eoraptor lunensis, the holotype of Panphagia protos, and other as yet undescribed species (
The reddish brown bones were covered by a coarse hematite crust, and the entire specimen was embedded in a grey-green, fine-grained sandstone matrix. The overall preservation of the specimen is good. All the bones are three-dimensionally preserved, and most are complete with the exception of the femora, which are partially distorted and lack fine details. The incompleteness of the skeleton is attributable to pre-burial processes, although the third, fourth and anterior part of the fifth dorsal vertebra were lost in the course of preparation. The specimen was prepared using a pneumatic air scribe and pin vice.
TerminologyWe employ traditional, or “Romerian, ” anatomical and directional terms rather than their veterinarian alternatives (
We used the stem-based phylogenetic definition for Herrerasauridae proposed by Sereno (
In order to asses the phylogenetic position of the new taxonamong basal Dinosauria, we added it (Table 1) and the recently described basal sauropodomorph Panphagia (
Character state scores for Sanjuansaurus gordilloi (PVSJ 605). Data lines inserted into the data matrix of Langer and Benton [33] with the addition of Martinez and Alcober [30]
| Sanjuansaurus | 0???? ?00?? ????? ????? ??000 00?11 11111 1?110 00111 1???? ????? ????? ??1?? ???2? ?1??0 010?? ?0001 00000 0000? ??? |
This published work and the nomenclatural acts it contains have been registered in ZooBank, the proposed online registration system for the ICZN. The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix “http://zoobank.org/”. The LSID for this publication is: urn:lsid:zoobank.org:pub:FB2AE660-C3EE-4348-BF9F-F4311C47E853
Institutional abbreviations:PVSJ Instituto y Museo de Ciencias Naturales, San Juan 5400, Argentina.
Results Systematic PaleontologySystematic hierarchy
Dinosauria Owen, 1842
Saurischia Seeley, 1887
Herrerasauridae Benedetto, 1973
urn:lsid:zoobank.org:act:DC75ADA0-0C6B-41D5-8E29-CD76725FD704
Sanjuan, in reference to San Juan Province, Argentina; saurus, lizard (Latin).
Sanjuansaurus gordilloi
urn:lsid:zoobank.org:act:84F081D6-4E0A-414E-83A3-D994263C1005
gordilloi, in honor of Raul Gordillo, head fossil preparator and artist in the laboratory of the San Juan Museum and team member during many years of excavation.
PVSJ 605, an incomplete skeleton including left maxilla, partial axial column, from the axis to the twelfth caudal vertebra, lacking the third, fourth, and anterior half of the fifth dorsal vertebrae, both scapulae, left ulna, ungual phalanx of left? digit III, preacetabular portion of the left ilium, proximal end of left and complete right pubis, both femora and tibiae, right fibula, right astragalus and calcaneum, and left metatarsal II.
The specimen was found in the Cancha de Bochas Member (
40 m above the base of the Ischigualasto Formation, Late Triassic, Carnian (ca. 231.4 Ma), Ischigualasto–Villa Unión Basin (
Diagnosed by the following autapomorphies: shelf-like, posterolaterally directed transverse processes on the posterior cervical vertebrae; neural spines of the sixth to eighth dorsal vertebrae, at least, bearing acute anterior and posterior processes; everted lateral margins of the glenoid; short pubis (63% of the femoral length); and pronounced, rugose scar on the medial surface of the femur at the level of the fourth trochanter.
