





(C) 2011 Radomir Jaskuła. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
The tiger beetle fauna of the Balkan Peninsula is one of the richest in Europe and includes 19 species or 41% of the European tiger beetle fauna. Assembled by their biogeographical origins, the Balkan tiger beetle species fall into 14 different groups that include, Mediterranean, Middle Oriental, Central Asiatic, Euro-Siberian, South and East European, Pannonian-Sarmatian, West Palaearctic, Turano-European and Afrotropico Indo-Mediterranean species. The Mediterranean Sclerophyl and the Pontian Steppe are the Balkan biogeographical provinces with the highest species richness, while the Balkan Highlands has the lowest Cicindelidae diversity. Most species are restricted to single habitat types in lowland areas of the Balkan Peninsula and only Calomera aulica aulica and Calomera littoralis nemoralis occur in respectively 3 and 4 different types of habitat. About 60% of all Balkan Cicindelidae species are found in habitats potentially endangered by human activity.
Balkan Peninsula, biodiversity, distribution, Europe, zoogeography
Tiger beetles occur world-wide, with the exception of the polar regions and some oceanic islands (
The Balkan Peninsula is part of the Mediterranean basin.
It is one of the 25 most important world hotspot areas of biodiversity (
The first data on the tiger beetle fauna of the Balkan
Peninsula were published at the end of the 19th and beginning of the
