





(C) 2010 Lawrence R. Kirkendall. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Invasive bark beetles are posing a major threat to forest resources around the world. DAISIE’s web-based and printed databases of invasive species in Europe provide an incomplete and misleading picture of the alien scolytines and platypodines. We present a review of the alien bark beetle fauna of Europe based on primary literature through 2009. We find that there are 18 Scolytinae and one Platypodinae species apparently established in Europe, from 14 different genera. Seventeen species are naturalized. We argue that Trypodendron laeve, commonly considered alien in Europe, is a native species; conversely, we hypothesize that Xyleborus pfeilii, which has always been treated as indigenous, is an alien species from Asia. We also point out the possibility that the Asian larch bark beetle Ips subelongatus is established in European Russia. We show that there has been a marked acceleration in the rate of new introductions to Europe, as is also happening in North America: seven alien species were first recorded in the last decade.
We present information on the biology, origins, and distributions of the alien species. All but four are polyphagous, and 11 are inbreeders: two traits which increase invasiveness. Eleven species are native to Asia, six to the Americas, and one is from the Canary Islands. The Mediterranean is especially favorable for invasives, hosting a large proportion of the aliens (9/19). Italy, France and Spain have the largest numbers of alien species (14, 10 and 7, respectively). We point out that the low numbers for at least some countries is likely due to under-reporting.
Finally, we discuss the difficulties associated with identifying newly invasive species. Lack of good illustrations and keys hinder identification, particularly for species coming from Asia and Oceania.
Invasive species, polyphagy, inbreeding, Ambrosiodmus, Ambrosiophilus, Coccotrypes, Cyclorhipidion, Dactylotrypes, Dryocoetes, Gnathotrichus, Hypocryphalus, Hypothenemus, Phloeosinus, Phloeotribus, Megaplatypus, Monarthrum, Xyleborinus, Xyleborus, Xylosandrus
The great British ecologist Charles Elton presciently
referred to the effect of invasive species as “one of the great historic
convulsions in the world’s fauna and flora” (
Introduced wood-borers are a major concern to regions
with significant forest resources. Around the world, dozens if not
hundreds of alien phytophagous insects become established every decade,
and wood-borers make up a significant proportion of these (
There has been over three decades of discussion of the problems posed by introduced bark beetles. Steve Wood’s 1977 paper was developed from a talk given at the XIVth International Congress of Entomology in 1972. Both this and the subsequent treatment of the topic in the introductory material of the 1982 monograph (pages 25–27) were from an American point of view: which species have been introduced to the Americas, and which North, Central or South American species have become established in Europe.
With respect to exotic wood-boring insects, for North
America, much is known about which invasive species are present and
where (
There are two sources of newcomers to a fauna: species originally from distant regions or other continents, and those from the same region or continent which are expanding or shifting their ranges. We consider here only established species immigrant to continental Europe. Within-Europe range expansions are of interest in themselves, but ecologically and evolutionarily are a distinct phenomenon from that of the establishment of truly exotic species. We will use the term ‘alien species’ here in the sense of alien to Europe, originating outside the bounds of continental Europe.
MethodsTerminology
The terminology of invasion biology is much disputed (e.g.
While we adopt the same definition of Europe
used in DAISIE and Fauna Europaea, we exclude the Macaronesian islands,
preferring to focus on continental Europe (including Ireland and the
United Kingdom). Consequently, we consider the Canary Island endemic Dactylotrypes longicollis to be alien to Europe, and we do not treat the alien species found on the Azores (
For brevity, in taxonomic contexts, we use bark beetle to include both Scolytinae and the closely related Platypodinae. Ambrosia beetles cultivate symbiotic fungi on the walls of their tunnel systems, which fungi are the sole food of larvae and adults. All Platypodinae are ambrosia beetles, as are many genera of Scolytinae.
