





(C) 2010 Munetoshi Maruyama. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Three myrmecophilous genera of Aleocharinae (Staphylinidae) associated with Leptogenys distinguenda (Emery, 1887) and Leptogenys mutabilis (Smith, 1861) are reviewed with descriptions of new taxa: Maschwitzia Kistner, 1989, Togpelenys Kistner, 1989 and Witteia Maruyama & von Beeren, gen. n. (type species: Witteia dentilabrum Maruyama & von Beeren, sp. n.). The following new combinations are proposed: Zyras (s. lat.) aenictophila (Kistner, 1997), comb. n. (ex Maschwitzia), Zyras (s. lat.) dichthadiaphila (Kistner in Kistner et al., 2003), comb. n. (ex Maschwitzia), Maschwitzia derougemonti (Pace, 1984), comb. n. (ex Wroughtonilla Wasmann, 1899), Maschwitzia watanabei (Maruyama, 2004), comb. n. (ex Wroughtonilla), Maschwitzia dilatata (Pace, 2005), comb. n. (ex Wroughtonilla), Witteia borneensis (Pace, 1986), comb. n. (ex Wroughtonilla). These genera belong to the Wroughtonilla genus group of the tribe Lomechusini.
Myrmecophily, Maschwitzia, Togpelenys, Witteia gen. n., Wroughtonilla genus group, new species, new combination, Malaysia, new host record
The ant genus Leptogenys Roger, 1861 belongs to the subfamily Ponerinae. Some of its members show army ant-like behavior (
In the present article, we revise some of the rove beetle genera associated with Leptogenys ants. This first part of the series reviews the genera which are associated with Leptogenys distinguenda and Leptogenys mutabilis, including descriptions of some new taxa and some new combinations.
Materials and methodsThe rove beetles were collected in spring and autumn 2008
and 2009 in a well regenerated dipterocarp rainforest in the Field
Studies Centre of the University of Malaya in Ulu Gombak, Malaysia (03°19.4796N; 101°45.1630E, altitude 230 m) and near the Institute of Biodiversity in Bukit Rengit, Malaysia (03°35.779N; 102°10.814E, altitude 72 m). Nests of the nocturnal host ants were located during the night by back-tracking Leptogenys
raiding trails. Since all rove beetles follow the host ant migrations,
we detected them on these occasions and collected them with aspirators
(for further information see
The methods of dissecting and line drawings followed
Leptogenys distinguenda is sometimes treated as a subspecies of Leptogenys processionalis Jerdon, 1851 known from India (

Host ants. 1 Leptogenys distinguenda, lateral view 2 ditto, head 3 Leptogenys mutabilis, lateral view 4 ditto, head.
Fig. 5
Maschwitzia ulrichi Kistner, 1989.
This genus is closely allied to Witteia in general appearance, especially pronotal shape, but may easily be distinguished from it by having a generalized labrum, not strongly sclerotized and without projections; the simple mandibles, their inner edges not emarginate at middle; the straight lateral projections of the labial apodeme; the much smaller eyes; the shorter antennae; and the shorter legs.

Type species of the genera Maschwitzia, Togpelenys and Witteia 5 Maschwitzia ulrichi, dorsal habitus 6 Togpelenys gigantea, dorsal habitus 7 Witteia dentilabrum gen. et sp. n., dorsal habitus 8 ditto, head and pronotum, dorsal view.
Zyras (s. lat.) aenictophila (Kistner in
Zyras (s. lat.) dichthadiaphila (Kistner in
Trachydonia
The following four species are known in Maschwitzia.
Fig. 5
Ulu Gombak, Selangor, Malaysia.
Ulu Gombak (University Malaya Field Studies Centre, 03°19.479N; 101°45.170E, 220–250 m alt.), Selangor, Malaysia, VIII 2008, C. von Beeren from the colony of Leptogenys distinguenda (10); same data, but III 2009, C. von Beeren and V. Witte (12); same data, but VIII 2009, C. von Beeren (10); same data, but IX 2009, Y. Nakase (6); same data, but III 2010, C. von Beeren from the colony of Leptogenys mutabilis (2); Bukit Rengit, Pahang, Malaysia (03°35.779N; 102°10.814E, altitude 72 m): C. von Beeren and V. Witte (8).
