





(C) 2011 Claudia Drees. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
We investigated the extent of poleward shifts in the distribution range of Agonum viridicupreum due to climate change in the western Palaearctic. Species’ records were obtained from extensive literature sources as well as from collections, and consistent amateur entomologists’ recordings. Within the general geographic range of the species, we analyzed in detail two parts of both, the northern and southern distribution range boundaries: (1 and 2) north-western Germany (leading or high-latitude edge), (3) Israel and (4) southern Italy (rear or low-latitude edge). Temporal changes in the occurrence data of the species indicated a northward shift of the leading edge of a minimum of 100 km within the last 50 to 100 years. In contrast, according to the data gathered, the rear edge has not changed during the last decades. Further studies are needed in order to fully understand the underlying mechanisms of the different behaviour of leading and rear range edges of Agonum viridicupreum in the current context of global change. Despite our incomplete understanding, chronosequences of the occurrence of the given species have the potential to optimize climate niche modelling to predict trends in the distribution range in the future.
chronosequence, climate change, distribution area, global change, wetlands, power of dispersal, migration, range shift
For about 250 years, man has released radiatively active
gases and particles in substantial amounts into the atmosphere. As one
of the consequences, the global mean near surface temperature has
increased, a phenomenon commonly referred to as ‘global warming’ or
‘climate change’. Deduced mainly from instrumental observations
initiated around 1860, the observed climate change can be attributed to
a large extent to human activities, which influence not only global
temperature, but also pH-values of the oceans, precipitation and the
general hydrological cycles on Earth (
For many animal and plant species, theoretical analyses
on the climate determination of the species’ occurrence have predicted a
general poleward shift and (in mountain areas) an uphill shift of the
given distribution areas and populations, respectively, as a response
to climate change. In agreement with theory, numerous range shifts have
been documented in the last years. Examples are known from vascular
plants, birds, and many insects such as butterflies, dragonflies and
damselflies (
Poleward shifts of distribution areas are very likely
also for widely distributed carabid species (in contrast to species with
restricted distribution areas, i.e. endemics), as their patterns of
geographic distribution are strongly determined by climatic factors (as
shown by a large-scale analysis of West Palaearctic ground beetle
diversity,
Moreover, ground beetles with their northern
distribution limit in Britain have moved about 50 km northwards within a
period of about two decades (
However, previous studies on poleward range margin
shifts of ground beetles have mainly focused on the leading (i.e.
current high altitudinal and latitudinal) edges of their distribution
areas (literature cited above). Changes occurring at the leading edge
are interesting, especially in the framework of dispersal biology, and
they enable us to understand many population biological processes (
Nevertheless, up to now, there is no available study
comparing the reaction of a ground beetle species at both margins of its
distribution range. Thus, in this study we aimed at investigating the
extent of poleward range shifts at both the leading and rear edges of
the distribution area of a carabid species due to recent climate
changes. We selected Agonum viridicupreum
as our study object because it fulfils many preconditions of a suitable
model species to assess potential margin shifts: The specimens can be
easily found in the field, they are fully winged and fly actively, and
the species’ habitat preferences are well-known. Furthermore, the
specimens are nicely coloured, stimulating many amateur entomologists
to record the species, and, therefore, allowing suitable faunistic
data from large parts of its distribution area. Moreover, the species
is not restricted to habitats that are influenced or even destroyed by
other drivers of global change, nor have been altered simultaneously by
the temperature increase in the last decades (e.g. oligotrophic peat
bogs affected by increased atmospheric nitrogen depositions due to
pollution,
Agonum viridicupreum
(Goeze, 1777) is a macropterous and thermophilous species restricted
to open, wet habitats such as meadows, fens and rain ponds. The
day-active beetle prefers sun-exposed muddy sites where it can be easily
detected by its green-bronze-coloured surface. Due to its occurrence in
floodplain areas (with high probability of diversion), the dispersal
of individuals is not only determined by the species’ ability to fly.
Specimens can be transported downstream by flooding events into areas
where the species might not be able to establish autochthonous
populations (
In the Levant (Middle East, see below), the beetle lives in wet habitats, mostly close to winter or rain ponds (personal observations). In southern Italy (Calabria) the species lives in river bank habitats around Typha swamps or in other wet vegetation types and crops, and on lake shores, until about 1400 m above sea level.
