





(C) 2010 Sergei I. Golovatch. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Polydesmidae are represented in Taiwan by seven species in two genera. Neither of the genera is endemic to Taiwan, but six of the species are, including five new: Nipponesmus minor sp. n., Epanerchodus bispinosus sp. n., Epanerchodus curtigonopus sp. n., Epanerchodus flagellifer sp. n. and Epanerchodus pinguis sp. n. In addition, the diagnosis of the hitherto enigmatic genus Nipponesmus Chamberlin & Wang, 1953 is refined vis-à-vis the especially similar, Central Asian, Siberian and Eastern European genus Schizoturanius Verhoeff, 1931, chiefly based on new material of the type-species Nipponesmus shirinensis Chamberlin & Wang, 1953; this species is adequately redescribed and represents still another Taiwanese endemic. A key to all three currently known species of Nipponesmus Chamberlin & Wang, 1953 is given. The highly speciose Central to East Asian genus Epanerchodus Attems, 1901 is represented in Taiwan by five species, all keyed, including Epanerchodus orientalis Attems, 1901, which is long known to be highly variable in Japan and found particularly polymorphous and apparently allochthonous in Taiwan. The following synonymy is formalized: Epanerchodus orientalis orientalis Attems, 1901 = Epanerchodus orientalis takakuwai Verhoeff, 1913, syn. n. The genus Usbekodesmus Lohmander, 1932 is formally synonymized with Epanerchodus Attems, 1901, syn. n., resulting in the following new formal transfers: Epanerchodus redikorzevi (Lohmander, 1932), Epanerchodus swatensis (Golovatch, 1991), Epanerchodus varius (Geoffroy & Golovatch, 2004), Epanerchodus anachoretus (Golovatch, 1986), Epanerchodus buddhis (Golovatch, 1986), Epanerchodus occultus (Golovatch, 1986), Epanerchodus sacer (Golovatch, 1987), Epanerchodus theocraticus (Golovatch, 1990) and Epanerchodus theosophicus (Golovatch, 1986), all comb. n. ex Usbekodesmus. The distributions of all seven species of Polydesmidae occurring in Taiwan are mapped and discussed.
millipede, Polydesmidae, taxonomy, new species, new synonymy, key, Taiwan
The millipede fauna of Taiwan is still quite poorly explored (
Prompted by the abundant new material collected during the last couple of decades all over the island, below we offer a revision of the Taiwanese Polydesmidae. Already seven species appear to be involved, representing two genera. These new samples also allow for the diagnosis of one genus to be refined, and the synonymy of another one formalized.
Material and methodsMaterial serving as the basis for the present contribution was preserved in 75% alcohol and is currently shared between the collections of the National Museum of Natural Science, Taichung, Taiwan (NMNS), Taiwan Forest Research Institute, Taipei, Taiwan (TFRI), Department of Biological Sciences, National Sun Yat-Sen University, Kaohsiung, Taiwan (NSYSU), Zoological Museum, State University of Moscow, Russia (ZMUM), Natural History Museum of Denmark, University of Copenhagen, Denmark (ZMUC), Muséum national d’Histoire naturelle, Paris, France (MNHN), and Institute of Biology and Soil Science, Far Eastern Branch, Russian Academy of Sciences, Vladivostok, Russia (IBSS), as indicated hereafter. Specimens were studied and illustrated using standard stereomicroscopic, photographic and drawing equipment.
In the catalogue sections, which mostly refer to the fauna of Taiwan alone, D stands for the original description, N for additional descriptive notes, and R for a mere mention or record.
Systematicshttp://species-id.net/wiki/Nipponesmus_shirinensis
Figs 1–132 juv. (NSYSU), Taiwan, Taipei City, Wenshan Distr., Chih-Nan Temple, 03.2002, leg. C.C. Chen et al.; 2 ♂, 2 ♀ (NMNS-6555-001), 1 ♂ (MNHN JC 331), Nantou County, Re-nai Township, Mei-Feng, 22.10.2001; 1 ♂ (ZMUC), 1 ♂ (NMNS-6555-002), same locality, 15.10.2001; 1 ♂ (TFRI), same locality, 19.02.2002; 1 ♂ (NMNS-6555-003), same locality, 15.10.2001; 1 ♂ (ZMUM), Nantou County, Huisun timber land, 20.09.1997; 1 ♂ (NMNS-6555-004), same locality, 10.1997; all leg. S.H. Wu; 3 ♂, 5 ♀ (TFRI), Nantou County, Lugu Township, Sitou, 15.11.2005, leg. J.D. Lee; 8 juv. (NSYSU), same locality, 6.07.2010, leg. H.W. Chang; 1 ♂, 1 ♀ (TFRI), Taitung County, Taimali Township, I-Ma forest road, 06.12.2004, leg. S.Y. Wu; 4 ♂, 1 ♀, 5 juv. (NSYSU), Chiayi County, Alishan Township, Nansi Forest road, ca 2, 000 m a.s.l., 29.10.2010, leg. H.W. Chang; 1 ♂ (ZMUM), Hsinchu County, Wufeng Township, Sakaru (Shilu) trail, 30.09.2005; 1 ♂ (NMNS-6555-005), same locality, 22.09.2005, all leg. H.D. Zhu; 1 ♂ (NSYSU), Kaohsiung County, Liouguei, Shanping Workstation, 04.2004, leg. M.J. Hung; 1 ♂ (NMNS-6555-006), Kaohsiung County, Taoyuan Township, Tengjhih, 1, 550 m a.s.l., 17.11.2010, leg. S. Golovatch; 2 ♀ (NMNS-6555-007), Kaohsiung County, Taoyuan Township, Chungchihkuan, 15.03.2005, leg. Y.C. Chang; 1 juv. (NSYSU), same locality, Taiwania cryptomeroides plantation, 15.10.2005, leg. M.H. Hsu; 1 ♂, 1 ♀ (IBSS), Pingtung County, Chunri Township, Mt Dahan, 1, 200 m a.s.l., 15.12.2009; 2 ♀ (NMNS-6555-008), same locality, ca 1, 850 m a.s.l., 15.12.2009, all leg. M.H. Hsu.
Differs from the other Polydesmidae in Taiwan by the larger body and often a dark coloration, from the only sympatric congener Nipponesmus minor sp. n. also in the collum being broader than the head and in the gonopod showing a strongly elongate, slender and falcate telopodite (see also Key below).
Length of both sexes ca 18–25 mm, usually about 20–23 mm; width of pro- and metazona varying between specimens from 1.5–2.2 to 2.9–4.8 mm, respectively, usually 1.8–2.0 and 3.0–4.3 mm, respectively. Usually ♂♂ somewhat smaller than ♀♀. Coloration in alcohol from uniformly pallid (faded?) to dark chocolate-brown; in the latter case, sides, venter and legs (light) grey-brown to dark brown (Figs 1–6). Body with 20 segments. Tegument mainly dull, at most slightly shining, texture very delicately alveolate. Frons and labrum densely pilose, vertex bare; epicranial suture distinct but thin; a paramedian pair of evident, oblique ridges above antennal sockets; isthmus between antennae considerably broader than diameter of antennal socket (Fig. 5). Antennae rather long and only slightly clavate (Fig. 5), either slightly overreaching segment 3 dorsally (♂) or slightly shorter (♀); antennomere 3 longest (Figs 3–5); antennomeres 5 and 6 each with a small, compact, distodorsal group of bacilliform sensilla; antennomere 7 with a minute dorsoparabasal cone and a distodorsal group of microscopic sensilla.
In width, head < collum < segment 2 < 3 < 4 < 5=15(16), thereafter body gradually tapering towards telson. Paraterga strongly developed, starting from collum, subhorizontal, set high but always lying slightly below a faintly convex dorsum, drawn clearly forward only on metatergum 2 (Fig 3). Starting from segment 8 or 9 (♂) or 16 (♀), paraterga extending increasingly beyond rear tergal contour, caudal corners invariably evidently rounded, only on paraterga 17–19 subspiniform, usually slightly more narrowly rounded in ♂ compared to ♀ (Figs 3, 6–9). Paraterga more (♀) or less (♂) evidently rounded laterally (Figs 3, 6–9); starting from segment 2, all poreless segments with three, all pore-bearing ones with four, minute, sometimes even obliterate incisions at lateral margin. Front edges of metaterga slightly bordered and upturned, straight, usually forming a distinct shoulder. Pore formula normal, ozopores evident, dorsal, located in front of posteriormost marginal indentation. Metatergal sculpture typical, well-developed, with three transverse rows of setiferous, polygonal bosses (Figs 3, 6–9). Tergal setae very short, mostly obliterate, partly retained only on collum and/or metatergum 19. Stricture between pro- and metazona wide, shallow and smooth (Figs 3, 4, 6–9). Limbus very thin, microdenticulate. Pleurosternal carinae absent (Fig. 4). Epiproct rather short, conical, preapical papillae very evident (Figs 7, 9). Hypoproct semi-circular; caudal, paramedian, setiferous papillae small and well-separated.
Sterna without modifications, very densely (♂) (Fig. 5) or poorly (♀) setose, ♀ ones often shining. Epigynal ridge very low. Legs rather long and slender (Figs 5, 10), ca 1.5–1.6 (♂) or 1.1–1.2 (♀) times as long as midbody height; ♂ legs evidently enlarged, prefemora only slightly swollen dorsally and, like femora and, partly, postfemora, ventrally beset with shorter bifid setae turning into short sphaerotrichomes on postfemora, tibiae and tarsi (Fig. 10).
Gonopods (Figs 4, 5, 11–13) with large, subquadrate, medially fused coxae carrying a few long setae ventrally. Telopodite elongated, slender, falcate, prefemoral (densely setose) portion almost half as long as entire telopodite; seminal groove running mesally over most of its extent, only distally moving frontally to recurve first laterad and then a little basad at base of both a somewhat shorter, more complex endomere (en) and a longer, simpler exomere (ex); en beset with long, bacilliform setae distally and supplied with an evident spine (s) mesally, as well as a prominent pulvillus near base, this pulvillus being likewise beset with bacilliform setae and marking the end of seminal groove devoid of any accessory seminal chamber; ex strongly unciform apically, with an additional lateral tooth distally to subapically.

Nipponesmus shirinensis Chamberlin & Wang, 1953, ♀ (1) and ♂ (2) from Mei-Feng, ♂ (3–5) from Tengjhih. 1, 2 habitus, lateral view 3–5 anterior portion of body, dorsal, lateral and ventral views, respectively. Photographed not to scale.

Nipponesmus shirinensis Chamberlin & Wang, 1953, ♂ (6, 7) from Tengjhih and ♀ (8, 9) from Mei-Feng. 6 middle portion of body, dorsal view 7, 9 posterior portion of body, dorsal view 8 anterior portion of body, dorsal view. Photographed not to scale.

