





(C) 2011 Carolina Cuezzo. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
A new genus and species of nasutitermitine termites are described and illustrated, based on soldier and worker characters. Sinqasapatermes gen. n., can be distinguished from all other nasutitermitine genera by its singular worker gut coiling and enteric valve characters: distal margin of the enteric valve not everted into the paunch but bending towards the ileum, that is, directed against the flow of food; enteric valve armature with one ring of six equal subtriangularly-shaped ridges, each ridge with short spines on the entire surface; enteric valve armature situatedonexternal face of cone, facing the internal ileum wall; enteric valve seating tri-lobed and separated from remaining portionof the paunch; paunch subdivided. Sinqasapatermes sachae sp. n., was collected on a tree in a very narrow flattened tunnel that was well concealed beneath lichens in a northern Peru rainforest (Arcadia, Loreto Province).
Taxonomy, Subulitermes-group , enteric valve configuration, gut coiling, Peruvian rainforest
The subfamily Nasutitermitinae is a monophyletic group, currently worldwide represented by 81 living genera and 575 living species (
Although Nasutitermitinae was never formally subdivided into minor taxonomic hierarchies, several authors (
Currently, there are 21 genera described as part of the Subulitermes-group, with 11 genera occurring in the Neotropical region (
In this study, an unusual termite sample collected from the rainforest of northern Peru is described and assigned to a new neotropical nasutitermitine genus and species, Sinqasapatermes sachae gen. n. and sp. n. The systematic position of the new genus is determined according to an evaluation of the internal morphology. Such characters provide enough evidence to place Sinqasapatermes gen. n., in the Subulitermes-group and also to distinguish it substantially from other known nasutitermitine genera within this group. The phylogenetic relationships among members of the Subulitermes-group remain unresolved (
The sample was subdivided and deposited in the Isoptera Collection of the Museo de Historia Natural, Universidad Nacional Mayor de San Marcos, Lima, Peru (MUSM), the National Museum of Natural History, Washington, DC, USA (USNM) and the Museu de Zoologia da Universidade de Sao Paulo, Brazil (MZUSP).
Morphometric characters used in this study and their correspondence with Roonwall’s system (
Description of the digestive tube morphology follows
Drawings were made with a camera lucida attached to a stereomicroscope.
Taxonomic treatmenturn:lsid:zoobank.org:act:15D04831-D8C0-4059-BC6D-75E77AAA7C46
Sinqasapatermes sachae sp. n.
The new genus is distinguished from other nasutitermitine genera by the unique combination of the following characteristics from the worker digestive tube: distal margin of the enteric valve (P2) not everted into the paunch (P3) but bending towards the ileum (P1), that is, directed against the flow of food; armed with one ring of six equal subtriangularly-shaped ridges, each ridge with short spines on entire surface (Fig. 14); enteric valve armature situatedonexternal face of cone, facing P1 internal wall (Fig. 15); enteric valve seating tri-lobed and separated from remaining portionof P3; P3 subdivided.
Imago. Unknown.
Soldier. Monomorphic. In dorsal view, head capsule with a constriction behind base of antenna. Maximum width behind constriction, at middle of posterior lobe. In profile, dorsal margin of head capsule nearly straight to base of nasus; weakly depressed at base of nasus. Nasus narrow, conical in dorsal view; about same length as head capsule; slanted slightly upward in profile. Mandibles vestigial, without points. Postclypeus moderately arcuate, in profile. Labrum wider than long, with rounded antero-lateral corners. Antenna with 11 articles. Pronotum shallowly saddle-shaped, anterior margin rounded, not emarginate. Tibial spurs 2:2:2.
Worker. Head capsule trapezoidal in dorsal view; maximum width of head capsule at base of mandibles. Fontanelle area depressed, situated at posterior third of head capsule.Postclypeus short and moderately inflated; median line weakly defined. Antenna with 12 articles. Pronotum shallowly saddle-shaped, anterior margin rounded, not emarginate. Tibial spurs 2:2:2.