Peninsular Malaysia.
Leptogenys distinguenda and Leptogenys mutabilis.
This species is closely similar to Maschwitzia watanabei in general appearance, but is distinguished from it by the pronotum being narrower around the posterior margin and the aedeagus being different in shape, especially with the apical part of the median lobe being strongly widened apically and not excavated paramerally.
Two specimens were collected for the first time from the end of a migration column of Leptogenys mutabilis (new host record).
Kalaw, Myanmar
Myanmar.
Unknown.
The aedeagal shape is clearly different from the other congeners in particular the parameral crest is larger and the apical lobe longer.
Only the holotype and one paratype are known. The original description by
Bolikhamsai (Borikhamxay), Laos.
Laos.
Unknown.
This species is closely similar to Maschwitzia ulrichi in general appearance, but is distinguished from it by the pronotum being wider around the posterior margin and by the different shape of the aedeagus, especially the apical part of the median lobe being less widened apically and largely excavated paramerally.
Only the holotype is known. The symbiotic host is probably Leptogenys distinguenda or a related species, although Leptogenys distinguenda is not recorded from Laos at present.
Umran, East Khasi Hills, Meghalaya, India.
Meghalaya, India.
Unknown.
This species is closely similar to Maschwitzia ulrichi in general appearance, but is distinguished from it by the pronotum being wider around the posterior margin and by the different shape of aedeagus, especially the apical part of the median lobe being less widened apically and largely excavated paramerally.
Only the holotype has been known. The original description by
Fig. 6
Togpelenys gigantea Kistner, 1989.
This genus is clearly distinguished from the other genera of Wroughtonilla group by the combination of the following character states: eyes extremely large; pronotum without superior marginal line of the pronotal hypomeron; pronotal disc quite convex, with a shallow and large longitudinal depression; pronotum and elytra covered with long, suberect macrosetae; and abdomen large, expanded, much wider than elytra.
Peninsular Malaysia.
Only the type species Togpelenys gigantea Kistner, 1989 has been known in the genus. Probably further species will be found from the regions around Peninsular Malaysia, e.g., Sumatra, Borneo and Java.
Fig. 6
Ulu Gombak, Selangor, Malaysia.
Bukit Rengit (03°35.779N; 102°10.814E; 72 m alt.), Pahang, Malaysia, III 2009, C. von Beeren and V. Witte (2 males).
Peninsular Malaysia.
Leptogenys distinguenda.
This species is easily distinguished from the other species of Wroughtonilla as well as its allied genera by the generic diagnosis.
Rare species, newly recorded from Pahang. In the type locality, Ulu Gombak, Selangor, no additional specimen has been collected despite more than 40 colonies having been examined in the last few years (von Beeren and Witte, personal observations; the type series were collected in 1982). We are not sure whether this is due to environmental changes in Ulu Gombak or simply due to rarity of this species.
urn:lsid:zoobank.org:act:B3D89EA2-4867-40F5-9817-E6E108D0BA2B
Figs 7–18Witteia dentilabrum sp. n.
Dedicated to Dr. Volker Witte for his contribution to the biology of Leptogenys ants and their symbionts. Gender, feminine.
This genus is similar to Maschwitzia Kistner, 1989 in body shape and punctation of body surface, but may easily be distinguished from it by the labrum being strongly sclerotized and with a pair of spines; the inner margins of the mandibles emarginate at middle; the lateral projections of the labial apodeme curved apically; the extremely large eyes; the longer antennae; and the longer legs.
Body (Fig. 7) elongate, flattened; surface of fore body (Fig. 8) weakly rugose, reticulated, somewhat matte.