Distribution area and temporal changesWe reviewed the available faunistic literature for
the western Palaearctic (Europe, the Mediterranean area) to determine
the general distribution area of the study species (
The situation of faunistic recordings is sufficient for one region at the northern distribution edge (north-western Germany) and for two regions at the southern distribution edge (Levant in the Middle East, mainly Israel, and Calabria in southern Italy).
– North-western Germany has been studied by numerous amateur entomologists who have greatly contributed to our knowledge on the geographic distribution of carabid beetles. We therefore analyzed the changes in the species’ distribution separately for (a) West Lower Saxony (west of river Weser) and for (b) East Lower Saxony (east of river Weser), Hamburg, and Schleswig Holstein. For these regions records from three periods (before 1950, between 1951 and 1980, after 1980) were summed up to document tendencies in the numbers of catches.
– For Israel, the first records date from the 1920s (the beginning of modern zoological exploration of the given region by local scientists, in former times only explorers from abroad collected beetles there). We therefore distinguished only two periods of collecting: before 1980 and after 1980.
– For southern Italy (Calabria) there are scarce
historical records (before 1980). However, after 1980, intensive
ecological surveys were carried out on populations in several sites of
the Crati river valley (
Consequently, within the global distribution area of
the study species, we analyzed the northern and southern range
boundaries by studying in detail the three above mentioned concrete
margin regions, where the coverage of the faunistic recordings is amply
and sufficient: (1 and 2) a part of the leading edge (north-western
Germany: Lower Saxony, Hamburg and Schleswig-Holstein, divided into
regions west and east of the river Weser), and (3 and 4) the only
continental areas at the rear edge that are not limited by the sea or by
the presence of Agonum fulgidicolle
(the Levant in the Middle East: mainly Israel, and Calabria,
(southern Italy)). For these areas we compiled numerous faunistic
records mostly published in local journals (
We surveyed climatological literature and compiled information about recent climate changes in the three regions north-western Germany, Israel and southern Italy (Calabria). We focussed only on changes in temperature and precipitation, the main factors influencing the ground beetles’ biology and distribution.
Results Climate changesNorth-West Germany – warmer springs with wetter winters and drier summers
Over the last 150 years a considerable increase of the global mean temperature
by about 0.8°C has been observed. Also for western Europe the
measurements show a warming trend. For Germany during the 20th century a
mean temperature rise of about 1.0°C was reported by
Linear 20th century precipitation trends for Germany have been reported to be about 8.5% (an increase from 750 mm to 800 mm,
An analysis for the period 1964 to 1994 of temperature
measurements at 40 stations evenly distributed over Israel came to the
conclusion that there appears to be a general warming trend, with some
local exceptions, i.e. in the south, which could be related to
enhanced aerosol emission (
Concerning climatological precipitation trends
the Levant has to be divided into a southern and northern part. An
analysis of winter half-year precipitation over the entire Mediterranean
region reveals predominating rainfall decreases during the last 50
years. The areas deviating from this general trend includes southern
Israel (
From the maps of linear trends in annual mean temperature for Europe compiled by
Overall it can be said that the most southern part of
Italy and especially Calabria has become drier over the last decades.
While the linear trend in annual precipitation for the entire
20th century for the Calabria region is almost zero, a pronounced trend
exists for the period from 1951 to 2000 with a decrease in
precipitation by about 20% in the annual mean with a decrease in summer
precipitation of about 40% (
The distribution area of the species within the western part of the Palaearctic is given in Fig. 1. Agonum viridicupreum occurs around the Mediterranean Sea (with a distribution gap in north-eastern Africa). The northern edge of the distribution area runs from the Netherlands through northern Germany and Poland (see also Fig. 2). In the south-east the species occurs in Turkey, Lebanon, and Israel.