Mandibles. Left mandible (Fig. 5) with apical tooth slightly as prominent as or slightly more prominent than M1+2, posterior margin of apical tooth weakly concave or truncate; right or acute angle between posterior margin of apical tooth and anterior margin of M1+2; posterior margin of M1+2 straight or slightly sinuous; third marginal tooth reduced to a vestigial node; molar tooth visible at V-shaped gap but apex hidden beneath molar prominence; molar prominence with very weakly-defined ridges. Right mandible (Figs 6–7) with apical tooth slightly more prominent than first marginal tooth, second marginal tooth not visible or vestigial (as a minute prominence); molar plate concave with very weakly defined ridges; basal notch well defined.
Digestive tube (Figs 8–15). Coiling gut in situ forming a short, broad mass. Crop (C) voluminous, partially visible in dorsal view; positioned to left halfof abdomen. Gizzard (G) with a strong musculature; well separated from crop; partially visible in ventral view. Mesenteron (M) passing through right side but not reaching medial line in ventral view. Mesenteric-proctodeal junction circular (mixed segment absent); visible in right lateral view. Malpighian tubules (TM) slightly dilated to form an ampulla at base, arranged in adjacent pairs with a common base at mesenteric-proctodeal junction on inner face. Ileum or first proctodeal segment (P1) tubular; shorter than mesenteron length. Enteric valve (P2) lying beneath P1, conical with distal margin not everted into P3 but bending towards P1, i.e., directed against the flow of food (Fig. 15); armed with one ring of six equal subtriangularly-shaped ridges, each ridge with short spines on entire surface (Fig. 14); enteric valve armature situatedonexternal face of cone, facing P1 internal wall (Fig. 15). P2 in same axis aspaunch (P3). Enteric valve seating, tri-lobed and separated from remaining portionof P3 by a constriction; another subdivision visible at P3 before protruding through mesenteric ring; distal part of P3 very prominent in dorsal view and joining colon (P4) on left side; isthmus conspicuous. Dorsal torsion well developed. P4 “U-turn” dilated. Distal colon tubular.
From Quechua, indigenous South American language, sinqa = nose and sapa = big, and Latin termes = termite, meaning termite with a big nose.
Among soldiers of neotropical genera, the long, narrow nasus and reduced number or absence of bristles on head capsule and thoracic nota are shared characters among species of Cyranotermes Araujo, Anhangatermes Constantino and Sinqasapatermes, but Cyranotermes and Anhangatermes species have antenna with 13 articles and a rounded head capsule. Soldiers of the latter two genera are also significantly larger than those of Sinqasapatermes. Agnathotermes Snyder and Paraconvexitermes Cancello & Noirot also have a conical nasus and antenna with 11 articles, but they differ from Sinqasapatermes by their chaetotaxy and head shape of the soldier.Soldiers of species of Angularitermes Emerson, Araujotermes Fontes, Atlantitermes Fontes, Coatitermes Fontes, Convexitermes Holmgren, Ereymatermes Constantino and Subulitermes Holmgren are genera that can be separated from Sinqasapatermes on the basis of different chaetotaxy arrangement and head capsule shape.
Among workers of neotropical genera, the third marginal tooth of left mandible and second marginal tooth of right mandible absent or vestigial are shared characters among species of Cyranotermes and Anhangatermes, however, molar area differs in Sinqasapatermes by having no ridges and being more concave. Reduced ridges on molar areas are found in Araujotermes, Atlantitermes, Coatitermes, Convexitermes, Ereymatermes, Paraconvexitermes, and Subulitermes, but none of these genera have lost the marginal teeth. The worker gut coiling and enteric valve features of Sinqasapatermes do not match any member of the Subulitermes-group (
urn:lsid:zoobank.org:act:86BBA560-790D-42D8-B3EF-FFB3864F534C
http://species-id.net/wiki/Sinqasapatermes_sachae
Figs 1–15Holotype soldier, in alcohol, separate in a microvial. Original typewritten label: “PERU, Loreto, Arcadia, 0°59.37'S, 75°18.55'W, 150m, 31 Oct-10 Nov. 1993 leg., D.A. Nickle & J. Lewis; Castana 227; E66". Holotype will be deposited at the Museo de Historia Natural, Universidad Nacional Mayor de San Marcos, Lima, Peru (MUSM). Paratypes: 12 soldiers and 14 workers with same data as holotype. Paratypes will be deposited as follows: five soldiers and six workers at the MUSM, five soldiers and six workers as part of lot no. USNM 10601 at the USNM; two soldiers and two workers as part of lot no. MZUSP 13095 at the MZUSP.
