





(C) 2012 Jukka Salmela. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
All available type material of Tipula stackelbergi Alexander, Tipula usuriensis Alexander and Tipula subpruinosa Mannheims were examined. Tipula (Yamatotipula) stackelbergi stat. rev. is elevated from a subspecies of Tipula (Yamatotipula) pruinosa Wiedemann to a valid species. Two new synonyms are proposed: Tipula usuriensis syn. n. proved to be a junior synonym of. Tipula (Yamatotipula) pruinosa and Tipula subpruinosa syn. n. a junior synonym of Tipula (Yamatotipula) freyana Lackschewitz. Tipula (Yamatotipula) stackelbergi is redescribed, male and female terminalia of Tipula (Yamatotipula) pruinosa are illustrated and discussed. Female terminalia of Tipula (Yamatotipula) freyana are described and illustrated for the first time. A key to both sexes of Tipula (Yamatotipula) stackelbergi and Tipula (Yamatotipula) pruinosa, and a key to females of Tipula (Yamatotipula) chonsaniana, Tipula (Yamatotipula) freyana and Tipula (Yamatotipula) moesta are provided. Subspecies are not uncommon among crane flies, but their ranges and traits are poorly known. An interdisciplinary approach (genetics, ecology, taxonomy) is suggested if subspecific ranks are to be used in tipuloid systematics.
Palaearctic region, crane flies, Tipula, new synonyms
The description of Tipula stackelbergi (Diptera, Tipulidae) was based on male holotype collected from Russian East Siberia (
Subspecies are traditionally held as geographically separate and genetically distinct populations within the species’ range, permitting gene flow in the area of contact (
In crane flies (Diptera, Tipuloidea) subspecific ranks are not uncommon. For example, out of 493 and 168 Palaearctic Tipulidae taxa described by C.P. Alexander (1889–1981) and E.N. Savchenko (1909–1994), respectively, 24 and 26 taxa are currently ranked as subspecies (data from
In this article I present the results of an examination of all available type material of Tipula stackelbergi, Tipula usuriensis and Tipula subpruinosa. I propose changes to the nomenclature of these species and I also review the morphology of Tipula pruinosa and Tipula stackelbergi, with an emphasis on male and female genitalia. In addition, female genitalia of Tipula (Yamatotipula) freyana are illustrated and a key to Tipula (Yamatotipula) freyana and females of Tipula (Yamatotipula) chonsaniana and Tipula (Yamatotipula) moesta are provided.
Material and methodsThe morphological terminology used here mainly follows
Layer photos were taken using an Olympus SZX16 stereomicroscope attached to an Olympus E520 digital camera. Digital photos were captured using the programmes Deep Focus 3.1 and Quick PHOTO CAMERA 2.3. Layer photos were finally combined with the program Combine ZP.
http://species-id.net/wiki/Tipula_stackelbergi
Figs 1, 2, 3e, 6a, c, dHolotype of Tipula stackelbergi: male, pinned specimen (ZISP). “Tigrovaja, Suchan./ rn.Uss.kr. 16.VI/ Stackelberg. 927” (white label, partly hand written, in Cyrillic letters). “81” (white label, handwritten). “HOLOTYPE/ Tipula stackelbergi/ C.P. Alexander” (red label, partly handwritten) (Fig. 1a).
With except of the male hypopygium, the holotype specimen is in rather good condition (Figs 1b, c). All legs are detached from the specimen, but four legs are glued to the pin below the specimen. Tips of wings are broken. Right antenna is broken, only scape and pedicel are left; left antenna has seven flagellomeres. Tip of abdomen is broken; apparently hypopygium is mounted on a celluloid strip, which is attached on a pin. The surface of this strip is heavily cracked, and the structure of the hypopygium cannot be examined.
Russia, Vladivostok, Nekrutenko leg, 2.VI. 1957, 1 male, 1 female (ZISP).
