





(C) 2010 Fourie René. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
The purse-web spider genus Calommata Lucas, 1837 is revised in the Afrotropical Region. Following examination of the female type material, Calommata transvaalica Hewitt, 1916 is removed from synonymy with Calommata simoni Pocock, 1903 and revalidated. The females of both species are redescribed and their males described for the first time. While Calommata simoni is very widespread across tropical Africa, Calommata transvaalica is endemic to northern South Africa. Four new species are described, all known only from males: Calommata megae sp. n. (Zimbabwe), Calommata meridionalis sp. n. (South Africa), Calommata namibica sp. n. (Namibia) and Calommata tibialis sp. n. (Ivory Coast and Togo). Notes are presented on the biology of each species.
conservation, endemic, Mygalomorpha, new species, revalidation
The spider family Atypidae includes a small group of
distinctive Mygalomorpha, commonly known as purse-web spiders.
Presently it comprises three genera, Atypus Latreille, 1804, Calommata Lucas, 1837 and Sphodros Walckenaer, 1835, representing 43 species in Africa, Europe, Asia and North America (
Atypidae represents an ancient branch of the
Mygalomorpha, with morphological and molecular phylogenetic analyses
placing the family basally in the suborder, sister to the
Antrodiaetidae in the clade Atypoidina = Atypoidea (
The Atypidae, with a residual dorsal abdominal scutum in males, can be characterised by (
While the taxonomy of the Atypidae of North America (
Until now, Calommata was thought to be represented in the Afrotropical Region by a single species described from Cameroon, Calommata simoni Pocock, 1903 (
In this paper the Calommata of the Afrotropical Region are revised, Calommata transvaalica is removed from synonymy with Calommata simoni, and four new species are described. While the biology of Calommata simoni and Calommata transvaalica has been studied (
All spiders were studied under an Olympus SZX10
stereomicroscope under 10× magnification. Measurements (body and legs)
were taken with a measuring eyepiece and are given in millimetres (mm).
The proportional indices used follow
Material of male specimens of Calommata meridionalis sp. n., Calommata namibica sp. n., Calommata simoni and Calommata tibialis sp. n. was prepared for Scanning Electron Microscopy (SEM). Specimens were transferred to absolute (100 %) ethanol and left overnight. After drying in hexamethyldisilazane (HMDS), the specimens were glued to rounded aluminium rivets using two-sided copper strips and then coated with gold for examination using a JEOL JSM-6480 scanning electron microscope at the MRAC. Digitised micrographs were taken.
Digital photographs of the dorsal habitus of a Calommata simoni female were taken with a Leica MZ16 and stacked using the LAS automontage software at the MRAC. Digital photographs of the dorsal habitus of the remaining species and a Calommata simoni male were taken using a Nikon Coolpix 8400 mounted on a Nikon SMZ800 stereomicroscope at the UFS. The extended focal range images were stacked using CombineZM software to increase depth of field (http://www.hadleyweb.pwp.blueyonder.co.uk/).
Abbreviations:AL abdomen length
ALS anterior lateral spinneret(s)
AMNH American Museum of Natural History (New York, U.S.A.)
AW abdomen width
BMNH BritishNatural History Museum (London, U.K.)
CL carapace length
CW carapace width
MRAC Royal Museum for Central Africa (Tervuren, Belgium)
NCA National Collection of Arachnida, Plant Protection Research Institute (Pretoria, South Africa)
NMBA National Museum (Bloemfontein, South Africa)
NMSA Natal Museum (Pietermaritzburg, South Africa)
PLS posterior lateral spinneret(s)
PMS posterior median spinneret(s)
UFS University of the Free State (Bloemfontein, South Africa)
TL total length
TMSA Ditsong National Museum of Natural History (Pretoria, South Africa)
USNM National Museum of Natural History (Washington D.C., U.S.A.)
Taxonomy
Family Atypidae
Subfamily Atypinae
Pachyloscelis fulvipes Lucas, 1835 from Java and Sumatra
Calommata can be distinguished from Atypus and Sphodros by three main characteristics: 1) Spermathecal structure. In Atypus, there are two broad plates each bearing two or more small receptacula (
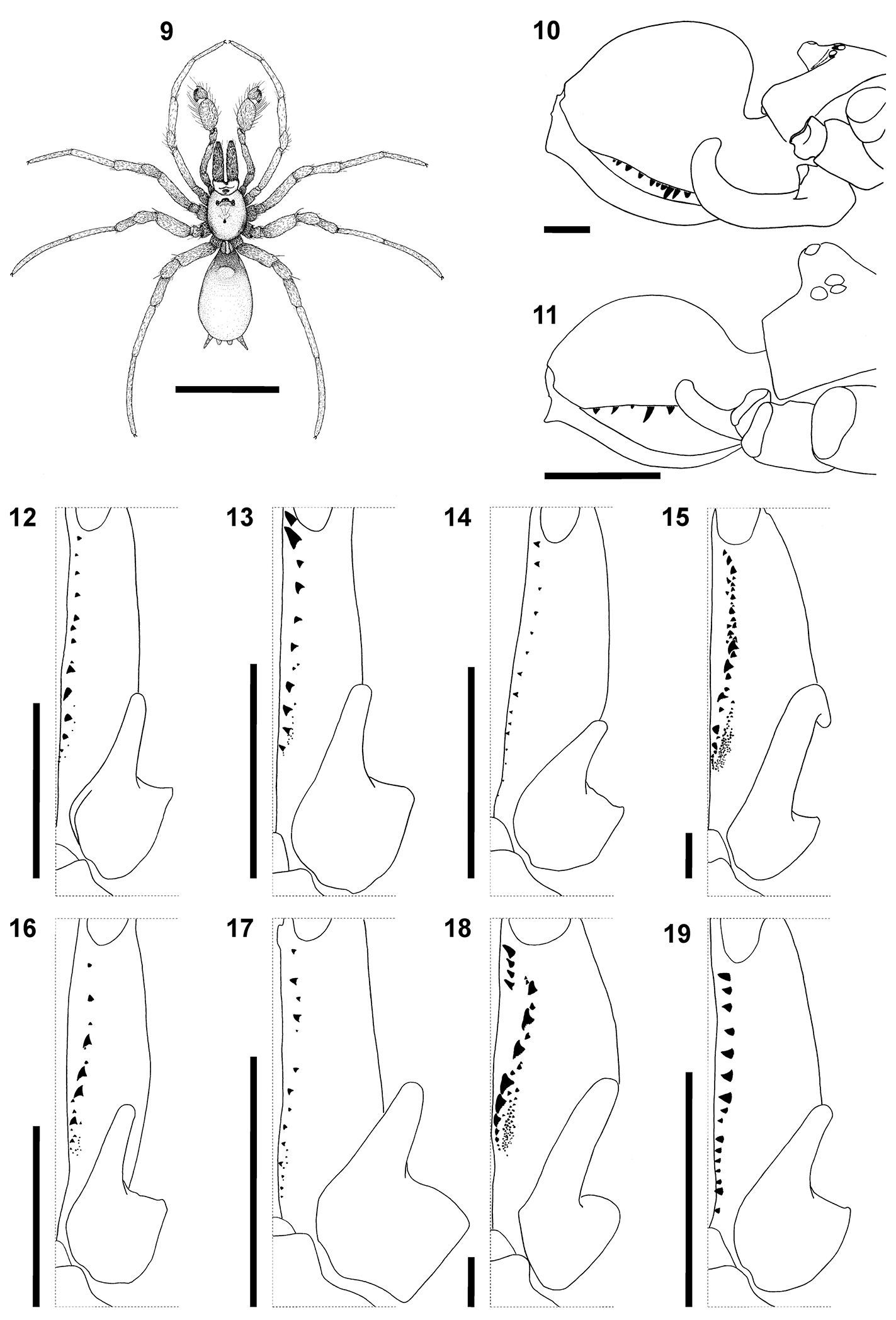
Medium to large sexually dimorphic spiders (Figs 1–9),
females 18.60–33.40 in length and males 5.10–9.35 in length. Carapace
with an anterior, strongly elevated median ocular tubercle (Figs 10, 11), with a flattened posterior part traversed by a longitudinal thoracic furrow with a small deep pit in the middle (Figs 1–8).
Three faint lines run from the ocular tubercle, converging at the
fovea. Chelicerae massive, dorsally expanded with flattened sides,
bearing sharp teeth usually in one (rarely two) rows, and a long arched
fang with a slit-like distal opening and distinctive basal ledge along
its outer margin (Figs 10–19, 20, 21, 26, 29).
Cheliceral dentition of both sexes often variable between specimens and
even between opposing chelicerae of a specimen. The general dentition
pattern of each species and sex is indicated in Figs 12–19.
Chelicerae of females have larger, more numerous teeth and an
extensive field of tiny denticles retrolateral of the teeth row near the
cheliceral base (Figs 15, 18). Chilum single, large, varying in shape, oval, rectangular or pentagonal (Figs 1–8). Endites strongly elongated and curved upwards on the prolateral side (Figs 10, 11, 22).
Sternum with a distinctive labio-sternal suture anteriorly. Legs of
females short and stout, leg formula of females 4231. Legs of males of
normal size, with the tarsi pseudosegmented (Figs 23, 24, 30) and tarsi of legs III and IV distinctly longer (Figs 1–3, 5, 6, 8, 9);
leg formula of males usually 4321, rarely 4132. Tarsi with three
claws, paired claws of males with single row of teeth and unpaired claw
without teeth (Figs 23, 27), contra

Dorsal habitus of Afrotropical Calommata species 1 Calommata megae sp. n., male (Harare, Zimbabwe) 2 Calommata meridionalis sp. n., male (Erfenis Dam, South Africa) 3 Calommata namibica sp. n., male (Etosha, Namibia) 4 Calommata simoni Pocock, female (Galim, Cameroon) 5 same, male (Dja Reserve, Cameroon) 6 Calommata tibialis sp. n., male (Bassari–Sokodé, Togo) 7 Calommata transvaalica Hewitt, female (Blouberg, South Africa) 8 same, male (Groenkloof, South Africa). Scale bars: 5mm.