20th century (
The aim of this paper is to summarize knowledge on the diversity of tiger beetles in the Balkan Peninsula with particular emphasis on total group diversity, zoogeographical composition, distribution, and ecological preferences of the species.
Study areaWe can define the Balkan Peninsula as a part of southeastern Europe with its northern boundary at the Danube, Sava and Kupa rivers. The rest of its margins are made up of the Black Sea in the east, the Adriatic Sea in the west, and the Mediterranean Sea (including the Aegean and Ionian seas) in the south (Fig. 1). The region has a combined area of ca. 550, 000 km2, which is nearly 5% of the entire European continent. The peninsula includes twelve countries, seven of which are completely confined to the Balkan Peninsula (Albania, Bulgaria, Greece, Macedonia FYR, Montenegro, Kosovo, and Bosnia-Herzegovina), and five (Romania, Serbia, Croatia, Slovenia, and Turkey) have only a part of their territories on the peninsula.
The largest surface of the Balkan Peninsula is
mountainous. Lowlands extend along the lower reaches of rivers that are
grouped into three catchments draining into the Adriatic, Aegean, and
Black Sea (
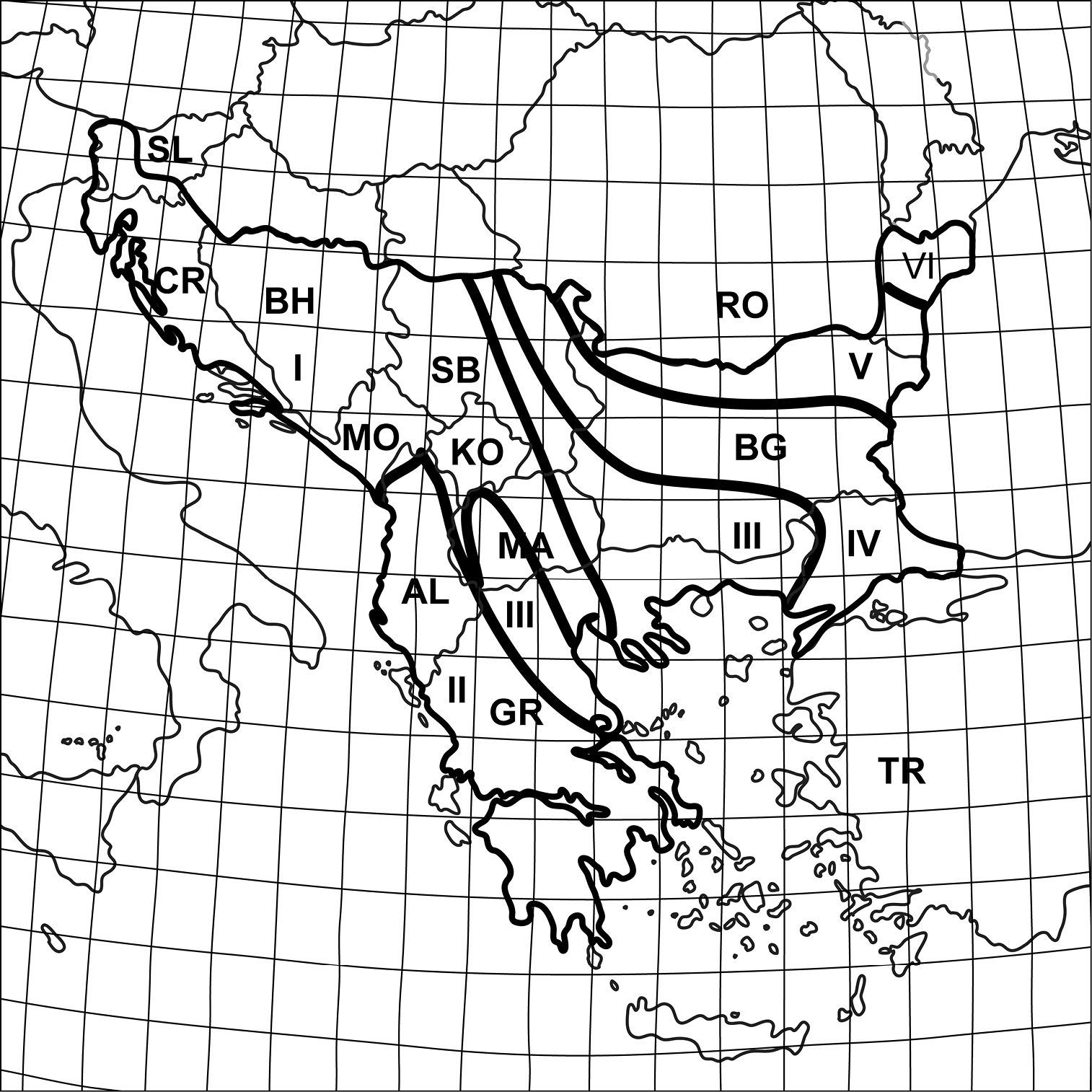
Geographical and administrative divisions of the Balkan Peninsula: I Dinaric region II Pindus region III Tracian-Macedonian region IV Balkanid region V Danubian plain region VI North-Dobroudzha region AL Albania BG Bulgaria BH Bosnia and Herzegovina CR Croatia GR Greece KO Kosovo MA Macedonia FYR, MO Montenegro RO Romania SB Serbia SL Slovenia TR Turkey.