Male. Head yellowish brown, with grey pruinosity. Rostrum yellowish, nasus distinct, bearing numerous light hairs. Palpi brown. Scape yellowish, elongate, length 387–450 μm, width 126–131 μm (n=2). Pedicel yellowish, globular, length 147 μm, width 139 μm (n=1). Flagellomere 1 yellowish brown, length 486 μm, width 91 μm (n=1). Flagellomere 2 length 464 μm, width 79 μm (n=1). Flagellomeres bear erect short hairs, giving silvery appearance. Flagellomeres 2–7 elongate, brown, with dark verticils (Fig. 1c).
Prescutum with four brown stripes (Fig. 1e). Pronotum, prescutum, scutum, anepisternum, katepisternum and meron brownish, with grey pruinosity. Scutellum, anepimeron and laterotergite yellowish. Anterior part of mediotergite yellowish, more brownish in posterior part, having two weak longitudinal brown stripes. Coxa 1 brown. Anterior part of coxa 2 brown, posterior part yellow. Coxa 3 yellow. Femorae yellowish brown, darkening toward tarsi. Wings without markings, pterostigma brown (Fig. 1f). Wing length 13.8 mm (n=1). Halter yellowish.
Abdominal tergites yellowish brown, slightly darkening toward tip of abdomen. 9th tergite with two median projections, densely covered by dark bristles. Lateral corners of 9th tergite glabrous, pointed (Fig. 2a). 9th sternite with median incision, bearing two fleshy and hairy outgrowths in the margin of the incision. Outer gonostylus worm-like, apical half covered by light hairs (Fig. 2b). Inner gonostylus elongate (Figs 2b, c, 3e); beak rounded, with ten stout apical bristles and four subapical weaker bristles; central ridge with few weak bristles along its length; lower beak roundish, not angular. Posterior immovable apodeme of sperm pump almost straight (Fig. 2d). Distal end of compressor apodeme of sperm pump club-shaped, roundish (Fig. 2f). Aedeagal guide as in Fig. 2e.
Female. In general similar to male. Scutellum brown, abdominal tergites brown. Wing length 18.4 mm (n=1). Female terminalia as in Fig. 6a. Basal part of hypogynial valves with dense black setae. Proximal ends of valves roundish, tapering toward bases (Fig. 6c). Genital fork of vaginal apodeme brown, rather narrow in its whole length (Fig. 6d). Dorsal view of vaginal apodeme as in Fig. 6d.

Tipula (Yamatotipula) stackelbergi Alexander a Label of the holotype b Holotype male, habitus, lateral view. Yellowish celluloid board is attached below the specimen; most probably C.P. Alexander dissected hypopygium on this board. The surface of the board is heavily cracked, no details of the hypopygium are discernible c Thorax and head, holotype, lateral view d Right wing, holotype e Thorax and head, holotype, dorsal view f Male (Russia, Vladivostok), habitus, lateral view. Scale bars: b, f 2 mm; c & d 1 mm; e 0.5 mm.