Head (Figs 7–8) transverse, with eyes extremely large, somewhat shorter than head, with a round depression above; clypeus truncate apically. Labrum (Figs 8–9) strongly sclerotized, with a pair of projections laterally (Figs 9: arrow). Mandibles asymmetrical, strongly curved, each apex acutely pointed; inner margin of left mandible (Fig. 10) shallowly emarginate; that of right (Fig. 11) rather largely emarginated. Mentum (Fig. 12) trapezoidal, with several thick setae, with sparse pseudopores. Labium (Fig. 13) broad; prementum with a setal pore and 2 real pores near base, with several pseudopores around inner ridges; apodeme without median projection, with lateral projection curved apically; ligula long, each lobe with a large setulum and three small setula; labial palpus with segment I long and apically dilated; segment II half as long as I; segment III thin, parallel-sided, slightly shorter than II.
Pronotum (Figs 7–8) with disc well margined, slightly convex, with a narrow longitudinal groove medially and a pair of depressions postero-laterally. Mesocoxal cavity well margined; process of metaventrite narrow, pointed at apex.
Elytra (Fig. 7) apically widened, laterally with a pair of rather high carinae, that are slightly curved inwards.
Legs (Fig. 7) very long, thin; femora slightly narrowed apically near apex; tibiae somewhat widened around middle to basal 1/3, their bases constricted; tibiae very thin, filiform.