According to
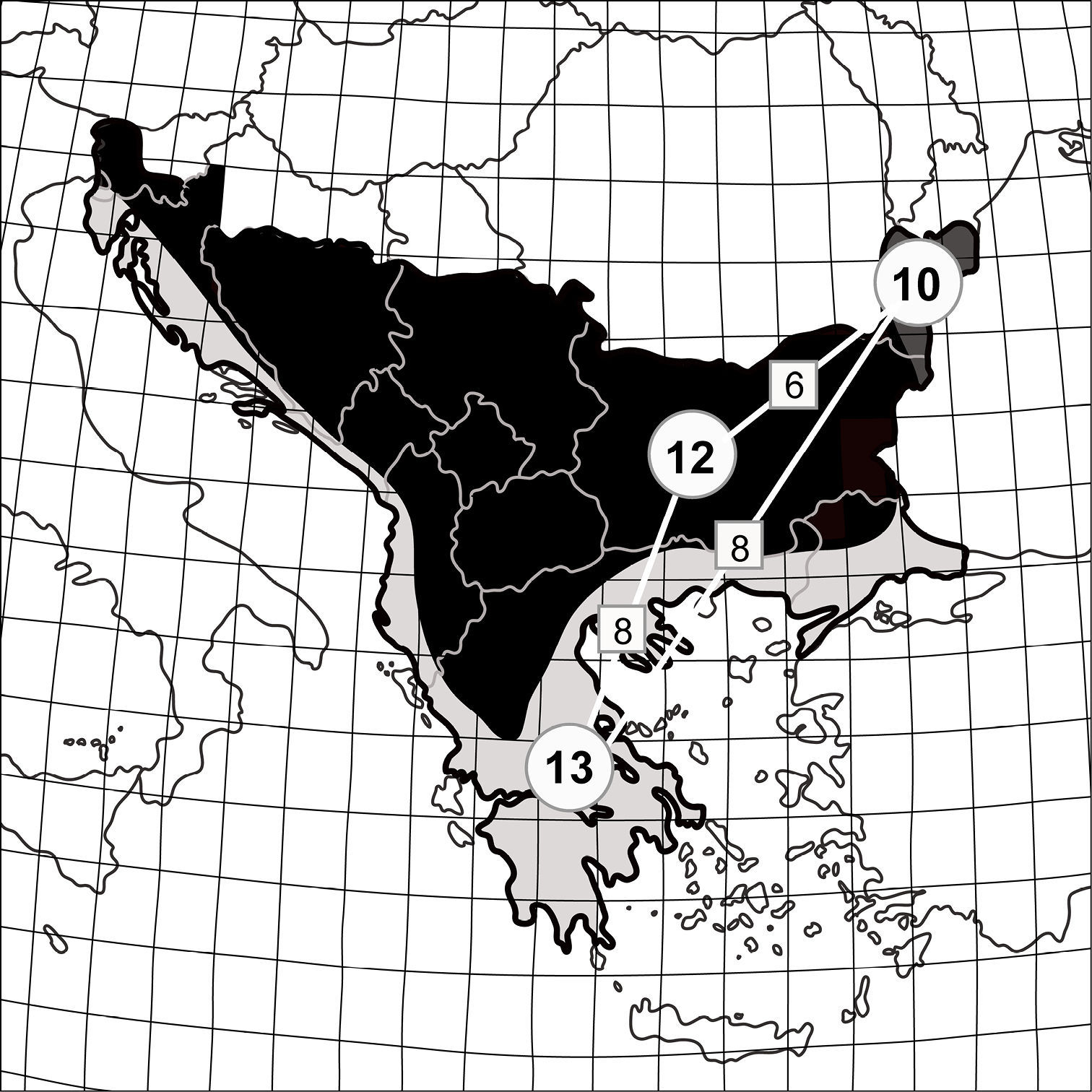
Tiger beetle faunas in the biogeographical provinces of the Balkan Peninsula (division after
The basis for this analysis of Balkan tiger beetles comes from published literature data; such as museum collections of the Museum and the Institute of Zoology, Polish Academy of Sciences (Warsaw, Poland), Royal Belgian Institute of Natural Sciences (Brussels, Belgium), Zoological Museum (Copenhagen, Denmark), Finnish Museum of Natural History (Helsinki, Finland), University of Montenegro (Podgorica, Montenegro); and original collections made by the author in the years 2005–2009 during five scientific expeditions covering almost all Balkan countries (“I-III Amphi-Balkan expeditions” and “Ist and IIIrd TB-Quest expeditions”).
Tiger beetle species richness and distribution of taxa
were analysed based on squares of 1o latitude and longitude. In each
square the number of all species recorded was summarized. Similarities
among tiger beetle fauna between geographical units were measured using
the Bray-Curtis index for presence/absence data (Primer v.2.0).
Jaccard’s (1902) index was used to present the degree of dissimilarity
between zoogeographic regions distinguished by
R= 100c/a+b-c
where: a = number of species in the richest fauna; b = number of species in the poorest fauna, c = number of species comon to both faunas.