These features distinguish Sanjuansaurus gordilloi from the previously described herrerasaurids, such as Herrerasaurus ischigualastensis and Staurikosaurus pricei, as well as other basal saurischians from Ischigualasto Formation, such as Eoraptor lunensis (
Although some bones of Sanjuansaurus gordilloi were disarticulated, the proximity of all elements, found in an area of only one square meter, their complementary size, and the absence of any duplicated elements suggest that they represent a single individual (Figure 2). In size and general proportions (Table 2, 3), the new specimen (Figure 3A) is comparable to a medium-sized Herrerasaurus (Figure 3B), and slightly larger than the only known specimen of Staurikosaurus (Figure 3C).
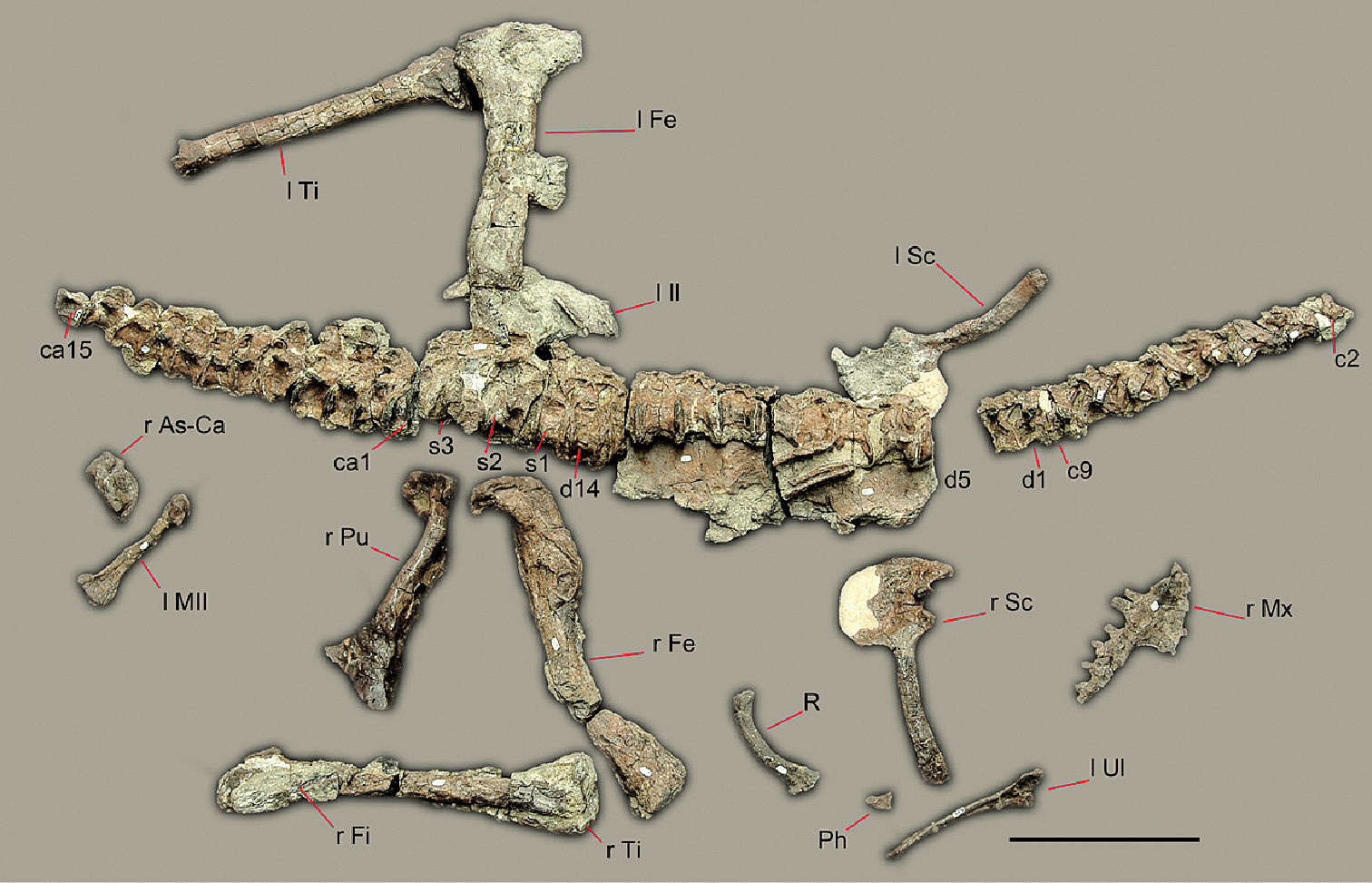
Preserved bones of Sanjuansaurus gordilloi (PVSJ 605), positioned as they were found in the field. Abbreviations: ca1–15, caudal vertebrae 1–15; c2–9, cervical vertebrae 2–9; d1–14, dorsal vertebrae 1–14; l Fe, left femur; l il, left ilium; l MII, left metatarsal II; l Sc, left scapula and coracoid; l Ti, left tibia; l Ul, left ulna; Ph, manual phalanx; R, rib; r As-Ca, right astragalus and calcaneum; r Fe, right femur; r Fi, right fibula; r Fe, right femur; r Mx, right maxilla; r Pu, right pubis; r Sc, right scapula and coracoid; r Ti, right tibia; s1–3, sacral vertebrae 1–3. Scale bars equals 20 cm.
Cranium. The left maxilla is the only cranial bone preserved (Figure 4A). It exhibits the anteroposteriorly elongated dorsal process and transversely narrow antorbital fossa as in Herrerasaurus (Figure 4B, C) but unlike the wide fossa of Eoraptor (
Dimensions (mm) of preserved vertebrae (Figures 2, 3, 5) from the holotypic specimen of Sanjuansaurus gordilloi (PVSJ 605).
| Vertebra | Centrum_length1 | Vertebra | Centrum_length1 |
|---|---|---|---|
| C2 | 37.0 | D13 | 36.0 |
| C3 | 42.5 | D14 | 34.5 |
| C4 | 44.0 | S1 | 38.0 |
| C5 | 46.1 | S2 | 48.0 |
| C6 | 47.5 | S3 | 46.2 |
| C7 | 45.6 | CA1 | 35.8 |
| C8 | 38.5 | CA2 | 36.8 |
| C9 | 38.0 | CA3 | 35.0 |
| D1 | 33.6 | CA4 | 37.0 |
| D2 | 28.3 | CA5 | 33.2 |
| D6 | 36.8 | CA6 | 32.3 |
| D7 | 38.0 | CA7 | 38.5 |
| D8 | 38.8 | CA8 | 38.5 |
| D9 | 36.6 | CA9 | 37.8 |
| D10 | 37.7 | CA10 | 37.8 |
| D11 | 37.5 | CA11 | 36.5 |
| D12 | 38.0 |
Abbreviations: C cervical; CA caudal; D dorsal; S sacral. 1Measured along ventral edge excluding anterior convexity of centrum when present.
Dimensions (mm) of girdle and limb bones (Figures 2, 3, 6-8) of the holotypic specimen of Sanjuansaurus gordilloi (PVSJ 605).
| Bone | Measurement | Length |
|---|---|---|
| Scapulocoracoid | Coracoid, maximum height | 63.0 |
| Posterior process (glenoid to tip of process) | 131.0 | |
| Scapular length | 185.0 | |
| Scapular blade, minimum width | 18.5 | |
| Scapular blade, distal width | 26.8 | |
| Height (glenoid to acromion) | 89.5 | |
| Ulna | Maximum length | 178.2 |
| Anteroposterior shaft diameter (mid-shaft) | 10.1 | |
| Pubis | Length (acetabulum to foot) | 260.0 |
| Iliac peduncle length | 32.0 | |
| Ischial peduncle length | 29.0 | |
| Pubic foot, maximum length | 100.2 | |
| Pubic foot, maximum width | 21.5 | |
| Femur | Maximum length | 395.0 |