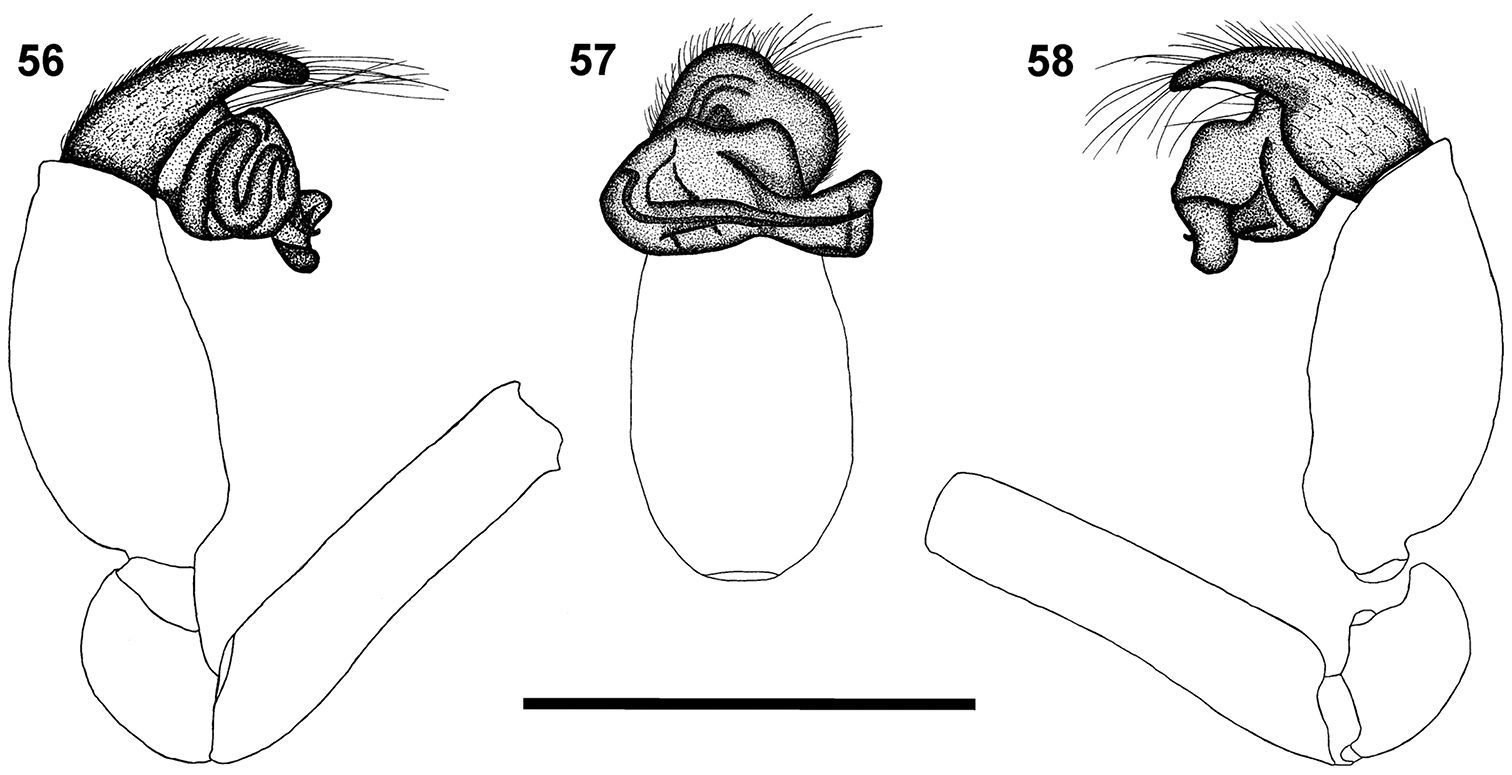
Somatic morphology of Calommata tibialis sp. n., male 9, 17 Calommata simoni Pocock, female 10, 15 and male 11, 16 Calommata megae sp. n., male 12 Calommata meridionalis sp. n., male 13 Calommata namibica sp. n., male 14and Calommata transvaalica Hewitt, female 18 and male 19: 9 dorsal habitus 10, 11 lateral view of chelicera, endites and anterior of carapace 12–19 left chelicera, ventral view, indicating dentition. Scale bars: 5mm (9), 1mm (10–19).

Scanning electron micrographs of Calommata meridionalis sp. n. 20–25 Calommata simoni Pocock 26–28 and Calommata tibialis sp. n. 29–31 males 20, 29 chelicera in ventral view 21 tip of fang 22 right endite, ventral view 23, 27 tarsus and claw, leg I (note pseudosegmentation of tarsus) 24, 30 tarsus IV, lateral and ventral view (note pseudosegmentation of tarsus) 25, 28, 31 detail of ventral scopulate setae on tarsus IV 26 chelicera in prolateral view.
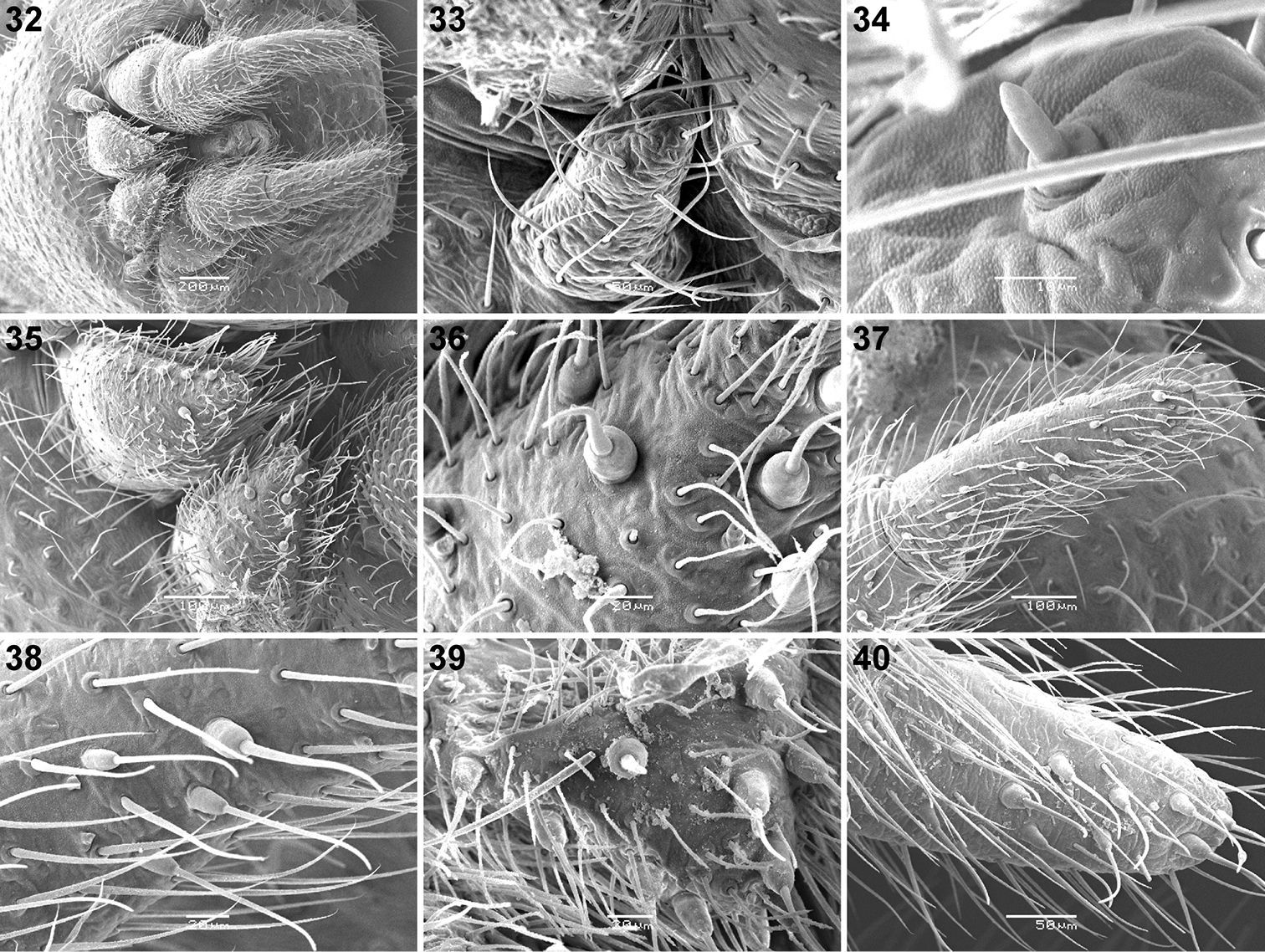
Scanning electron micrographs of Calommata simoni Pocock 32–38 and Calommata meridionalis sp. n. 39, 40 male spinnerets 32 spinneret field, ventral view 33 ALS 34 detail of single ALS spigot 35 PMS 36, 39 detail of PMS spigots 37, 40 PLS 38 detail of PLS spigots.