Nipponesmus shirinensis Chamberlin & Wang, 1953, ♂ from Sitou. 10 leg 9 11, 12 right gonopod, lateral and mesal views, respectively 13 distal part of gonopod telopodite, enlarged, mesal view. Scale bar: 0.5 mm (10–12) and 0.25 mm (13). See text for explanation of labels.
Remarks. This species is the largest, as well as one of the most conspicuous and common among the polydesmids in Taiwan. It inhabits a wide range of habitats, mainly montane woodlands above 1, 200 m a.s.l. (Map 1).
urn:lsid:zoobank.org:act:225E19E9-CF6B-46A1-A16F-7EFAD3A91A58
http://species-id.net/wiki/Nipponesmus_minor
Figs 14–23Holotype ♂ (TFRI), Taiwan, Ilan County, Tatong Township, near Lakes Jialuohu, ca 2, 250 m, 27.04.2003, leg. Y.M. Chen. Paratypes: 3 ♂, 1 ♀ (TFRI), same locality, 14.03.2003; 12 ♂, 8 ♀, 1 fragm. (TFRI), same locality, 28.04.2003; 1 ♂ (TFRI), same locality, 27.04.2003; 2 ♂ (ZMUM), 1 ♂ (IBSS), 2 ♂ (NSYSU), same locality, 25.12.2002; 1 ♂ (ZMUC), 1 ♂, 1 ♀ (TFRI), same locality, 7.11.2001; 1 ♂, 1 ♂ fragm. (TFRI), same locality, 4.06.2003, all leg. Y.M. Chen; 1 ♂ (ZMUM), same locality, 24.10.2002, leg. J.T. Chao; 1 ♂ (TFRI), Taichung County, Shengguang, 24.09.2002; 1 ♂ (TFRI), same locality, 24.01.2003; 1 ♀ (TFRI), same locality, 26.03.2003; 1 ♂ (MNHN JC 332), same locality, 24.01.2003; 1 ♂ (TFRI), same locality, 24.09.2002, all leg. W.C. Yeh; 1 ♂ (NMNS-6556-001), Nantou County, Ren-ai Township, Mei-Feng, 19.02.2002; 1 ♂ (NMNS-6556-002), same locality, 11.07.2002; 4 ♂ (NMNS-6556-003), same locality, 15.04.2002, all leg. S.H. Wu.
To emphasize the smaller body size and the shorter gonopod telopodite.
Differs from Nipponesmus shirinensis, the only other congener known from Taiwan, in the smaller size, as well as in the collum being narrower than the head and in the gonopod telopodite being stouter and shorter (see also Key below).
Length of both sexes ca 12–16 mm; width of pro- and metazona varying between specimens from 0.8–1.3 to 1.5–2.0 mm, respectively. Holotype ca 12 mm long, and 0.8 and 1.5 mm wide on pro- and metazona, respectively. Usually ♂♂ somewhat smaller than ♀♀. Coloration in alcohol from uniformly pallid (faded?) to yellowish to reddish-brown; in the latter case, sides, venter and legs light grey-brown (Figs 14–19).
All characters as in Nipponesmus shirinensis, except as follows.
Antennae a little shorter, usually reaching midway of segment 3. In width, collum < head < segment 2 < 3 < 4 < 5=15(16), thereafter body gradually tapering towards telson. Starting from segment 16 (♂, ♀), paraterga extending increasingly beyond rear tergal contour, caudal corners invariably evidently rounded, but even on paraterga 17–19 not spiniform (Figs 15, 18). Paraterga less evidently rounded laterally even in ♀, lateral edges mostly subparallel in ♂ (Figs 15, 18), incisions being visible. Metatergal sculpture typical, rather superficial, with three transverse rows of setiferous, polygonal bosses (Figs 15, 18). Tergal setae very short, usually retained at least on collum and/or metatergum 19 (Figs 15, 16, 18).
Gonopod telopodite (Figs 20–23) much stouter; endomere (en) slightly longer than exomere (ex), beset with long, bacilliform setae nearly throughout, usually supplied with an evident spine (s) or tooth mesally, as well as a usually somewhat less prominent pulvillus near base; ex often not so strongly unciform apically, often with an additional lateral tooth in distal part.
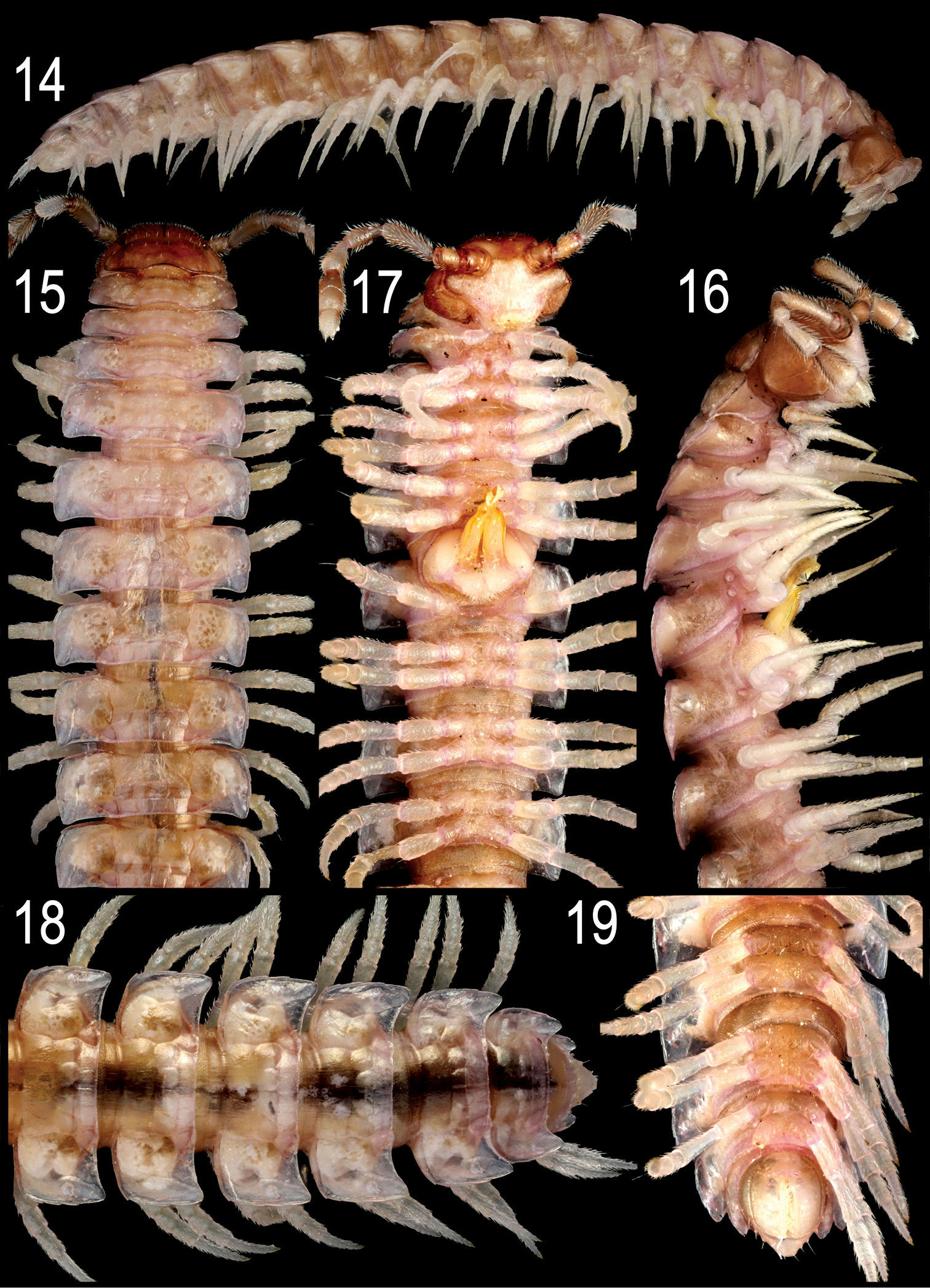
Nipponesmus minor sp. n., ♂ paratype from Jialuohu. 14 habitus, lateral view 15–17 anterior portion of body, dorsal, lateral and ventral views, respectively 18, 19 posterior portion of body, dorsal and ventral views, respectively. Photographed not to scale.
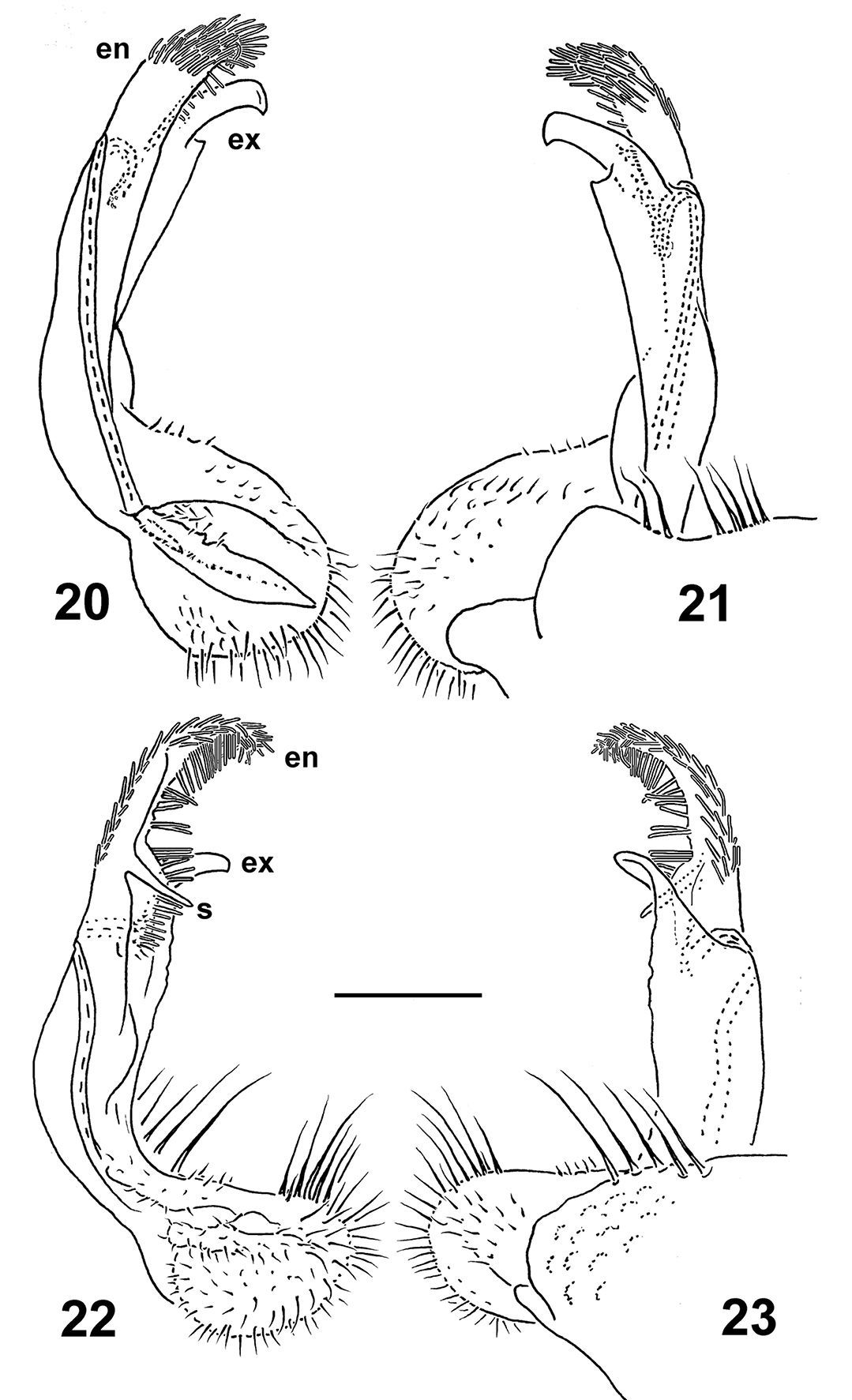
Left gonopods of Nipponesmus minor sp. n., ♂ paratypes from Mei-Feng (20, 21) and Jialuohu (22, 23), mesal, lateral, mesal and lateral views, respectively. Scale bar: 0.2 mm.