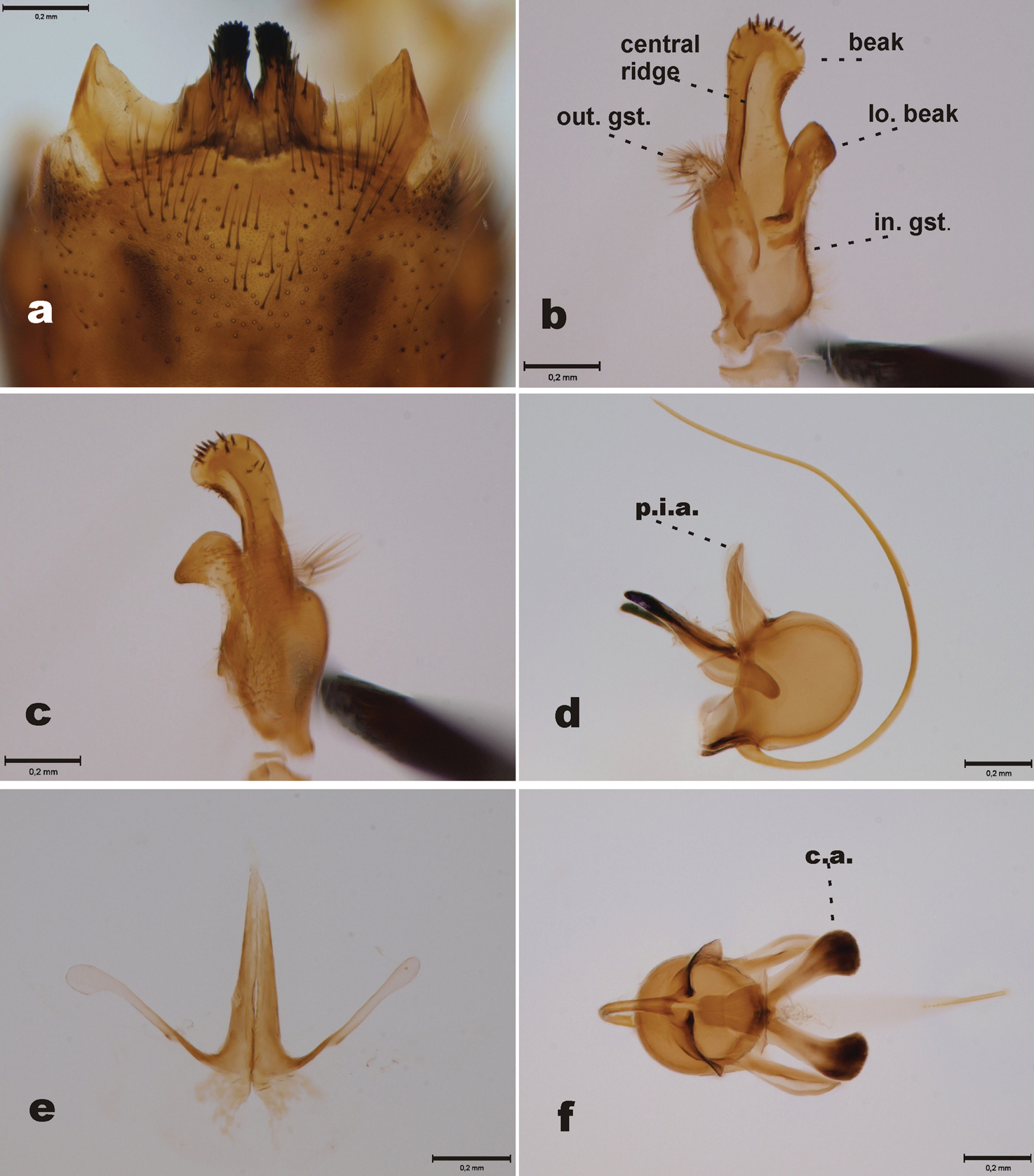
Tipula (Yamatotipula) stackelbergi Alexander, male (Russia, Vladivostok) a 9th tergite, dorsal view b Outer and inner gonostylus, posterior view; abbreviations: out. gst.= outer gonostylus, in. gst. = inner gonostylus, lo. beak = lower beak c Inner gonostulys, anterior view d Sperm pump, lateral view; abbreviation: p.i.a. = posterior immovable apodeme e Aedeagal guide, dorsal view f Sperm pump, ventral view; abbreviation: c.a. = compressor apodeme. Scale bars: 0.2 mm.

