





(C) 2010 Golovatch Sergei I.. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
The hitherto monotypic genus Caucasodesmus is new to the Ukrainian list due to the discovery of Caucasodesmus tauricus sp. n. in a cave in the Crimea. The new species is easily distinguished from Caucasodesmus inexpectatus Golovatch, 1985, the type, and only other, known species of this genus, in the abundantly setose collum and following metaterga, and more elaborate gonopods. The status of Caucasodesmus, which shows in the superfamily Trichopolydesmoidea where it definitely belongs such evident generic-level apomorphies as the absence of bacilliform sensilla on antennomeres 5 and 7, of a cannula on the gonocoxite, and of a seminal groove on a biramous gononod telopodite (apparently, both latter characters are functionally correlated to each other), is refined by formally reassigning it to the family Trichopolydesmidae.
millipede, Trichopolydesmidae, taxonomy, new species, cave, Crimea
The millipede fauna of the Crimea has recently been reviewed (
The more so important is the present discovery of still another new millipede in a cave in the Crimea. Unlike all other faunal elements, this new species might prove to represent the first truly relict, palaeoendemic in the diplopod list of that peninsula.
Material serving as the basis for the present contribution was captured with pitfall traps, later transferred into 75% alcohol and is currently deposited in the collection of the Zoological Museum, State University of Moscow, Russia. Specimens were studied and illustrated using standard stereomicroscopic, photographic and drawing equipment.
SystematicsTrichopolydesmidae Verhoeff, 1910
Caucasodesmus inexpectatus Golovatch, 1985, by original designation.
urn:lsid:zoobank.org:act:66FEBC49-89EC-432E-A0E2-561179B19B24
http://species-id.net/wiki/Caucasodesmus_tauricus
Figs 1–8Holotype ♂, Ukraine, Crimea, Mt Villya-Burun, Cave Villyaburunskaya, pitfall traps, 12.05.2008–12.10.2010, leg. A. Koval. – Paratype: 1 ♀, same locality, 19.07.2004–17.07.2006, leg. A. Koval.
Easily distinguished from Caucasodesmus inexpectatus Golovatch, 1985, the type, and only other, known species of this genus, by the abundantly setose metaterga and more elaborate gonopods.
Length of both sexes ca 8 mm, width of midbody pro- and metazona 0.8 and 1.5 mm, respectively. Coloration in alcohol from uniformly pallid to light yellowish.
Body with 20 segments. Tegument mainly dull, at most slightly shining, texture very delicately alveolate. Head densely pilose throughout; epicranial suture distinct but thin; isthmus between antennae ca 1.5 times broader than length of antennomere 1, still broader than diameter of antennal socket. Antennae rather short, evidently clavate due to a considerably enlarged antennomere 6, slightly overreaching segment 2 dorsally; antennomeres 2, 3 and 6 longest, subequal in length (Figs 1, 2); only antennomere 6 with a large, compact, roundish, distodorsal group of bacilliform sensilla.
In width, collum < segment 2 = 3 < head = 4 < 5=16 (♂) or head = collum = segment 2 = 4 < 5=16 (♀), thereafter body gradually tapering towards telson. Paraterga moderately developed, starting from collum, subhorizontal to slightly declivous, set high but always lying slightly below a faintly convex dorsum, devoid of shoulders frontally (Figs 1–3). Caudal corner of postcollum paraterga invariably spiniform, pointed, starting from segment 4 extending increasingly further than rear tergal margin. Lateral edge of paraterga with neither marginal groove nor thickening, with 5–6 clear setigerous indentations. Pore formula normal, ozopores evident, round, located laterally in front of caudalmost incision. Collum and following metaterga beset with numerous medium-sized setae set on minute knobs, polygonal bosses missing (Figs 1–3). Stricture between pro- and metazona wide, shallow and smooth. Limbus very thin, microdenticulate. Pleurosternal carinae absent (Fig. 2). Epiproct short, conical, directed caudoventrally; preapical papillae small (Fig. 4). Hypoproct subtrapeziform, setiferous papillae at caudal corners evident, rather well separated.
Sterna without modifications, poorly setose. Epigynal ridge very low. Legs rather short (Figs 2, 5), ca 1.2–1.3 (♂) or 0.9–1.0 (♀) times as long as midbody height; ♀ legs slightly slenderer; ♂ legs with clearly enlarged prefemora and femora; tarsi especially long and slender, claw long, ca 1/4 length of tarsus; sphaerotrichomes missing (Fig. 5).
Gonopod aperture large, transversely oblong-oval, taking up nearly all of ventral part of metazonite 7. Gonopods (Figs 6–8) with large, globose, medially fused coxae carrying rather numerous setae laterally, but no trace of a cannula. Telopodite subfalcate, distally of a rather short prefemoral (= setose) portion split into two branches: exomere (ex) largest and longest, more simple, whereas endomere (en) shorter, more complex in shape; an evident, tooth-like, mesal process (d) at base between both ex and en; no trace of a seminal groove.

Caucasodesmus tauricus sp. n., holotype. 1, 2 anterior half of body, dorsal and lateral views, respectively 3, 4 posterior portion of body, dorsal and ventral views, respectively. Photographed not to scale.