This species is not so widely distributed in Taiwan. Allopatry is prevailing, as it is only at Mei-Feng that both the congeners co-occur (Map 1). However, Nipponesmus minor sp. n. quite often lives even syntopically together with Epanerchodus orientalis (see below).
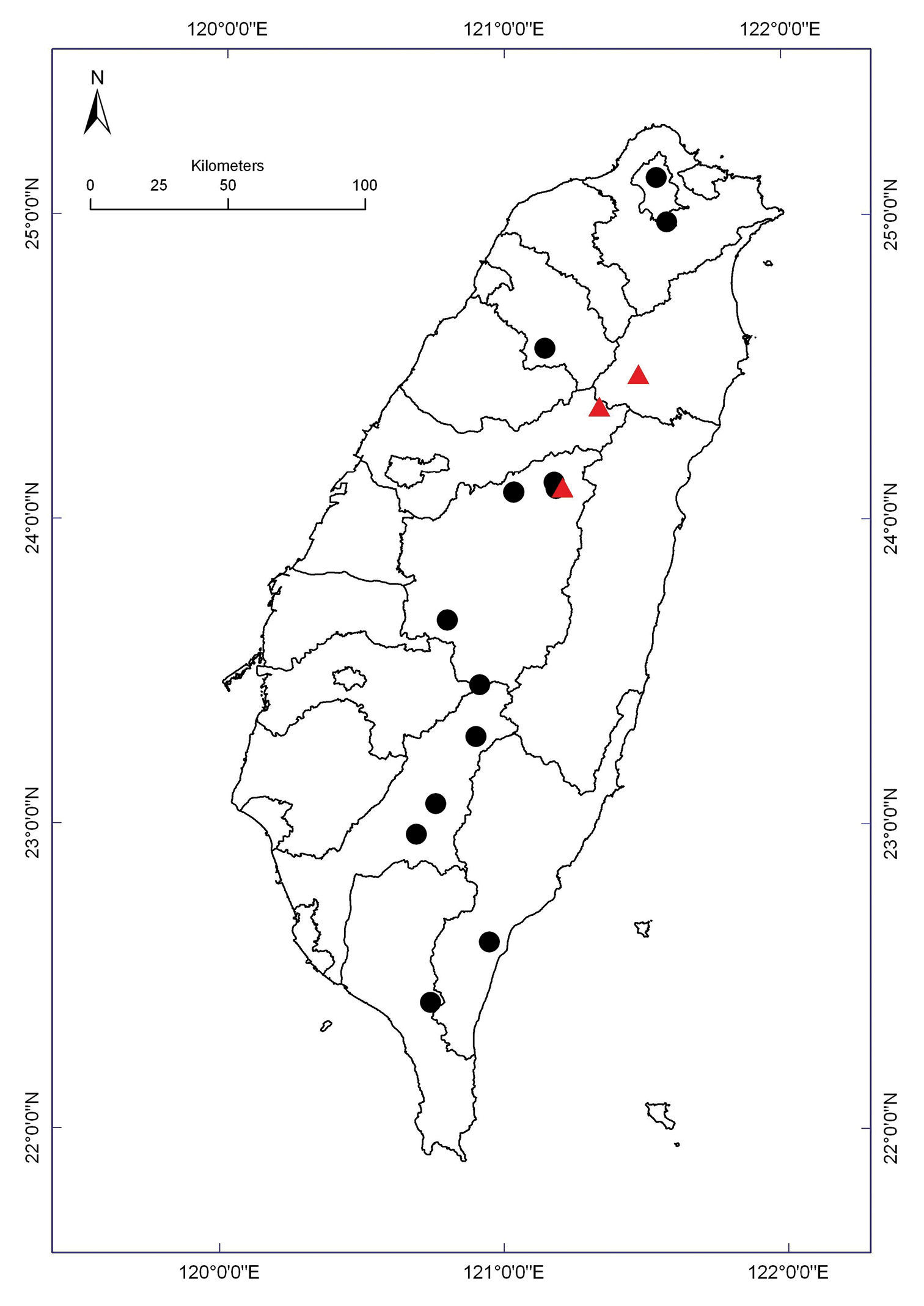
Distribution of Nipponesmus species in Taiwan. Borderlines show borders between the counties. Nipponesmus minor sp. n.: filled red triangles; Nipponesmus shirinensis: filled black circles.
A study of the ample material representing not only the type species Nipponesmus shirinensis, but also the above new congener from Taiwan allows for the identity of Nipponesmus Chamberlin & Wang, 1953 to finally become clarified.
This genus has hitherto remained enigmatic, originally described too poorly (
In Nipponesmus shirinensis, the gonopod telopodite (Figs 11–13) is indeed biramous only distally, being divided into subequally prominent endo- and exomere. Furthermore, the endomere (en) is beset with a characteristically bacilliform trichome, whereas the exomere (ex) is simple. The seminal groove runs mostly mesally to recurve neatly between ex and en
and then to debauch somewhat basally into a prominent hairy pulvillus
which is also beset with the same peculiar trichome, and is devoid of
an accessory seminal chamber. The same general pattern is observed both
in Nipponesmus tangonis and Nipponesmus minor (Figs 20–23). What we term here as endomere is the branch which
As regards Golovatch’s (1991) placement of Nipponesmus also near Epanerchodus, following
This results in the following new formal transfers: Epanerchodus redikorzevi (Lohmander, 1932), from Uzbekistan, Tajikistan and Afghanistan, Epanerchodus swatensis (Golovatch, 1991), from Swat Province, northern Pakistan, Epanerchodus varius (Geoffroy & Golovatch, 2004), from Hubei Province, southern China, as well as the Nepalese Epanerchodus anachoretus (Golovatch, 1986), Epanerchodus buddhis (Golovatch, 1986), Epanerchodus occultus (Golovatch, 1986), Epanerchodus sacer (Golovatch, 1987), Epanerchodus theocraticus (Golovatch, 1990) and Epanerchodus theosophicus (Golovatch, 1986), all comb. n. ex Usbekodesmus.
The following key can serve to separate all three currently known species of Nipponesmus.
| 1 | Adult body smaller, ≤ 10 mm long and ≤ 1.4 mm wide. Collum as wide as head. Endomere strongly enlarged, lobe-shaped, fringed by a very dense bacilliform trichome; exomere slender, much longer than endomere. Honshu, Japan | Nipponesmus tangonis |
| – | Adult body larger, at least 12 mm long and ≥ 1.5 mm wide. Collum either broader or narrower than head. Endomere at most slightly enlarged, digitiform, beset with a dense bacilliform trichome nearly all over; exomere rather short and stout, at most only slightly longer than endomere. Taiwan | |
| 2 | Adult body 18–25 mm long and 1.55–2.0 mm wide. Collum broader than head. Gonopod telopodite considerably longer (Figs 11–13) | Nipponesmus shirinensis |
| – | Adult body 12–16 mm long and 2.9–4.8 mm wide. Collum narrower than head. Gonopod telopodite considerably stouter (Figs 20–23) | Nipponesmus minor sp. n. |
http://species-id.net/wiki/Epanerchodus_orientalis
Figs 24–36Complete catalogue information referring to this species can be found in
10 ♂ (TFRI), 1 ♂ (NMNS-6557-001), 1 ♀ (NSYSU), 2 ♂ (MNHN JC 333), Taiwan, Taichung County, Shengguang, 25.04.2003; 2 ♀ (TFRI), same locality, 26.03.2003; 1 ♀ (TFRI), same locality, 24.09.2002; 4 ♂ (TFRI), 1 ♂ (NMNS-6557-002), same locality, 24.07.2003, all leg. W.C. Yeh; 2 ♂, 1 ♀ (TFRI), 2 ♂, 1 ♀ (IBSS), Ilan County, Fushan, 23.03.2003; 2 ♀ (NMNS-6557-003), same locality, 18.05.2001; 5 ♂, 1 ♀ (TFRI), same locality, 19.06.2001; 1 ♂ (TFRI), same locality, without date, all leg. J.T. Chao; 1 ♂ (ZMUM), same locality, 20.11.2001, leg. S.S. Lu; 52 ♂, 17 ♀, 4 fragm. (TFRI), 2 ♂ (ZMUC), 2 ♂ (NSYSU), Ilan County, Tatong Township, near Lakes Jialuohu, ca 2, 250 m, 4.06.2003; 1 ♀ fragm. (TFRI), same locality, 28.04.2003; 8 ♂, 6 ♀ (TFRI), same locality, 20.06.2002; 1 ♂ (ZMUM), same locality, 25.04.2003; 1 ♂ (TFRI), same locality, 23.07.2002, all leg. Y.M. Chen; 1 ♀ (NSYSU), Taitung County, Yanping Township, Yanping Forest road, ca. 1, 250 m, 3.06.2003, leg. M.H. Hsu; 2 ♂ (NSYSU), Hualien County, Fongbin Township, Guangfong Highway, ca 300 m, 6.05.2009; 1 ♀ (NSYSU), same County, Shoufang Township, ca. 230 m a.s.l., 5.05.2009, all leg. M.H. Shu; 2 ♀ (NMNS-6557-004), Pingtung County, Chaozhou Township, Si-Lin, 1.06.1999, leg. ?; 1 ♀ (NMNS-6557-005), Kaohsiung County, Taoyuan, Provincial Road No. 20, near 142.5 km road-sign, 3.09.2009, leg. C.Y. Huang.
Differs from the other Epanerchodus species known from Taiwan in the variable, mostly medium size, coupled with the caudal corner of most of the paraterga being pointed, and in the gonopod telopodite being relatively slender, complex, highly variable in shape and armature (see also Key below).