Abdomen (Fig. 7) fusiform, flattened, widest around apex of segment III; surface smooth, sparsely punctured, shining. Median lobe of aedeagus (Figs 16–17) with long and narrow apical part, with a small projection near base of apical part (Fig. 16: arrow). Paramere with apical lobe slightly widened apically.
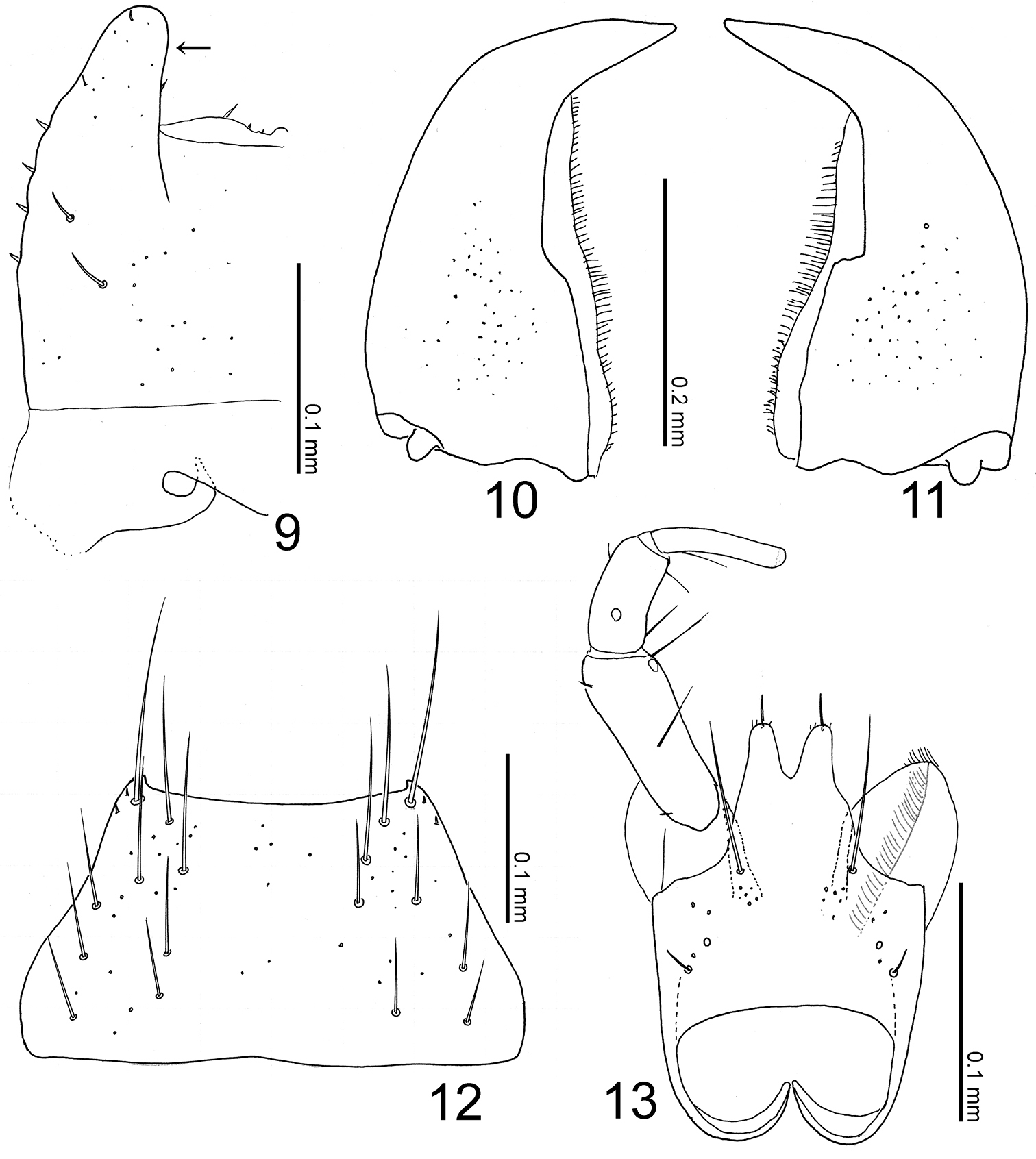
Mouthparts of Witteia dentilabrum gen. et sp. n. 9 labrum, left side, dorsal view 10 left mandible, dorsal view 11 right mandible, dorsal view 12 mentum, ventral view 13 labium, ventral view.
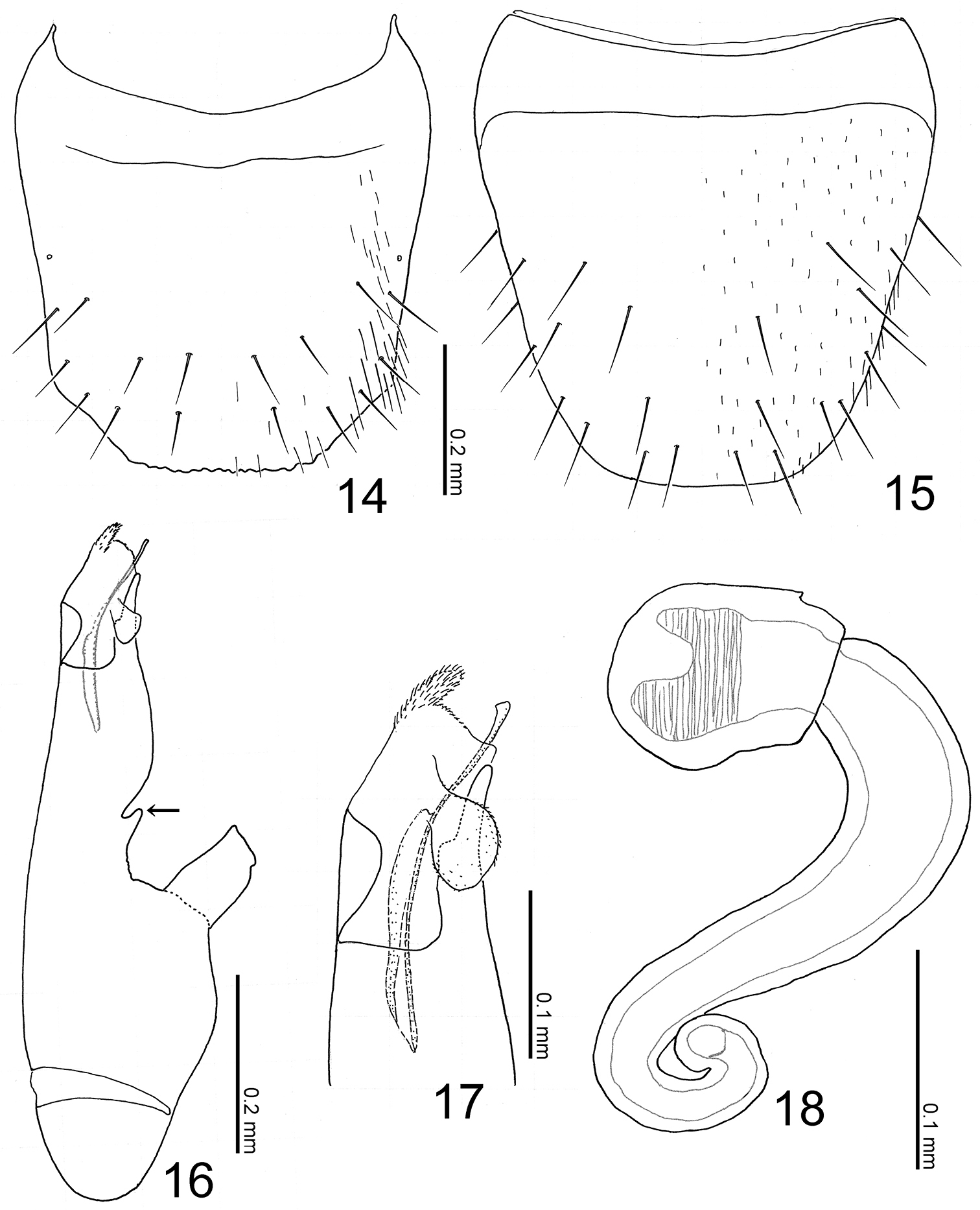
Terminalia of Witteia dentilabrum gen. et sp. n. 14 Male tergite VIII, dorsal view 15 male sternite VIII, ventral view 16 median lobe of aedeagus, lateral view 17 ditto, apical part 18 spermatheca.
urn:lsid:zoobank.org:act:EF84731E-0E8D-486B-859B-DBD89510FC8A
Figs 7–18Referring to the lateral projections on the labrum which is a unique character state in Witteia.
Holotype, male, Ulu Gombak (University Malaya Field Studies Centre, 03°19.479N; 101°45.170E, 220–250 m alt.), Selangor, Malaysia, III 2009, C. von Beeren and V. Witte, from a colony of Leptogenys distinguenda (KUM). Paratypes: same data as holotype (2 males, 1 female); same data, but VIII 2008, C. von Beeren (10); same data, but VIII 2008, C. von Beeren (8); same data, but IX 2009, Y. Nakase (2).
Ulu Gombak, Selangor, Malaysia.
Peninsular Malaysia.
Leptogenys distinguenda.