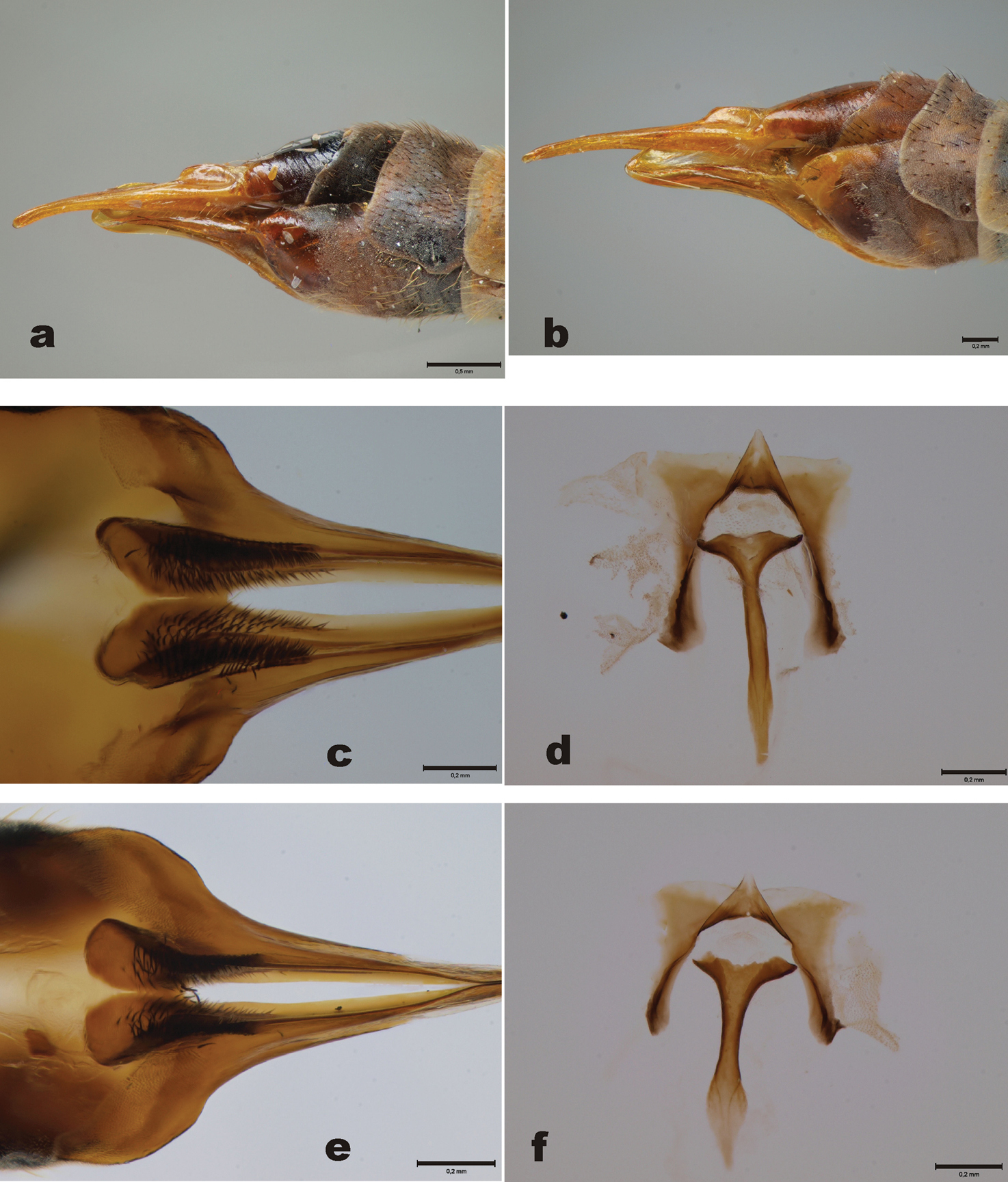
Tipula usuriensis Alexander (=syn. of Tipula (Yamatotipula) pruinosa Wiedemann), holotype male a Label of the holotype b Slide mounted wing c Habitus, lateral view d Sperm pump (lateral view) and aedeagal guide (dorso-lateral view). Scale bars: c 1mm; d 0.5 mm.