PERU, Loreto Province: Arcadia, 0°59.37'S, 75°18.55'W, 190m. Type material was collected on a tree in a very narrow flattened tunnel that was well concealed beneath green/white lichens at the rainforests of northern Peru. The tunnel was not obvious.
Monotypic genus – see generic diagnosis.
Imago. Unknown.
Soldier (Figs 1–2). Head capsule with scattered long bristles and a few short hairs. Nasus with four shorter bristles and a few microscopic hairs near apex. Labrum with four short hairs. Postclypeus and postmentum glabrous. Thoracic nota and abdominal tergite I glabrous. Tergite II with a few long bristles at posterior margin; tergites III–X with several long bristles at posterior margin; IX–X tergites with bristles and hairs distributed on entire surface. Sternites with more bristles than tergites. Legs with two long bristles, first proximal and second at middle on external surface of all tibiae; short hairs on internal surface of all tibiae. Head capsule yellow; nasus reddish; antenna yellow with articles III–IV reddish; thoracic nota pale yellow; digestive tube visible through abdominal sclerites. Measurements (mm) of six soldiers are given as ranges, followed by holotype values in parentheses: LH, 0.85–0.90 (0.88); LHp, 0.46–0.49 (0.48); WH, 0.43–0.48 (0.48); HH, 0.30–0.32 (0.32); WP, 0.25–0.27 (0.26); LT, 0.48–0.51 (0.49). Ratios: LH/WH, 1.83–1.98 (1.83); LH/LT, 1.74–1.80; LHp/LT, 0.94–0.98 (0.98).