This species is closely similar in general appearance to Witteia borneensis (Pace, 1986), comb. n., from Sabah, Borneo, but is distinguished from it by the larger body and the smaller apical part of the spermatheca.
Body (Fig. 7) color reddish brown, but head completely black, mouthparts, legs, apex of abdomen lighter, medial areas of abdominal segments V and VI infuscate. Head (Figs 7–8) sparsely covered with setae; surface finely reticulated. Antennae (Figs 7–8) long, filiform; all segments longer than wide; segments III-X almost twice as long as wide; segment XI elongate. Pronotum (Figs 7–8) subquadrate, subparallel-sided, slightly wider than long (width/length = 1.11–1.18); surface moderately covered with minute setae, with some minute macrosetae laterally. Abdomen with anterior margins of sternites IV-VI produced medially; tergite VIII (Fig. 14) crenulate apically, with 8 macrosetae; sternite VIII (Fig. 15) rounded apically.
Male: sternite VIII with 11–12 macrosetae. Median lobe of aedeagus (Figs 16–17) with large parameral crest; apical part roundly convex paramerally; apical lobe slightly trilobed; copulatory piece with a short flagellum.
Female: sternite VIII with 8–9 macrosetae. Spermatheca (Fig. 18) with basal part dilated apically, coiled near base, curved near apex; apical part short.
Measurements: BL, ≈ 4.2–5.1; FBL, ≈ 1.8–2.0; HL, 0.660–0.738; HW, 0.887–0.988; AL, ≈ 2.8–3.1; PL, 0.806–0.950; PW, 0.725–0.800; HTL, 1.238–1.438.