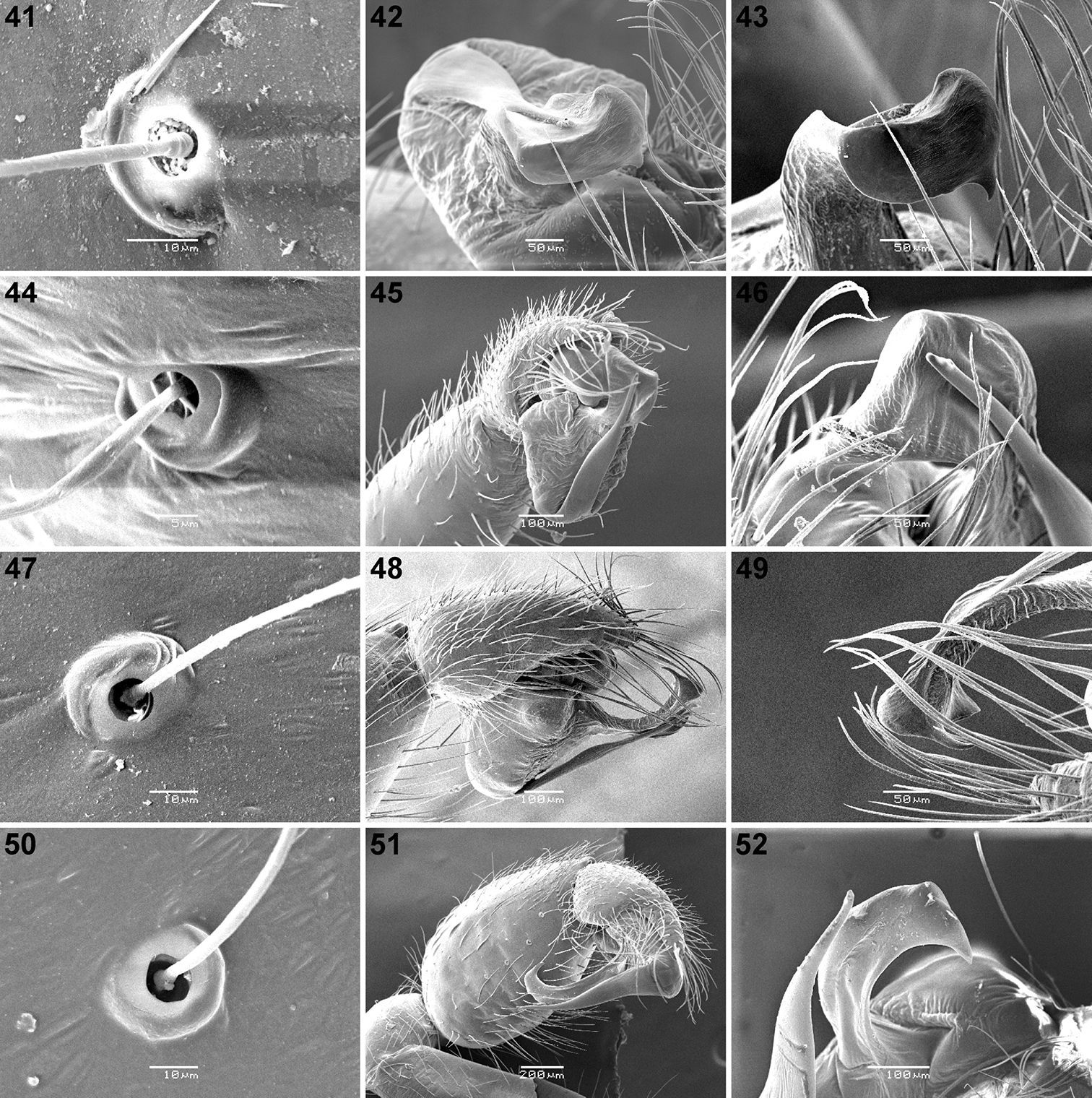
Scanning electron micrographs of Calommata meridionalis sp. n. 41–43, Calommata namibica 44–46, Calommata simoni Pocock 47–49 and Calommata tibialis sp. n. 50–52 males 41, 44, 47, 50 trichobothrial bases on palpal tarsus 42, 43, 46, 49, 52 distal end of conductor and embolus 45 palpal organ in prolateral ventral view 48, 51 palpal organ in prolateral view.
Key to the Afrotropical Calommata
[females of Calommata megae sp. n., Calommata meridionalis sp. n., Calommata namibica sp. n. and Calommata tibialis sp. n. unknown]
| 1 | Females | 2 |
| – | Males | 3 |
| 2 | Chelicerae with a single row of teeth along promargin running from fang base close to cheliceral base (Fig. 15); epigyne with two pairs of spermathecae (Fig. 62) (tropical Africa) | Calommata simoni |
| – | Chelicerae with an additional row of 2–4 teeth along promargin placed close to fang base, prolateral of main teeth row (Fig. 18); epigyne with one pair of transversely oval spermathecae (Fig. 69) (South Africa) | Calommata transvaalica |
| 3 | CL subequal to CW | 4 |
| – | Carapace clearly longer than wide, CL>1.15CW | 5 |
| 4 | Embolus and conductor in ventral view directed obliquely and retrolaterally towards endites (Fig. 54); palpal tibia clearly longer than wide (Fig. 53), patella-tibia index ~1.3 (Zimbabwe) | Calommata megae |
| – | Embolus and conductor in ventral view directed obliquely and retrolaterally towards distal end of chelicerae (Fig. 67); palpal tibia swollen, only slightly longer than wide (Fig. 66), patella-tibia index ~0.9 (Cote d’Ivoire and Togo) | Calommata tibialis |
| 5 | Embolus and conductor in ventral view directed transversely across palpal axis | 6 |
| – | Embolus and conductor in ventral view directed obliquely relative to palpal axis | 7 |
| 6 | Chelicerae with one or two very large teeth near fang base, remaining teeth distinctly smaller; several small denticles retrolateral of teeth row near base of chelicerae (Fig. 13); cymbium tip in ventral view broad and rounded (Fig. 57) (South Africa) | Calommata meridionalis |
| – | Chelicerae without one or two large teeth near fang base, teeth gradually decreasing in size from fang base to cheliceral base; denticles absent near base of chelicerae (Fig. 19); cymbium tip in ventral view tapering to rounded point (Fig. 71) (South Africa) | Calommata transvaalica |
| 7 | Cheliceral teeth variable in size, with at least three large teeth in midsection of teeth row and several denticles retrolateral of teeth row (Fig. 16); conductor and embolus not projecting beyond cymbial margin in ventral view (Fig. 64); conductor with distinct tooth and second ridge resembling a tooth distally on its dorsal surface (Fig. 49) (tropical Africa) | Calommata simoni |
| – | Cheliceral teeth tiny, subequal, without denticles retrolateral of teeth row (Fig. 14); conductor and embolus projecting far beyond cymbial margin in ventral view (Fig. 60); conductor with single tooth distally on its dorsal surface (Fig. 46) (Namibia) | Calommata namibica |
urn:lsid:zoobank.org:act:ED324676-F4FB-4F0D-816E-4670E7EE2577
http://species-id.net/wiki/Calommata_megae
Figs 1, 12, 53–55ZIMBABWE: Harare, Highlands [17°48'S, 31°05'E], 13.I.2003, on soil, by hand, M. Cumming (NCA 2004/362).
ZIMBABWE: 1imm.: same locality as holotype, 21.II.2000, in garden, M. Cumming (NCA 2004/1361).
The male of this species is recognised by the conductor that narrows and makes a half twist before broadening distally, and the obliquely orientated embolus and conductor (Figs 53–55). This species shares with Calommata tibialis sp. n. the carapace that is subequal in length and width (longer than wide in the other four species).
The specific epithet is a patronym in honour of the collector of the holotype, Meg Cumming, in recognition of her contributions to African arachnology, particularly in Zimbabwe.
Male holotype. Measurements: CL 2.00, CW 2.05, AL 2.95, AW 1.80, TL 7.00. Length of leg segments, and total: I 2.40 + 0.90 + 1.70 + 1.90 + 1.70 = 8.60; II 2.15 + 0.95 + 1.40 + 2.20 + 2.15 = 8.85; III 1.80 + 0.95 + 1.00 + 2.15 + 3.55 = 9.45; 2.35 + 1.10 + 1.28 + 2.50 + 4.35 = 11.58. Carapace index 0.98; patella-tibia index 1.30.
Carapace and chelicerae brown in colour (Fig. 1). Carapace flat and robust. Median ocular tubercle raised, darker in colour.Chelicerae with a single row of small teeth, increasing in size from fang base to base of chelicerae, with a few denticles near base of chelicerae (Fig. 12). Sternum and coxae light brown, remainder of legs dark brown, fading to light yellow-brown at tarsi. Legs weakly covered with bristles; prolateral side of patellae, tibiae and metatarsi of legs II–IV covered with spinules (thicker and shorter than bristles). Abdomen dark brown, with an irregular brown scutum present anteriorly (Fig. 1). Palpal cymbium short with rounded distal margin; embolus and conductor orientated obliquely, pointing retrolaterally towards base of chelicerae; conductor narrow, making a half twist before broadening distally, with single small curved tooth on its dorsal surface distally; embolus short and straight, slightly curved near tip (Figs 53–55).
Female. Unknown.