Length of both sexes ca 11–19 mm; width of pro- and metazona varying between specimens from 0.8–1.8 to 1.2–3.0 mm, respectively. Usually ♂♂ somewhat smaller than ♀♀. Coloration in alcohol from uniformly pallid (faded?) to light (reddish- to grey-) brown; in the latter case, venter and legs light grey to yellow (Figs 24–31). Body with 20 segments. Tegument mainly dull, at most slightly shining, texture very delicately alveolate. Labrum, frons and vertex densely pilose; epicranial suture clear but thin; no ridges above antennal sockets; isthmus between antennae considerably broader than diameter of antennal socket (Fig. 29). Antennae long and only slightly clavate (Figs 24, 25, 28), reaching behind end (♂) or midway (♀) of segment 4 dorsally; antennomere 3 longest, 5th highest (Figs 25, 28); antennomeres 5 and 6 each with a small, compact, distodorsal group of bacilliform sensilla; antennomere 7 with a minute dorsoparabasal cone and a distodorsal group of microscopic sensilla.
In width, either collum ≤ segment 2 = 3 < 4 < head = 5(6)-15(16), or segment 2 = 3 < collum < head = 4 < 5–15(16) (both sexes), thereafter body gradually tapering towards telson. Paraterga strongly developed, starting from collum, subhorizontal, set high but always lying slightly below a faintly convex dorsum; front shoulders and caudal edge nearly straight on several postcollum metaterga, caudal corner always pointed. Collum (Fig. 28) nearly regularly elliptical, with an incision near caudal corner and three transverse rows of setae (6+6, 4+4 and 3+3). Starting from segment 5, paraterga extending increasingly beyond rear tergal contour, mostly subspiniform (Figs 25–28). Starting from segment 2, all poreless segments with three, all pore-bearing ones with four, small but evident incisions, each usually bearing a small seta on top at lateral margin. Pore formula normal, ozopores evident, dorsal, located in front of 4th indentation. Metatergal sculpture typical, rather well developed, with three transverse rows of setiferous, polygonal bosses (Figs 25–28). Tergal setae short, mostly retained, a little longer only on collum and in rear row on metatergum 19 (Figs 25, 27). Stricture between pro- and metazona rather obscure, wide, smooth and polished. Limbus very thin, microdenticulate. Pleurosternal carinae absent. Epiproct rather short, conical (Fig. 31), preapical papillae evident. Hypoproct semi-circular; caudal, paramedian, setiferous papillae evident and well-separated.
Sterna without modifications (Figs 29, 31), densely setose. Epigynal ridge very low, regularly rounded. Legs long and slender (Figs 28, 29), ca 1.8–1.9 (♂) or 1.2–1.3 (♀) times as long as midbody height; ♂ legs evidently enlarged, prefemora only slightly swollen dorsally, femora ventro-parabasally with shorter bifid setae replaced by sphaerotrichomes in remaining parts of femora, as well as on entire postfemora, tibiae and tarsi.
Gonopods (Figs 32–36) with large, subquadrate, medially fused coxae carrying a few long setae ventrally. Telopodite stout to rather slender, subfalcate, prefemoral (densely setose) portion one-third to half as long as entire telopodite; seminal groove running mesally over much of its extent, only distally moving frontally to recurve first laterad and then mesad, squeezing neatly between a simple, more or less rudimentary to completely reduced exomere (ex) and a more complex, branching and always well-developed endomere (en), then groove continuing to a considerable extent basad to end into a large accessory seminal chamber lying near base of a prominent excavation, the latter carrying an evident hairy pulvillus; en often but not alwayswith a strong mesal process (s), either slenderer and simple or larger and more complex in shape, as well as usually with a distinct, often again branching, laterobasal outgrowth (p); distal remainder of en elongate, often branching and enlarged, sometimes fringed with short setae or spines.

Epanerchodus orientalis Attems, 1901, ♂ from Fushan. 24 habitus, lateral view 25 anterior portion of body, subdorsal view 26 middle portion of body, dorsal view 27 posterior portion of body, dorsal view. Photographed not to scale.
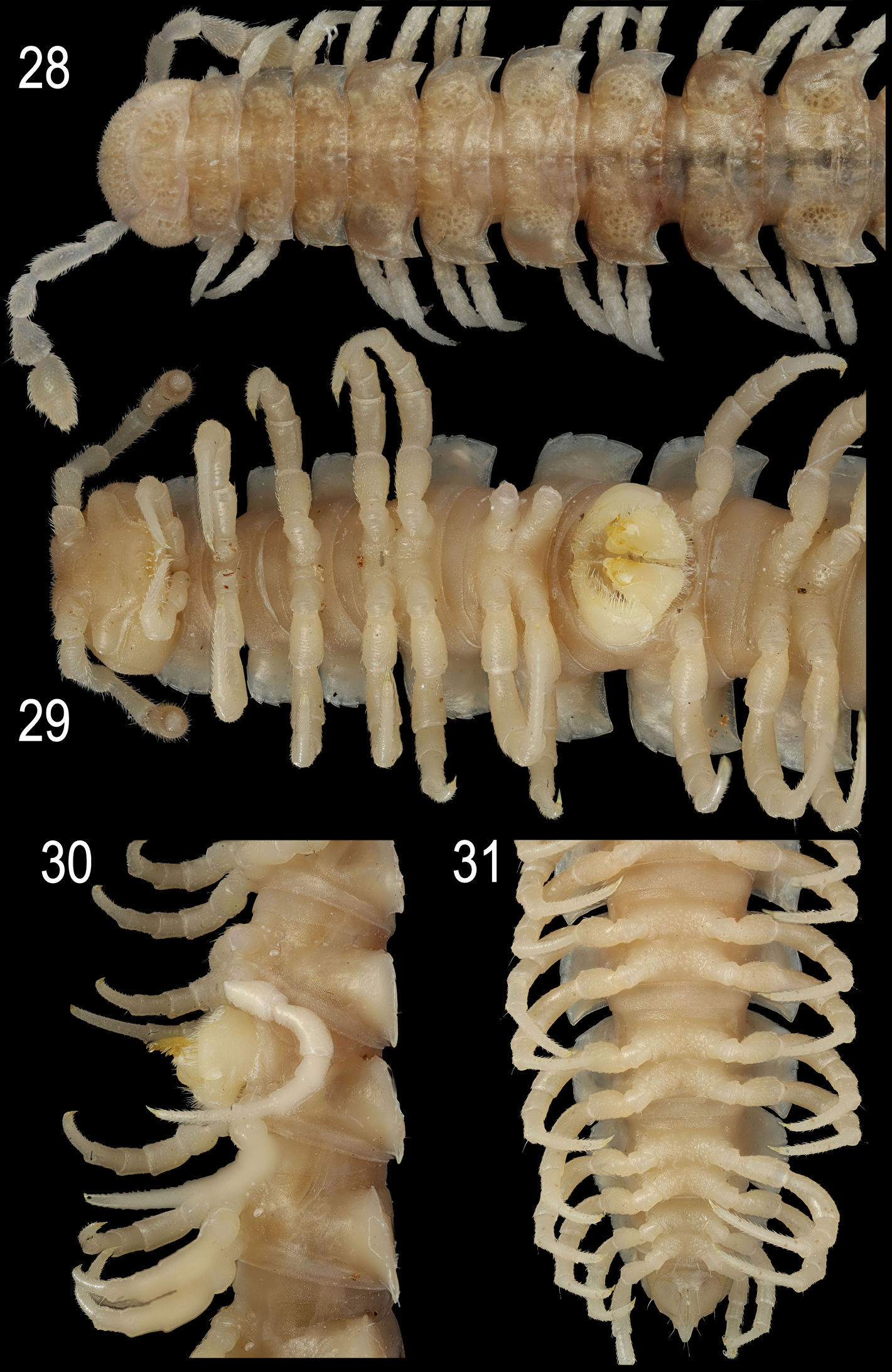
Epanerchodus orientalis Attems, 1901, ♂ from Jialuohu. 28, 29 anterior portion of body, dorsal and ventral views, respectively 30 middle portion of body, lateral view 31 posterior portion of body, ventral view. Photographed not to scale.
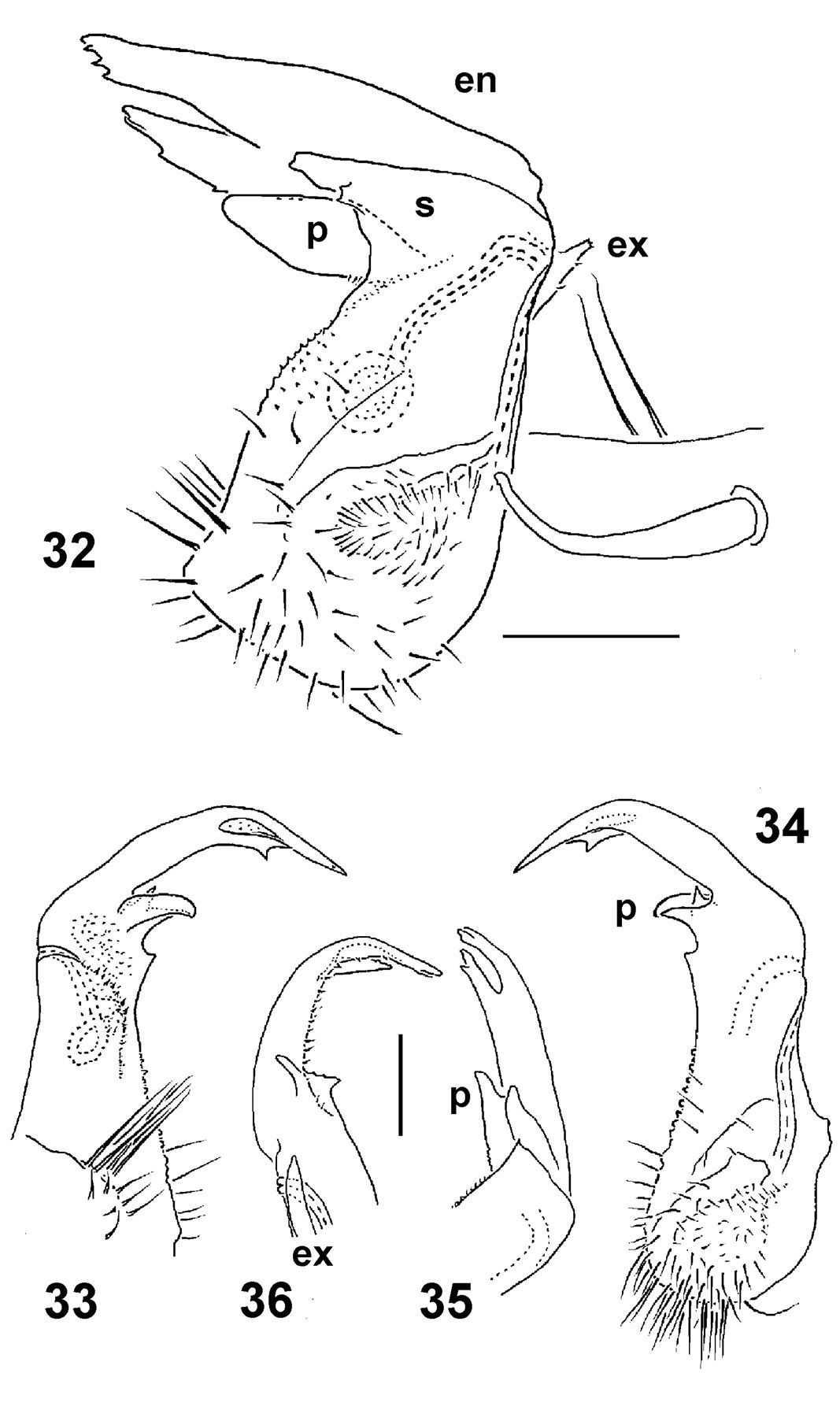
Right gonopods of Epanerchodus orientalis Attems, 1901, ♂♂ from Shengguang (32), Jialuohu (33–35) and Guangfong Highway (36), mesal, lateral, mesal, sublateral and submesal views, respectively. Scale bar: 0.1 mm.
