





(C) 2011 Lucija Šerić Jelaska. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Carabid beetle diversity and mean individual biomass (MIB) were analysed in three different successional stages of beech tree stands (60, 80 and 150 years old). Carabid beetles were captured using pitfall traps placed at nine sites (three per age class) in the Papuk Mountain of East Croatia during 2008. A cluster analysis identified three groupings that corresponded to the beech age classes. MIB values increased with stand age, ranging from 255 in 60-year-old stand to 537 in the oldest forests. The 80-year-old stand showed the highest species richness and diversity values. With respect to species composition, large species such as Carabus scheidleri and Carabus coriaceus were dominant only in the oldest forests. Furthermore, species that overwinter in the larval stage were more abundant in the oldest forests (45% of the total number of individuals from the 150-year-old stand) than in the younger ones (20% of individuals from 60-year-old, and 22% of individuals from 80-year-old stands). Our results showed that the analyses of species composition and life history traits are valuable for estimating the conservation values of older forests. Although the investigated sites form part of a continuous forested area and are only a couple of kilometres apart, MIB values detect significant differences associated with forest age and can be a useful tool in evaluating the degree to which a forest reflects a natural state.
carabid beetles, MIB, over-wintering stages, beech forests succession, Papuk Nature Park
In forest management, sustainability is an
internationally accepted goal. One technique by which sustainability is
assessed is through the monitoring of indicator species (
Generally, it is assumed that older forests have less
species richness than younger stages, but analysing species richness
alone without considering species composition (e.g., forest
specialists, saproxilic species) would underestimate the conservation
value of older forests (
According to changes in species body sizes as well as
their community composition, the Mean Individual Biomass (MIB) index of
carabid beetles has been proposed as a good indicator of succession (
The aim of this study is to compare carabid beetle
assemblages (species composition and richness) and MIB index values in
mesophyllous beech forest stands of various successional ages (60-, 80-
and 150-year-old forests). Our hypotheses are that the Mean Individual
Biomass index will increase with forest age and species with
over-wintering larvae will be more abundant in the older forests.
Similar trends were evident from previous studies (
The study was performed in the Papuk Nature Park in continental Croatia (Fig. 1). The area was proclaimed protected in 1999 and represents a hilly and forested wildlife area within a lowland, agricultural region. The highest peak is 954 m above sea level (a.s.l.). The total area of Papuk Nature Park is 336 km2 and is mostly (96%) covered by forest, and the rest of the habitat consists of settlements and small agricultural areas. Sessile oak (Quercus petraea (Mattuschka) Liebl.) forests dominate areas up to 350 m a.s.l. Beech trees (Fagus sylvatica L.), depending upon geological substrate and microclimatic conditions, grow in several different forest associations and cover more than 50% of the forested area, whereas mixed beech-fir forests grow in areas higher than 700 m a.s.l. Forests are of natural origin but are influenced by forestry. Deciduous forests are managed as even-age stands. Beech-fir forests are managed following the selection cutting system. Logged areas are mainly under natural regeneration. Most forests in the Park are 60 to 80 years old. Jankovac forest (660 ha), which has been left unmanaged, is the only large beech-forest stand that is more than 150 years old. The carabid fauna was compared among three different beech forest stand ages: younger (60-year-old forest), middle-aged (80-year-old forest) and old beech forest (150 years old). For this study, we selected nine sampling sites (three per age class) (Fig. 1).