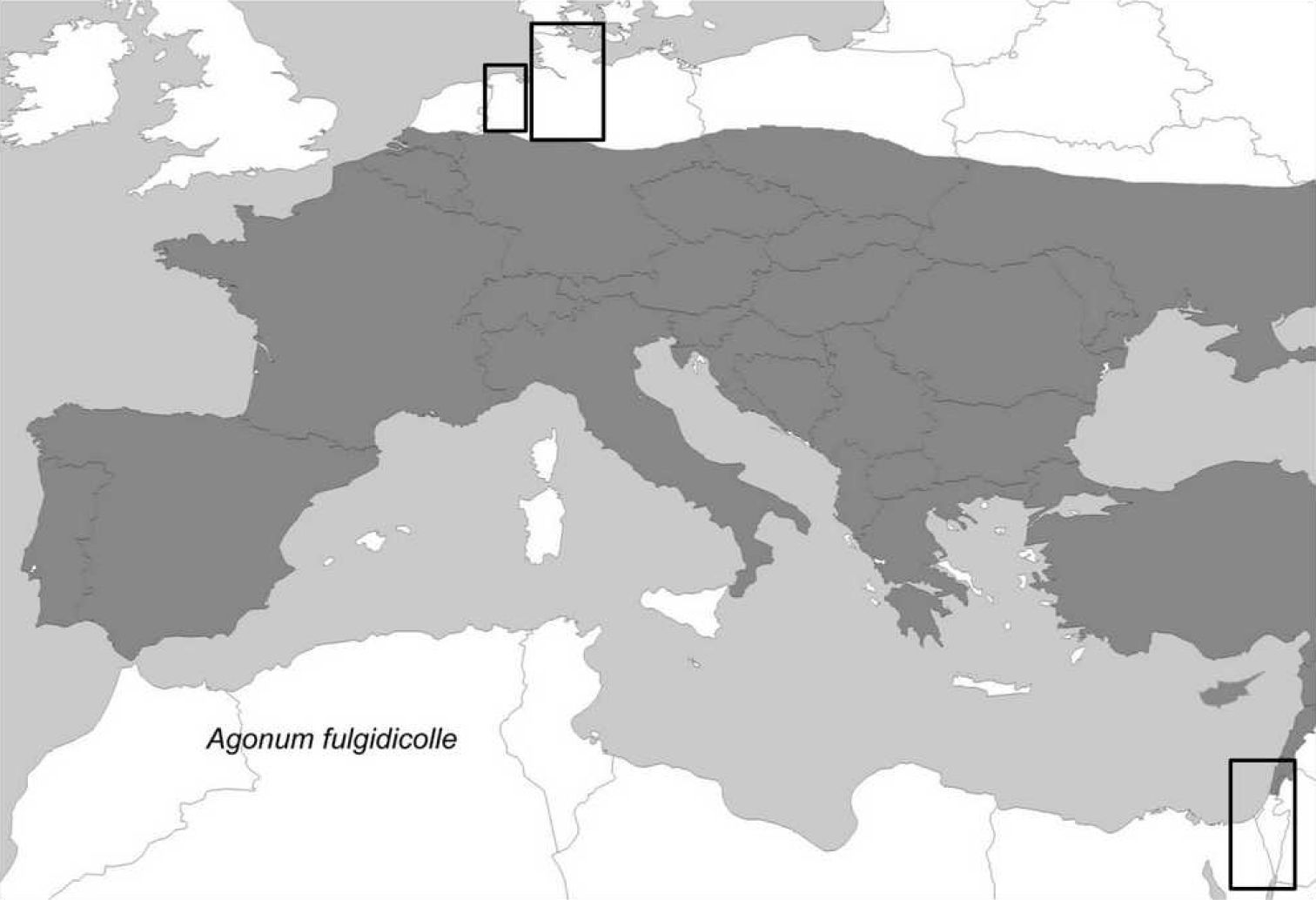
Distribution of Agonum viridicupreum (shaded in grey) and its sister taxon Agonum fulgidicolle in the western Palaearctic. Map modified after
In the southern Iberian Peninsula and Morocco, the beetle prefers mountainous areas (
The south-eastern distribution edge in the Levant virtually coincides with the border of the Mediterranean climate (Fig. 3). In this study, we report the first record for Egypt ([(T)El Arish, Sinai, leg. L. Fishelsohn, 12.03.1956], record in TAU, Fig. 3). However, the single specimen collected is not a proof of the existence of an autochthonous population here. The same is true for records obtained from the desert regions (e.g. Dead Sea Region, where no suitable habitats for the species occur, cf. Fig. 3).
Faunistic analyses of the distribution margins West Lower Saxony (west of the river Weser):Although Agonum viridicupreum
has been known from the Netherlands since the 19th century, no
specimens were recorded from West Lower Saxony until the 1980s (Table 1).