| Maximun distal width | 90.5 | |
| Tibia | Length | 260.0 |
| Maximun proximal dimension | 91.0 | |
| Proximal width | 41.5 | |
| Distal transverse width | 40.0 | |
| Distal anteroposterior width | 43.5 | |
| Astragalus-Calcaneum | Proximodistal depth | 36.0 |
| Transverse width | 83.0 | |
| Medial anteroposterior width | 37.5 | |
| Metatarsal II | Length | 147.5 |
| Maximun proximal dimension | 40.0 | |
| Distal transverse width | 32.0 |
Axial skeleton. The cervical vertebrae are preserved from the incomplete atlas to the last vertebra of the series (Figure 5A, B). Sanjuansaurus
has nine cervical vertebrae. The last cervical vertebra differs from
the first dorsal vertebra in the presence of a ventral keel and being
nearly 30% longer, with the paraphophysis being more anteriorly located
and the capitulum of the associated rib being more slender (Figure 2, 3, 5A). Although ten cervical vertebrae have been reported in Herrerasaurus (
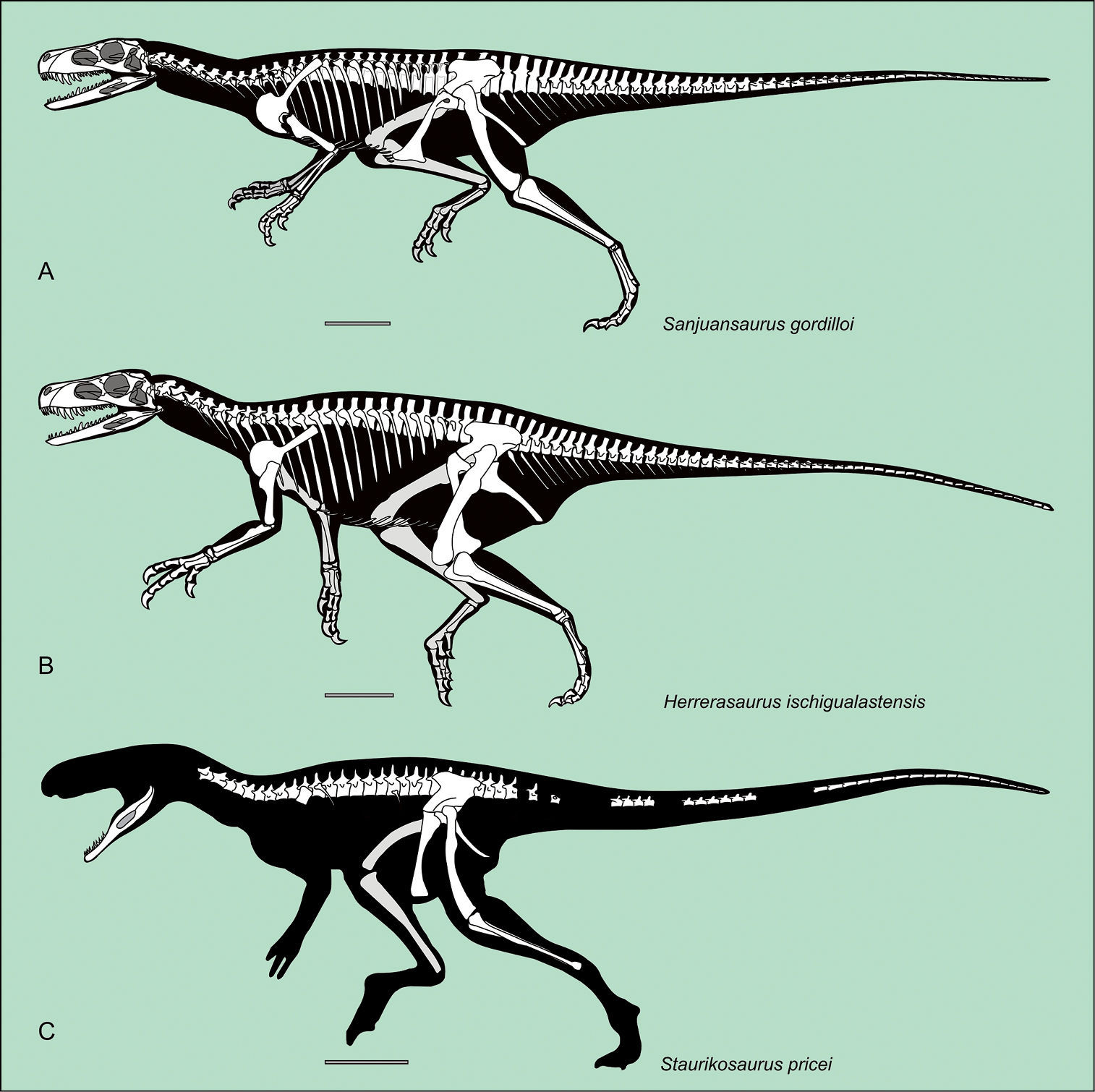
Silhouette reconstruction of the skeletons of the best known herrerasaurids.Sanjuansaurus gordilloi (PVSJ 605) (A) Herrerasaurus ischigualastensis (B) Staurikosaurus pricei (C) Scale bars equals 20 cm. A: missing bones of Sanjuansaurus modified from

Maxilla of Sanjuansaurus gordilloi (PVSJ 605). Left maxilla of Sanjuansaurus in lateral view (A) Skull of Herrerasaurus (PVSJ 407) in left lateral view (B) Left maxilla of Herrerasaurus (PVSJ 053) in lateral view (C) Abbreviations: af, antorbital fossa; dp, dorsal process; ip, internarial process; J, jugal; jp, jugal process; M, maxilla; of, oval fenestra; pf, promaxillary fenestra; PO, postorbital. Scale bars equals 5 cm.
The atlas is represented only by its centrum, the odontoid (Figure 5A),
which is completely co-ossified with the axial centrum. It is
subcircular in dorsal view with a pointed median projection as in Herrerasaurus (
The axial intercentrum (Figure 5A) is fused to the axial centrum. It is much broader than the anterior end of the axial centrum as in Herrerasaurus. In lateral view it is subquadrangular rather than triangular as in Herrerasaurus (
The axial centrum (Figure 5A) is twice as long as it is deep and bears a pronounced ventral keel as in Herrerasaurus (
The postaxial cervical centra are spool-shaped, amphicoelous, and have pronounced ventral keels. In Herrerasaurus,
by contrast, the lateral and ventral sides of the centrum are less
concave, and the ventral keel diminishes progressively in more distal
vertebrae (
The dorsal vertebrae are articulated. Only the third,
fourth and anterior half of the fifth vertebra are lacking and were
accidentally lost during preparation (Figure 5A–C). Thus we can say with confidence that there are 14 dorsal vertebrae as in Herrerasaurus (
The sacrum (Figure 5C, D) of Sanjuansaurus comprises three vertebrae. The first is a dorsosacral whereas the second and third represent the primordial sacral pair as in Herrerasaurus (
The first sacral vertebra is 10% longer than the last
dorsal vertebra, and the position of the infradiapophyseal laminae and
the transverse processes are somewhat different. The posterior
centrodiapophyseal lamina is displaced anteriorly toward the anterior
centrodiapophyseal lamina, so that ventral to the transverse process
they both extend nearly vertically (Figure 5C, D).