Chorotypes follow
According to
Comparison of area and tiger beetle species richness of some European regions [based on
| Region | Area (km2) | Number of species | Species density (species number/1000 km2) |
|---|---|---|---|
| Balkan Peninsula | 550 000 | 19 | 0.034 |
| Iberian Peninsula | 580 000 | 19 | 0.033 |
| Italian Peninsula | 150 000 | 13 | 0.086 |
| Scandinavian Peninsula | 800 000 | 5 | 0.006 |
| France (mainland) | 675 000 | 14 | 0.021 |
| Ukraine | 603 700 | 19 | 0.031 |
| Russia (European part) | 4 268 850 | 23 | 0.005 |
The number of Balkan tiger beetle species is high compared with the number noted from other European regions with similar sized areas, especially north of the Balkan Peninsula (Table 1). Moreover, the diversity of the tiger beetle fauna in the studied area is similar to the fauna known from the entire territory of the European part of Russia. Among European regions with a similar area, only the Iberian Peninsula and the Ukraine exhibit similar numbers of tiger beetle species.
Balkan Cicindelidae belong to 14 different groups according to their geographical origin (
Chorotypes of Balkan tiger beetles (after
| Balkan endemics | Cicindela campestris oliviera, Cicindela monticola albanica |
|---|---|
| Mediterranean | Calomera littoralis nemoralis, Cephalota circumdata circumdata, Calomera aulica aulica |
| East Mediterranean | Calomera concolor concolor |
| West Mediterranean | Cylindera trisignata trisignata |
| Middle Oriental | Calomera fischeri fischeri |
| Central Asiatic | Cephalota chiloleuca, Cylindera contorta contorta |
| Northeast Mediterranean (Aegean) | Cephalota turcica, Cylindera trisignata hellenica |
| East European | Cephalota elegans stigmatohora |
| West Palaearctic | Cicindela campestris campestris, Cylindera germanica germanica, Cicindela hybrida |