This species seems to be the most widespread and
variable among congeners. It has heretofore been accepted as being split
into two nominal subspecies: Epanerchodus orientalis orientalis and Epanerchodus orientalis takakuwai
Verhoeff, 1913, but, given a similarly profound variation range in
body size and shape, and, especially, in gonopod structure as
observed in Japan alone, the subspecific status of takakuwai has long been questioned (e.g.
It is noteworthy that, formally, Epanerchodus orientalis fails to occur in southernmost Japan, i.e. on Kyushu Island and in the Ryukyus (
Looking at such a profound variation as observed in a number of peripheral and gonopod characters in Epanerchodus orientalis,
one cannot ignore several further nominal congeners described from
Japan, including Kyushu, in which the gonopods look especially similar
to those of Epanerchodus orientalis: Epanerchodus inferus Verhoeff, 1941, Epanerchodus lobatus Verhoeff, 1941, Epanerchodus satoi Takakuwa, 1954, Epanerchodus tenuis Takakuwa, 1954, Epanerchodus aculeatus Miyosi, 1954, Epanerchodus etoi Miyosi, 1955, Epanerchodus chichibensis Haga, in
Based on the great variation observed in the populations of Epanerchodus orientalis in Japan and, especially, Taiwan, the statuses of Epanerchodus polymorphus, Epanerchodus acuticlivus and Epanerchodus aster as further congeners highly similar to Epanerchodus orientalis are likewise to be questioned, as at least some of them might also prove to be the latter’s junior synonyms. Yet no formal synonymies are advanced here as obviously being too premature at this purely descriptive taxonomic stage.
That not all of the congeners similar in gonopod structure to Epanerchodus orientalis are synonyms of the latter is proven at least by Epanerchodus koreanus
Verhoeff, 1937, a species common throughout Korea, in Kyushu, Japan
and in the southern Russian Far East. At least in Russia, it often
occurs syntopically together with Epanerchodus polymorphus. Yet Epanerchodus koreanus,
despite its minor variations in gonopod conformation, is easily
recognizable even superficially through its much wider paraterga
(3.3–3.8 versus 2.6–3.0 mm); in addition, these species never mate (
In other words, several, but definitely not all, of the nominate species of Epanerchodus that show their gonopods particularly similar in structure to those of Epanerchodus orientalis are jeopardized as potential junior synonyms of the latter species.
In Taiwan, Epanerchodus orientalis occurs in various habitats ranging from lowlands to the mid-montane belt (up to 1, 250 m a.s.l.) all over the island (Map 2).
urn:lsid:zoobank.org:act:B0E8A6F1-335A-4961-9876-0328480D6D47
http://species-id.net/wiki/Epanerchodus_pinguis
Figs 37–44Holotype ♂ (head missing) (NMNS-6558-001), Taiwan, Nantou County, Huisun timber land, 15.04.2002, leg. S.H. Wu. Paratype ♂ (NMNS-6558-002), same locality, 24.03.1998, leg. S.H. Wu.
To emphasize the stout gonopod telopodite.
Differs from the other Epanerchodus species, in particular from the apparently especially similar Epanerchodus orientalis, in the mostly square, broader and slightly upturned paraterga, coupled with the gonopod showing an unusually densely setose coxa and a remarkably stout telopodite (see also Key below).
Length ca 16 (paratype) or 20 mm (holotype); width of midbody pro- and metazona 1.5 and 2.9 mm (paratype) or 1.5 and 3.0 mm (holotype), respectively. Coloration in alcohol uniformly light red-brown to red-brown; venter and legs yellowish (Figs 37–39).
All characters as in Epanerchodus orientalis except as follows.
Antennae rather long, slender, only slightly clavate, reaching behind segment 3 dorsally; antennomere 3 longest, clearly longer than highest 5th; antennomeres 5 and 6 each with an evident, compact, distodorsal group of bacilliform sensilla; antennomere 7 with a minute dorsoparabasal cone and a distodorsal group of microscopic sensilla.
In width, collum < head ≤ segment 2 ≤ 3 < 4 < = 5–16, thereafter body gradually tapering towards telson (Fig. 39). Paraterga strongly developed, starting from collum, slightly upturned, set high, mostly level to a very faintly convex dorsum, slightly below dorsum only on collum and segment 2; paraterga on collum small, subtriangular, a small lateral incision in front of a narrowly rounded caudal corner; front shoulders drawn forward only paraterga 2 and 3, straight and subrectangular on paraterga 4, onward straight but directed increasingly caudolaterad; caudal edge nearly straight on paraterga 2–7, thereafter caudal corner increasingly acutangular, nearly always narrowly rounded, only on segment 19 spiniform and nearly pointed, mostly lying nearly level to rear tergal contour until segment 15, onward extending increasingly beyond it (Figs 37–39). Paraterga 2 with 3–4 small lateral incisions, all following poreless segments with three, all pore-bearing ones with four, small but evident incisions, each usually bearing a small seta on top at lateral margin. Metatergal sculpture typical, rather obliterate, with three indistinct transverse rows of setiferous, polygonal bosses (Figs 37–39). Tergal setae very short, mostly retained, a little longer only on collum and in rear row on metatergum 19 (Figs 37, 39). Stricture between pro- and metazona wide and smooth. Limbus very thin, microdenticulate. Epiproct rather short, conical (Fig. 39), preapical papillae prominent. Hypoproct semi-circular; caudal, paramedian, setiferous papillae evident and well-separated.
Sterna without modifications, very densely setose (Figs 40, 41). Legs long and slender, evidently enlarged (Figs 42), ca 1.6–1.7 times as long as midbody height, prefemora not swollen dorsally, acropodite (femur+postfemur+tibia+tarsus) with sphaerotrichomes ventrally (Fig. 42).
Gonopods (Figs 40, 43, 44) with large, subquadrate, medially fused coxae carrying numerous long setae ventromedially. Telopodite mostly hidden inside gonocoel, unusually stout, subfalcate, prefemoral (densely setose) portion about half as long as entire telopodite; endomere (en) short, simple, with a subapical knob laterally and a rounded tip, as well as with two subunciform processes (s and p); hairy pulvillus very evident, exomere totally suppressed.

Epanerchodus pinguis sp. n., ♂ paratype. 37, 40 anterior portion of body, dorsal and ventral views, respectively; 38, middle portion of body, dorsal view 39, 41 posterior portion of body, dorsal and ventral views, respectively. Photographed not to scale.
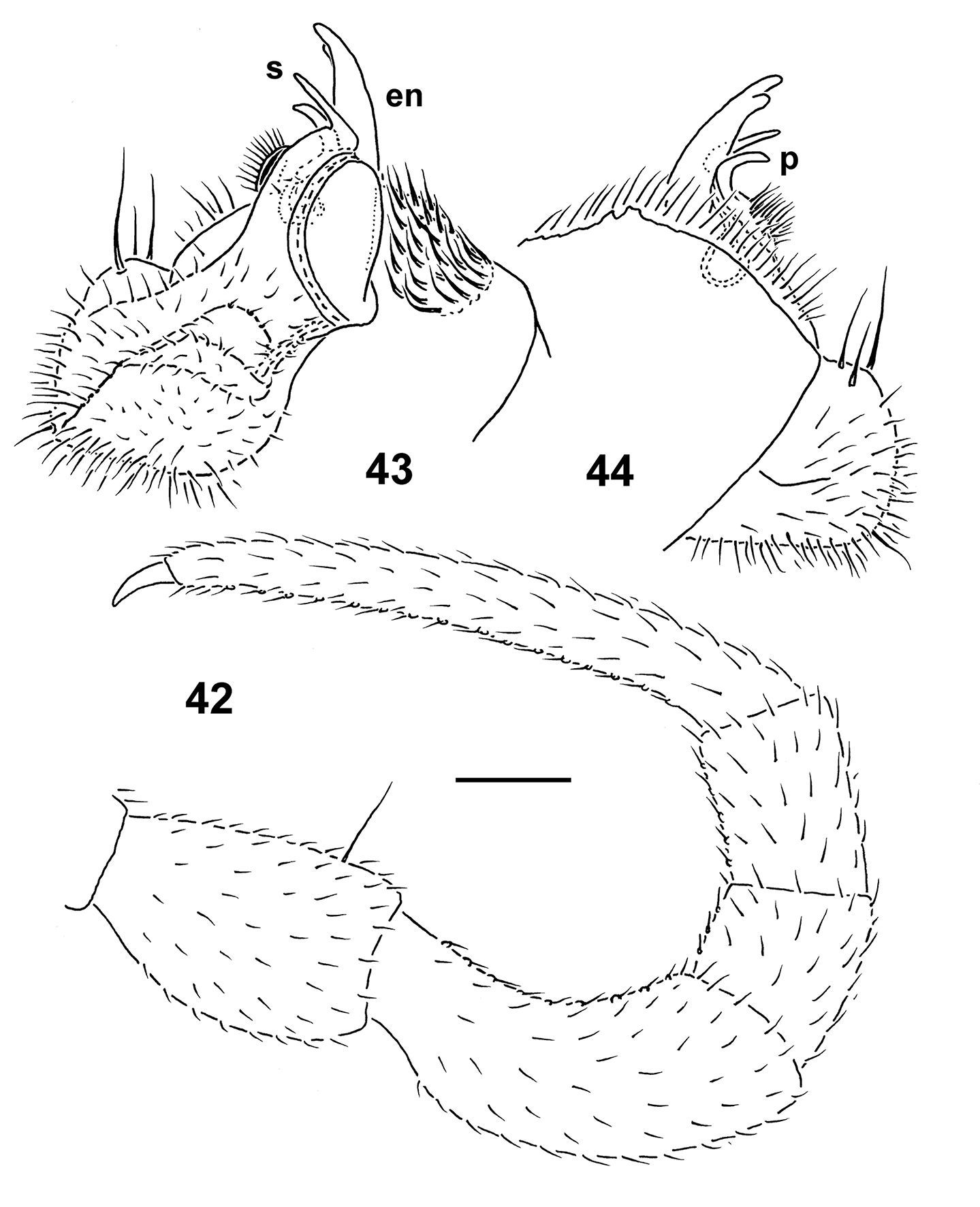
Epanerchodus pinguis sp. n., holotype. 42 leg 9 43, 44 right gonopod, mesal and lateral views, respectively. Scale bar: 0.2 mm.