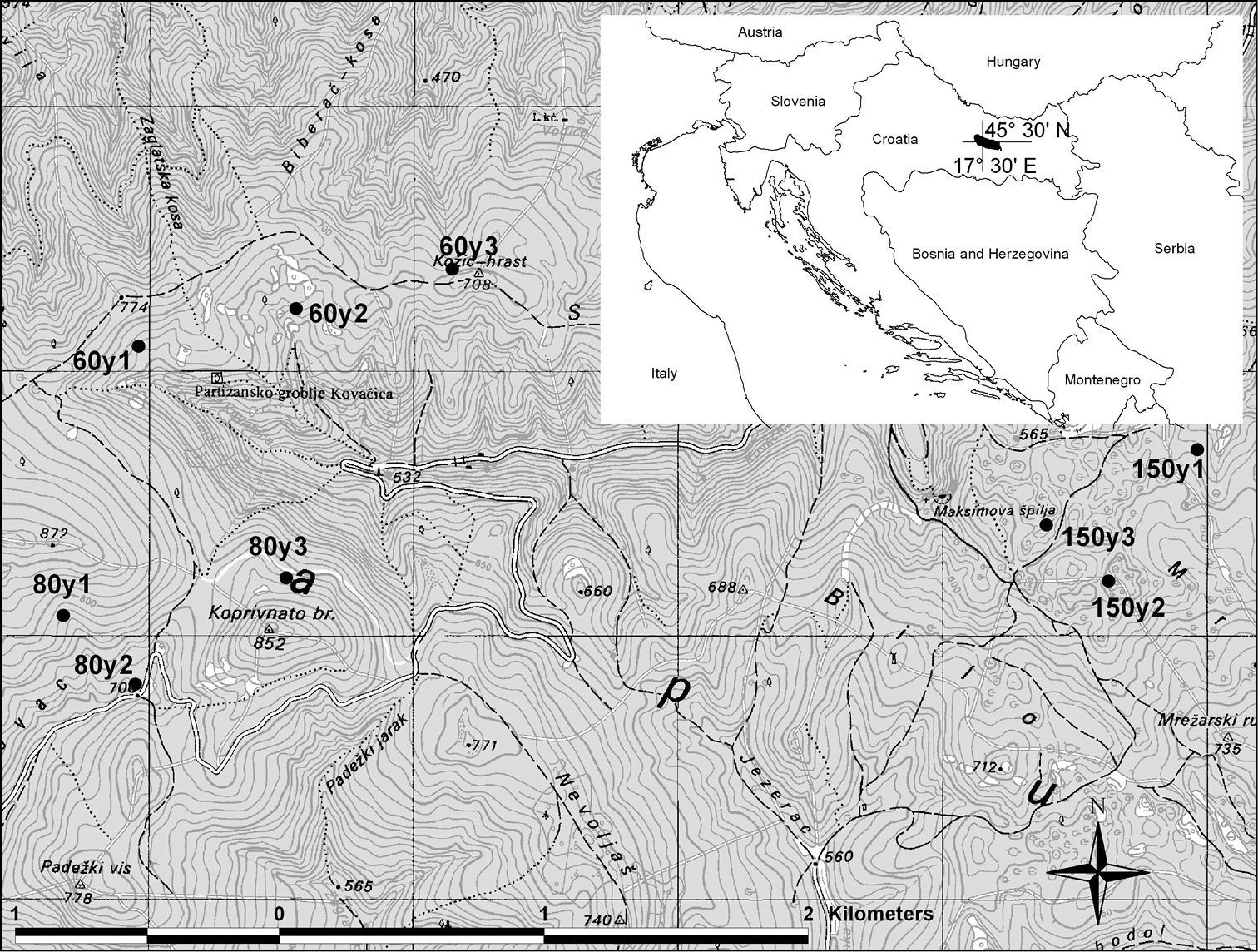
Position of investigated sites labelled according to the age of the forest (“60y 1–3” denote sites 1 to 3 in the 60-year-old forest, “80y 1–3” denote sites 1 to 3 in the 80-year-old forest and “150y 1–3” denote sites 1 to 3 in the 150-year-old forest). Insert: location of Papuk Nature Park in Croatia.
Carabid beetles were collected from mid May to mid September in 2008. We used plastic pitfall traps (0.5 L volume, 10 cm diameter) filled with a mixture of 96% ethanol, 9% acetic acid and water in equal proportions and covered with roofs for protection against rain and evaporation. At each sampling site, three individual traps were placed 10 m apart to form a triangle. Traps were emptied on the same day approximately every three weeks (a total of six visits).
Carabid beetles were collected using 27 traps from 162 trapping events altogether (3 traps x 9 sampling sites x 6 visits). Samples were pooled per site across all visits, resulting in 9 sampling sites (three per age class) for analyses.
The species collected were identified using keys (
To compare carabid diversity, we calculated Margalef
species richness (number of species per standardised number of
individuals), Shannon diversity index and Pielou’s evenness. For
similarity between sites, Bray-Curtis indices using presence/absence
data, and the number of individuals were calculated and used in cluster
analyses with the complete linkage method for constructing dendrograms (
ln y = -8.92804283 + 2.5554921ln x,
where x is the body length of a specimen and y is the live estimated body weight of the individual. The estimated biomass of a species was calculated by multiplying the estimated body weight of the individual times its abundance for all sampled individuals per sampling site.