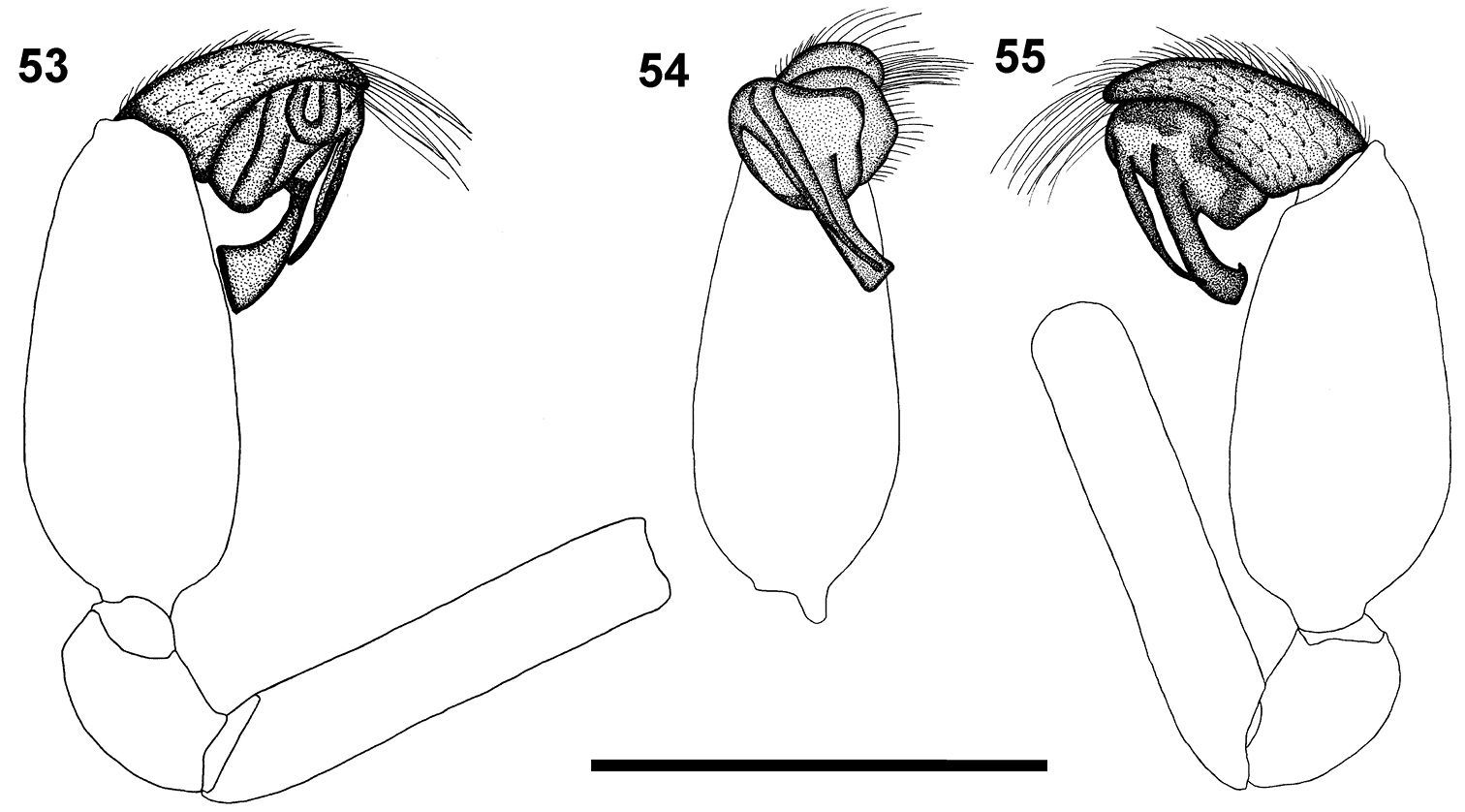
Left palp of male Calommata megae sp. n. 53 prolateral view 54 ventral view 55 retrolateral view. Scale bars: 1mm.
Known only from the type locality (Fig. 73).
Poorly known. The holotype male was collected in
mid-summer on the soil surface. The second instar juvenile specimen was
captured while ballooning and landing on the porch of a house (M.
Cumming, pers. comm.). Amongst atypids, ballooning has previously been
recorded in both Atypus and Sphodros (Wiehle 1953 cited in
urn:lsid:zoobank.org:act:7E2F6F05-6995-4C3B-AC59-30A0CE451914
http://species-id.net/wiki/Calommata_meridionalis
Figs 2, 13, 20–25, 39–43, 56–58Holotype male. SOUTH AFRICA: Free State Province: Erfenis Dam Nature Reserve, 28°29.888'S, 26°48.488'E, 21.IX–22.X.2005, pitfalls, unburned site 1, C. Haddad, S. Otto & R. Poller (NCA 2009/3488).
SOUTH AFRICA: Free State Province: 3♂: Bloemfontein, National Botanical Gardens, 29°03.006'S, 26°12.701'E, 27.X–16.XI.2009, pitfalls, grassland, C. Haddad (NMBA 13981); 7♂: Same data, 16–21.XI.2009 (NMSA 22616); 1♂: Same locality, 21.XI–7.XII.2009, pitfalls, grassland, C. Haddad & R. Fourie (MRAC 229029); 1♂: Erfenis Dam Nature Reserve, 28°30.373'S, 26°48.437'E, 21.IX–22.X.2005, pitfalls, burned site 1, C. Haddad, S. Otto & R. Poller (NCA 2009/3663); 4♂: Same locality, 28°30.134'S, 26°48.427'E, 22.X–22.XI.2005, pitfalls, burned site 2, C. Haddad, S. Otto & R. Poller (NCA 2007/3142); 1♂: Same locality, 28°29.741'S, 26°48.065'E, 21.IX–22.X.2005, pitfalls, unburned site 3, C. Haddad, S. Otto & R. Poller (NCA 2009/3664); 1♂: Oranjeville district, Vaal Dam, 26°59.523'S, 28°15.737'E, 1–29.X.2009, pitfalls, grassland, R. Fourie & A. Grobler (NCA 2009/3539).
SOUTH AFRICA: Free State Province: 1♂: Erfenis Dam Nature Reserve, 28°30.134'S, 26°48.427'E, 21.IX–22.X.2005, pitfalls, burned site 2, C. Haddad, S. Otto & R. Poller (MRAC, prepared for SEM).
The male of this species can be easily recognised from African congeners by the presence of one or two very large teeth at the fang base (Fig. 13), and the transversely orientated curved embolus with distally broadened conductor bearing a single tooth on its dorsal surface (Figs 42, 43, 57). The raised median ocular tubercle is broader than in the other species.
Left palp of male Calommata meridionalis sp. n. 56 prolateral view 57 ventral view 58 retrolateral view. Scale bars: 1mm.
This specific epithet is Latin for southern, referring to the distribution of the species, southernmost in the genus.
Male holotype. Measurements: CL 1.60, CW 1.25, AL 3.15, AW 1.68, TL 6.35 (5.60–7.80). Length of leg segments, and total: I 1.40 + 0.55 + 1.00 + 1.15 + 1.25 = 5.35; II 1.30 + 0.55 + 0.85 + 1.24 + 1.45 = 5.39; III 1.05 + 0.64 + 0.64 + 1.20 + 2.40 = 5.93; IV 1.45 + 0.64 + 0.75 + 1.44 + 2.90 = 7.18. Carapace index 1.28; patella-tibia index 0.97.
Carapace and chelicerae dark brown in colour (Fig. 2). Median ocular tubercle raised, darker in colour; median ocular tubercle broader than in other species. Chelicerae with single prolateral row of teeth, one very large tooth close to fang base, sometimes accompanied by second large tooth, remaining teeth distinctly smaller and subequal in size, with several denticles retrolateral of teeth row close to cheliceral base (Fig. 13). Sternum and coxae pale brown, remainder of legs brown, gradually fading to yellow at tarsi. Legs weakly covered with bristles; prolateral side of patellae, tibiae and metatarsi of legs II–IV covered with spinules. Abdomen grey-brown, with a round brown scutum anteriorly (Fig. 2). Palpal cymbium short with rounded distal margin; embolus and conductor orientated transversely across palpal axis, pointing retrolaterally; conductor broadened distally, with a single small tooth on its dorsal surface distally; embolus long, slightly curved in a S-form along its length (Figs 42, 43, 56–58).
Female. Unknown.
Endemic to central and northern Free State Province, South Africa (Fig. 73).
The species was collected exclusively by pitfalls in spring and early summer (September to early December) in the Grassland Biome of South Africa. Specimens were only collected in dark vertic clay and loamy-clay soils and not from sites with sandy soils. Most of the specimens were collected from sites near to freshwater streams and dams. Despite exhaustive attempts to locate burrows in the vicinity of pitfall sites (Erfenis Dam Nature Reserve and Botanical Gardens) none could be found.
urn:lsid:zoobank.org:act:099D7F5A-BBF3-482E-B570-3D8053C88E93
http://species-id.net/wiki/Calommata_namibica
Figs 3, 14, 44–46, 59–61NAMIBIA: Etosha National Park, Beisebvlakte, 18°32'S, 17°02'E, 10–14.XI.1996, A. Russell-Smith (MRAC 215409).
None.
Diagnosis. The male of this species can be recognised by the tiny cheliceral teeth in a single row, without retrolateral denticles (Fig. 14), and the obliquely orientated conductor and embolus projecting far beyond the retrolateral cymbial margin (Fig. 60).
The specific epithet refers to the country of the type locality.
Male holotype. Measurements: CL 1.80, CW 1.24, AL 2.75, AW 1.54, TL 6.30 (5.10–6.30). Length of leg segments, and total: I 1.21 + 0.50 + 0.83 + 1.09 + 1.35 = 4.98; II 1.40 + 0.51 + 0.62 + 1.13 + 1.40 = 5.06; III 1.05 + 0.49 + 0.47 + 1.16 + 1.95 = 5.12; IV 1.44 + 0.60 + 0.65 + 1.38 + 2.40 = 6.47. Carapace index 1.45; patella-tibia index 0.74.
Carapace and chelicerae dark brown (Fig. 3). Median ocular tubercle raised, narrow, darker in colour. Chelicerae with single prolateral row of tiny teeth, without denticles near cheliceral base (Fig. 14). Sternum and coxae light brown, femora dark brown; subsequent segments fading to light yellow at tarsi. Legs weakly covered with bristles; prolateral side of patellae, tibiae and metatarsi of legs II–IV covered with spinules. Abdomen dark grey, nearly black, with brown scutum present in the anterior half (Fig. 3). Palp with elongate cymbium, with tapering pointed distal margin; embolus and conductor orientated obliquely, pointing retrolaterally and distally, projecting far beyond retrolateral cymbial margin; conductor short, slightly broadened distally, with a very prominent, very long and slender tooth distally on its dorsal surface; embolus long, with slight bend in distal half (Figs 45, 46, 59–61).
Female. Unknown.