Sources of data
Our starting point for listing alien bark beetles
was the European database for alien organisms DAISIE (Delivering Alien
Invasive Species Inventories for Europe). The DAISIE project encompasses
over 11, 000 species of all types of organisms, and is meant to be a
central clearing house for information on biological invasions in
Europe, and the database is continually updated. The geographic and
taxonomic information in DAISIE is intended to play a key role in future
national and international efforts to monitor and combat the spread of
harmful non-native organisms. This information comes in two forms, the
web-based database (
In addition to the DAISIE website, we consulted
We also searched ISI Web of KnowledgeSM (and Internet
more generally), but quickly found that almost none of the literature
on alien bark beetles can be found by searching the web. The sources for
the data in DAISIE are not given. To investigate the validity of the
records available in the online databases, we searched the literature
at our disposal, including the world catalog for bark beetles (
Treatments of data
We have attempted to classify the phase of establishment of each species (Table 1),
given the collection localities and dates which are available in the
literature. Phases range from Phase 1 (newly collected or intercepted,
no evidence of establishment) to Phase 5 (apparently distributed
throughout currently suitable habitat in Europe). (Since this paper
focuses on aliens for which there is evidence of establishment, we do
not treat species which are in Phase 1.) We did not feel that enough was
known about alien bark beetle populations (in particular, about local
abundances) to apply the Stages system of
The population phases which we apply to alien species in Europe.
| Stage | Population level in Europe | Examples of evidence (not exhaustive) |
|---|---|---|
| Phase 1 | Interception, recently arrived (no evidence of establishment) | Collected from imported plant material; trapped at port or near imported logs; unique, old literature records |
| Phase 2 | local colony persisting | One area: many specimens; repeated collections; collections in natural forests far from ports of entry |
| Phase 3 | >1 colony, not spreading. | Disjunct populations, but no sign of expanding |
| Phase 4 | more than one large colony, spreading | Disjunct populations: Well established in several areas and still spreading |
| Phase 5 | established throughout suitable habitats | Distributed throughout region with currently suitable climate and host plants |
Problems with data quality
As we quickly discovered to our dismay, literature documenting the discovery and spread of alien species is scattered and mostly published in obscure and difficult to obtain journals and newsletters, in a bewildering variety of languages: few of these publications are peer reviewed and almost none indexed in ISI Web of KnowledgeSM. Much of the knowledge of new discoveries seems to have been transmitted by word of mouth, in Europe.
Adding to the confusion is the fact that old names die hard. Much of the literature on introduced species promulgates names used in the original papers but which are no longer used. This is especially true of review papers and invasive species databases.
Many articles lack information on who identified the specimen(s) and what criteria were used. New locality records (even country records) seem to occasionally be based on similarity with a species which is known to be in nearby countries, or based on old, incomplete keys; both methods can easily lead to mistakes in difficult taxa, such as Hypothenemus or Coccotrypes, which only experienced specialists can identify with any degree of confidence. Almost never is information on the deposition of voucher specimens stated; to confirm the identity of the species, one must try to find and contact an author in order to locate specimens.
Results and discussionWhich alien species are established in Europe?
The species present. There are 19 alien species established in continental Europe, according to our sources (Table 2). One of these, Megaplatypus mutatus, is a platypodine; the remainder are scolytines. Of these 19, we classify 14 as potentially expanding (Phases 2 – 4), 5 as probably currently spreading (Phases 3 – 4). All but one are considered naturalized: Xylosandrus morigerus is not known to have established populations in the wild, but seems to have a permanent presence in European greenhouses.
The alien Scolytinae and Platypodinae of Europe, and the countries in which they are established. First: first record or first publication. Phase: phase of colonization, see Table 1.