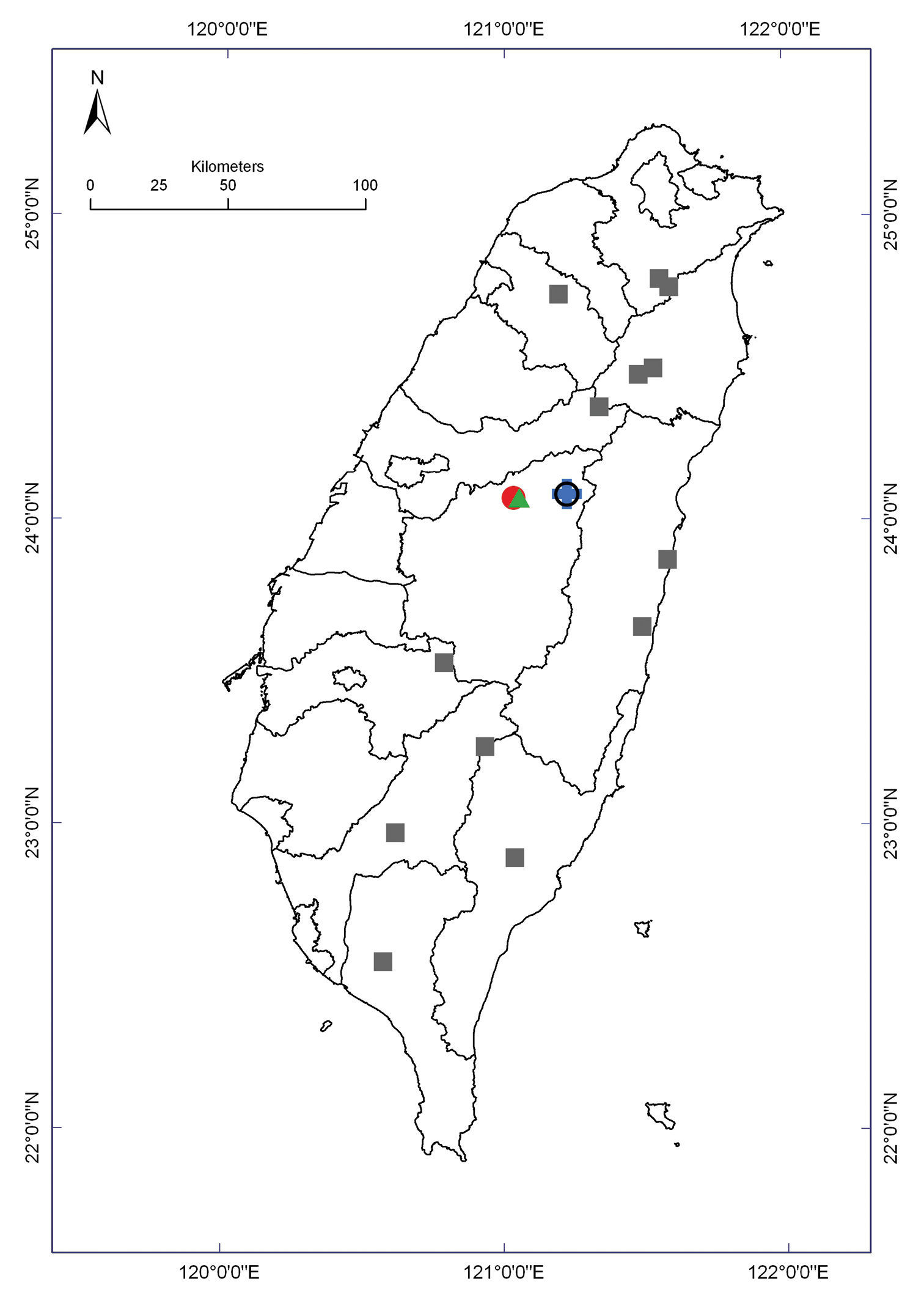
Distribution of Epanerchodus species in Taiwan. Borderlines show borders between the counties. Epanerchodus bispinosus sp. n.: empty black circle; Epanerchodus curtigonopus sp. n.: blue cross; Epanerchodus flagellifer sp. n.: filled green triangles; Epanerchodus orientalis: filled grey squares; Epanerchodus pinguis sp. n.: filled red circle.
This species is apparently very local in distribution (Map 2) and seems to be allopatric with Epanerchodus orientalis.
urn:lsid:zoobank.org:act:5944975E-495B-4578-8145-779D40B9C80A
http://species-id.net/wiki/Epanerchodus_bispinosus
Figs 45–53Holotype ♂ (NMNS-6559-001), Taiwan, Nantou County, Ren-ai Township, Mei-Feng, 15.04.2002, leg. S.H. Wu. Paratypes: 3 ♂, 1 ♂ fragm., 4 ♀, 1 ♀ juv. (NMNS-6559-002), 1 ♂, 1 ♀ (ZMUM), 1 ♂ (without head), 1 ♀ (ZMUC), 1 ♂ fragm., 1 ♀ (MNHN JC 334), 1 ♂ (TFRI), 1 ♂ (NSYSU), same locality, together with holotype.
To emphasize the distal half of the gonopod telopodite being like two long spines.
Due to the presence of a strong and spiniform exomere, this new species joins the few congeners hitherto referred to the erstwhile genus Usbekodesmus (see above the synonymy with Epanerchodus), but differs in the distal half of the gonopod telopodite being represented by only two spiniform branches showing no additional outgrowths (see also Key below).
Length of both sexes ca 8–11 mm; width of pro- and metazona varying between specimens from 0.8–1.3 to 1.1–1.7 mm, respectively. Holotype ca 9 mm long, and 0.8 and 1.1 mm wide on pro- and metazona, respectively. Coloration in alcohol from pallid to uniformly light grey, yellow or very light red-brown, sometimes head faintly marbled light red-brown; venter and legs yellowish to greyish (Figs 45–49).
All characters as in Epanerchodus orientalis except as follows.
Antennae rather long and evidently clavate, reaching behind end (♂) or midway (♀) of segment 3 dorsally; antennomere 3 longest, considerably longer than a relatively stout, yet highest, 5th (Figs 45); antennomeres 5 and 6 each with a small, compact, distodorsal group of bacilliform sensilla; antennomere 7 with a minute dorsoparabasal cone and a distodorsal group of microscopic sensilla.
In width, collum < segment 2 = 3 < 4 < head = 5–15 (Fig. 46), thereafter body gradually tapering towards telson (♂, ♀) (Fig. 48). Paraterga moderately developed, starting from collum, set high but invariably lying slightly below a faintly convex dorsum; paraterga on collum small, subtriangular, with a small lateral incision in front of a rounded caudal corner; front shoulders drawn forward and slightly convex only paraterga 2–4, thereafter straight, increasingly well rounded and directed increasingly caudolaterad; caudal corner on pataterga 2–5 subrectangular and narrowly rounded, starting from 6th increasingly acutangular and beak-shaped, starting from segment 9 first faintly and then extending increasingly beyond rear tergal contour (Figs 45–48). All poreless segments with three, all pore-bearing ones with four, small but evident incisions, each usually bearing a small seta on top at lateral margin. Metatergal sculpture typical, moderately developed, with three indistinct transverse rows of setiferous, polygonal bosses (Figs 46–48). Tergal setae very short, mostly retained, a little longer only on collum and in rear row on metatergum 19. Stricture between pro- and metazona wide and smooth. Limbus very thin, microdenticulate. Epiproct rather short, conical (Fig. 48), only slightly bent ventrad, preapical papillae evident. Hypoproct semi-circular; caudal, paramedian, setiferous papillae evident and well-separated.
Sterna without modifications, very densely setose. Legs rather short, slender, in ♂ evidently enlarged (Figs 45), ca 1.5–1.6 (♂) or 1.1–1.2 times (♀) as long as midbody height, ♂ prefemora not swollen dorsally, together with femora beset with bifid setae ventrally, acropodite (postfemur+tibia+tarsus) with sphaerotrichomes ventrally (Fig. 51).
Gonopods (Figs 50, 52, 53) with large, subquadrate, medially fused coxae carrying only a few long setae ventrally. Telopodite slender and simple, prefemoral portion relatively short, exomere (ex) spiniform, endomere (en) only a little longer than ex, apex bent unciform mesally, devoid of any outgrowths near base; hairy pulvillus evident.

Epanerchodus bispinosus sp. n., ♂ paratype. 45 habitus, lateral view 46 anterior portion of body, dorsal view 47 middle portion of body, dorsal view 48, 49 posterior portion of body, dorsal and ventral views, respectively 50 gonopods, lateral view. Photographed not to scale.

Epanerchodus bispinosus sp. n., ♂ paratype. 51 leg 9 52, 53 left gonopod, submesal and sublateral views, respectively. Scale bar: 0.4 mm (51) and 0.2 mm (52, 53).
This species is remarkable in showing an exomere,
albeit usual and spiniform, almost as long as a particularly simple
endomere. This condition nicely bridges the weak distinction which has
hitherto remained to formally keep Usbekodesmus and Epanerchodus as independent genera (
Epanerchodus bispinosus sp. n. in Taiwan is apparently very local in distribution, having been encountered only at a single locality (Map 2).
urn:lsid:zoobank.org:act:CBDF0100-0AEF-4D12-B88A-3A59119AD5C0
http://species-id.net/wiki/Epanerchodus_curtigonopus
Figs 54–60Holotype ♂ (NMNS-6560-001), Taiwan, Nantou County, Ren-ai Township, Mei-Feng, 15.10.2001, leg. S.H. Wu.
To emphasize the remarkably short gonopod telopodite.
Differs from other Epanerchodus species in the distal half of the gonopod telopodite being particularly stout, with the endomere represented only by a short spine supplied with a similarly short spine at its base (see also Key below). This new species seems to be particularly similar to Epanerchodus pinguis sp. n. in sharing the gonopod which shows a densely setose coxa and a very stout telopodite (apparently, both being synapomorphies), but differs in the entire leg telopodites (prefemur+femur+postfemur+tibia+tarsus) being supplied with sphaerotrichomes ventrally, and the endomere and its basal process rudimentary and spiniform.
Length ca 15 mm; width of pro- and metazona 1.3 and 2.5 mm, respectively. Coloration in alcohol pallid.