Worker (Figs 3–15). Head capsule with numerous erect bristles and few short hairs over entire surface. Postclypeus with six bristles at anterior margin; labrum with numerous bristles. Pronotum with few short hairs on both margins; mesonotum with four bristles and metanotum with six bristles. Tergites with decumbent bristles over surface plus few erect bristles toward posterior margin. Sternites with decumbent bristles over surface plus erect ones at posterior margin. Legs with two long bristles, first proximal and second at middle on external surface of all tibiae among shorter ones. Head capsule and thoracic nota whitish; digestive tube visible through abdominal sclerites. Mandibles and digestive tube, under genus description. Measurements (mm) of seven workers are given as ranges: WH, 0.48–0.50; DA–M1+2, 0.05–0.06; LT, 0.44–0.48. Ratio: WH/LT, 1.04–1.14.
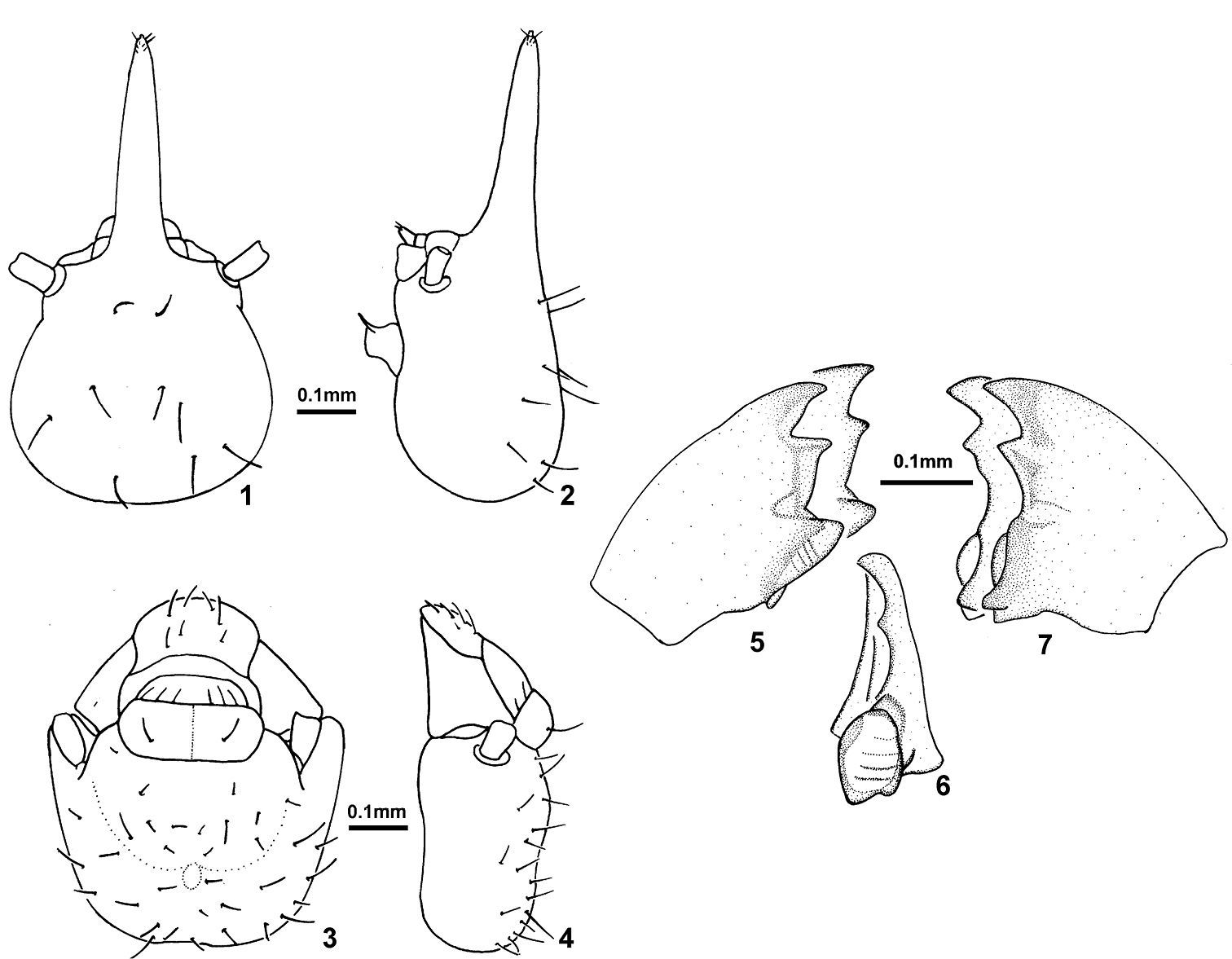
Sinqasapatermes sachae 1 soldier head in dorsal view 2 soldier head in profile 3 worker head in dorsal view 4 worker head in profile 5 worker left mandible in dorsal view 6 worker right mandible, showing molar plate in frontal view 7 worker right mandible in dorsal view.
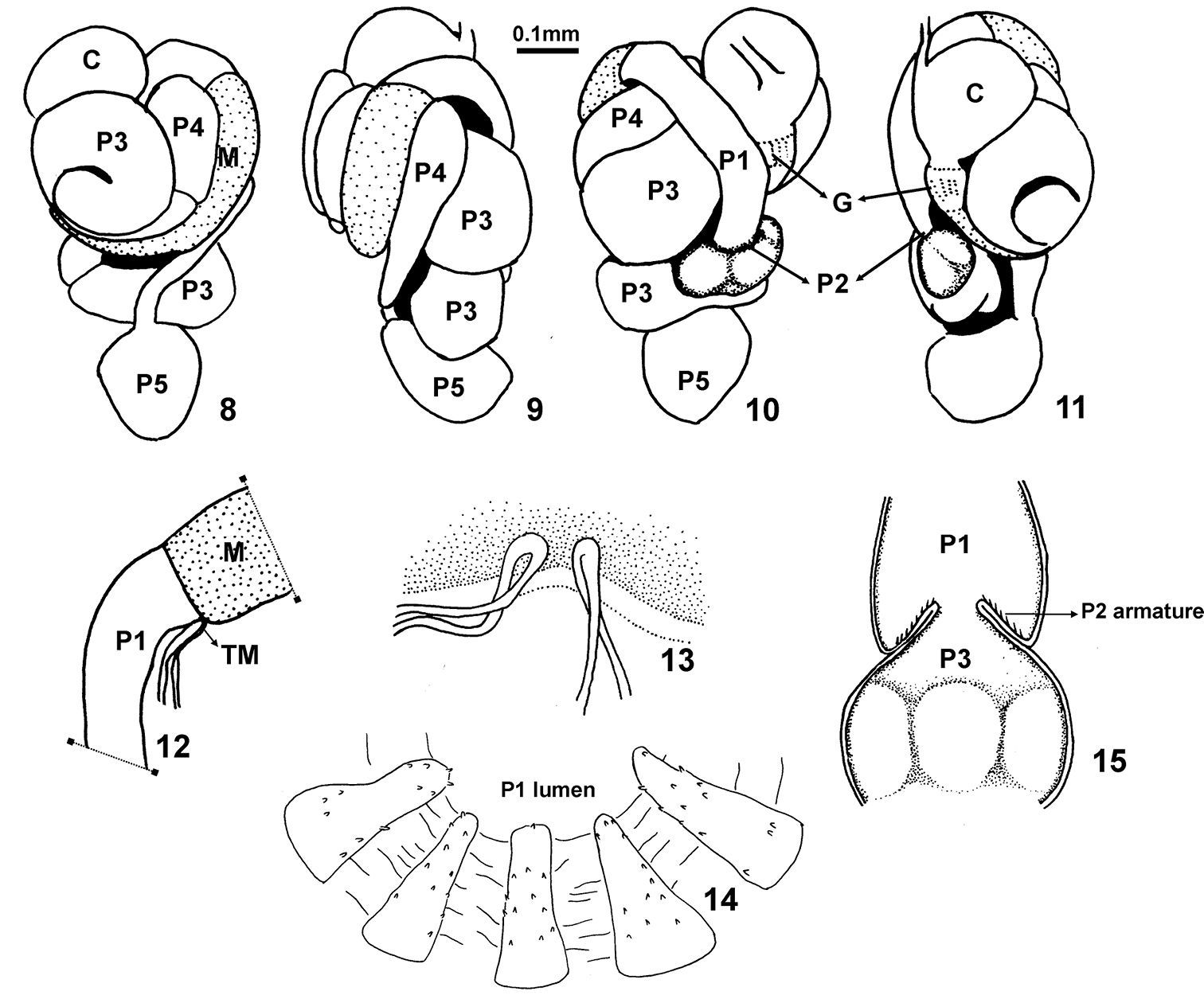
Sinqasapatermes sachae. 8–11 worker gut in situ respectively from dorsal, right, ventral and left views 12–13 Malpighian tubules attachment 14 worker enteric valve armature 15 scheme of worker enteric valve configuration. C crop G gizzard M mesenteron, stippled P1 first proctodeal segment P2 enteric valve P3 paunch P4 colon P5 rectum TM Malpighian tubules.