The transverse process is not a single flat process as on the posterior
dorsal vertebrae but rather is composed of two laminae, one horizontal
and the other one vertical, which join to form an inverted L-shape
near the contact with the ilium. This configuration is similar to that
shown on dorsal vertebra 15 in Herrerasaurus (
The second sacral vertebra is 25% longer than the first and is the longest in the sacrum (Figure 5C, D). This is true for one individual of Herrerasaurus (PVSJ 373), whereas in another (PVL 2566) the third sacral vertebra is longest (
The third sacral vertebra, the most robust of the sacrum, is dorsolaterally flattened and transversally expanded as in Herrerasaurus (
The sacral ribs of the second and third sacral vertebrae
have broad distal attachment surfaces that are continuous and, in
lateral view (Figure 5D)
form a C-shape that opens dorsally. Large subcircular openings are
present between the articular ends of the sacral ribs and the centra as
in Herrerasaurus (
The first 15 caudal vertebrae of PVSJ 605 are preservedin
articulation. The posterior end of the fifteenth vertebra and all the
hemal arches are lacking (Figure 5E). The preserved centra are strongly constricted, or spool-shaped, more so than in Herrerasaurus (
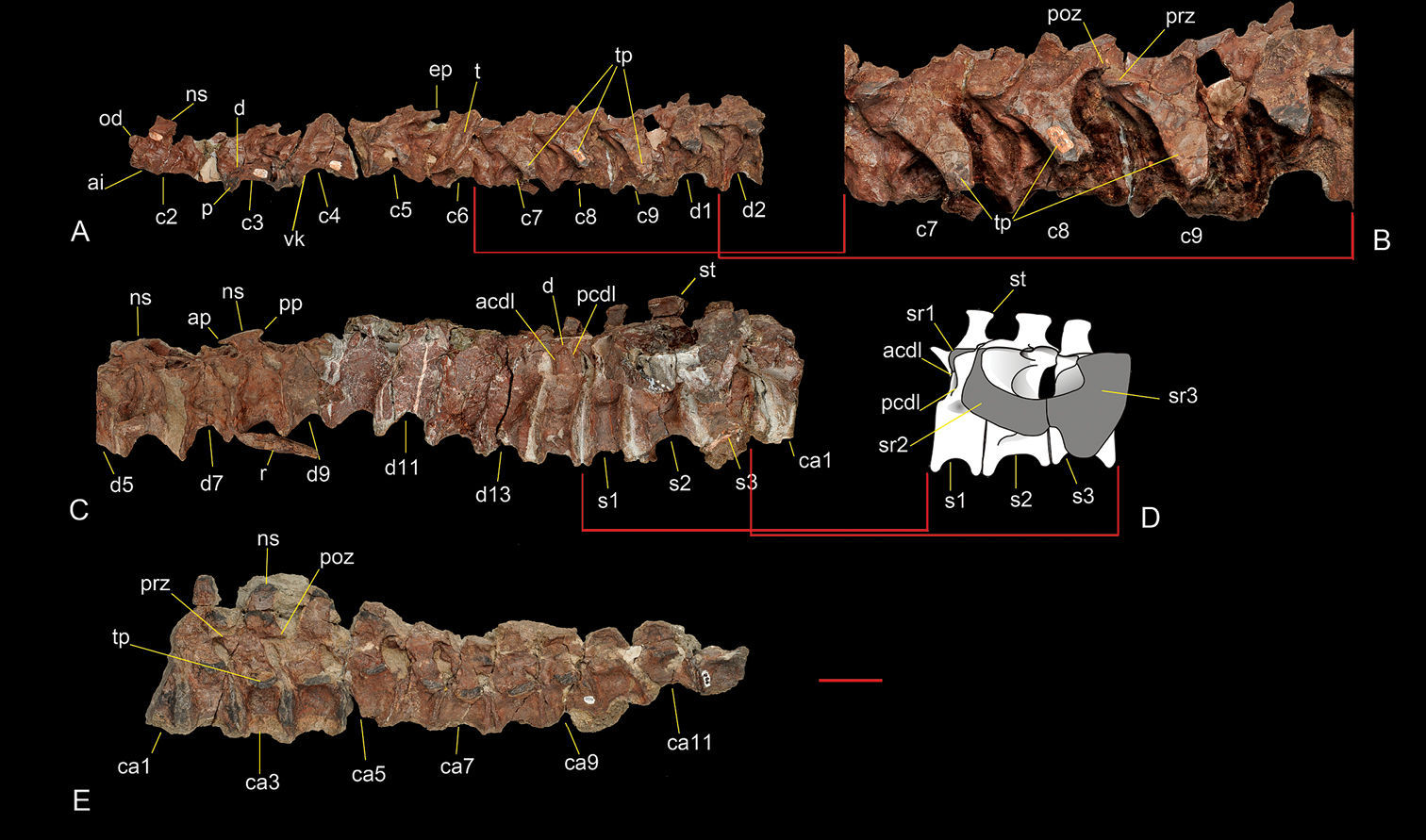
Axial skeleton of Sanjuansaurus gordilloi (PVSJ 605). Nine anterior cervical and two anterior dorsal vertebrae in left lateral view (A). Cervical vertebrae 7, 8 and 9 in left lateral view (B). Dorsal vertebra 1 to caudal vertebra 1 in left lateral view (C). Reconstruction of the sacrum in left lateral view (D). Caudal vertebra 1 to 12 in left lateral view (E). Abbreviations: acdl, anterior centrodiapophyseal lamina; ai, axis intercentrum;ap, anterior process of dorsal neural spine; ca1–12, caudal vertebrae 1 to 12; c2–9, cervical vertebrae 2 to 9; d, diapophysis; d1–13, dorsal vertebrae 1 to 13; ep, epipophysis; ns, neural spine; od, odontoides; p, parapophysis; pcdl, posterior centrodiapophyseal lamina; prz, prezygapophysis; poz, postzygapophysis; pp, posterior process of dorsal neural spine; s1–3, sacral vertebrae 1 to 3; sr1–3, sacral ribs 1 to 3; st, spine table; t, tooth; tp, transverse process; vk, ventral keel. Scale bar equals 5 cm.