| Species | Established in countries | First | Phase | Notes, References |
|---|---|---|---|---|
| *Ambrosiodmus rubricollis (Eichhoff) | IT | 2008 | 2 |
|
| *Ambrosiophilus atratus (Eichhoff) | IT | 2007 | 2 |
|
| Coccotrypes dactyliperda (Fabricius) | ES, FR, GR, HU (cultivated palms), IT, MA | 1884 | 5 |
First mention is GR, |
| *Cyclorhipidion bodoanum (Reitter) | BE, CH, DE, FR, IT, NE | 1960 | 4 |
BE, NE, |
| Dactylotrypes longicollis (Wollaston) | CA, ES, FR, IT | 1949 | 4 | ES, |
| *Dryocoetes himalayensis Strohmeyer | CH, FR | 2009 | 4 | Knížek, unpub., CH and FR, established. |
| Gnathotrichus materiarius (Fitch) | BE, CH, CZ, DE, ES, FI, FR, IT, NE, SE | 1933 | 5 | BE, |
| *Hypocryphalus scabricollis (Eichhoff) | MA | 1991 | MA, |
|
| *Hypothenemus eruditus Westwood | ES, FR, IT, MA | 1924 | 5 |
|
| Megaplatypus mutatus (Chapuis) a | IT | 2000 | 2 | IT, |
| *Monarthrum mali (Fitch) | IT | 2008 | 2 | IT, |
| *Phloeosinus rudis Blandford | FR, NE | 1940 | 3 | FR, |
| Phloeotribus liminaris (Harris) | IT | 2004 | 2 | IT (only), |
| Xyleborinus attenuatus (Blandford) b | AT, CH, CZ, DE, ES, NE, PL, SE, RU, SK, UN | 1987 | 5 | AT: |
| Xyleborus affinis Eichhoff | AT | 2006 | 3 | HU: found in imported Dracaena, no recent records (Merkl Otto, email, |
| *Xyleborus pfeilii (Ratzeburg) | AT, BG, CH, CA, CZ, DE, ES, FR, HU, IT, PL, SI, SK, UN | 1837 | 5 |
Infrequently collected, but widespread in Europe and N. Africa. AT and DE, “Gallia”, |
| Xylosandrus crassiusculus (Motschulsky) | IT | 2003 | 2 | IT, |
| Xylosandrus germanus (Blandford) | AT, BE, CH, CZ, DE, ES, FR, IT, NE | 1950 | 5 | AT, |
| Xylosandrus morigerus (Blandford) | AT, CZ, FR, IT, UK | 1916 | 3 | UK, FR, AT and CZ (Bohemia), |
Country abbreviations: AT Austria; BE Belgium; BG Bulgaria; CA Croatia, CH Switzerland; CZ Czech Republic; DE Germany; ES Spain; FI Finland; FR France; GR Greek; HU Hungary; IT Italy; MA Malta; NE Nederland; PL Poland; RU Russia; SE Sweden; SI Slovenia; SK Slovakia; UK United Kingdom; UN Ukraine.
a The only Platypodinae; treated as Platypus sulcatus or Platypus mutatus in most earlier literature. b Treated as Xyleborinus alni (Niijima, 1909) in most literature. *Species not treated as established extra-European aliens in DAISIE.
Nine of our 19 species are not classified as
established aliens in DAISIE. We explain their inclusion here briefly.
Five on our list are classified by DAISIE as “status unknown”. In two,
this is probably due to simple “coding errors”: there is no doubt that
widely distributed species as (1) Coccotrypes bodoanum and (2) Hypothenemus eruditus are well-established aliens. That three more restricted species are established aliens is less widely known. (3) Phloeosinus rudis was collected in 1940 from Thuja japonica branches in St. Tropez (
Xyleborus affinis is tentatively included in our list, because of the Malaise trap catch in Austria (
Two ambrosia beetles on our list but not in DAISIE are only recently discovered: Ambrosiodmus rubricollis (
The last species on our list of alien species, Xyleborus pfeilii,
is currently considered to be indigenous. This ambrosia beetle is
considered rare but found in much of Europe as well as in northern
Africa and Turkey (
Finally, there is one species which we did not
include but which may have recently made its first inroads into Europe.
The highly aggressive Asian larch bark beetle Ips subelongatus (Motschulsky) has long considered synonymous with the European larch bark beetle Ips cembrae (Heer) (
Two species are listed by
The written list of aliens (
The genera present. Clearly, a wide variety of bark beetles are capable being transported to Europe, and there is a surprisingly high diversity which have succeeded in colonizing the continent: the 18 alien species comprise 16 different genera (15 of Scolytinae, 1 of Platypodinae), of which only five are present in the native fauna. Only two genera, Xylosandrus and Xyleborus,are represented by more than one exotic species; the Xyleborini (these two, plus Ambrosiodmus, Ambrosiophilus, Cyclorhipidion, and Xyleborinus) make up half of all adventive species.