Commonly found in Leptogenys distinguenda colonies together with Maschwitzia ulrichi, but less frequent than the latter species.
Pangi, Sabah, Malaysia.
This species is closely similar in general appearance to Witteia dentilabrum sp. n. from Selangor, Malaysia, but is distinguished from it by the smaller body (3.0 mm) and the larger apical part of the spermatheca.
Sabah, Borneo; Burma.
This species was described based on a single specimen from Sabah, Borneo. In the original description
The genera Maschwitzia, Togpelenys and Witteia are closely allied to Wroughtonilla (one species from India, Sri Lanka, Malaysia) and they belong to the Wroughtonilla genus-group (here proposed) of the subtribe Myrmedonina of the tribe Lomechusini, together with the genera Aenictonia Wasmann, 1900 (10 species from tropical Africa, one species from Thailand), Anommatochara Wasmann, 1915 (one species from tropical Africa), Leptogenoxenus, 1975 (one species from Philippines) and Neowroughtonilla, 1989 (one species from Malaysia). All members of this genus-group are associated with Leptogenys ants as far as known, except for species of Aenictonia and Anommatochara which are associated with Dorylus Fabricius, 1793 and/or Aenictus Shuckard, 1840 ants. The genera of this group share the following apomorphic character states: head with “neck”, a constricted postoccipital suture; pronotum with a longitudinal median groove; elytra with a pair of carinae laterally; and apical lobe of aedeagal median lobe elongate.
When Hlaváè and Janda (2009) described the genus Leptogenopapus (species from Papua New Guinea, associated with Leptogenys breviceps Viehmeyer, 1914), they stated that it is closely related to Leptogenoxenus. However, Leptogenopapus does not share the character states mentioned above. Because the type species Leptogenopapus mirabilis is in its general appearance extremely modified for myrmecomorphy, it is possible that the apomorphic character states in the Wroughtonilla group cited above have been secondarily lost or modified in this species. However, the aedeagal shape, which is normally not modified along with modification of external morphology to the myrmecophilous habitat, of Leptogenopapus mirabilis is very different from those of the Wroughtonilla group. Leptogenopapus does not belong to the Wroughtonilla genus-group.
Witteia is established as a new genus due to a strong autapomorphy, the presence of the projections on the labrum, by which it is clearly distinguished from Maschwitzia which is similar overall in general appearance. This character state is unique in the Lomechusini, possibly not present in any other aleocharine genera. Several other character states further distinguish Witteia from Maschwitzia (see Diagnosis of both the genera), though their polarities remain uncertain. We were reluctant to establish Witteia based on this single autapomorphy. However, considering generic concepts of the Wroughtonilla genus-group, which seem relatively narrow, the present establishment of the new genus seems reasonable at present. Phylogeny-based, ideally molecular phylogeny-based systematic revisions will be needed in the future.
Leptogenys distinguenda has been known as the only host of Maschwitzia ulrichi. However, it was collected also from colonies of Leptogenys mutabilis (new record above). This is the only confirmed example of Leptogenys-associated aleocharine species that parasitizes more than one species of Leptogenys ants. Leptogenys distinguenda and Leptogenys mutabilis are morphologically very similar which suggests their close phylogenetic relationships, and this probably allows Maschwitzia ulrichi to parasitize both species. In other myrmecophilous aleocharine species of Lomechusini, some of the Asian species of the genus Pella Stephens, 1835 have been known to host more than one ant species of Lasius spp. that are also very closely related, i.e., they belong to the same subgenus (
We thank Mr. Peter Hlaváč (Košice), Dr. Alfred F. Newton (Field Museum of Natural History, Chicago) and Dr. Volker Witte (Ludwig-Maximilians-Universität München) for their comments on the manuscript and Dr. Fuminori Ito for information on Leptogenys ants. This paper is supported by a Grant-in-Aid for Scientific Research from JSPS (Start-up 20870031) as well as a grant of the Deutsche Forschungsgesellschaft (Project WI 2646/3).