Appendicular Skeleton. Each scapula is firmly
fused to its respective coracoid. The anterior margin is broken away on
both scapulae, although its curved margin can be restored. The acromial
process diverges from the blade at an angle slightly greater than 90°,
as in Herrerasaurus (
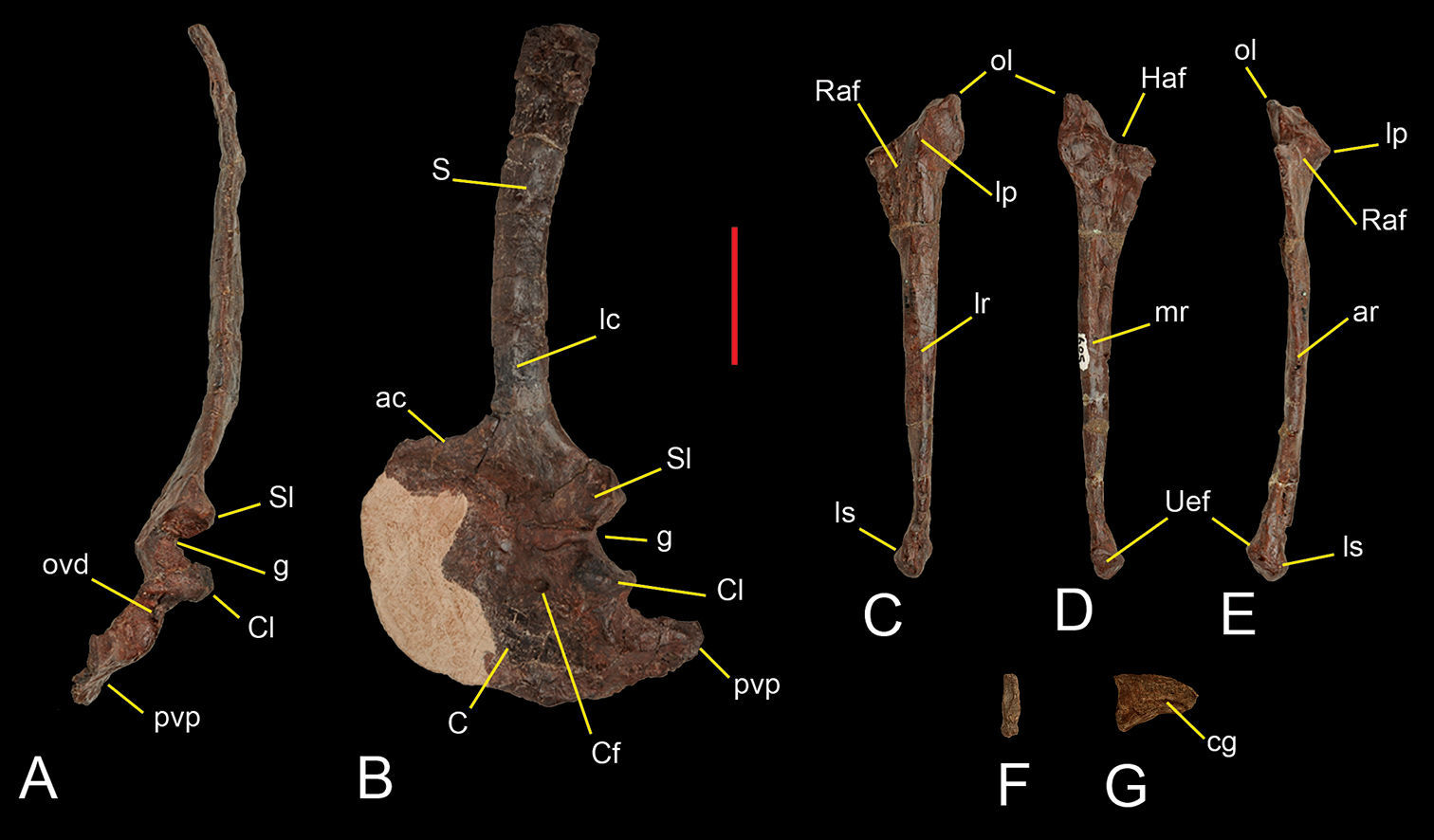
Preserved shoulder blade and forelimb bones of Sanjuansaurus gordilloi (PVSJ 605). Left scapula and coracoid inposterior(A), and lateral (B) views. Left ulna in lateral (C), medial (D), and anterior views (E). Ungual phalanx of digit III in proximal (F) and lateral view (G). Abbreviations: ac, acromion; ar, anterior ridge; C, coracoid; cg, collateral groove; Cf, coracoid foramen; Cl, coracoid lip; g, glenoid surface; Haf, humeral articular facet; lc, lateral crest; lp, lateral prominence; lr, lateral ridge; ls, ligament scars; lp, lateral prominence; ol, olecranon ; ovd, oval depression; pvp, posteroventral process; Raf, radius articular facet; S, scapula; Sl, scapular lip; Uaf, ulnare articular facet. Scale bar equals 5 cm.
The semicircular, plate-shaped coracoid (Figure 6A, B) is broader anteroposteriorly than dorsoventrally as in Herrerasaurus (
The ulna is more gracile than that of Herrerasaurus (
Of the manus only one manual ungual (Figure 6F, G) is preserved. The ungual has been crushed transversely. Its curvature and transversely narrow proportions (Figure 6F) identify it as pertaining the the manus rather than the pes. Its small size, short and deep proportions, and ventral position of the attachment groove suggest that it is probably the fourth (terminal) phalanx of the third digit. The ungual has a well developed flexor tubercle more deeply grooved on the left side (Figure 6G). Although we considered that it is the smaller ungual of the hand, it is still small compared to individuals of Herrerasaurus (PVSJ 373) smaller in overall size than Sanjuansaurus.
A fragment of the left ilium comprises the distal portion of the pubic peduncle (Figure 7D).
It is fused with the proximal end of the pubis, and preserved in
articulation with the left femur (although the latter is dorsally
rotated from its natural position). Medially it is also fused to a
distal fragment of the second sacral rib. The pubic peduncle is stout,
anteroventrally directed and forms the anterior border of a wide
perforate acetabulum as in Herrerasaurus (
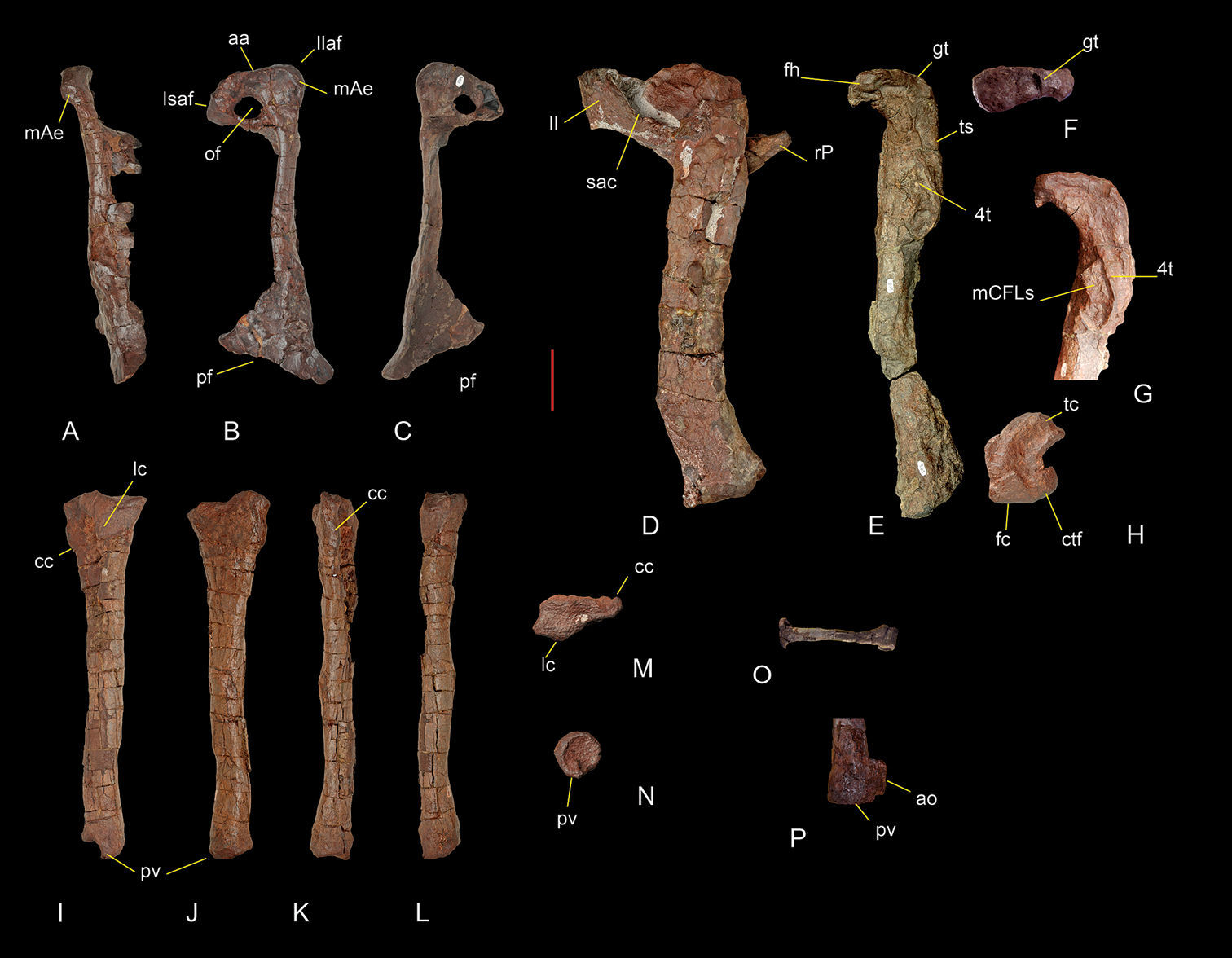