When did they arrive?
The precise date of arrival in Europe is not known
for most species because the introductions of bark beetles (and of most
animals) are unintentional, and up to several decades can go by before
newly established exotics – especially those which are not pests – are
noticed (
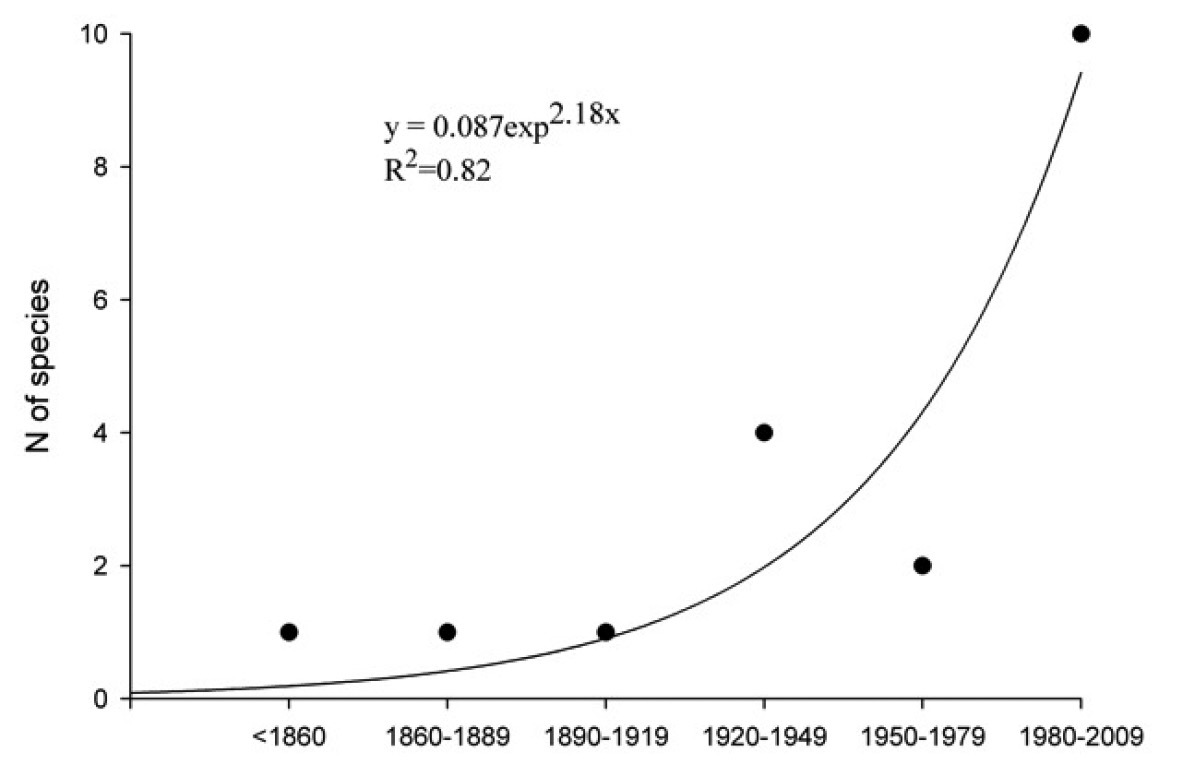
The accelerating rate of discovery of introduced Scolytinae and Platypodinae in Europe, shown as numbers of new species found in each 30-year period (data from Table 2).
How were they transported?
Many wood-boring insects, particularly scolytine and
platypodine beetles, are transported between continents. While the
majority of introductions of alien insects to Europe is via trade in
ornamental plants (
Biology of Europe’s alien bark beetles
Whether or not alien insects succeed in establishing breeding populations depends on a number of factors, including suitability of local climate and hosts, appropriate phenology, and the effects of potential competitors and natural enemies. Immigrants which are host generalists or which use host species which are abundant and widespread where they have arrived should have a good chance of establishing permanent populations, given appropriate climatic conditions.