Differences in MIB values and diversity parameters among the three forest age classes were tested using analyses of covariance (ANCOVA), where mean individual biomass, species richness, evenness and Shannon diversity index were used as dependant variables, forest ages as categorical predictors and proportion of species with respect to overwintering strategy as the covariate, followed by the Scheffé post-hoc test. Pearson product-moment correlation coefficient was used for analyses. Statistical tests were performed using Primer 6 (PRIMER E Inc 2002) and Statistica 8 (Statsoft Inc 2008).
ResultsA total of 1244 carabid beetles belonging to 31 species were collected (Table 1).
Between 9 and 18 species were collected per sampling site. Similar
species richness levels have been recorded in other beech forests in
nearby countries as well as in similar forest communities in Croatia (
List of species, their mean body sizes (authors’
measurements), over-wintering stage (a-adults, l-larvae), estimated
body weight values and number of individuals in 60- , 80- and
150-year-old forests. Body weight values were calculated according to
| Species | Mean body size /mm | Over-winteringstage | Body weight /mg | Number of individuals | ||
|---|---|---|---|---|---|---|
| 60 y | 80y | 150y | ||||
| Abax carinatus (Duftschmid 1812) | 16 | a | 158.4 | 139 | 228 | 133 |
| Abax parallelepipedus (Piller & Mitterpacher 1783) | 20 | a | 280.1 | 1 | 32 | 62 |
| Abax parallelus (Duftschmid 1812) | 17 | a | 184.9 | 0 | 26 | 15 |
| Aptinus bombarda (Illiger 1800) | 12 | a | 75.9 | 1 | 36 | 3 |
| Calosoma inquisitor (Linné 1758) | 21 | l | 317.3 | 2 | 1 | 1 |
| Carabus arcensis Herbst 1784 | 20 | a | 280.1 | 36 | 38 | 1 |
| Carabus convexus Fabricius 1775 | 17 | a | 184.9 | 1 | 0 | 1 |
| Carabus coriaceus Linné 1758 | 39 | l | 1543.7 | 0 | 0 | 29 |
| Carabus intricatus Linné 1761 | 30 | a | 789.5 | 3 | 0 | 1 |
| Carabus irregularis Fabricius 1792 | 25 | a | 495.5 | 1 | 0 | 0 |
| Carabus nemoralis O.F.Müller 1764 | 25 | a | 495.5 | 4 | 0 | 5 |
| Carabus scheidleri Panzer 1799 | 30 | l | 789.5 | 0 | 16 | 119 |
| Carabus ullrichii Germar 1824 | 25 | a | 495.5 | 3 | 53 | 20 |
| Carabus violaceus Linné 1758 | 32 | l | 931.1 | 42 | 75 | 46 |
| Cychrus attenuatus (Fabricius 1792) | 17 | l | 184.9 | 0 | 6 | 1 |
| Cychrus semigranosus Palliardi 1825 | 20 | l | 280.1 | 0 | 9 | 4 |
| Harpalus affinis (Schrank 1781) | 12 | l | 75.9 | 0 | 1 | 1 |
| Leistus piceus Frölich 1799 | 8 | a | 26.9 | 0 | 1 | 1 |
| Licinus hoffmannseggii (Panzer. 1797) | 13 | l | 93.2 | 0 | 4 | 0 |
| Limodromus assimilis (Paykull 1790) | 11 | a | 60.8 | 0 | 1 | 0 |
| Molops elatus (Fabricius 1801) | 19 | a | 245.7 | 0 | 0 | 1 |
| Molops piceus (Panzer 1793) | 11 | a | 60.8 | 3 | 5 | 4 |
| Myas chalybeus (Palliardi 1825) | 16 | 158.4 | 4 | 1 | 0 | |
| Notiophilus rufipes Curtis 1829 | 6 | a | 12.9 | 0 | 0 | 1 |
| Platyderus rufus (Duftschmid 1812) | 7 | l | 19.2 | 1 | 0 | 0 |
| Platynus scrobiculatus (Fabricius 1801) | 11 | l | 60.8 | 0 | 1 | 0 |
| Pseudoophonus rufipes (DeGeer 1774) | 16 | l | 158.4 | 2 | 1 | 1 |
| Pterostichus aethiops (Panzer 1796) | 14 | 112.6 | 0 | 0 | 1 | |
| Pterostichus niger Schaller 1783 | 23 | l | 400.4 | 0 | 4 | 0 |
| Pterostichus oblongopunctatus (Fabricius 1787) | 12 | a | 75.9 | 0 | 8 | 0 |
| Pterostichus transversalis (Duftschmid 1812) | 16 | 158.4 | 0 | 3 | 0 | |
| N (species) | 15 | 22 | 22 | |||
| N (individuals) | 243 | 550 | 451 | |||
The 80- and 150-year-old forests yielded 22 species each, whereas 15 species were collected from the 60-year-old forest sites. Nine species (29%) were present in all three forest age classes; one species was recorded only from the 60-year-old stand, six species only from the 80-year-old stand, and three only from the oldest forests.