Pelvic and hind limb bones of Sanjuansaurus gordilloi (PVSJ 605). Right pubis in anterodorsal (A), lateral (B), and medial views (C). Left femur articulated with the preacetabular portion of the ilium and proximal end of the pubis in lateral view (D). Right femur in medial view (E). Right femur head in proximal view (F). Proximal end of the right femur in anteromedial view (G). Distal end of the left femur in distal view (H). Left tibia in lateral (I), medial (J), anterior (K), posterior (L), proximal (M), and distal views (N). Left metatarsal II in anterior view (O). Distal end of the right tibia in posterior view (P). Abbreviations: aa, acetabular area; ao, abnormal outgrowth in posteroventral process; cc, cnemial crest; ctf, crista tibiofibularis; 4t, fourth trochanter; fc, fibular condyle; fh, femoral head; gt, greater trochanter; Isaf, ischium articular facet; Il, illium; Isaf, ischium articular facet; lc, lateral condyle; mae, ambiens muscle eminence; mCFLs, scars for attachment of muscle caudifemoralis longus; of, obturator foramen; pf, pubic foot; pv, posteroventral process; rP, right pubis; sac, supraacetabular crest; tc, tibial condyle; ts, trochanteric shelf. Scale bar equals 5 cm.
The pubis is relatively short, its proximodistal length comprising only 63% the length of the femur (Figure 7A–C). By contrast, in Herrerasaurus and Staurikosaurus,
the length of the pubis equals 91% (PVL 2566) and 70% the length of
the femur, respectively. The two proximal articular surfaces are set at
an angle of 130°. The first faces posteriorly and articulates with the
ischium whereas the second faces posterodorsally and includes an
acetabular section and an articular facet for the ilium. A marked
prominence, more distinct than that seen in Herrerasaurus (
Both femora are poorly preserved (Figure 7D–H). The left femur is complete and articulated with the ilium, although rotated dorsally from its natural articulation (Figure 7D). Only the proximal and distal ends of the right femur are preserved (Figure 7E). The femur is sigmoid in lateral and anterior views (Figure 7D, E).
The anteromedially projecting head lies at an angle of approximately
65° to the transverse axis of the distal end. This is slightly more
divergent than in Herrerasaurus (55°;
The left tibia of Sanjuansaurus is complete and well preserved (Figure 7I–N).
The right tibia is preserved in articulation with the fibula. The
distal ends of these bones as well as the right astragalus and calcaneum
exhibit some features that appear to be abnormalities rather than
artifacts of postmortem compression or crushing. The tibia is slightly
shorter than the femur. The tibiofemoral ratio is 0.89, which lies
within the range recorded for Herrerasaurus (0.87–0.91) (
The relatively slender fibula is subequal to the tibia in length and has transversally flattened proximal and distal ends, the former twice the anteroposterior width at the mid shaft. The shaft is slightly bowed anterolaterally, and has a subtriangular cross-section at mid shaft. Poor surface preservation and breakage of the distal end obscure further details.