Niche breadth. The vast majority of bark
beetles (particularly phloeophagous species) are monophagous, breeding
in one genus of host plants, or oligophagous, breeding in one family
of host plants (
Source and biology of alien bark beetles of Europe. Data from sources in Table 2,
| Species | Native to | Additional distribution | Zone | Feeds/Breeds | Host use |
|---|---|---|---|---|---|
| Ambrosiodmus rubricollis | east Asia | eastern North America, Australia | T | Xm/inbreeding | Polyphagous, broadleaf trees |
| Ambrosiophilus atratus | east Asia | North America | T | Xm/inbreeding | Polyphagous, broadleaf trees |
| Coccotrypes dactyliperda | ? (Old World) | globally distributed, tropics & subtropics | M | Spm/inbreeding | Polyphagous, mainly palm seeds in Europe |
| Cyclorhipidion bodoanum | north Asia | North America | T | Xm/inbreeding | Oligophagous, Fagaceae |
| Dactylotrypes longicollis | Canary Islands | Madeira, North Africa | M | Spm/outbreeding | Oligaphagous, palm seeds |
| Dryocoetes himalayensis | India | T | Phl/outbreeding | Polyphagous, Juglans regia, Pyrus lanata | |
| Gnathotrichus materiarius | eastern N. America | T | Xm/outbreeding | Polyphagous, conifers | |
| Hypocryphalus scabricollis | east Asia | M | Phl/outbreeding | Polyphagous, broadleaf trees | |
| Hypothenemus eruditus | American tropics? | globally distributed, tropics & subtropics | M | Phl/inbreeding | Polyphagous |
| Megaplatypus mutatus | South America | M | Xm/outbreeding | Polyphagous, broadleaf trees | |
| Monarthrum mali | eastern N. America | T | Xm/outbreeding | Polyphagous, broadleaf trees | |
| Phloeosinus rudis | east Asia | T | Phl/outbreeding | Oligophagous, Cupressaceae | |
| Phloeotribus liminaris | eastern US | M | Phl/outbreeding | Monophagous, Prunus | |
| Xyleborinus attenuatus | east Asia | North America | T | Xm/inbreeding | Polyphagous, broadleaf trees |
| Xyleborus affinis | Neotropics? | globally distributed, tropics & subtropics | M | Xm/inbreeding | Polyphagous |
| Xyleborus pfeilii | east Asia | North America | T, M | Xm/inbreeding | Polyphagous, broadleaf trees in Europe* |
| Xylosandrus crassiusculus | tropical & subtropical Asia | globally distributed, tropics & subtropics | M | Xm/inbreeding | Polyphagous |
| Xylosandrus germanus | east Asia | North America | T | Xm/inbreeding | Polyphagous |
| Xylosandrus morigerus | Asian tropics? | globally distributed, tropics | gh | Xm/inbreeding | Polyphagous; in Europe, greenhouse orchids |
Importance of reproductive system.
Particularly important to recently established, small populations are
Allee effects, the acute demographic, ecological and genetic problems
posed by low densities (
Both inbreeding and polyphagy should favor invasiveness. Interestingly, 10/15 polyphagous species are inbreeders, and 10/11 inbreeders are polyphagous (Table 4).
The relationship between feeding habits and reproductive systems, for alien Scolytinae and Platypodinae established in Europe. Data from Table 3.
| Reproduction type | Polyphagous | Not polyphagous |
|---|---|---|
| Inbreeding | 10 | 1 |
| Outbreeding | 5 | 3 |
Biogeography: Where are alien species established, and where did they come from?
Climatic zones of Europe. Though smaller in area, the Mediterranean zone is disproportionately rich in alien bark beetles (Table 3). Mediterranean ecosystems are particularly rich in biodiversity (
Country records. Although 22 European
countries recorded exotic species, large differences exist among the
numbers of alien insects recorded per country (Fig. 2).