http://species-id.net/wiki/Tipula_pruinosa
Figs 3a–d, 3f–g, 4, 6b, e, fHolotype of Tipula usuriensis: male, pinned specimen (USNM). “Kudia River/Amagu Siberia/Cockerell/July 1923” (white label, printed). “HOLOTYPE /Tipula/ usuriensis/ C.P. Alexander” (red label, partly handwritten). Slide, permanently mounted wing. “Tipula usuriensis Alex./ ♀ Siberia, Amagu, / Kudia River/ July 1923, (T.D.A. Cockerell) / The Alexander Collection of Crane-Flies/HOLOTYPE 2967” (white label, partly handwritten). (Figs 3a, b). The holotype specimen of Tipula usuriensis is in quite bad condition (Fig. 3c). Half of the abdomen (distal part) and four legs are glued to a card. One wing (length 14.0 mm) is slide mounted and one wing is glued to a white card, one leg is also glued to the same card. Scape, pedicel and three flagellomeres of antennae are present. The holotype is also laterally flattened, perhaps due to compression of the freshly collected specimen. Hypopygium was detached by the author from the cardboard, macerated in KOH and finally preserved in glycerol in a microvial.
Finland. Savonia borealis: Kiuruvesi, Jynkänjärvi 63.5194°N; 26.6941°E, 13.VII. 2008, J. Salmela leg, 2 males (ZMUT); Ostrobottnia australis: Ilmajoki, Kivistönmäki 62.8492°N; 22.6623°E, 1 female, V.-M. Mukkala leg (PVM); Regio aboensis: Taivassalo, Orikvuori 60.6027°N; 21.6653°E, 26.VI. 2005 V.-M. Mukkala leg, 1 female (PVM); Regio aboensis: Turku, Piipanoja 60.4918°N; 22.3017°E, 22.VI. 2011 A. Teräs leg, 1 female, 4 males (ZMUT).
Male. 9th tergite (Fig. 4a) essentially similar to Tipula (Yamatotipula) stackelbergi. 9th tergite with two median projections, densely covered by dark bristles, lateral corners of the tergite glabrous, pointed (Fig. 4a). 9th sternite with median incision, bearing two fleshy and hairy outgrowths. Outer gonostylus worm-like, apical half covered by dark hairs (Figs 4b, c). Inner gonostylus elongate. Beak rounded, rather wide, resembling helmet (Figs 4b, c, 3f–g). Apical portion of beak bearing around 20 stout bristles, central ridge with numerous weak bristles, along the whole length of the ridge. Lower beak angular. Posterior immovable apodeme of sperm pump curved in lateral and ventral view (Figs 4d, f). Distal end of compressor apodeme of sperm pump truncated (Fig. 4f). Aedeagal guide as in Fig. 4e.
Female. Female terminalia as in Fig. 6b. Basal part of hypogynial valves with dense black setae, proximal ends of valves rounded, widest sub-basally, not tapering toward proximal end (Fig. 6e). Stalk of genital fork gradually widening toward caudal and proximal ends, being narrowest around midpoint (Fig. 6f). Dorsal view of vaginal apodeme as in Fig. 6f.
Geographical variation: The above mentioned description of male terminalia suites well to European specimens. The beak of the inner gonostylus among specimens from Asia is somewhat more i) sinuous, ii) slender and iii) with fewer stout bristles. Variation related to the geographical origin of the specimens is not detected in the structure of sperm pump. It is likely that Tipula (Yamatotipula) pruinosa sinapruinosa Yang & Yang, 1993 is similar to the holotype of Tipula usuriensis and to a male from Russia, Altay. These eastern Palaearctic specimens could perhaps be given a subspecific or infrasubspecific rank under Tipula (Yamatotipula) pruinosa. However, one widespread species with slight geographic variation in the coloration of head and abdomen (see

Tipula (Yamatotipula) pruinosa Wiedemann, male (Finland, Turku) a 9th tergite, dorsal view b Outer and inner gonostylus, posterior view c Inner gonostulys, anterior view d Sperm pump, lateral view e Aedeagal guide, dorsal view f Sperm pump, ventral view. Scale bars 0.2 mm.