The greatest number of individuals (44%) was recorded from the 80-year-old forest sites, 36% from the 150-year-old forest sites, and 20% from the 60-year-old forest sites (Table 1). Two species, Abax carinatus and Carabus violaceus, were collected from all nine sites with the greatest number of individuals, accounting for 40.2 and 13.1% of the total catch, respectively. Although Abax carinatus and Carabus violaceus were numerous at all three forest ages, two large species, Carabus scheidleri and Carabus coriaceus, were mainly collected from the 150-year-old sites. Carabus scheidleri and Carabus coriaceus accounted for 33% of the 451 individuals in the oldest forest and 12% of the total catch.
The 80-year-old sites showed the highest average Shannon-Wiener index value, whereas the lowest value was recorded from the youngest sites. The variability of data in the whole sample could be explained by the second order polynomial with R2 = 0.54. The same trend was observed for the average standardised species richness (Margalef’s index) (Table 2). MIB values increased with forest age. Beech forests that are 150 years old have the highest mean individual biomass values for carabids (Table 2, Fig. 2). Similar trends were recorded for proportion of species that hibernate as larvae. There were more carabid beetles that hibernate as larvae (45% of individuals collected) in the oldest forest than in the younger forest stages; in the 60- and 80-year-old forests, 80 and 78% of specimens hibernate as adults, respectively (Fig. 3). MIB was highly correlated with the proportion of carabids that hibernate as larvae (r = 0.84, p < 0.05).

MIB values (mg) compared to the age of the forests stands (years).

Proportion of species according to their hibernation strategies (larvae – black columns, adults – white columns) in relation to forest age (years).
Number of species and individuals, diversity indices and MIB (mg) values from the nine sites in Papuk Nature Park, Croatia.
| Plots | Total species | Total number of individuals | Margalef species richness | Pielou’s evenness | Shannon-Wiener index | MIB (mg) |
|---|---|---|---|---|---|---|
| 60y1 | 9 | 43 | 2.13 | 0.72 | 1.59 | 458 |
| 60y2 | 9 | 118 | 1.68 | 0.48 | 1.06 | 255 |
| 60y3 | 10 | 82 | 2.04 | 0.63 | 1.45 | 367 |
| 80y1 | 15 | 80 | 3.19 | 0.80 | 2.18 | 343 |
| 80y2 | 11 | 342 | 1.71 | 0.62 | 1.49 | 260 |
| 80y3 | 17 | 128 | 3.30 | 0.74 | 2.11 | 496 |
| 150y1 | 9 | 110 | 1.70 | 0.81 | 1.77 | 515 |
| 150y2 | 11 | 175 | 1.94 | 0.76 | 1.82 | 537 |
| 150y3 | 18 | 166 | 3.33 | 0.66 | 1.90 | 532 |
| Average values per age | ||||||
| 60y | 9.33 | 81.00 | 1.95 | 0.61 | 1.37 | 360 |
| 80y | 14.33 | 183.33 | 2.74 | 0.72 | 1.92 | 366 |
| 150y | 12.67 | 150.33 | 2.32 | 0.74 | 1.83 | 528 |
MIB values, Shannon–Wiener indices, Margalef’s indices and Pielou’s evenness did not differ significantly among forests of various age groups (ANCOVA, p<0.05), (Table 3). The covariate, hibernation strategy as larvae, was significantly related to the Mean Individual Biomass and Shannon–Wiener indices values (ANCOVA, p<0.05), (Table 3). Although they differed among forest age-groups at significance level slightly higher than p=0.05, performed post hoc comparison confirmed significant differences in MIB values of carabids between 150 year old forests and younger sites (60 and 80 year old forests) and for Shannon–Wiener indices between the 80-year-old sites from the youngest and the oldest sites (Scheffé test, p<0.05, Table 3).
Based on presence/absence data, a cluster analysis identified three groups: one with all 60-year-old forest sites, one with all 80-year-old sites as well as one 150-year-old site, and a third group with two 150-year-old forest sites (Fig. 4). A cluster analysis based on the number of individuals, grouped 60-year-old sites with two 80-year-old sites and all 150-year-old sites with one 80-year-old site (figure not shown).