| Turano-European | Cicindela monticola rumelica |
| South European | Cicindela sylvicola, Cylindera germanica muelleri |
| Pannonian-Sarmatian | Cicindela soluta pannonica |
| Euro-Siberian | Cicindela sylvatica, Cylindera arenaria viennensis |
| Afrotropico Indo-Mediterranean | Myriochila melancholica melancholica |
Within the Balkan Peninsula, species richness of
particular regions differs both in number of taxa and species
composition. Records from the literature and my own observations within
single squares of 1o latitude and longitude show that the highest
numbers of tiger beetle species are along sea coasts (Fig. 3). Moreover, within biogeographic provinces as definied by
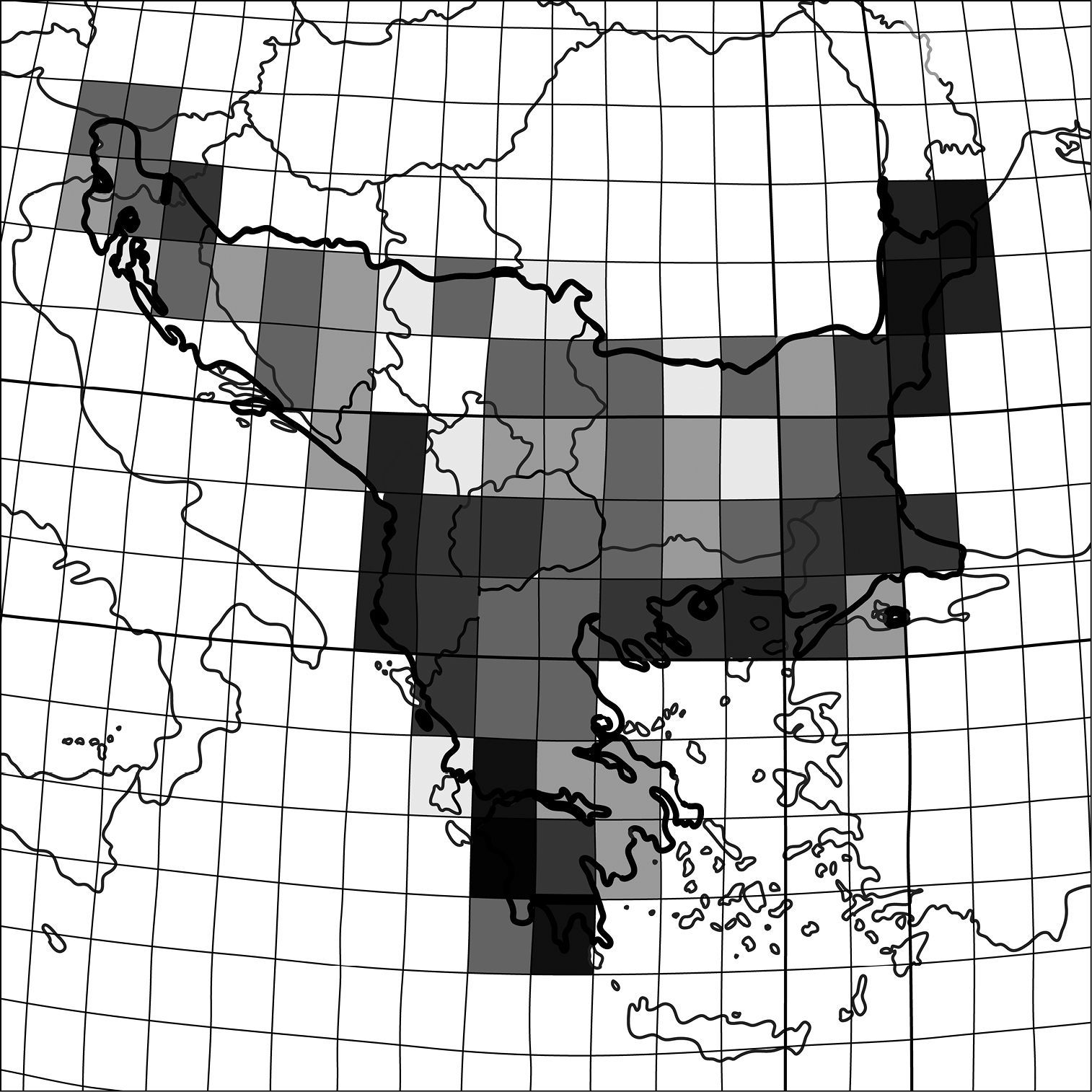
Species richness of tiger beetles within the Balkan Peninsula. The colour gradient indicates an enhanced diversity from one species (white square) to eight (black square).
Bray-Curtis analysis of similarities among tiger beetle faunas from different Balkan geographical regions shows the presence of three main clades (Fig. 4). The Dinaric, Tracio-Macedonian and Balkanic regions group mainly mountain areas, with lowlands only as very small parts, and covers a great part of Udvardy’s (1975) Balkan Highlands. The Danubian plain and North-Dobroudzha regions compose the second group, mentioned in biogeographic studies as the Pontian Steppe and north-eastern part of the Balkan Highlands. Clearly different is the Pindus area, which covers a large area of the Mediterranean Sclerophyl province. The Jaccard’s similarity index for Mediterranean Sclerophyl – Pontian Steppe was 53%, for Mediterranean Sclerophyl – Balkan Highland was 47%, and for Pontian Steppe – Balkan Highland was 38% (Fig. 4).