Indeed, in the 1950s the northern distribution limit of the species’
range was located southwards of Lower Saxony, in the Westphalian
Lowlands (
Number of records of Agonum viridicupreum in the different periods of time in north-western Germany (leading edge) and Israel (rear edge).
| Study region | Number of records | ||
|---|---|---|---|
| before 1950 | 1950–1980 | after 1980 | |
| Leading edge: | |||
| North-West Germany – West Lower Saxony | 0 | 0 | 26 |
| North-West Germany – East Lower Saxony and Schleswig Holstein | 11 | 12 | 24 |
| Rear edge: | |||
| Israel | 12 | 14 | |
2 Pevestorf [river Elbe, south-east of Lüneburg]
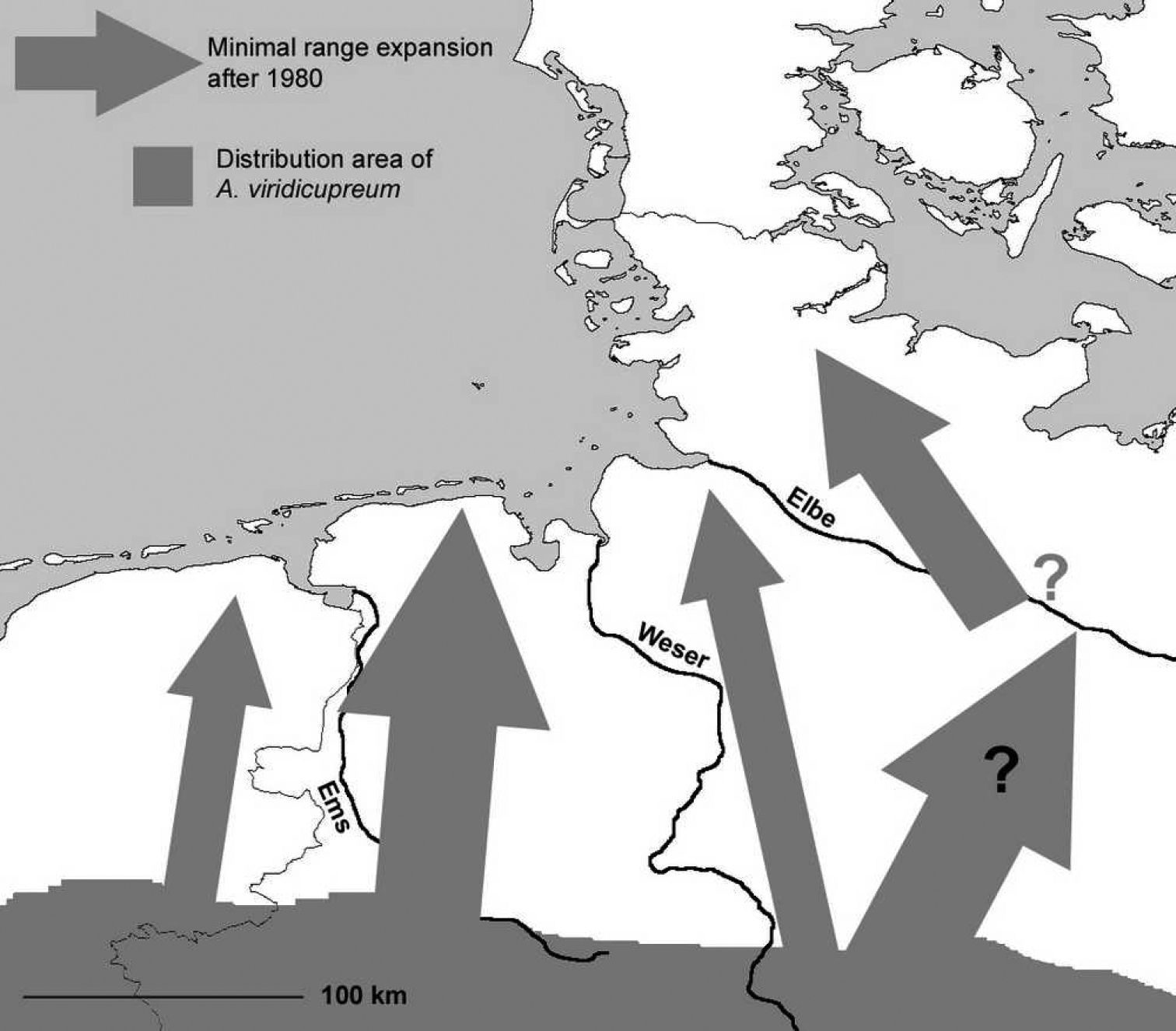
Distribution of Agonum viridicupreum (shaded in grey) in North-West Germany with eastern parts of the Netherlands. Arrows indicate minimum range expansion in the last three decades (for explanation and records see text). Range expansion in the Netherlands indicated after Turin, pers. comm.