The astragalus and calcaneum of Sanjuansaurus are fused as in some dinosauromophs (Dromomeron romeri;
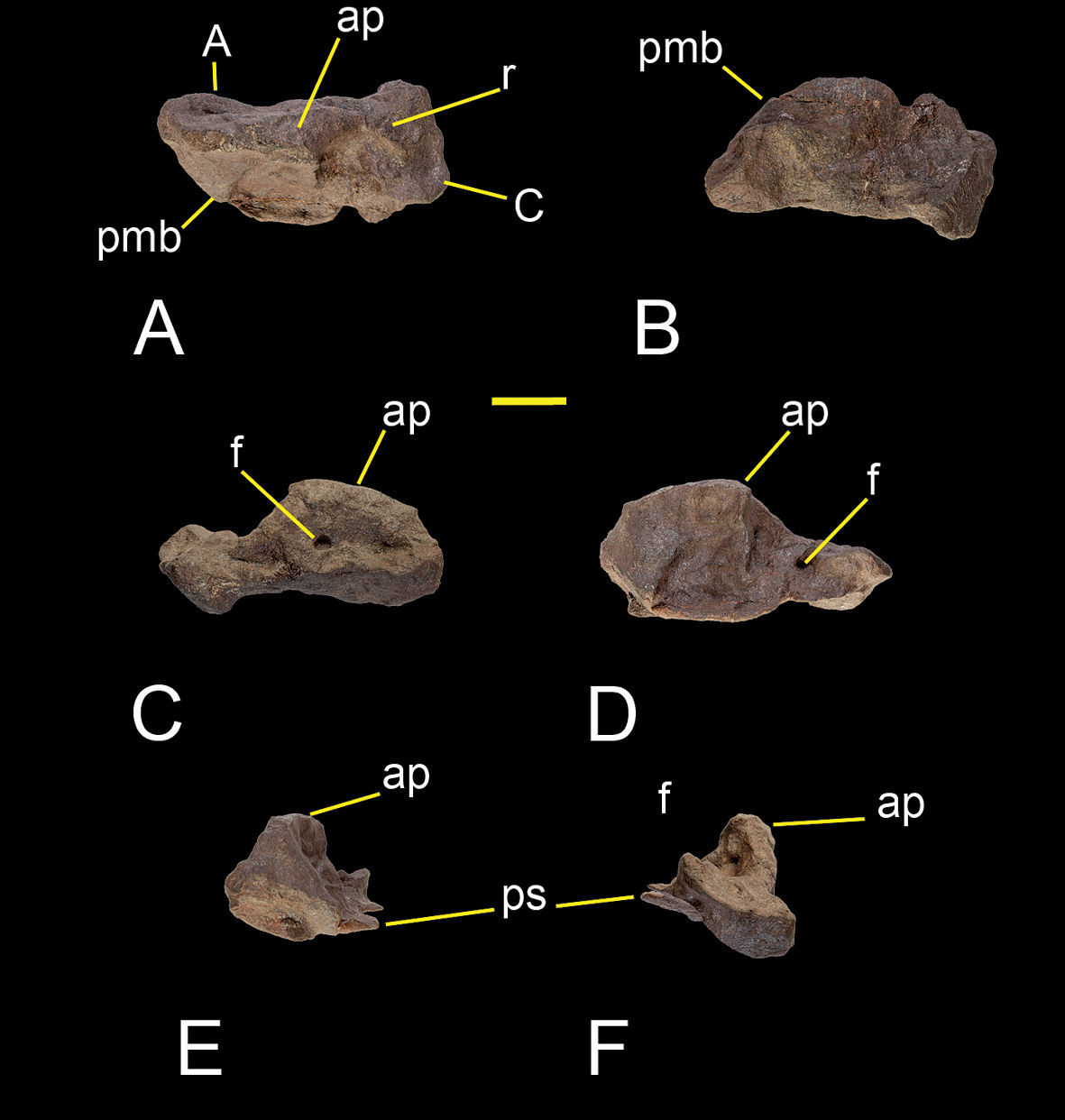
Right astragalus and calcaneum of Sanjuansaurus gordilloi (PVSJ 605). Right astragalus and calcaneum in proximal (A), distal (B), anterior (C), posterior (D), lateral (E), and medial views (F). Abbreviations: A, astragalus; ap, ascending process; C, calcaneum; f, foramen; pmb, posteromedial border; ps, posterior shelf; r, ridge. Scale bar equals 2 cm.
A complete left second metatarsal is the only pedal bone preserved. It is straight in dorsal view (Figure 7O). The proximal end is transversely flattened with the long axis directed anterolaterally as in Herrerasaurus (
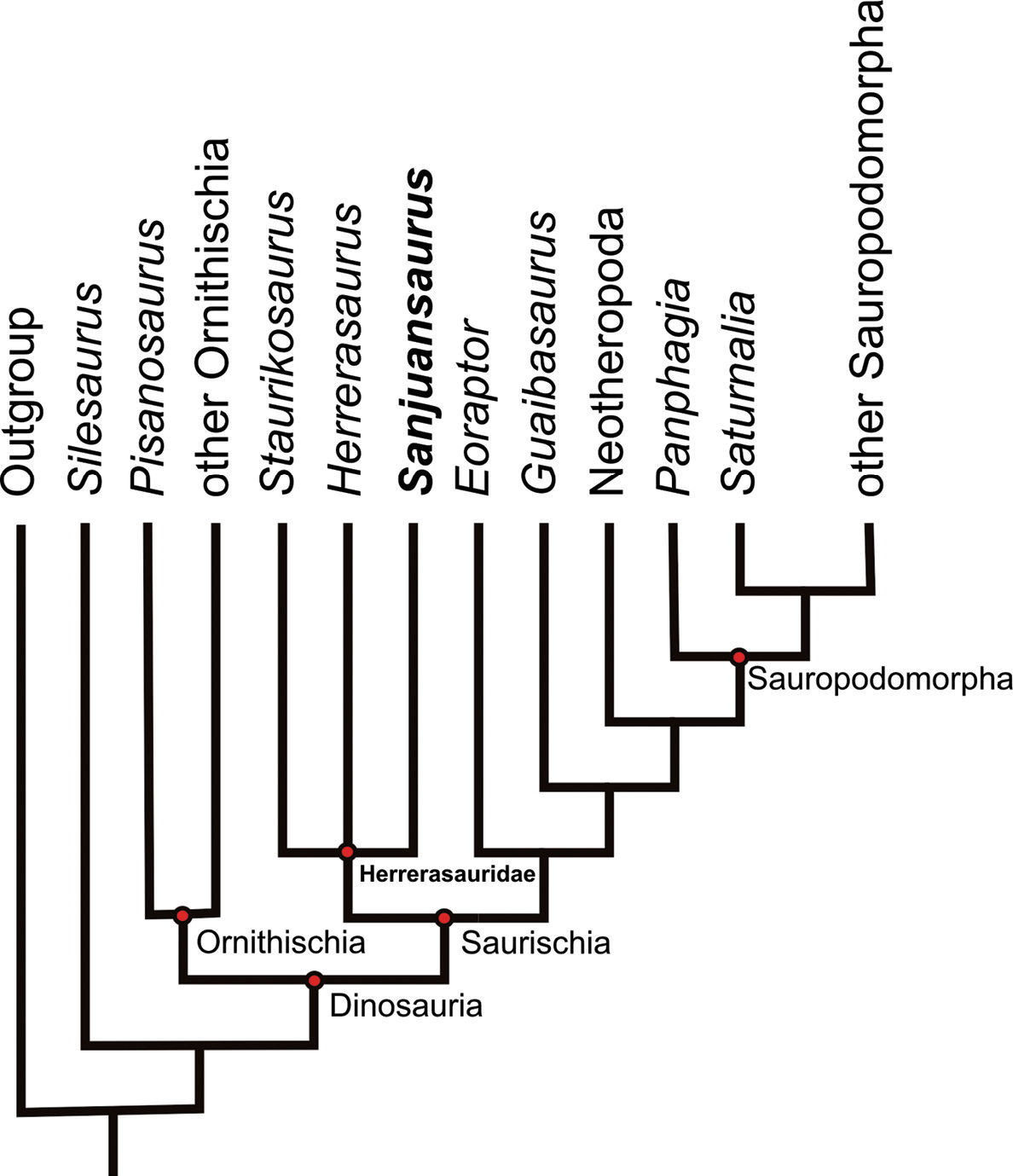
Single most parsimonious tree (MPT) resulting from the present parsimony analysis (tree length 190 steps; consistency index 0.553, retention index 0.593).