Dendrogram of cluster analyses among forest sites using presence/absence carabid beetle data. Two distinct clusters are formed at roughly 30% similarity. Marks 60y 1–3, 80y 1–3 and 150y. 1–3 denote investigated sites placed in the 60-, 80- and 150-year-old forests.
ANCOVA results for the effects of forest age on mean individual biomass, the Shannon-Weiner diversity index, Pielou’s evenness and Margalef’s richness of carabid beetles, followed by the Scheffé post-hoc test.
| Variables | SS | DF | MS | F | P | Scheffé test |
|---|---|---|---|---|---|---|
| Dependant variable: MIB (g) | ||||||
| Corrected Model | 0.100 | 3 | 0.033 | 39.883 | 0.001 | |
| Intercept | 0.079 | 1 | 0.079 | 95.071 | 0.000 | |
| Species that hibernate as larvae | 0.045 | 1 | 0.045 | 54.156 | 0.001 | 150>80=60 |
| Forest age (years) | 0.010 | 2 | 0.005 | 5.712 | 0.051 | |
| Error | 0.004 | 5 | 0.001 | |||
| R2 = 0.960 (Adjusted R2 = 0.963) | ||||||
| Dependant variable : Shannon-Wiener index | ||||||
| Corrected Model | 0.796 | 3 | 0.265 | 7.268 | 0.028 | |
| Intercept | 2.184 | 1 | 2.184 | 59.831 | 0.001 | |
| Species that hibernate as larvae | 0.265 | 1 | 0.265 | 7.251 | 0.043 | 80>150=60 |
| Forest age (years) | 0.338 | 2 | 0.169 | 4.634 | 0.073 | |
| Error | 0.183 | 5 | 0.037 | |||
| R2 = 0.813 (Adjusted R2 = 0.702) | ||||||
| Dependant variable: Pielou’s evenness | ||||||
| Corrected Model | 0.055 | 3 | 0.018 | 2.786 | 0.149 | |
| Intercept | 0.414 | 1 | 0.414 | 63.233 | 0.001 | |
| Species that hibernate as larvae | 0.026 | 1 | 0.026 | 3.948 | 0.104 | |
| Forest age (years) | 0.011 | 2 | 0.005 | 0.804 | 0.498 | |
| Error | 0.033 | 5 | 0.007 | |||
| R2 = 0.626 (Adjusted R2 = 0.401) | ||||||
| Dependant variable: Margalef richness | ||||||
| Corrected Model | 1.651 | 3 | 0.550 | 1.099 | 0.431 | |
| Intercept | 3.558 | 1 | 3.558 | 7.106 | 0.045 | |
| Species that hibernate as larvae | 0.722 | 1 | 0.722 | 1.442 | 0.284 | |
| Forest age (years) | 0.919 | 2 | 0.459 | 0.918 | 0.458 | |
| Error | 2.504 | 5 | 0.501 | |||
| R2 = 0.397 (Adjusted R2 = 0.036) | ||||||
MIB values increased with forest age, confirming MIB as a
useful indicator in evaluating later succession stages. An increase in
MIB values was accompanied by changes in community structure, i.e., a
decreasing portion of smaller carabids and an increasing proportion of
the largest species, as well as their abundance, with forest age. A
similar trend has been reported in
Although evenness slightly increased with forest age,
other diversity parameters showed nonlinear patterns, with the highest
average values for forests of mid-age. Cluster analyses of carabid
composition in this study clearly divided the youngest and the oldest
forests, whereas middle-aged forests occupied an intermediate position.
Differences within 80-year-old forest sites were more pronounced than
between sites from other age classes that show a more diverse data set.
Similar results were found by
Forest age could affect the carabids assemblages due to
changes in habitat structure, where early and later forest successional
stages differ in vegetation structure, and in the accumulation of dead
and decaying wood (
To summarise, MIB values showed significant differences associated with forest age and can be a useful tool in evaluating whether a forest reflects a natural succession. Furthermore, our study indicates that for preserving stable communities and overall carabid diversity, it is important to have part of the forest unmanaged or at least leave some stands to reach the decomposition phase, taking into account the spatial connectivity of stands enabling the migration of species.
We would like to thank Papuk Nature Park for technical support for this research, Edin Lugić for field and laboratory assistance, and Sven Jelaska, Johan Kotze and two anonymous referees for critical reading of the manuscript and providing many valuable comments and suggestions.