





(C) 2011 Jan Szyszko. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Possibilities of the assessment of a landscape with the use of succession development stages, monitored with the value of the Mean Individual Biomass (MIB) of carabid beetles and the occurrence of bird species are discussed on the basis of an example from Poland. Higher variability of the MIB value in space signifies a greater biodiversity. Apart from the variability of MIB, it is suggested to adopt the occurrence of the following animals as indicators, (in the order of importance), representing underlying valuable landscapes: black stork, lesser spotted eagle, white-tailed eagle, wolf, crane and white stork. The higher number of these species and their greater density indicate a higher value of the landscape for biodiversity and ecosystem services, especially carbon sequestration. All these indicators may be useful to assess measures for sustainable land use.
Carabidae, indicators, landscape evaluation, Mean Individual Biomass (MIB)
As part of the ongoing discussion on the sustainable use of landscapes and ecosystem services we give here an overview on the relationship between biodiversity and carbon sequestration in forests as revealed by a long-term study in north-western Poland with particular reference to carabid beetle diversity.
From the very beginning of life, environmental resources
on Earth have been shaped by natural succession processes, influenced
by climate, and by various disturbances such as orogenic movements,
windfall, floods and fires. Depending on latitude and the geological
base, these disturbance factors vary among various places on the globe.
The lack of catastrophes entails the buildup of organic substances,
which can be measured simply with the content of carbon. The content of
carbon in a single hectare of natural forest of the moderate climatic
zone shaped by succession processes for thousands of years, exceeds 350
tons/ha with upper limits estimated between 500 and 700 tons/ha (
In such old forests (old ecosystems created by nature),
ecosystems have a specific composition of plant, animal and fungal
species. Species that are linked to old trees, old decaying wood, and
well developed soil, occur here (Fig. 1).
In such forests in the European geographic zone the full range of
carabid species with a narrow geographic arrangement, low dispersal
power and large body size occurs. The Mean Individual Biomass (MIB) of
the carabids beetles might exceed 350 mg in such forests (

A natural forest with a carbon content within the limit of 350 tons per ha and a mean individual biomass (MIB) for carabid beetles exceeding 350 mg and with species characteristic for that environmental system. Lucanus cervus – a species linked to old decaying oak wood (top right), Cerambyx cerdo – a species linked to living old oaks (top left), Boletus erythropus – a mycorrhizal species occurring in soils with a historically well developed soil profile (bottom right), Carabus intricatus – a species occurring in old forest environmental systems with an easily decomposing duff (bottom left).

Succession of carbon content (left y-axis and black
line) and species diversity (right y-axis and striped line) in a stand
after the destruction of the trees by fire. After the fire, the carbon
content is low with Cicindela hybrida and the sticky bun (Suillus luteus) as characteristic species (left photographs). In 100 years old stands the carbon content is high with Carabus intricatus and the dotted stem bolete Boletus erythropus as characteristic species (right photographs) (from
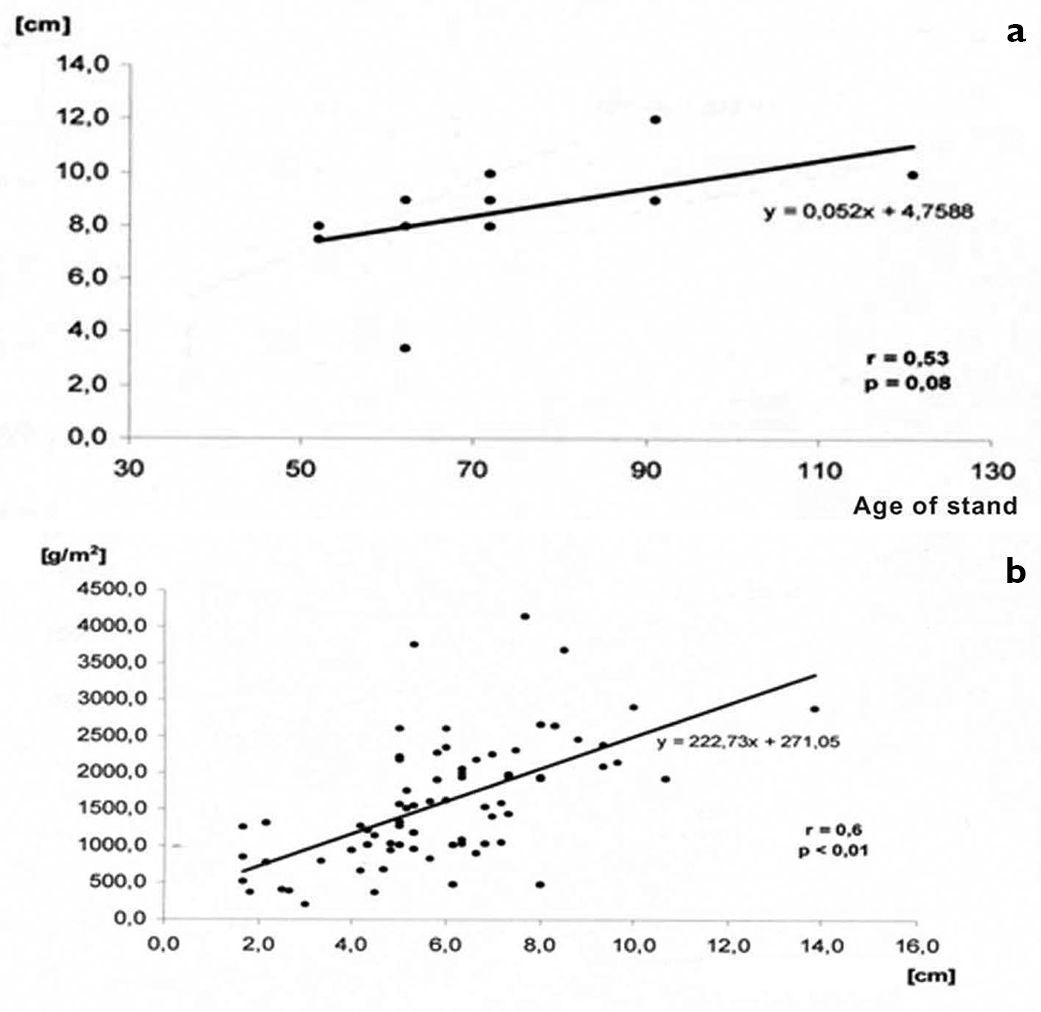
a Relationship between the age of a forest stand and thickness of the litter layer b relationship between the thickness of the litter layer and the weight of carbon per 1 sq. m (from
Humans play a similar role in shaping ecosystems and
landscapes. Ecological catastrophes can destroy the effects of our
economic activities and constitute a danger our its safety. This is why
humans manage the environment by trying to reduce floods, fires and
windfalls. However, to preserve the full range of biodiversity, humans
replace the forces of nature and mimic its destructive role by active
management. This can clearly be illustrated for forests in Poland. If it
was not for the interference with natural succession and the reduction
of the carbon content of up to a few dozen tons per ha (Fig. 4a) due to clearcuts, no pine cultivations could have emerged in forest areas. Consequently, bird species such as the nightjar (Camprimulgus europaeus) and woodlark (Lullula arborea)
would have had no place to nest and the majority of birds of prey
nesting in old trees would not have been able to hunt. Carabid beetles
like Carabus nitens, Bembidion nigricorne, Pterostichus lepidus, Calathus erratus, Masoreus wetterhallii and Harpalus rufitarsis (resulting in a MIB value of about 50 mg) would not have occurred in this region (