Sanjuansaurus gordilloi exhibits several features that allow its distinction from all other known basal dinosaurs:
1. Shelf-like, posterolaterally directed transverse processes in posterior cervical vertebrae. In Sanjuansaurus
from the fifth cervical vertebra back, the transverse processes
increase in anteroposterior width and transverse length, and project
posteroventrolaterally (Figure 5B), an unusual shape among basal dinosaurs. In other basal dinosaurs the transverse process are shorter or triangular (e.g, , Herrerasaurus, Staurikosaurus, Eoraptor, Tawa, Adeopapposaurus;
2. Neural spines of the sixth to eighth dorsal vertebrae, at least, bearing acute anterior and posterior processes. Sanjuansaurus has distinctive pointed processes extending anteriorly and posteriorly from the apex of the preserved anterior dorsal spines (Figure 5C, D). This feature is unique among known dinosaurs.
3. Coracoid with long posteroventral process. In Sanjuansaurus the hook-shaped posteroventral process of the coracoid is long and pointed (Figure 6B), similar to that in ornithomimids such as Gallimimus (
4. Everted lateral margins of the glenoid. In Sanjuansaurus the rim of the glenoid protrudes laterally in a conspicuous manner (Figure 6A).
In the glenoid portion of the bones the margins represent 50% of the
thickness of the scapula and the 60% the thickness of the coracoid. In
other basal dinosaurs the glenoid portions of the scapula and coracoid
are the thicker portions of the respective bones, but they lack the
everted lateral margins (e.g, Herrerasaurus, Eoraptor, Panphagia, Tawa, Guaibasaurus, Saturnalia;
5. Short pubis (63% of the femoral length). In Sanjuansaurus the pubis is very short (63% of the femoral length), shorter than that present in other basal surischians (e.g, Herrerasaurus (91%), Staurikosaurus (70%), Tawa (90%), Eoraptor (80%).
6. Pronounced rugose scar on the medial surface of the femur at the level of the fourth trochanter. In Sanjuansaurus the scar presumably for the insertion of M. caudifemoralis longus is very large, pronounced and rugose protuberance (Figure 7G). In other basal dinosaurs this scar is relatively small and smooth (e.g., Herrerasaurus, Eoraptor, Panphagia, Tawa, Guaibasaurus, Saturnalia;
These autapomorphies allow us to distinguish Sanjuansaurus gordilloi from other known basal dinosaurs. Furthermore, although Sanjuansaurus and Herrerasaurus
are similar, the new taxon can be further distinguished from the
latter form by three other characters: (1)The scapular blade of Sanjuansaurus is straplike in lateral view (Figure 6A, B), similar in shape than that present in Herrerasaurus, but it is narrower in Sanjuansaurus.
The minimum width of the blade (near the base) is 54% the width of the
acromial margin, less than that found in either small (83%; PVSJ 407)
or very large individuals (60%; PVSJ 053) of Herrerasaurus. (2) The pubis shaft of Sanjuansaurus lacks the strong proximal curvature characteristic of Herrerasaurus (
Phylogenetic analysis resulted in a single most
parsimonious tree of 190 steps (consistency index 0.553, retention
index 0.593). An implicit enumeration search (
The analysis supports the hypotheses that Dinosauria and Herrerasauridae (Staurikosaurus pricei + Herrerasaurus ischigualastensis + Sanjuansaurus gordilloi) are monophyletic and that Herrerasauridae is positioned at the base of Saurischia outside of Eusaurischia, a result similar to that presented by Langer and Benton (
Seven synapomorphies support the clade Herrerasauridae
in the consensus tree (characters 20.1, 39.1, 45.1, 46.1, 47.1,
69.2, and 77.1). Only four of those (characters 39, 45, 46, and 77)
can be scored in all herrerasaurids, and character 20 is the only one
that cannot be determined in Sanjuansaurus. The unambiguous synapomorphies that unite the herrerasaurids in this analysis are the same as those previously identified (
Other features that support a grouping Sanjuansaurus + Herrerasaurus,
but are ambiguous at present include: a narrow “U” shaped antorbital
fossa with a promaxillary fenestra located on the anterodorsal border;
centrum of the sixth cervical vertebra longest in the cervical series;
spine tables on the distal end of the last dorsal and the sacral neural
spines; two sacral vertebrae with the addition of one incipient
dorsosacral; strap-shaped scapular blade that forms an angle of more
than 90° with the acromion; size and shape of the pubic foot; and
sinuous lateral border of the pubis in anterior view. There are a few
ambiguous character-states shared by Sanjuansaurus and Staurikosaurus but not with Herrerasaurus:
shortness of the pubis relative to the femur and the subcircular distal
end of the tibia in distal view. The latter character was originally
considered as an autapomorphy of Staurikosaurus (
On the other hand, many characters of Sanjuansaurus are unlike those inother herrerasaurids (where known): long band-shaped transverse processes of the distal cervicals; deep lateral and ventral concavity in cervical and dorsal centra; short pubis with wide subcircular obturator foramen; and fused astragalus and calcaneum with a tabular ascending process (if this character-state is not pathological in origin).
Faunal ConsiderationsThe co-occurrence in the basal portion of the Ischigualasto Formation, of two herrerasaurids (Sanjuansaurus, Herrerasaurus), a basal saurischian (Eoraptor), and two sauropodomorphs (Panphagia, Chromogisaurus)
suggests that saurischian dinosaurs were already highly diversified in
southwestern Pangea early in the Late Triassic. The new herrerasaurid
also represents another large-bodied predatory dinosaur in the
Carnian-age Ischigualasto fauna, contrasting with the rarity of
carnivorous dinosaurs in the successive Norian assemblage of the
overlaying Los Colorados Formation. In that richly represented
assemblage, only a single specimen of a coelophysoid theropod has been
recovered to date (
We are especially grateful to Jeffrey Wilson and Augusto Haro for their suggestions on the manuscript. We thank Paul Sereno and Fernando Novas who improved the quality of the manuscript with their comments. We are indebted to the field crew and Earthwatch volunteers of 1994/1995. Fieldwork funding was provided by Earthwatch Institute. We also thank Raul Gordillo and Diego Abelín for skillfull preparation of the fossil material and Ivan Sabrodski for photography.