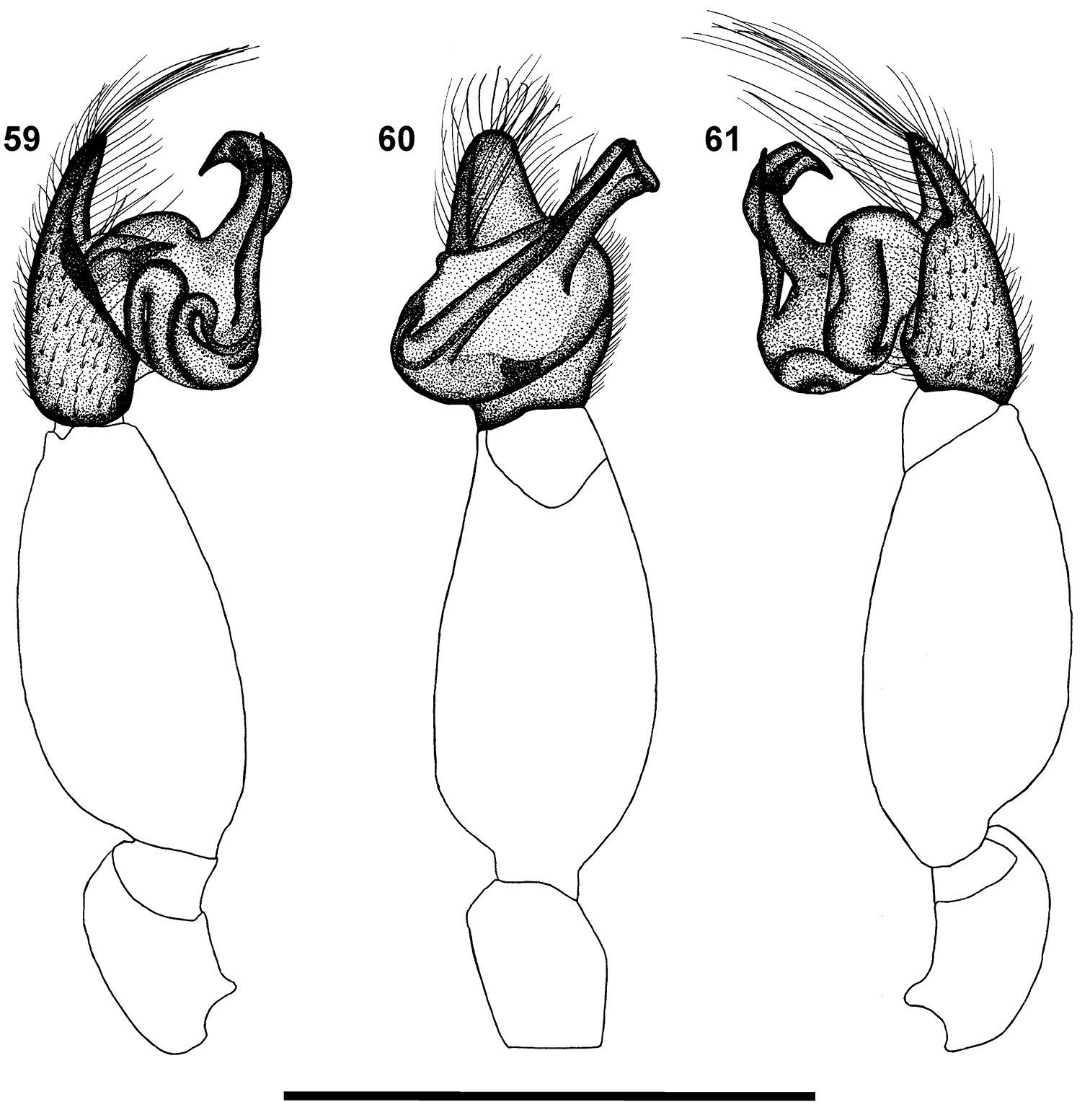
Left palp of male Calommata namibica sp. n. 59 prolateral view 60 ventral view 61 retrolateral view. Scale bars: 1mm.
Known only from the type locality (Fig. 73).
Poorly known. This species was collected in late spring in arid savanna.
http://species-id.net/wiki/Calommata_simoni
Figs 4, 5, 10, 11, 15, 16, 26–28, 32–38, 47–49, 62–65CAMEROON: Efulen [02°46'N, 10°43'E], G.L. Bates (BMHN, examined).
CAMEROON: 1♀: Galim [07°06'N, 12°28'E], 15.VIII–19.VIII.1971, F. Puylaert (MRAC 143671); 3♂: N of Dja Reserve, 03°41'N, 13°14'E, 8.III.2005, pitfalls, old secondary forest, I. Deblauwe (MRAC 220674); 1♂: Same locality, 8.III.2005, pitfalls, riverine forest, I. Deblauwe (MRAC 220663); 1♂: Same data, 6.V.2005 (MRAC 219754); 6♂: Same locality, 8.III.2005, pitfalls, young secondary forest, I. Deblauwe (MRAC 220659). CONGO D.R.: 1♂: Kisangani, Masako Forest, 00°35'N, 25°11'E, 25.II.2003, J.-L. Juakaly (MRAC 216031); 1♂: Same locality, 11.III.2003, pitfalls, primary forest, J.-L. Juakaly (MRAC 214347); 1♂: Same data, 11.III.2003 (MRAC 214354); 1♂: Same data, 11.III.2003 (MRAC 214385); 1♀: Kisantu, 05°08'S, 15°06'E, 1927, R. Vanderyst (MRAC 5201); 1♀: Zambi [05°51'S, 12°52'E], 1909–1915, American Museum Congo Expedition (AMNH). CÔTE D’IVOIRE: 1♂: Appouesso, Bossematié Forest, 06°35'N, 03°28'W, 19.IX.1994, pitfalls, rain forest, R. Jocqué & N. Séabé (MRAC 202481); 1♂: Same locality, 12.III.1995, pitfalls, forest, R. Jocqué & Tanoh (MRAC 205382); 1♂: Same data, 26.III.1995 (MRAC 205383); 1♂: Mankono, Ranch de la Marahoué, 08°27'N, 06°52'W, III.1980, riverine forest, J. Everts (MRAC 172117); 1♂: Same data (MRAC 172118); 1♂: Same data (MRAC 172119); 2♂: Same data (MRAC 172120). GUINÉE: 3♂: F.C. de Ziama, 08°24'N, 09°17'W, 22.IV.1998, pitfalls, rain forest, D. Flomo (MRAC 216239); 2♂: Same data, 5.V.1998 (MRAC 216248); 2♂: Same data, 18.V.1998 (MRAC 216249); 1♂: Same data, 14.VI.1998 (MRAC 216247); 1♂: Same data, 14.VI.1998 (MRAC 216250); 1♂: Same data, 14.VI.1998 (MRAC 216251); 1♂: Same data, 31.III.1999 (MRAC 216245); 2♂: Same data, 26.IV.1999 (MRAC 216243); 1♂: Same data, 26.IV.1999 (MRAC 216244); 1♂: Same data, 9.V.1999 (MRAC 216242); 2♂: Same data, 22.V.1999 (MRAC 216240); 1♂: Same data, 17.VI.1999 (MRAC 216241); 1♂: Same data, 31.III.2000 (MRAC 216246). KENYA: 1♂: Kakamega Forest, 00°13'N, 34°54'E, 24.I.2002, pitfalls, D. Shilabira Smith (MRAC 228141); 1♂: Same data, 13.IV.2002 (MRAC 220536). LIBERIA: 1imm.: Mount Coffee, Bensonville [06°29'N, 10°38'W], II.1894, constructs a tube-like nest under a log, collector unknown (USNM). MALAWI: 1♂: Chisasira Forest, 25km S of Chintheche, 11°50'S, 33°13'E, 1.XII.1977, pitfalls, Brachystegia woodland, R. Jocqué (MRAC 169498); 1♂: Same data, 1.XII.1977 (MRAC 169499). TANZANIA: 1imm.: Bunduki, Uluguru Mountains, 07°02'S, 37°38'E, 2.V.1957, nest, forest ground in litter, P. Basilewsky & N. Leleup (MRAC 111792).
The female of this species has the cheliceral teeth in a single row (Fig. 15), while an additional row is found in Calommata transvaalica (Fig. 18). The female genitalia have two pairs of small spermathecae (Fig. 62), while Calommata transvaalica only has a single pair of large transverse spermathecae (Fig. 69). The male of this species is recognised by the conductor ending broadly with a prominent tooth and sharp edge, appearing to be a second tooth, on its dorsal surface (Fig. 49). In Calommata simoni females the patella-tibia index is double that of Calommata transvaalica.
Female (measurements provided for female lectotype from Efulen, colouration for female from Galim). Measurements: CL 7.70, CW 6.10, AL 13.10, AW 8.85, TL 27.80 (25.80–33.40). Length of leg segments, and total: I 3.95 + 1.62 + 1.75 + 2.10 + 1.38 = 10.80; 3.70 + 2.25 + 1.70 + 2.23 + 1.60 = 11.48; III 3.65 + 2.75 + 1.55 + 1.78 + 1.39 = 11.12; IV 3.90 + 2.90 + 2.10 + 2.15 + 1.50 = 12.55. Carapace index 1.26; patella-tibia index 0.44.
Robustly built with short legs, carapace faded to creamy brown (Fig. 4). Median ocular tubercle raised, narrow, sloping sharply at fovea (Fig. 10). Single median line running from anterior of eye area to approximately middle of chilum. Chelicerae pale orange brown, darker laterally; chelicerae with a single row of small and medium sized teeth along promargin running from fang base close to cheliceral base, with extensive denticle field retrolateral of teeth row near cheliceral base (Fig. 15). Endites strongly elongated prolaterally, strongly curved upwards (Fig. 10). Sternum and legs light yellowish brown. Legs short and stout, leg formula 4231; legs III and IV more robust than legs I and II; leg I without bristles or spinules; leg II with few spinules distally on tibiae, and dorsal and lateral spinules on metatarsi and tarsi; legs III and IV with spinules from patellae to tarsi (mainly dorsal and prolateral) and covered in bristles. Abdomen globose and pale grey, with indistinct median heart marking in anterior half (Fig. 4). Epigyne forming a broad, weakly sclerotised plate ventrally, in dorsal view with two pairs of spermathecae; median pair subrectangular, rounded anteriorly, lateral pair subtriangular (Fig. 62). Female palp short, tibiae and tarsi flattened.