http://species-id.net/wiki/Tipula_freyana
Figs 5, 7a–cHolotype of Tipula subpruinosa: female, pinned specimen (MZHF). “Suomi/ KemL./ Pallastunturit/ 1.8.1951/ leg J. Kaisila” (white label, partly handwritten; backside: “Pyhäkuru” handwritten). “Tipula (Oreom.)/ stigma n. sp./ Mannheims det. 1953” (white label, partly handwritten). “Holotypus” (red label, printed). “Museum/ Helsinki/ Frey” (white label, handwritten). “Mus. Zool. H:fors/ Spec. typ. No 14227/ Tipula/ subpruinosa Mann.” (grayish label, partly handwritten) (Fig. 5a). Pyhäkuru is located in NW Finland, Muonio, Pallas-Yllästunturit National Park, rough coordinates of the type locality are 68.079°N; 24.083°E.
The holotype specimen is in good condition (Figs 5b, c, d). Left mid leg is missing, other legs are intact. Right wing has minor rupture proximal to the pterostigma, Costa is slightly damaged. Abdominal terminalia of the specimen were detached by me, macerated in KOH and later preserved in glycerol in a microvial. This microvial is attached to the same pin as the specimen. The name “stigma” has never been published, and it has most probably been a working title by Mannheims while compiling his first account of Finnish tipulids (
Paratype: female, pinned specimen (ZMUC). “Lpl Sorsele/ Vallnäs tr / 18.7.1925 / S. Gaunitz” (white-gray label, unclear hand writing) “ex coll./ Peder Nielsen” (white label, printed) “Tipula (Tipula) / subpruinosa n sp.) / Mannheims det 1953” (white label, partly handwritten) “Tipula (Tipula) / subpruinosa n sp.) / Mannheims det 1953” (white label, partly handwritten) “Paratypoid” (red label, printed). The paratype specimen is in rather bad condition. Left antenna has nine and right antenna ten segments. All legs are broken, remnants of two legs are glued to a card below the specimen.
Other material. Finland. Karelia borealis: Lieksa, Nurmespuro 63.4030°N; 28.1972°E, 19.VI.–14.VII. 2008, J. Salmela leg, 2 females (ZMUT); Lapponia kemensis pars occidentalis: Kittilä, Palovaara E 68.0054°N; 24.7736°E, 23.VI. 2009 J. Salmela leg, 1 female (ZMUT); Lapponia enontekiensis: Enontekiö, Tarvantovaara, Pahtavaara SE 68.6518°N; 22.5909°E, 11.VI.–19.VII. 2009, J. Salmela leg, 2 males, 1 female.

Tipula subpruinosa Mannheims (=syn. of Tipula (Yamatotipula) freyana Lackschewitz), holotype female a Label b Habitus, lateral view c Left wing d Thorax and head, dorsal view. Scale bars: 1 mm.
Female terminalia. Tipula (Yamatotipula) stackelbergi Alexander (Russia, Vladivostok) a Female cerci, lateral view, pinned specimen c Hypogynial valves, dorsal view d Vaginal apodeme and genital fork, dorsal view. Tipula (Yamatotipula) pruinosa Wiedemann (Finland, Turku) b female cerci, lateral view, pinned specimen e Hypogynial valves, dorsal view f Vaginal apodeme and genital fork, dorsal view. Scale bars: a 0.5 mm; b, c, d, e, f 0.2 mm.