The occurrence of characteristic birds, carabid beetles and fungi as well as the structure of the carbon content in tons per ha in a forest stand, litter and mineral soil up to 10 cm in depth in a: a 10 year old pine stand with a MIB value of about 50 mg, created after the clear-cut of a timber pine stand (more than 100 years old) b ca. 60 year old pine stand with a MIB value about 250 mg. c ca. 80 year old beech stand created from the undergrowth after the clear-cut of a pine stand with a MIB value of about 350 mg. In all graphs the annual accumulation of carbon in that stand, the value of such accumulation and the value of the entire carbon content (forest stand + litter + mineral soil) are expressed in carbon dioxide at the prices of the European Emission Trade System on 15.08.2008 (all graphs from Szyszko 2007).
Data on different forests presented above suggest that a
greater spatial differentiation of the carbon content in environmental
systems, or in other words a greater differentiation of successional
stages as measured using the MIB value of carabid coenoses, entails
greater biodiversity (Fig. 5,
Landscape quality includes biodiversity (highest in mosaic landscapes) and carbon storage (highest in peat bogs and old forests) and sequestration (highest in regenerating forests), as well as other ecosystem services. Sustainable land use is essential in maintaining or creating landscapes with high natural qualities. There is a need to assess measures of sustainable land use, and various animals mentioned in this paper, including carabid beetles and their MIB values, may serve as important indicators for this.

Examples of several habitats with different carbon
contents and different characteristic carabid species, building a
heterogenous landscape. Top left – a natural forest with a carbon
content of 350 ton per ha with Carabus coriaceus, top right – arable land with a carbon content 20 tons per ha with Cicindela campestris, in the middle – a peatbog with a very high content of carbon per ha with Panagaeus bipustulatus, bottom left – a clear-cut with a carbon content of ca. 90 tons per ha with Harpalus rufitarsis, Bottom right – a timber stand with the carbon content 124 tons with Carabus nemoralis (from

Landscape species in a heterogenous landscape. The lesser spotted eagle (Aquila pomarina) nests in old trees in natural and cultivated forests, and hunts in wastelands. The crane (Grus grus) nests in peat bogs and hunts in wastelands. The kestrel (Falco tinnunculus) nests in old trees in natural and cultivated forests and hunts in clear-cut areas. The black stork (Ciconia nigra) nests in old trees in natural and cultivated forests and hunts in peat bogs (from
The authors wish to thank an anonymous reviewer for valuable comments on an earlier drafts of the manuscript. This paper is communication number 246 of the Laboratory of Evaluation and Assessment of Natural Resources, Warsaw University of Life Sciences, and the Association for Sustained Development of Poland.