Italy, France and Spain have the largest numbers of alien species (14,
10 and 7, respectively); for the former two countries, this
corresponds to about 10% of their national bark beetle fauna (
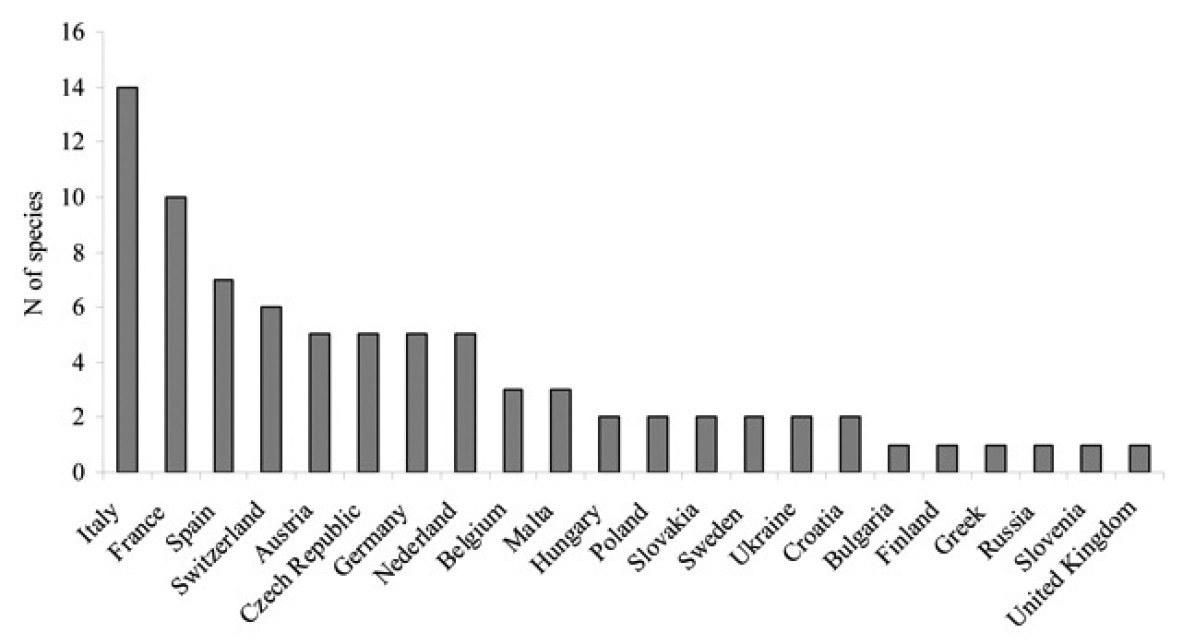
The numbers of alien bark beetles and pinhole borers per European country (data from Table 2).
The great differences among countries could be due to
several reasons. The number of alien insects is positively correlated
with country surface area (Roques et al., 2009). Furthermore bark
beetles show a latitudinal gradient in species richness: the number of
alien scolytines and platypodines generally decreases with the
increasing latitude (
While some of the differences between countries are real – Sweden does have fewer invasives than Switzerland – others are due to under-reporting at the country level; certainly, many of the differences among countries are due to differences in collecting effort and to the presence (or absence) of researchers with a special interest for Scolytinae and Platypodinae. Many sub-Scandinavia European countries are represented by zero or few records of alien bark beetle species but do have the requisite habitats. We found it particularly difficult to find detailed information on the bark beetle faunas of Portugal, eastern Europe, the Balkan countries, and countries of the eastern Mediterranean. Alien species doubtlessly can be found in these areas. The true ranges of alien bark beetles will not be known as long as there remain such gaps in our knowledge.
Unfortunately, here, too, the publicly available information on alien species in Europe is largely incorrect. Only for those recent arrivals established only in Italy are the country records in DAISIE accurate. Even species which have been established for over half a century and are well studied are not correctly reported in DAISIE: for both Gnathotrichus materiarius and Xylosandrus germanus, we can document at least three country occurrences missing from DAISIE.
The data in Fauna Europaea are similarly flawed. Three species are missing from the database, four country occurences (for three species) cannot be verified, and country records are incomplete for most alien species, including for Gnathotrichus materiarius (2 missing) and Xylosandrus germanus(3).