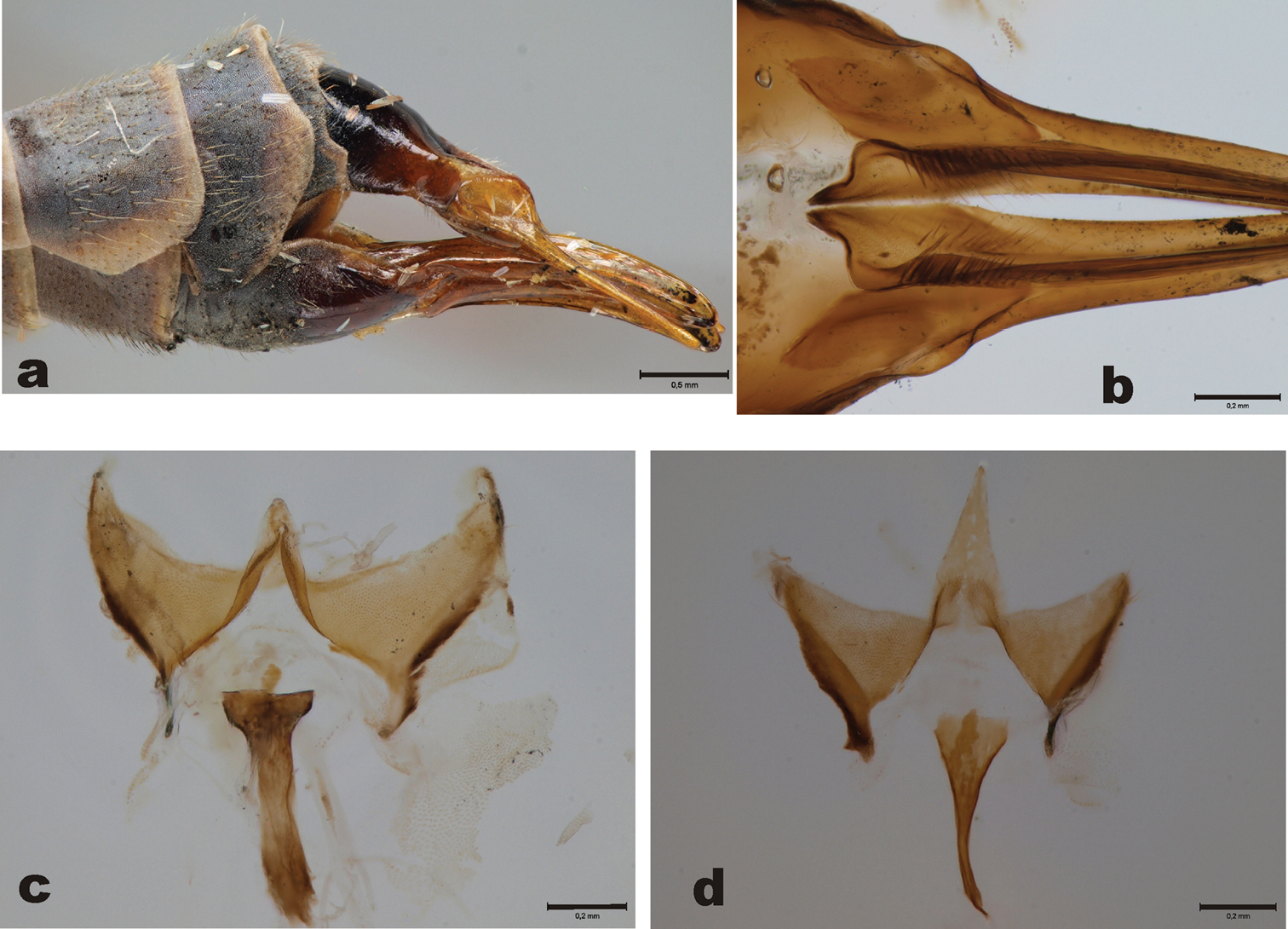
Female terminalia. Tipula (Yamatotipula) freyana Lackschewitz (holotype of Tipula subpruinosa Mannheims (Finland, Pallastunturit) a Female cerci, lateral view, pinned specimen b Hypogynial valves, dorsal view c Vaginal apodeme and genital fork, dorsal view. Tipula (Yamatotipula) chonsaniana Alexander (Finland, Taivalkoski) d Vaginal apodeme and genital fork, dorsal view.
Female terminalia as in Fig. 7a. Basal part of hypogynial valves with modest setosity, proximal ends of valves pointed (Fig. 7b). Genital fork of vaginal apodeme dark brown, slightly sinuous in lateral view. Dorsal view of vaginal apodeme and genital fork as in Fig. 7c.