Male from Cameroon. Measurements: CL 2.20, CW 1.90, AL 3.80, AW 2.32, TL 8.20 (5.60–9.35). Length of leg segments, and total: I 2.30 + 0.75 + 1.64 + 1.91 + 1.55 = 8.15; II 2.25 + 0.86 + 1.45 + 2.16 + 1.88 = 8.60; III 1.84 + 0.90 + 0.98 + 2.23 + 2.95 = 8.90; IV 2.50 + 1.05 + 1.43 + 2.60 + 3.90 = 11.48. Carapace index 1.16; patella-tibia index 1.09.
Carapace and chelicerae brown (Fig. 5). Median ocular tubercle raised, narrow, darker in colour (Fig. 11).Chelicerae with single prolateral row of teeth, largest teeth in midsection of teeth row interspersed with smaller teeth anteriorly and posteriorly, with several denticles retrolateral of teeth row close to cheliceral base (Fig. 16). Endites elongated prolaterally, curving upwards (Fig. 11). Sternum and coxae light yellowish brown, rest of leg segments brown, fading to light yellow at tarsi. Legs weakly covered with bristles; prolateral side of patellae, tibiae and metatarsi of legs II–IV covered with spinules. Abdomen grey brown, with elongate brown scutum present in anterior half (Fig. 5). Palp with short cymbium, with rounded distal margin; embolus and conductor orientated obliquely, pointing retrolaterally and distally, not projecting beyond retrolateral cymbial margin; conductor short, broadened distally, with a prominent elongate tooth and sharp edge opposite the tooth, appearing as a second tooth; embolus short and straight (Figs 48, 49, 63–65).
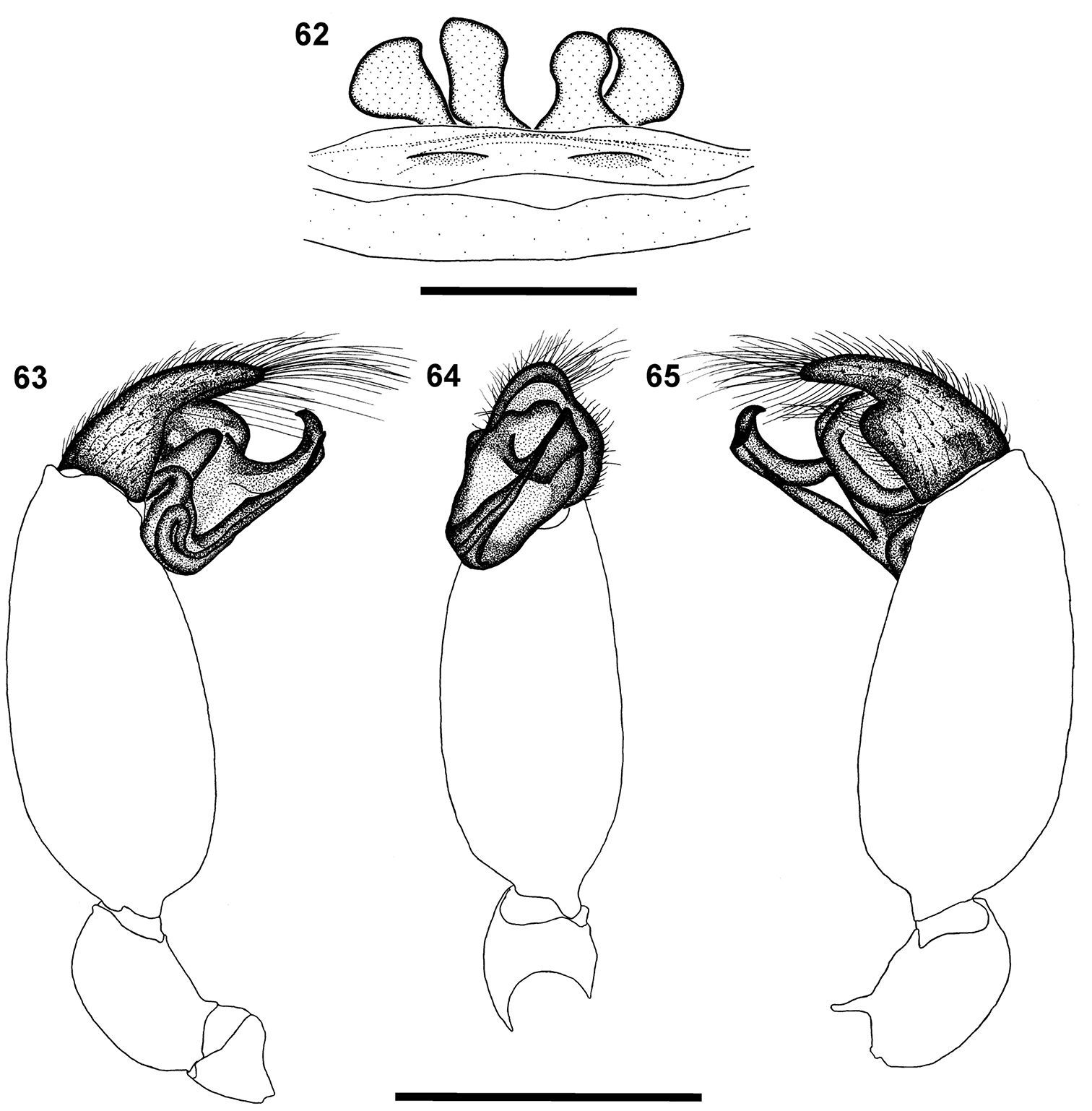
Calommata simoni Pocock female genitalia 62 and left palp of male 63–65: 62 dorsal view 63 prolateral view 64 ventral view 65 retrolateral view. Scale bars: 1mm.
Benoit’s (1967: 286, figs 1–4) drawings of a male “allotype” of Calommata simoni correspond with the males we have studied. However,
Widespread across tropical Africa in forests and savanna woodlands (Fig. 73).
The biology of “Calommata simoni” was studied by
Most of the specimens studied here from the MRAC collected in Guinée, Côte d’Ivoire, Kenya, Tanzania and Congo D.R. were collected in contrasting forest types across tropical Africa, indicating that Calommata simoni is tolerant and adaptable to a wide range of soil, vegetation and climatic variables.
urn:lsid:zoobank.org:act:E31586AD-B428-49F1-8F43-100356C90DCA
http://species-id.net/wiki/Calommata_tibialis
Figs 6, 9, 17, 29–31, 50–52, 66–68Holotype male. TOGO: Between Bassari and Sokodé, 09°15'N, 00°47'E, V–VII.1984, pitfalls, wooded savanna, P. Douben (MRAC 169501).
1♂: together with holotype. CÔTE D’IVOIRE: 1♂: Kossou, 06°57'N, 04°58'W, 28.IV.1975, pitfalls, wooded savanna, R. Jocqué (MRAC 169500).
CÔTE D’IVOIRE: 1 subadult ♀: Lamto, 06°12'N, 05°20'W, 6.III.1968, savanna with Borassus aethiopum, C. Girard (MRAC 232547); 1♂: Same locality, 11.II.1974, dirt road near biological station, P. Blandin (MRAC 232548).
The male of the species can be recognised by the carapace that is subequal in length and width (Fig. 6), the short, swollen palpal tibia, and the narrow conductor ending in a thick prominent tooth (Figs 51, 52).
The specific epithet refers to the palpal tibia of the male, which is distinctly shorter and more swollen compared to that of other African congeners.
Male holotype. Measurements: CL 2.13, CW 1.95, AL 3.45, AW 2.02, TL 6.65 (6.00–6.65). Length of leg segments, and total: I 1.98 + 0.65 + 1.35 + 1.71 + 1.63 = 7.32; II 1.80 + 0.75 + 1.10 + 1.60 + 1.80 = 7.05; III 1.49 + 0.80 + 0.75 + 1.70 + 2.45 = 7.19; IV 1.94 + 0.90 + 1.03 + 2.00 + 3.03 = 8.90. Carapace index 1.10; patella-tibia index 0.94.
Carapace and chelicerae orange brown (Fig. 6). Chelicerae with single row of alternating small and medium sized teeth, gradually decreasing in size from fang base to cheliceral base, with several denticles retrolateral of teeth row near cheliceral base (Fig. 17). Eye area raised, narrow, darker in colour. Sternum and coxae yellow, remainder of legs orange, fading to pale yellow at tarsi. Legs weakly covered with bristles; prolateral side of patellae, tibiae and metatarsi of legs II–IV covered with spinules. Abdomen grey, with pale orange-brown scutum anteriorly (Fig. 6). Palp with short cymbium, with rounded distal margin; tibia shorter and broader than in the other five species, slightly longer than wide; embolus and conductor orientated obliquely, pointing retrolaterally and distally, not projecting beyond retrolateral cymbial margin; conductor narrow with a thick prominent tooth distally on its dorsal surface; embolus short and slightly curved (Figs 51, 52, 66–68).
Female. Unknown.