Where are the exotics from?By far, the vast
majority of recent interceptions of non-indigenous plant pests in
European countries are from Asia or from Europe, with an order of
magnitude fewer interceptions originating in North America (
The tropical affinities of one-third of the species (Table 3) might come as a surprise to some. However, all but Xylosandrus morigerus range into temperate climes – and that one exception is only found in greenhouses, in Europe.
Taxonomy and invasives
Increasingly, governments at all levels realize the
severity of threat posed by alien insects, and national and
international programs have been set in motion throughout the world to
address the problem (e.g.
Taxonomy plays a fundamental but often
underappreciated or overlooked role in strategies for monitoring,
intercepting, and managing both exotic and indigenous organisms,
including wood borers. Phytosanitary efforts to monitor or control new
invasive species will fail without correct taxonomic and biogeographic
information (and the latter is dependent on the former). Cryptic species
often differ in key elements of their biology, such as in phenology,
host preferences, pheromone behavior, susceptibility to natural
enemies (including diseases), and in the species or strains of
microorganisms which they carry with them. When such differences exist
between look-alike species, control measures will often be ineffective
if the species is misidentified. For example, similar appearing species
may originate from different regions; incorrect identification in such
an instance could lead to fruitless searches for key biological control
agents. Occasionally, taxonomists themselves have overlooked minute
morphological differences between sister species, but more often the
incorrect identifications are by nonspecialists relying on published
databases, keys, and illustrations rather than on consultation with
taxonomic experts (
The taxonomic impediment is often three-fold: difficult access to taxonomic specialists; poor taxonomic knowledge of the group involved; lack of user-friendly keys and illustrations. Taxonomic specialists are few and overworked; quarantine agencies, foresters and other instances must compete with taxonomists’ own research projects (and more and more with specimen-rich biodiversity surveys). Taxonomic knowledge can be inadequate in several ways: many genera of wood-boring insects (including scolytines and platypodines) have not been recently revised (some, never so); for some regions of the world, the wood-boring fauna is poorly known; and for some species groups which are highly successful as colonists, species-boundaries and proper nomenclature are inadequately understood. Finally, even where the wood-borers are fairly well known and keys do exist (e.g. Central America), for many genera the keys can only be used by specialists with access to reference material; illustrations sufficient for species-level identification (drawings or high-resolution photographs) exist only for a very limited number of species groups or genera.
A way out of this impasse is two-fold: use of adequate photographic documentation of subtle morphological differences, especially when coupled with expert intelligence software for developing illustration-rich keys; and the development of inexpensive molecular methods (fragment profile- or sequence-based) for separating species difficult to identify by morphology (DNA barcoding). Fortunately, tools for both are becoming increasingly well known and more widely accessible, as are possibilities to publish new finds rapidly via highly accessible electronic journals. Consequently, we are already seeing that new discoveries are being documented, identified, illustrated and published much more rapidly.
In the future there will be more and more Asian wood-borers colonizing Europe and North America. Currently there are no modern resources for identifying bark beetles from Asia, the Orient, or Oceania. What is needed is the methodical, thorough monographic work which Steve Wood was so good at, preferably including DNA sequencing. Until we have monographs for China, Southeast Asia, and Oceania – and the young taxonomic talents capable of applying them –many future immigrants will long remain enigmas.
This paper would not have been possible without the generous cooperation of our many European and American colleagues who responded promptly to our requests for information and research articles: Tom Atkinson, Maria Louisa Dal Cortivo, Arturo Goldarazena, Jean-Marc Henin, Miloš Knížek, Frank Köhler, Åke Lindelöw, Maria Josefa Lombardero, Sergio Lopez, Michail Mandelshtam, Leen Moraal, Merkl Otto, Alain Roques, Wojciech Solarz, Christian Stauffer, Oscar Vorst. Robert Rabaglia provided specimens of Xyleborus californicus. Finally, we thank two anonymous reviewers and the editor for improvements to the text. MF’s work has received funding from the European Community’s Seventh Framework Programme (FP7/2007–2013) under grant agreement n° 212459 PRATIQUE. The views in this paper do not necessarily reflect the European Commission’s views and in no way anticipate the Commission’s future policy in this area.