Key to the Tipula (Yamatotipula) pruinosa and Tipula (Yamatotipula) stackelbergi
| 1 | Males | 2 |
| – | Females | 3 |
| 2 | Beak of inner gonostylus relatively wide, helmet-like, with numerous (ca. 20) stout bristles (Figs 4b, c). Sperm pump dark, posterior immovable apodeme curved in lateral and ventral view (Figs 4d, f) | Tipula (Yamatotipula) pruinosa |
| – | Beak of inner gonostylus rather narrow, with ca. 10 stout bristles (Figs 2 b, c). Sperm pump lighter, posterior immovable apodeme almost straight in lateral and ventral view (Figs 2d, f) | Tipula (Yamatotipula) stackelbergi |
| 3 | Basal part of hypogynial valves widest sub-basally, not tapering toward base (Fig. 6f). Stalk of genital fork gradually widening toward caudal and proximal ends, being narrowest around midpoint (Fig. 6g) | Tipula (Yamatotipula) pruinosa |
| – | Basal part of hypogynial valves roundish, tapering toward base (Fig. 6d). Genital fork rather narrow in its whole length (Fig. 6e) | Tipula (Yamatotipula) stackelbergi |
| 1 | Body coloration dark; scape, pedicel and 1st flagellomere dark brown | Tipula (Yamatotipula) moesta |
| – | Body coloration lighter; scape, pedicel and 1st flagellomere yellowish | 2 |
| 2 | Stalk (proximal 2/3) of genital fork very narrow, needle-like (Fig. 7d) | Tipula (Yamatotipula) chonsaniana |
| – | Stalk (proximal 2/3) of genital fork wider, as in Fig. 7c | Tipula (Yamatotipula) freyana |
In the present paper I suggest three changes to the nomenclature of Palaearctic Tipulidae: i) Tipula (Yamatotipula) stackelbergi is a valid species, not a subspecies of Tipula (Yamatotipula) pruinosa ii) Tipula usuriensis is neither a valid species nor a synonym of Tipula (Yamatotipula) stackelbergi, it is instead a junior synonym of Tipula (Yamatotipula) pruinosa and iii) Tipula subpruinosa is not a synonym of Tipula (Yamatotipula) stackelbergi, it is a junior synonym of Tipula (Yamatotipula) freyana. It remains questionable whether Tipula (Yamatotipula) pruinosa sinapruinosa is a valid subspecies. Based on the original description (
Tipula (Yamatotipula) pruinosa and Tipula (Yamatotipula) stackelbergi are closely related but valid species. The species pair is well separated due to the differences in male genitalia (see the key to the species), but less so regarding female genitalia. More females of Tipula (Yamatotipula) stackelbergi should be studied in order to firmly validate the diagnostic differences presented here. Tipula (Yamatotipula) stackelbergi is a very rarely collected species, known only from East Siberia and the Russian Far East (
Tipula subpruinosa, described from Finland and Sweden, was thought to be a synonym of Tipula (Yamatotipula) stackelbergi (
Compared to Tipula (Yamatotipula) stackelbergi and Tipula (Yamatotipula) pruinosa, Tipula (Yamatotipula) freyana is phylogenetically rather distant to these two species, being instead close to Tipula (Yamatotipula) moesta Riedel and Tipula (Yamatotipula) chonsaniana Alexander (e.g.
I am indebted to the following persons for information and/or the loan of specimens: Nikolai Paramonov (St. Petersburg), Pekka Vilkamaa (Helsinki), Jon Gelhaus (Philadelphia), David G. Furth, Patricia Gentili-Poole (Washington), Thomas Pape (Copenhagen), Valentin Pilipenko (Moscow) and Veli-Matti Mukkala (Kaarina). Julia Kuosmanen and Liisa Puhakka (Turku) are thanked for translations of Russian texts. Constructive comments by Fenja Brodo (Ottawa) and John Kramer (Leicester) improved the manuscript.