Superficially, also very similar to Epanerchodus pinguis sp. n., except as follows.
In width, collum < head < segment 2 = 3 < 4 < 5 < 6–15 (Figs 54), thereafter body gradually tapering towards telson (Fig. 55). Paraterga strongly developed, starting from collum, set high, subhorizontal to very faintly upturned, lying slightly below dorsum only on collum and segment 2; mid-dorsum invariably extremely faintly convex, nearly flat; front shoulders mostly straight, drawn forward only on paraterga 2 and 3, thereafter directed increasingly caudolaterad and increasingly well rounded anterolaterally; caudal corner on paraterga 3–12 subrectangular and narrowly rounded, starting from 13th increasingly acutangular and evidently extending increasingly beyond rear tergal contour, but nearly pointed and beak-shaped only on segment 19 (Figs 54, 55). Paraterga 2–5 and all following pore-bearing segments with four, following poreless ones with three, small but evident incisions, each usually bearing a small seta on top at lateral margin. Metatergal sculpture typical, moderately developed, with three indistinct transverse rows of setiferous, polygonal bosses (Figs 54, 55). Tergal setae very short, mostly retained, a little longer only on collum and in rear row on metatergum 19. Stricture between pro- and metazona wide and smooth. Limbus very thin, microdenticulate. Epiproct rather short, conical (Figs 55, 56), only slightly bent ventrad, preapical papillae evident. Hypoproct semi-circular; caudal, paramedian, setiferous papillae distinct and well-separated (Fig. 56).
Sterna without modifications, densely setose. Legs long and rather slender, only slightly enlarged (Figs 56, 57), ca 1.7–1.8 times as long as midbody height, prefemora not swollen dorsally, entire telopodite (prefemur+femur+postfemur+tibia+tarsus) with sphaerotrichomes ventrally (Fig. 58).
Gonopods (Figs 57, 59, 60) with large, subquadrate, medially fused coxae carrying numerous setae ventrally. Telopodite especially stout, prefemoral portion relatively prominent, taking up about one-third of telopodite length, endomere (en) like a short spine supplied with another, somewhat shorter spine (p) near base; both accessory seminal chamber and hairy pulvillus evident.
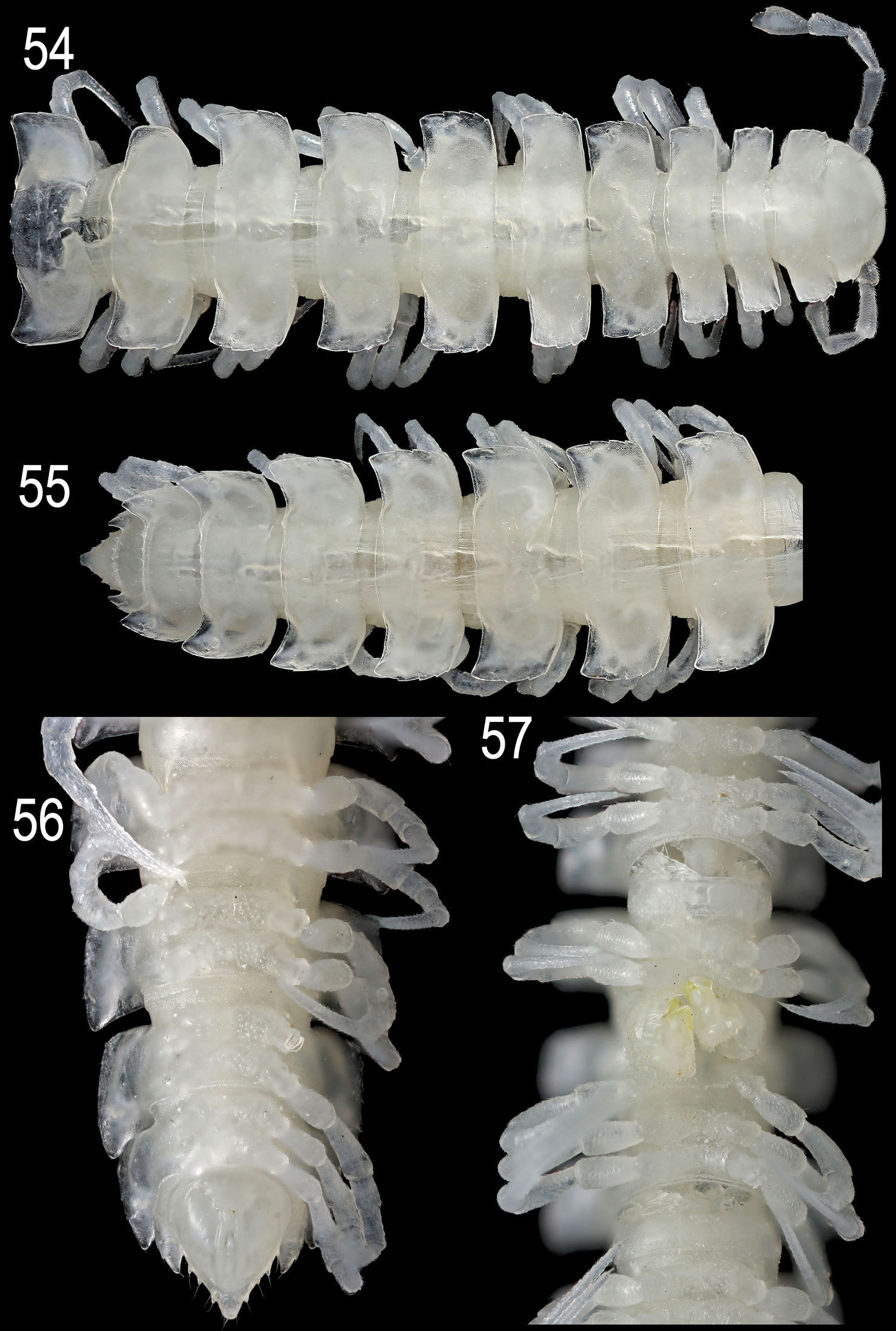
Epanerchodus curtigonopus sp. n., holotype. 54 anterior portion of body, dorsal view 55, 56 posterior portion of body, dorsal and ventral views, respectively 57 middle portion of body (with gonopods), ventral view. Photographed not to scale.
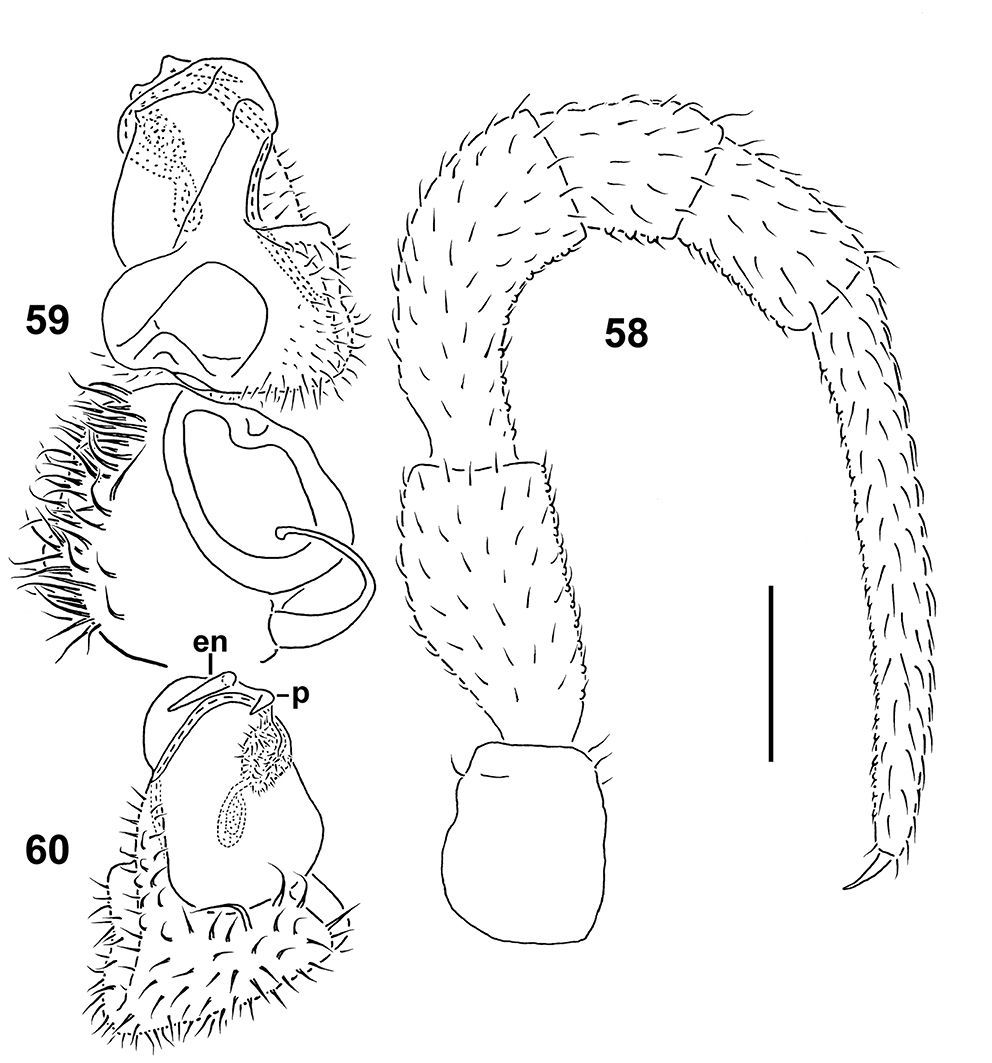
Epanerchodus curtigonopus sp. n., holotype. 58 leg 9 59, 60 left gonopod, dorsomesal and ventrolateral views, respectively. Scale bar: 0.2 mm.
This species is remarkable in showing an extremely short, simple, spiniform endomere (en), coupled with a single, even more rudimentary process (p) at en base.
In Taiwan, Epanerchodus curtigonopus sp. n. occurs very locally, having been found only at a single locality (Map 2).
urn:lsid:zoobank.org:act:35EAB5D4-7E11-496E-8292-4ED5C0676C83
http://species-id.net/wiki/Epanerchodus_flagellifer
Figs 61–68Holotype ♂ (NMNS-6561-001), Taiwan, Nantou County, Huisun timber land, 24.04.1998, leg. S.H. Wu. Paratypes: 1 ♂ (NMNS-6561-002), same locality, together with holotype; 1 ♂ (NMNS-6561-003), same locality, 04.1998; 1 ♂ (ZMUM), same locality, 25.04.1999; 1 ♂ (TFRI), same locality, 24.04.1999, all leg. S.H. Wu.
To emphasize the remarkably long, flagelliform distal part of the gonopod telopodite.
Differs from other Epanerchodus species in the distal half of the gonopod telopodite being particularly long, flagelliform, coupled with the presence of two rounded teeth at the base of the endomere (see also Key below). From the other congeners known from Taiwan, this new species differs also in the absence of sphaerotrichomes on ♂ legs.