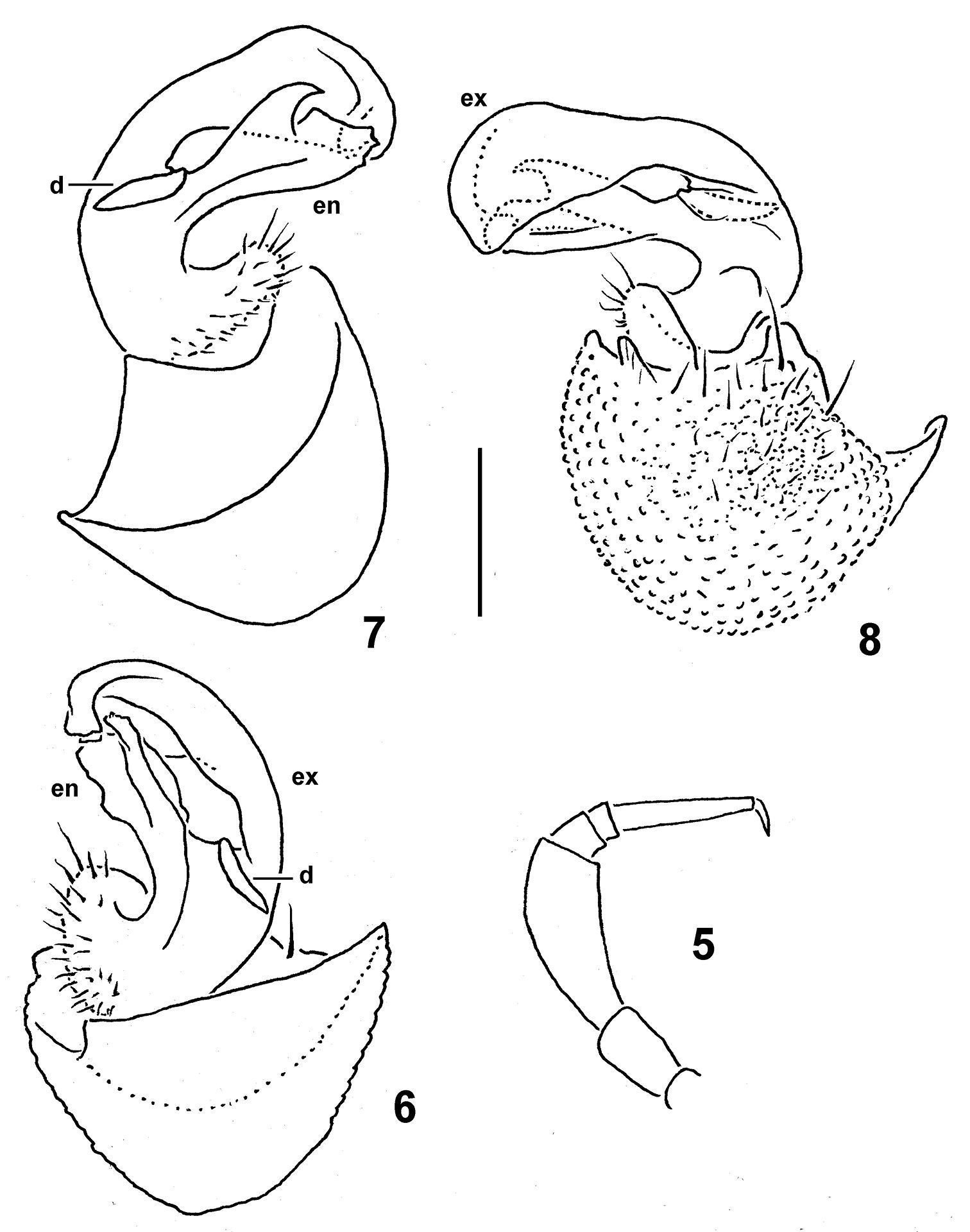
Caucasodesmus tauricus sp. n., holotype. 5 leg 9 (setae not shown) 6 rightgonopod, anteromesal view 7, 8, left gonopod, mesal, and lateral views, respectively. Scale bar: 0.4 (5) and 0.2 mm (6–8).
This species is an unquestioned relict troglobite and, based on its zoogeographical traits, might well represent the first palaeoendemic in the diplopod fauna of the Crimea.
Only one species of Caucasodesmus has hitherto been known: Caucasodesmus inexpectatus from Cave Nyvjin Lagat (= Nyvdzhinlagat, = Tagardonskaya) in North Ossetia, central Caucasus Major (
Originally, Caucasodesmus was treated as a genus of the small Holarctic family Macrosternodesmidae (
The superfamily Trichopolydesmoidea
can be defined by its gonopod prefemoral (= setose) part orientated
mostly transversely to the body’s main axis, extending mesally across
the entire width of the coxae (
Macrosternodesmidae (
Nearctodesmidae (
Trichopolydesmidae (
Mastigonodesmidae (
Fuhrmannodesmidae (
Based on the above diagnoses and distributions, it appears to be quite difficult to unequivocally reallocate Caucasodesmus. The correlated absence of both a cannula and a seminal groove is probably a sufficiently strong apomorphy to erect still another family of Trichopolydesmoidea for the accommodation of solely this genus, but I refrain here from doing so pending more information becomes available. New taxa are still being regularly described, new synonymies established, and old types revised. Instead I reassign Caucasodesmus to Trichopolydesmidae as a family not only representing the oldest taxon in the superfamily, but also one which shows the same basic traits of gonopod structure and a coherent distribution pattern.
I am most grateful to Alexandr Koval, St Petersburg, Russia, who has kindly allowed me to study his invaluable material, as well as to deposit it entirely in the Moscow Museum. I am also deeply obliged to Kirill Makarov, Moscow, Russia who skillfully took the pictures, and to Igor Muratov, Pietermaritzburg, Republic of South Africa for his kind technical assistance.