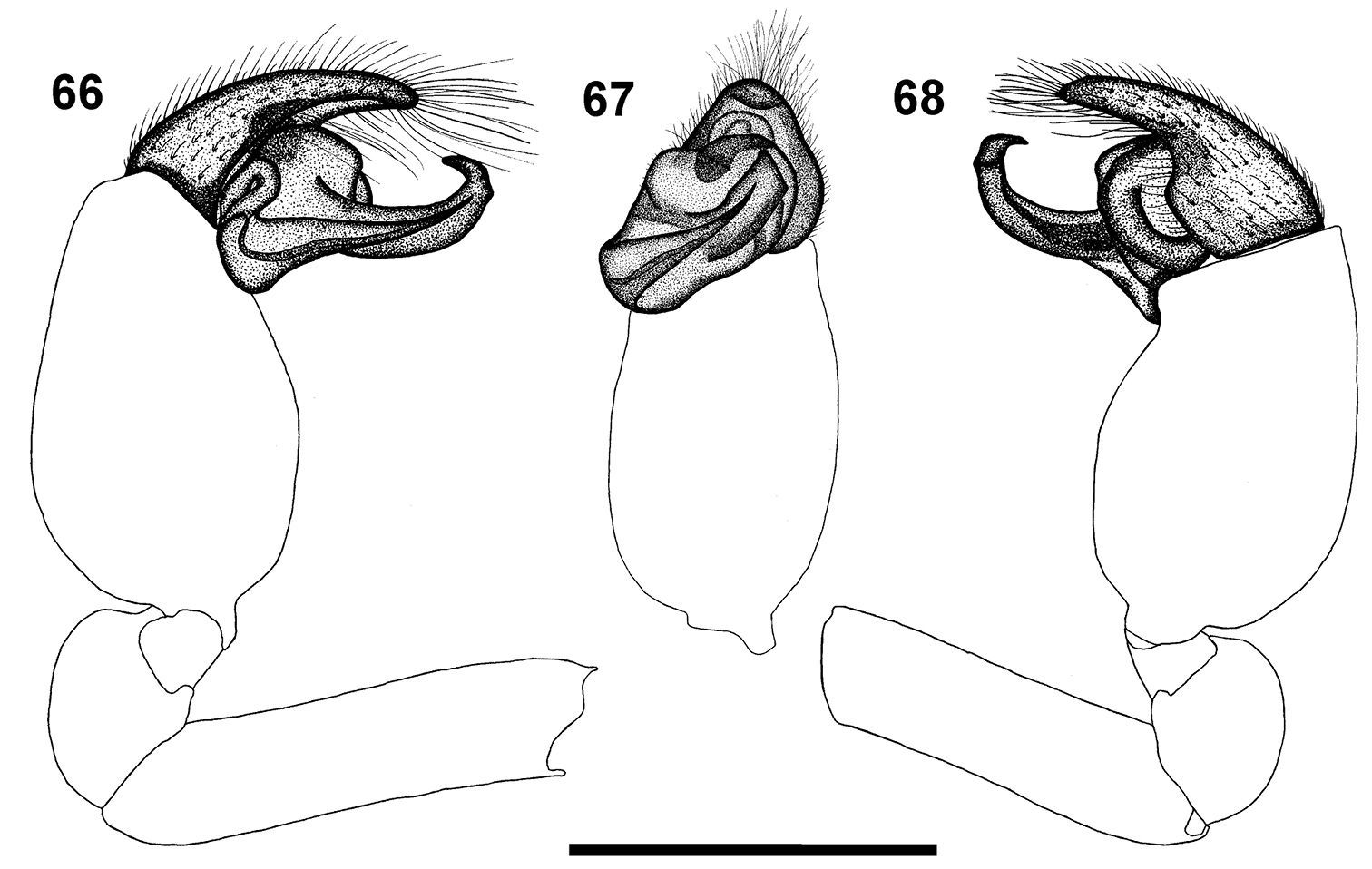
Left palp of male Calommata tibialis sp. n. 66 prolateral view 67 ventral view 68 retrolateral view. Scale bars: 1mm.
The specimens may possibly have faded over time in 70% ethanol, which can only be confirmed should fresh material become available. Although a subadult female is available it will not be described as the genitalic structure cannot be studied.
Central Côte d’Ivoire and northern Togo (Fig. 73).
Present data indicates that Calommata tibialis sp. n. occurs in woodland savannah habitats and avoids forests, where Calommata simoni
has been collected. This may indicate some degree of ecological
separation between the species but requires further study. Taking this
into account we consider the studies of
The burrow of Calommata tibialis sp. n. slants obliquely downwards into the soil, and was estimated to be 25–30cm deep by
http://species-id.net/wiki/Calommata_transvaalica
Figs 7, 8, 18, 19, 69–72SOUTH AFRICA: Gauteng Province: Pretoria, Roodeplaat [25°38'S, 28°21'E], 3.IV.1915, G. van Dam (TMSA 2999 – examined).
SOUTH AFRICA: Gauteng Province: 1♂: Groenkloof Nature Reserve [25°47'S, 28°12'E], 21.I.2003, reptile trap, M. Forsythe (NCA 2004/750); 1♀: Pretoria district, Hatfield [25°45'S, 28°15'E], 25.IV.1915, G. van Dam (TMSA 4639); 13♂: Zwartkoppies Farm 364, Portion 2, ca. 21km E of Pretoria, 25°45'23.6"S, 28°24'47.7"E, 1347m a.s.l., 29.X.2010, open pitfall traps, I. Engelbrecht & GDARD Field Staff (TMSA 23875). Limpopo Province: 1♂: Blouberg Nature Reserve, 23°00.065'S, 29°03.855'E, 29.XI.2005, searching below the knee, Philenoptera violaceae, A. Dawood (NCA 2009/3665); 1♀: Soutpansberg district, Blouberg, Wilhan’s Hohe [not traced], 28.VIII.1923, G. van Dam (TMSA 2772); 1♀: Same data, 29.VIII.1923 (TMSA 2773).
The female of this species has an additional row of two to four large prolateral teeth close to the fang base in addition to the main row of teeth, which are larger and more strongly curved than in Calommata simoni. The epigyne comprises a single pair of transversely oval spermathecae, while Calommata simoni possesses two pairs of smaller spermathecae. The male of the species shares with Calommata meridionalis the transversely orientated embolus and conductor, but the conductor of Calommata transvaalica is clearly narrower at the tip and the embolus is straight and not slightly curved as in Calommata meridionalis. The male chelicerae of Calommata transvaalica also lack the one or two large teeth found near the fang base in Calommata meridionalis.
Female from Blouberg Nature Reserve. Measurements: CL 6.72, CW 5.80, AL 13.70, AW 10.50, TL 25.20 (18.60–27.00). Length of leg segments, and total: I 1.85 + 0.60 + 0.74 + 0.85 + 0.60 = 4.64; II 1.60 + 1.05 + 0.69 + 0.90 + 0.72 = 4.96; III 1.45 + 1.28 + 0.60 + 0.80 + 0.55 = 4.68; IV 1.58 + 1.41 + 0.90 + 0.90 + 0.47 = 5.26. Carapace index 1.16; patella-tibia index 0.20.
Robustly built with short legs (Fig. 7), carapace pale creamy brown. Median ocular tubercle raised, narrow, sloping sharply at fovea. Single median line running from front of median ocular tubercle to middle of chilum. Chelicerae orange, darker laterally; chelicerae with a row of two to four large teeth close to fang base, prolateral of promarginal teeth row; teeth row comprising very large teeth curved at tips, interspersed with small teeth, with extensive denticle field retrolateral of teeth row near cheliceral base (Fig. 18). Endites strongly elongated and slender prolaterally, strongly curved upwards. Sternum and legs pale yellow-brown. Legs short and stout, leg formula 4231; legs III and IV more robust than legs I and II; leg I with three to five spines on patellae and two on tibiae; leg II with few spinules on patellae and several spinules on tibiae and metatarsi; legs III and IV with spinules from patellae to tarsi (mainly dorsal and prolateral); legs II to IV covered in bristles. Abdomen globose and pale grey, with indistinct median heart marking anteriorly (Fig. 7). Epigyne forming a broad, weakly sclerotised plate ventrally, in dorsal view with single pair of large, transversely oval spermathecae (Fig. 69). Female palp short, tibiae and tarsi flattened.
Male from Groenkloof Nature Reserve. Measurements: CL 2.10, CW 1.85, AL 2.70, AW 1.70, TL 6.40 (6.20–6.40). Length of leg segments, and total: I 1.66 + 0.60 + 1.10 + 1.43 + 1.28 = 6.07; II 1.67 + 0.70 + 0.95 + 1.50 + 1.47 = 6.29; III 1.45 + 0.70 + 0.70 + 1.55 + 1.95 = 6.35; IV 1.85 + 0.78 + 1.05 + 1.85 + 1.90 = 7.43. Carapace index 1.14; patella-tibia index 0.81.
Carapace and chelicerae dark brown in colour (Fig. 8). Chelicerae with single row of large teeth, gradually decreasing in size from fang base to cheliceral base, without denticles near cheliceral base (Fig. 19). Carapace oval in shape. Median ocular tubercle raised, narrow, darker in colour. Sternum and coxae yellow-brown, femora, patellae and tibiae brown, metatarsi yellow-brown, tarsi yellow. Legs weakly covered with bristles; prolateral side of patellae, tibiae and metatarsi of legs II–IV covered with spinules. Abdomen dark grey, nearly black, with dark orange-brown scutum anteriorly (Fig. 8). Palp with short cymbium, cymbium tip in ventral view tapering to rounded point; embolus and conductor orientated transversely to palpal axis, pointing retrolaterally, distal ends projecting beyond retrolateral cymbial margin; conductor short, slightly broadened distally, with single sharp, curved tooth on its dorsal surface; embolus long and straight (Figs 70–72).