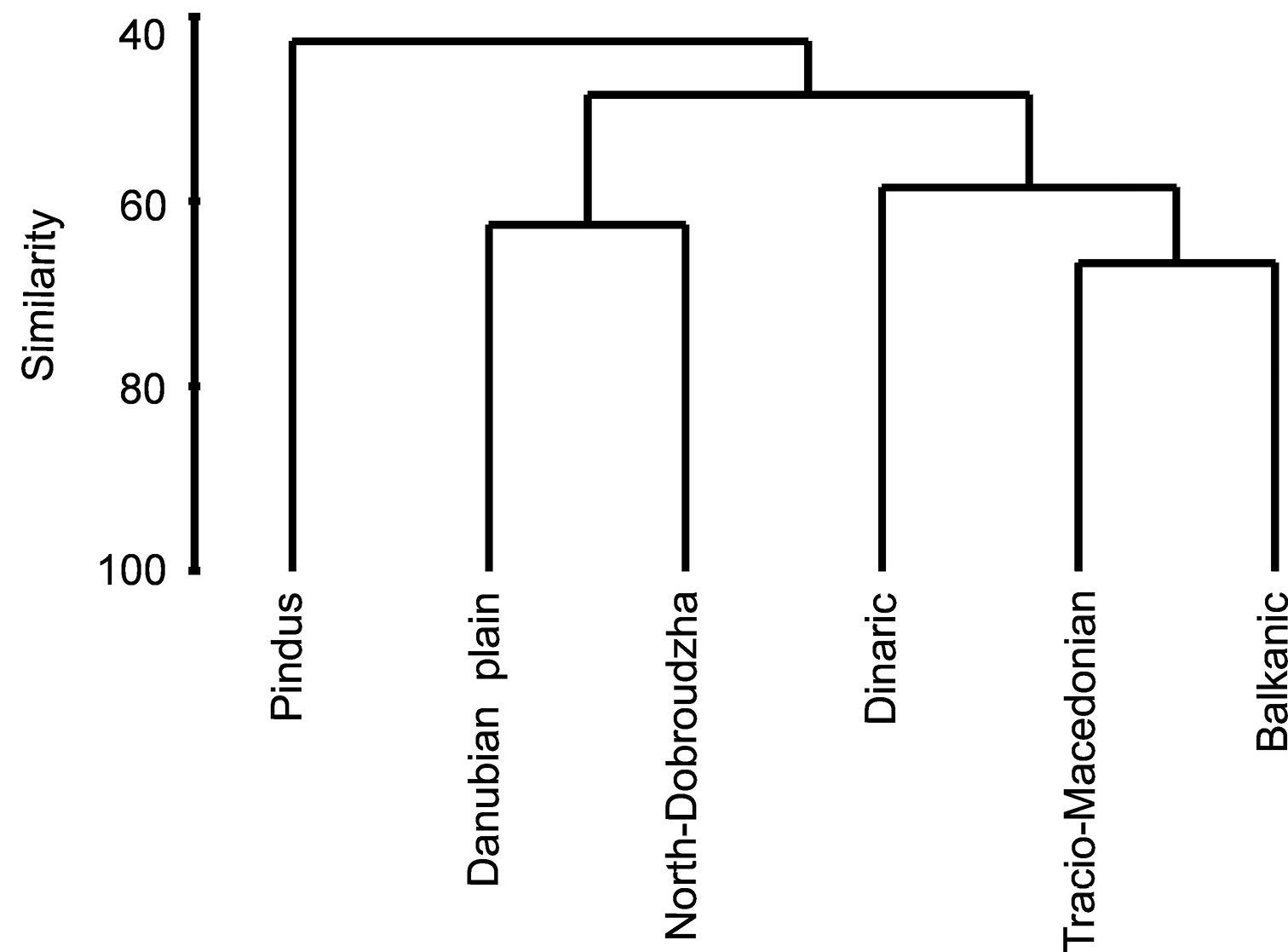
Similarities among tiger beetle faunas inhabiting regions of the Balkan Peninsula (Bray-Curtis similarity index for presence/absence data).
The most eurytopic species are Calomera littoralis nemoralis and Calomera aulica aulica (Table 3), occupying four and three habitats respectively. Ten species have been found occurring in only one type of habitat, including three Cephalota species in saltmarshes, three species restricted to sandy sea coasts (Calomera concolor, Cylindera contorta, Cylindera trisignata), four to river banks (Calomera fischeri, Cicindela sahlbergii albanica, Cylindera soluta pannonica, Cylindera arenaria viennensis), and one – Cicindela sylvatica, to forested sandy areas. Another five species were noted as occurring only in two types of habitat. Among all these tiger beetles, fifteen species (79% of the fauna) can be classified as coastal and riverine taxa, occurring in habitats adjacent to water, such as sea coasts, salt marshes (including lagoons and estuaries), and banks of rivers and freshwater lakes.