Between 1951 and 1980, only one record from another site in the Lower Saxonian floodplain area of the river Elbe is known (Table 1). Records from sites outside the given floodplain are exclusively known since 1981, when the number of records greatly increased.
Today, the species is found northwards, up to central Schleswig-Holstein (www.entomologie.de/hamburg/karten/fhl_02/_agovir1.htm), and reaches also the north-western parts of the considered area. Interpreting the old records from the floodplain area of the river Elbe as autochthonous populations leads us to think that the species’ geographic range has experienced a northward shift of about 100 km during the last century. Even if these records were not seen as autochthonous populations, the shift would have spanned over about 200 km (Fig. 2).
Levant: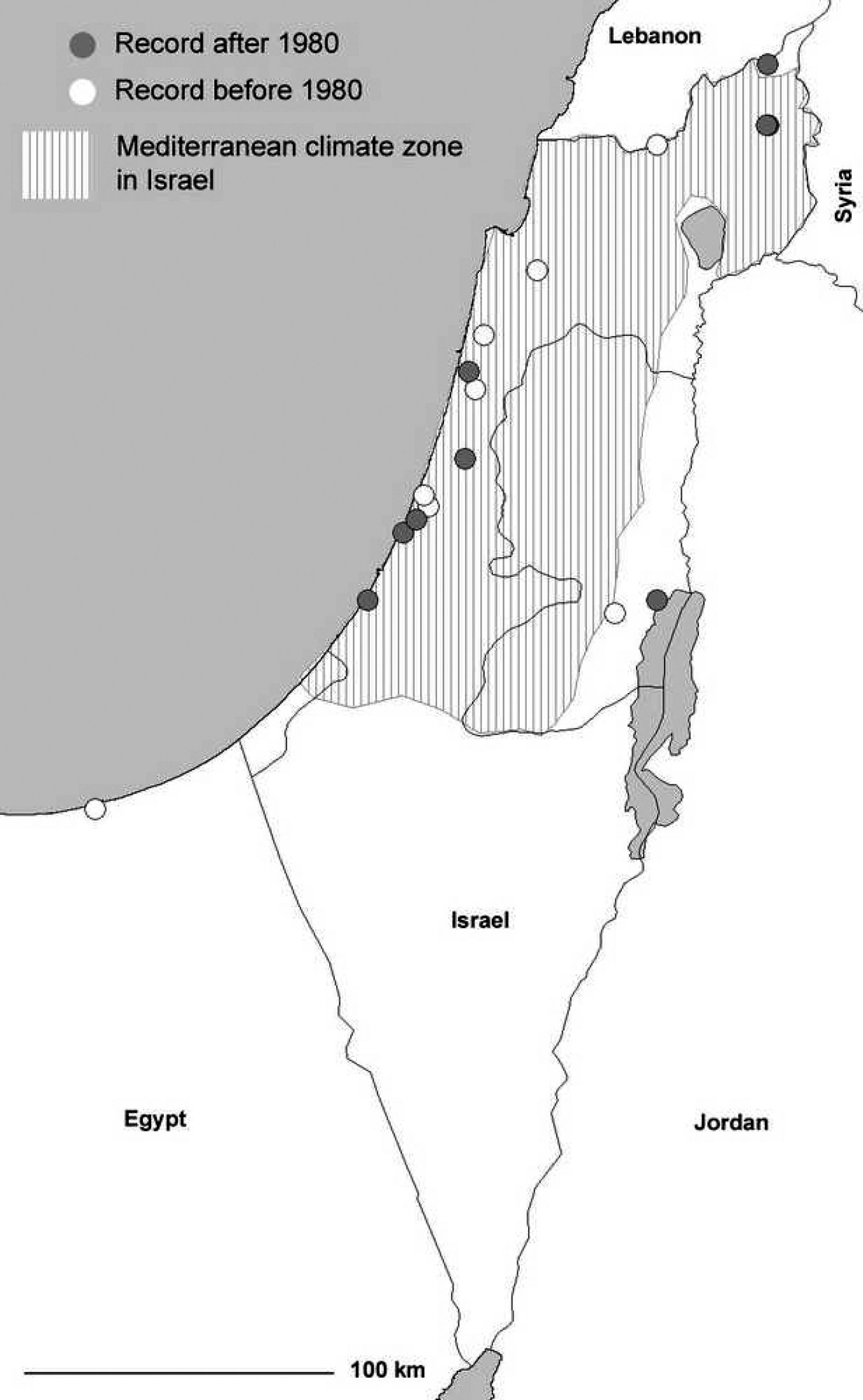
Distribution of Agonum viridicupreum in Israel. The striped area indicates Mediterranean climate zone (according to
For southern Italy (Calabria) there are scarce historical records (before 1980;
The compilation of the faunistic data showed that the distribution range of Agonum viridicupreum
had significantly shifted northwards within the last 50 to 100 years.