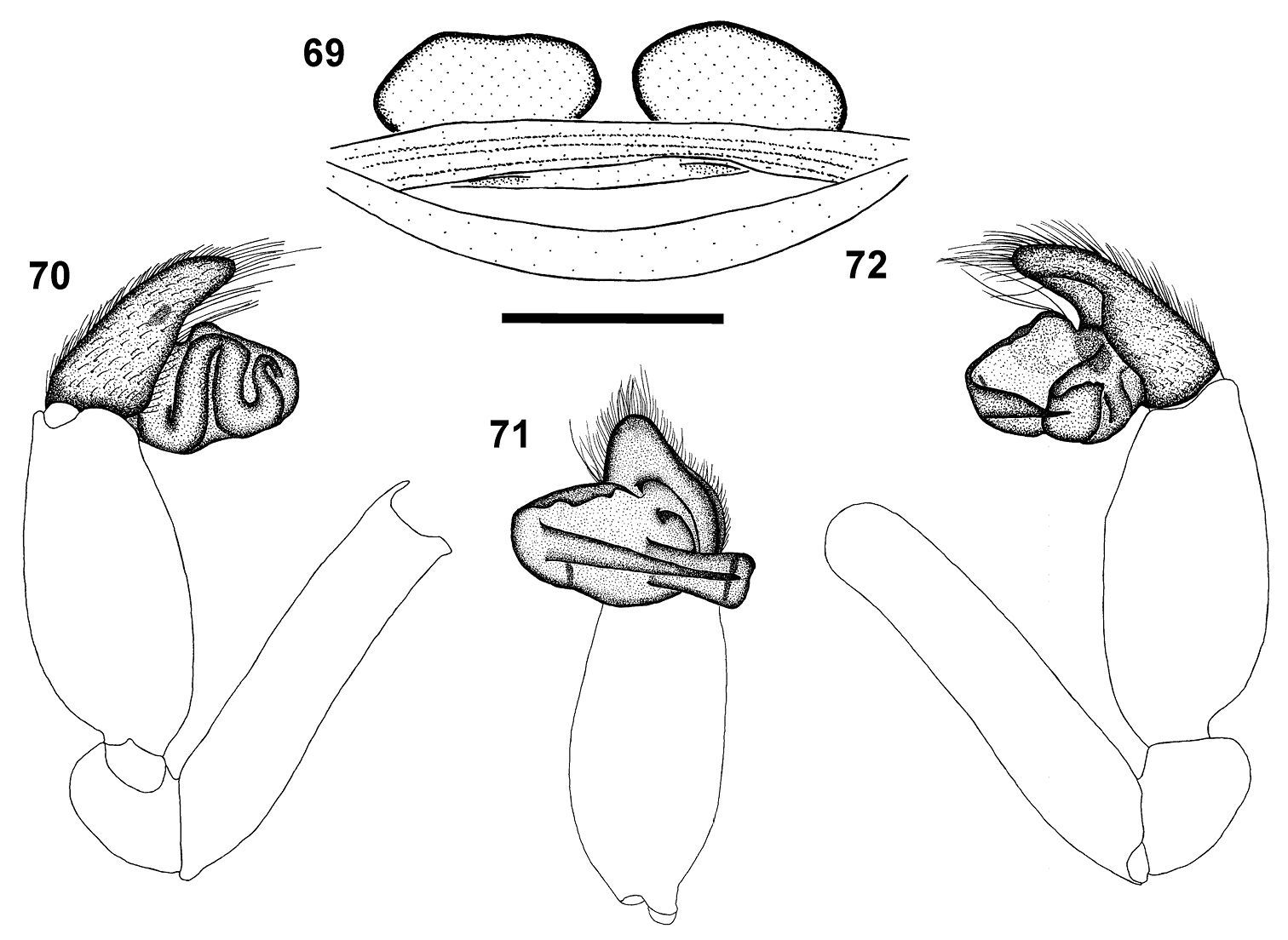
Calommata transvaalica Hewitt female genitalia 69 and left palp of male 70–72) 69 dorsal view 70 prolateral view 71 ventral view 72 retrolateral view. Scale bars: 1mm.
It is clear from the redescriptions of both sexes of Calommata simoni, and the redescription of the female and first description of the male of Calommata transvaalica
provided here, that the two species have distinct differences in their
somatic and genitalic morphology, most notably regarding their
cheliceral dentition, number of spermathecae in the female epigyne and
orientation and length of the male embolus and conductor. Consequently,
we reject Benoit’s (1967) synonymy of the two species and propose the
revalidation of Calommata transvaalica. The specimens collected by
The abdomen of the female holotype of Calommata transvaalica is damaged and therefore the specimen was not redescribed. The holotype is the smallest of the known females of this species (18.60mm long). The colour of all available female specimens has likely faded over time in 70 % ethanol.
Limpopo and Gauteng Provinces, South Africa (Fig. 73).
Biology. The biology of Calommata transvaalica was studied by
The recently collected series of males from Zwartkoppies Farm (TMSA 23875) was collected in open pitfalls without preservative from a site in open Acacia karroo woodland on red structured clay soils (30–45% clay in A horizon, Shortlands form). The site has a gentle slope and had recently been burned. The activity of the males appears to be related to heavy rainfall that had fallen two days prior to the collection of the males. It thus seems that males do not emerge on the night immediately following a heavy shower, but instead on the night thereafter (Ian Engelbrecht, pers. comm.).
Calommata is a small but widespread genus that is known from Africa, Israel and South East Asia. Two species are present in West Africa, one species in Central Africa and four species in southern Africa. The type of habitat seems to play a role in separating the species biogeographically. Calommata simoni is perhaps the most flexible in terms of habitat requirements, and occurs in forests, savannahs and grasslands in tropical Africa. In contrast, Calommata tibialis was only found in wooded savannahs in West Africa. In southern Africa, Calommata megae, Calommata namibica and Calommata transvaalica occur in the Savannah biome, but with contrasting vegetation structures and climatic variables. Lastly, Calommata meridionalis occurs only in the Grassland biome of central South Africa (Fig. 73).
Atypid spiders are widely regarded as being of
conservation importance due to their generally specific environmental
requirements, low rates of dispersal and general scarcity (
Calommata transvaalica (previously as Calommata simoni
in South Africa) was until recently presumed nationally extinct in
South Africa as the species had last been reported in the 1920’s (
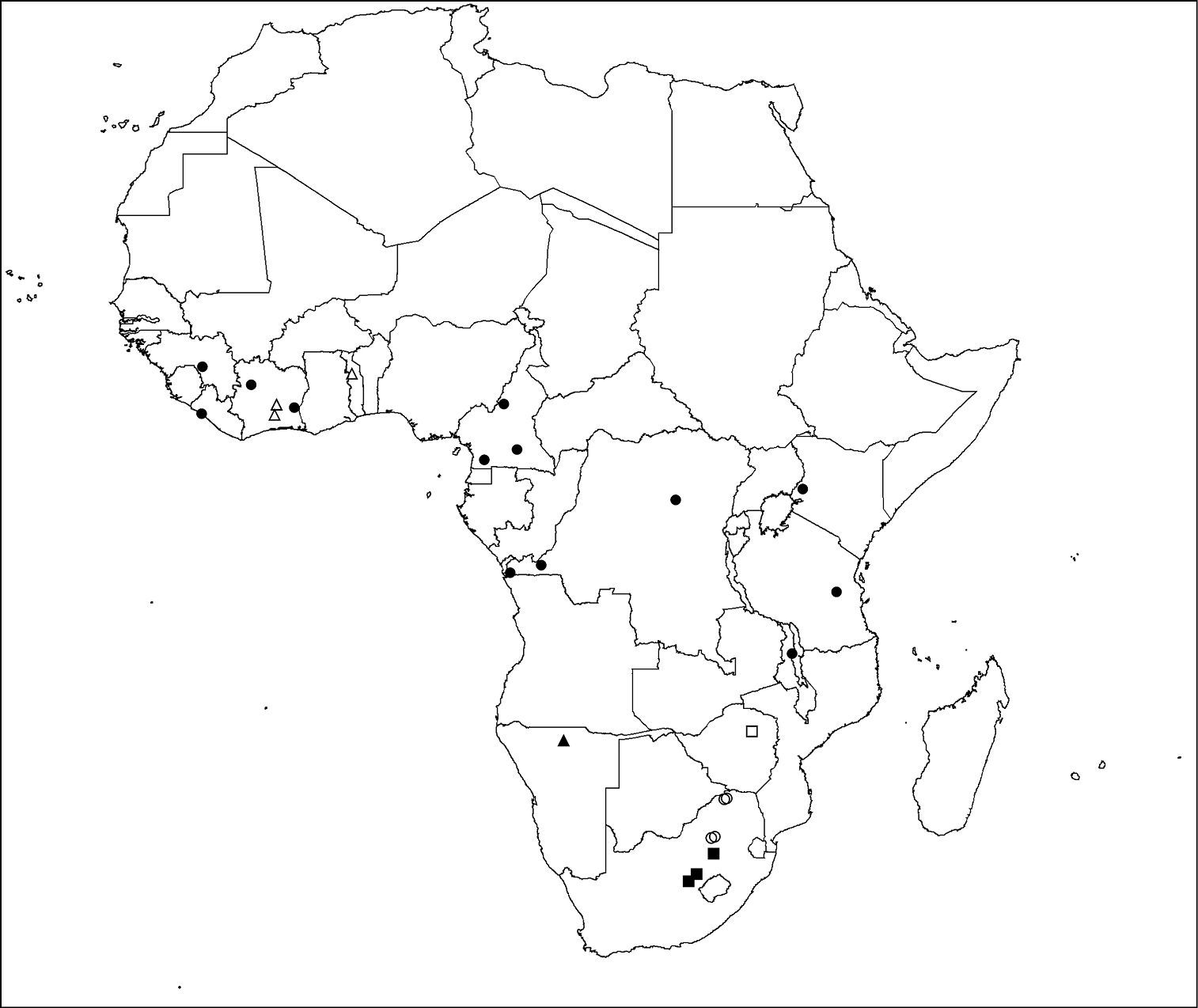
Distribution of Calommata species in the Afrotropical Region: Calommata megae (open square), Calommata meridionalis (solid squares). Calommata namibica (solid triangle), Calommata simoni (solid circles), Calommata tibialis (open triangles) and Calommata transvaalica (open circles).
This study forms part of the M.Sc of the senior author at the University of the Free State and was funded through a National Research Foundation of South Africa (NRF) grant in the NRF Thuthuka program to the second author (TK2008050500003). The NRF also provided a student travel grant to the senior author through its SABI program to visit the MRAC (GUN 69525). The following curators of the collections are thanked for the loan of material that made this study possible: Norman Platnick (AMNH), Janet Beccaloni (BMNH), Ansie Dippenaar-Schoeman (NCA), Leon Lotz (NMBA), Audrey Ndaba (NMSA), Robin Lyle (TMSA) and Jonathan Coddington (USNM). Ansie Dippenaar-Schoeman is thanked for providing some rare taxonomic and biological literature, and she and Ian Engelbrecht are thanked for useful discussion on the conservation importance of the South African species. Meg Cumming kindly provided biological information on the Calommata megae specimens that she collected in Zimbabwe and Ian Engelbrecht on Calommata transvaalica specimens collected in South Africa. Field work by Ian Engelbrecht was funded by the Gauteng Dept. of Agriculture and Rural Development and Duncan MacFadyen of E. Oppenheimer & Son. Richard Gallon and Jeremy Miller are thanked for their constructive comments that helped improve the manuscript.