Tiger beetles of the Balkan Peninsula and their ecological distribution: 1 salt marshes 2 sandy sea beaches 3 banks of rivers 4 banks of lakes 5 forest roads 6 mountain and highland pastures 7 flat coastal rocks (based on literature data and personal observations).
| No. | Species | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|---|
| 1 | Calomera aulica aulica | + | + | + | ||||
| 2 | Calomera concolor concolor | + | ||||||
| 3 | Calomera fischeri fischeri | + | ||||||
| 4 | Calomera littoralis nemoralis | + | + | + | + | |||
| 5 | Cephalota (Cephalota) turcica | + | ||||||
| 6 | Cephalota (Taenidia) chiloleuca | + | ||||||
| 7 | Cephalota (Taenidia) circumdata circumdata | + | ||||||
| 8 | Cephalota (Taenidia) elegans stigmatophora | + | + | |||||
| 9 | Cicindela (Cicindela) campestris | + | + | |||||
| 10 | Cicindela (Cicindela) hybrida | + | + | |||||
| 11 | Cicindela (Cicindela) monticola albanica | + | ||||||
| 12 | Cicindela (Cicindela) soluta pannonica | + | ||||||
| 13 | Cicindela (Cicindela) sylvatica | + | ||||||
| 14 | Cicindela (Cicindela) sylvicola | + | + | |||||
| 15 | Cylindera (Cylindera) germanica | + | + | |||||
| 16 | Cylindera (Eugrapha) arenaria viennensis | + | ||||||
| 17 | Cylindera (Eugrapha) contorta contorta | + | ||||||
| 18 | Cylindera (Eugrapha) trisignata | + | + | |||||
| 19 | Myriochila (Myriochila) melancholica melancholica | + | + | |||||
| Total | 9 | 5 | 8 | 1 | 3 | 4 | 1 | |
Compared to the area size of other regions of Europe,
the diversity of tiger beetles of the Balkan Peninsula is high and
constitutes about 41% of all European tiger beetle species. This result
confirms an important role of the Balkans as a biodiversity hotspot
noted earlier for many other groups of organisms (
The Balkan Peninsula is inhabited by a mixed tiger
beetle fauna with representatives of 19 species belonging to 14
different groups according to their geographical origin (Table 2).
Such a mosaic of faunal elements clearly suggests an important role of
the Balkan Peninsula as a natural geographic „bridge” between Europe and
Asia Minor for this group in the past. Similar patterns have been noted
also among other groups of insects (
A high level of landscape heterogeneity also helps in
explaining the general distribution pattern of tiger beetle species
within the Balkan Peninsula and their higher species richness in the
lowlands. Sandy habitats preferred both by larvae and adult are more
diverse at sea coasts than those found in mountain areas. This patterns
for Balkan tiger beetles is similar to that reported from other regions
of the Mediterranean area (

Balkan tiger beetle species: A Calomera littoralis nemoralis B Calomera fischeri fischeri C Cephalota chiloleuca D Cephalota chiloleuca circumdata E Cicindela sylvicola F Cicindela campestris oliviera G Cylindera trisignata hellenica H Myriochila melancholica melancholica.
The narrow specialization to habitat type recorded
for most of the Balkan tiger beetle species is similar to that in tiger
beetles occurring in other regions of the world, both for adults and
larvae. For example, of the 151 species noted on the Indian
subcontinent by
Such narrow specialization to habitat/microhabitat types among tiger beetle species is explained by physiological (
Most Balkan tiger beetles occupy sandy habitats localized in lowlands, mainly on the sea coasts and in river deltas (Table 3). More than 90% of south-east European salt marshes are found in the Balkan Peninsula (
Hopefully the plight of these tiger beetles will help focus the attention of biologists, ecologists, and nature conservationists on the Balkan Peninsula as an important European hotspot area for conserving biodiversity of the European fauna.
I would like to express my thanks to Michał Grabowski who first focused my attention on the Balkan tiger beetle fauna. Piotr Jóźwiak prepared maps used in this paper (both from the University of Łódź, Łódź, Poland). Finally thanks are due to two anonymous reviewers for their valuable remarks and language corrections. The research was partly financially supported by SYNTHESYS Project AT-TAF-418.