Up to 1950, in the analyzed region, the northern edge of the species’
distribution had stretched from the Netherlands (Nijmegen, Enschede;
Today, in the western part of Lower Saxony, Agonum viridicupreum can be found up to the North Sea, confirming a northwards range expansion of about 100 km. Similarly, in the neighbouring Netherlands the beetle has expanded its distribution range northwards and can nowadays be recorded close to the city of Groningen (Turin, pers. comm.). Also in the eastern part of Lower Saxony the species has spread northwards a minimum distance of 100 km and it reaches the centre of Schleswig-Holstein today. These results allow the assumption that a further temperature rise will make the species’ occurrence in Denmark highly probable in the near future.
Stable rear edgeUnlike the northern distribution edge, the southern range margin (rear edge) of Agonum viridicupreum has not changed within the last decades. Indeed, there are still populations with numerous individuals south of Tel Aviv, which is close to the southern limit of the Mediterranean climate. Consequently, in Israel we expect the beetle to occur in most of the regions characterized by Mediterranean climate. In southern Italy (Calabria) Agonum viridicupreum shows a stable rear edge north of the 39th parallel, with permanent populations in the Crati Valley in the Cosenza province, in areas marked by Submediterranean or Mediterranean climate.
Different behaviour of leading and rear edgesIn accordance with numerous other authors (e.g.
On the contrary, extrinsic factors may be acting differently at the leading and rear edge of the distribution range of Agonum viridicupreum. Our compilation of recent climate trends, however, reveals generally rising temperatures in all regions under study. In contrast, less consistent changes in precipitation can be observed. The populations in both, Calabria and the Levant will have to deal with a reduction in mean annual precipitation. In the face of the virtual exclusive occurrence of the species close to rain ponds in the southern edge of its distribution, it seems likely that the southern populations are limited by the given ground water tables which predominantly result from the annual amount of precipitation (mainly in the winter months).
In the Levant, larval development takes place during
winter and early spring, as revealed by numerous tenerals, even at
higher altitudes, e.g. 900 m a.s.l., in the Golan Heights, in April
and May (personal observation). In contrast, the northern populations
in Central Europe are unlikely to be limited by the amount of
precipitation, but rather by temperatures during the species’ activity
period. In fact, in this region, larval development takes place during
summer and tenerals occur in late summer and autumn (August to October;
Nonetheless it is possible that the northern and southern limits of Agonum viridicupreum are determined by different climatic factors: increasing summer temperature in the north and increasing precipitation during the winter in the south can explain the poleward shift of the leading and the stable rear edge of the given species’ distribution.
Potential of Agonum viridicupreum for further ecological research on global changeThis study is the first one that investigates
simultaneously the possible shifts of the northern and southern margins
of a carabid species’ distribution due to climate change. Undoubtedly,
at present we are not able to fully understand the underlying mechanisms
of the different behaviours of the leading and rear boundaries of the
geographic range of Agonum viridicupreum
in the actual context of global change. However, our analysis suggests
that the reaction of the study species to climate change may be more
intricate than expected at first. For this reason, we think that the
more complex situation in Agonum viridicupreum
has important potential for further carabidological investigations at
the interface of global change ecology and conservation biology. For
instance, predictions based on climate envelope modelling, which has
become both commonplace for many other animal species and the object of
an intensive (and critical) scientific discourse (
We thank Pascale Zumstein (Lüneburg) for her help with compiling records, the Hamburg Coleopterists group for providing additional records and Ariel-Leib-Leonid Friedman (Tel Aviv University) for helping with reading old Hebrew labels. Claudia Drees is a VATAT funded post-doctoral fellow at the Tel Aviv University Zoological Museum.