Noun in apposition taken from Quechua, sacha, meaning forest.
Figs 5 and 7 illustrate two sets of mandibles, which represent variation among workers of Sinqasapatermes sachae from the same sample. Those workers have not differentiation in coloration, pilosity, size or any other morphological characteristic to assume they belong to different instars.
This work was supported by Project grants Nos. 87–008, 89–163, 90–010, 92–032 from the Earthwatch Institute to DAN to conduct field research on termites in Peru. Additional support was provided by grants from the Department of Entomology, U.S. Museum of Natural History to DAN and J. E. Lewis to participate in the Amazon Expeditions Project, which helped further continuous work on Peruvian termites. DAN thanks the government of Peru for hosting scientists from USNM and SEL, USDA to study in Loreto Province, Peru. CC received support from PIP CONICET No. 6502, The Linnean Society through an SRF Award and a doctoral scholarship from CONICET. Thanks are extended to SEL, USDA, for providing space and equipment during aone-month visit in 2008. We are indebted to Eliana M. Cancello, MZUSP, who provided valuable advice regarding the taxa herein described, Lelia I. Albarracin de Alderetes, ADILQ (Asociación de Investigadores en Lengua Quechua) for confirming the meaning of words in the Quechua language, and J. E. Lewis, SEL, USDA, for helping with field work.
Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the USDA. The USDA is an equal opportunity provider and employer.