Length of both sexes ca 8–11 mm; width of pro- and metazona varying between specimens from 0.8–1.0 to 1.4–1.5 mm, respectively. Holotype ca 9 mm long, and 0.9 and 1.4 mm wide on pro- and metazona, respectively. Coloration in alcohol pallid to light grey- to red-brown (Figs 61–65).
All characters as in Epanerchodus orientalis except as follows.
Antennae rather long and evidently clavate (Figs 61, 64), reaching behind segment 3 dorsally.
In width, collum < segment 2 < head = 3 = 4 < 5–15(16) (Fig. 62), thereafter body gradually tapering towards telson (Fig. 63). Paraterga well-developed, starting from collum, set high, subhorizontal to very faintly upturned, mostly nearly level to a very faintly convex dorsum, only on collum and segment 2 lying clearly below dorsum; paraterga on collum small, subtriangular, a small lateral incision in front of a rounded caudal corner; front shoulders drawn forward and slightly convex only paraterga 2 and 3, straight on paraterga 4, thereafter increasingly well rounded and directed increasingly caudolaterad; caudal edge straight on segments 2–4, thereafter increasingly bisinuate and acutangular caudolaterally; starting from segment 10, caudal corners extending increasingly beyond rear tergal contour, spiniform on segments 17–19 (Figs 61–65). All poreless segments with three, all pore-bearing ones with four, small but evident incisions, each usually bearing a small seta on top at lateral margin. Metatergal sculpture typical, moderately developed, with three indistinct transverse rows of setiferous, polygonal bosses (Figs 62, 63). Tergal setae very short, mostly retained, a little longer only on collum and in rear row on metatergum 19. Stricture between pro- and metazona wide and smooth. Limbus very thin, microdenticulate. Epiproct rather short, conical (Fig. 65), only slightly bent ventrad, preapical papillae evident. Hypoproct semi-circular (Fig. 65); caudal, paramedian, setiferous papillae evident and well-separated.
Sterna without modifications, very densely setose. Legs rather long, slender, only slightly incrassate (Figs 61, 64–66), ca 1.7–1.8 times (♀) as long as midbody height, prefemora not swollen dorsally, sphaerotrichomes missing (Fig. 66).
Gonopods (Figs 67, 68) with large, subquadrate, medially fused coxae carrying only a few long setae ventrally. Telopodite simple and especially slender, prefemoral portion relatively short, endomere (en) flagelliform, with two small dentiform outgrowths at base; hairy pulvillus evident.

Epanerchodus flagellifer sp. n., ♂ paratype. 61 habitus, lateral view 62, 64 anterior portion of body, dorsal and ventral views, respectively 63, 65 posterior portion of body, dorsal and ventral views, respectively. Photographed not to scale.
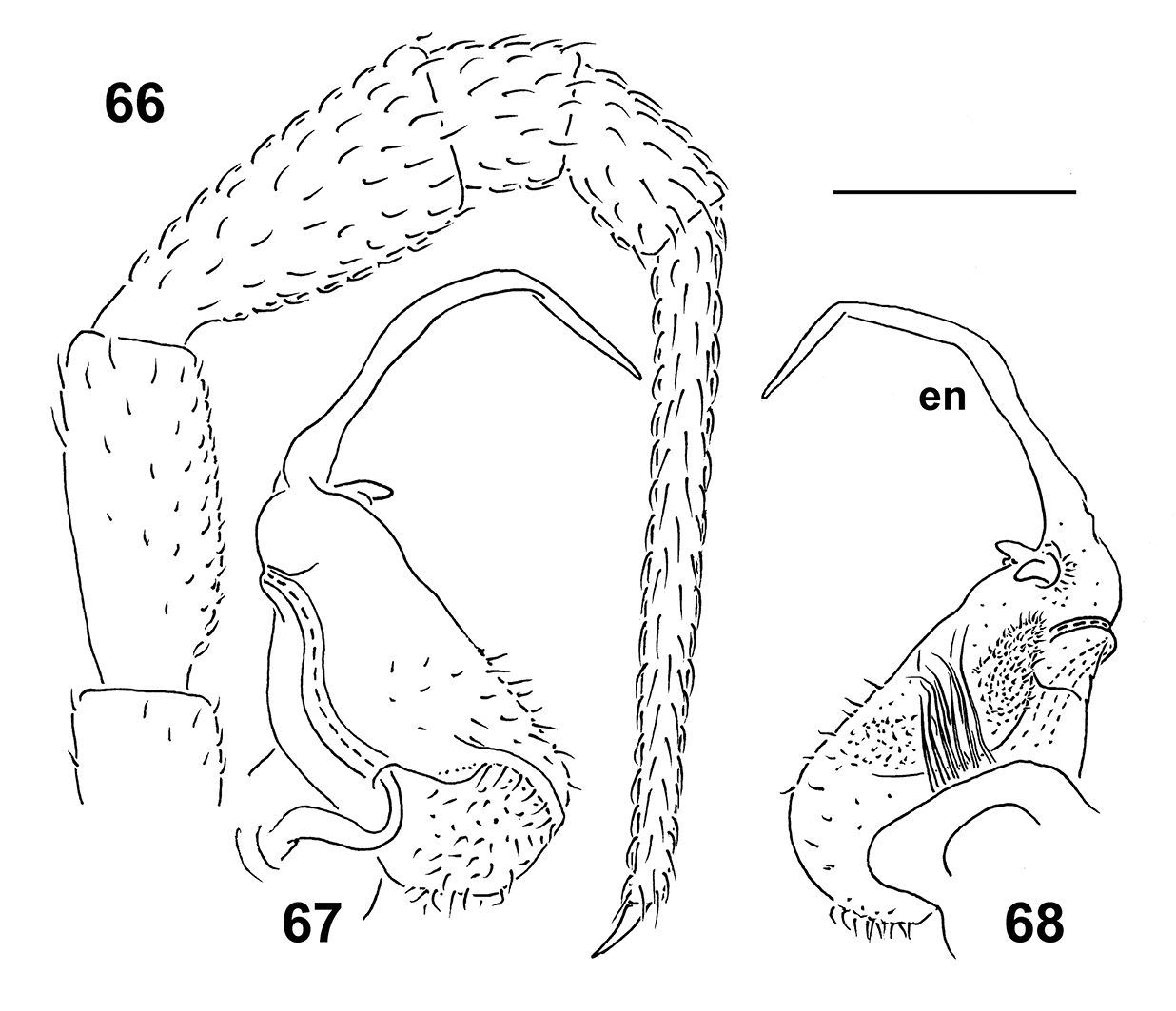
Epanerchodus flagellifer sp. n., ♂ paratype. 66 leg 9 67, 68 left gonopod, mesal and lateral views, respectively. Scale bar: 0.2 mm.
This small species is remarkable in showing a particularly long, flagelliform endomere, coupled with only two short outgrowths at its base.
In Taiwan, Epanerchodus flagellifer sp. n. occurs very locally, having been encountered only at a single locality (Map 2).
| 1 | Gonopod coxae unusually densely setose, while telopodite particularly stout: prefemoral, setose part at least half as long as entire telopodite (Figs 43, 44, 59, 60) | 2 |
| – | Gonopod coxae usual, with only a few strong setae, while telopodite elongate: prefemoral, setose part less than half as long as entire telopodite (Figs 32, 52, 53, 67, 68) | 3 |
| 2 | ♂ legs devoid of sphaerotrichomes (Fig. 66). Gonopod endomere (en) like a short spine (Figs 67, 68) | Epanerchodus curtigonopus sp. n. |
| – | all ♂ telopoditomeres with sphaerotrichomes (Fig. 42). Gonopod endomere (en) like a strong tooth (Figs 40, 43, 44) | Epanerchodus pinguis sp. n. |
| 3 | Body usually larger, 11–19 mm long. Gonopod telopodite highly variable, complex: exomere (ex), if present, rudimentary, while endomere (en) branching (Figs 29, 30, 32–36) | Epanerchodus orientalis |
| – | Body usually smaller, 8–11 mm long. Gonopod telopodite simple: exomere (ex), if present, strong and spiniform, while endomere (en) like a strong spine or a long flagellum | 4 |
| 4 | Gonopod telopodite biramous, exomere (ex) strong and spiniform, while endomere (en) like a strong spine ending in a strong uncus (Figs 52, 53) | Epanerchodus bispinosus sp. n. |
| – | Gonopod telopodite uniramous, exomere absent, endomere (en) flagelliform (Figs 67, 68) | Epanerchodus flagellifer sp. n. |
At present we can state that Taiwan supports a reasonably
rich and quite peculiar polydesmid fauna represented by seven species
in two genera. Of these, six species seem to be endemic, being largely
restricted to the mountains in the central and northern parts of the
island. Considering the distributions demonstrated by the Taiwanese Polydesmidae (Maps 1 and 2),
allopatry is prevailing while sympatry, probably even syntopy, has
only been observed in two places. Thus, the normally much larger Nipponesmus shirinensis, the slightly smaller Epanerchodus pinguis sp. n. and the smallest Epanerchodus flagellifer sp. n. co-occur at Huisun. Similarly, Nipponesmus shirinensis, Nipponesmus minor sp. n., Epanerchodus orientalis and Epanerchodus bispinosus
sp. n., which also form a comparable succession of size decrease, are
sympatric if not syntopic at Mei-Feng. Such examples seem to be best
explained in terms of local niche segregation long documented elsewhere
for a number of insular groups of Diplopoda (e.g.
The only polydesmid that is remarkably widespread and polymorphous in Taiwan, Epanerchodus orientalis, appears to be confined to lowland to foothill habitats (Map 2). For this reason alone, this species, which is also known to be extremely widely distributed, variable and lowland-dwelling across at least most of Japan, seems to be the only allochthonous element in the fauna of Polydesmidae of Taiwan, likely a later colonizer from Japan.
We are most grateful to the National Science Council, Taiwan, Republic of China and to the Russian Academy of Sciences, Moscow, Russian Federation for the support rendered to the Taiwanese and Russian teams, headed by H. W. Chang and S.I. Golovatch, respectively, to actively collaborate in our joint ecofaunistic studies on the Myriapoda of Taiwan.
Special thanks go to K. S. Lee (NMNS) and J. T. Chao (TFRI) for kindly loaning material under their care. K. Makarov (Moscow, Russia) very skillfully took the pictures, N. Likhitrakarn (Chulalongkorn University, Bangkok, Thailand) kindly helped in preparing the plates with colour photographs, while H.Y. Hsu (NSYSU) executed the maps. We are greatly obliged to C. C. Chen, S. Y. Wu, M. H. Hsu, J. D. Lee, H. D. Zhu, M. J. Hung, Y. M. Chen, W. C. Yeh, C. Y. Huang and S. H. Wu for their help in collecting material. We also thank the administration of the National Parks of Taiwan, and all departments of the county and city governments of Taiwan for permitting the collection of millipedes.