





(C) 2010 Anthony I. Cognato. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
A cladistic analysis based on 20 morphological characters was conducted for 11 species representing two valid and two synonymized Phloeotribini genera. One hundred-eighty most-parsimonious trees were recovered and the Dryotomicus Wood species were monophyletic in a mostly unresolved strict-consensus tree. The unusual antennal morphology, with the length of the first two funicular segments equal to the last three segments and a scape which is twice the length of the funicle, distinguish Dryotomicus from the other Phloeotribini genera. Hence this genus is resurrected because of monophyly and diagnostic characters. Dryotomicus oenophilis sp. n. and Dryotomicus woodrex sp. n. are described from Guyana and Peru, respectively. In the male specimen of Dryotomicus oenophilis, the frons has one median and two large lateral carinae and in the male specimen of Dryotomicus woodrex, the frons has three smaller median tubercles arranged transversely. Phloeotribus puberulus Chapuis and Phloeotribus tuberculatus (Eggers) were monophyletic with the new Dryotomicus species and thus are transferred to this genus. Keys to the Phloeotribini genera and Dryotomicus species are given.
Scolytidae, bark beetle, taxonomy, tropical biodiversity
The most diverse and unknown scolytine fauna lies in the
tropics. Although a recent monograph of the South American scolytines
has been published, approximately another 2500 species remain
undiscovered in the Neotropics (
In this study, we assembled specimens of Neotropical, Nearctic, and Australian Phloeotribini and conducted a cladistic analysis, which justified the resurrection of Dryotomicus and the description of two new species.
Materials and methodsSpecimens of one Chramesus (outgroup), one Aricerus and
11 Phloeotribus species, which included all species described as
Dryotomicus and Eulytocerus (
Characters and their states used in the phylogenetic analysis.
| Character 1 | Antennal club. 0: segments fused, 1: segments articulated. |
| Character 2 | Antennal club, shape of first segment. 0: not applicable, 1: asymmetrical with a constant width, 2: Asymmetrical, expanded at base (j- shaped), 3: symmetrical chevron-shaped. |
| Character 3 | Second funicular segment. 0: not longer than total length of segments 3-5, 1: longer than total length of segments 3-5. |
| Character 4 | Length of scape. 0: not extending beyond the anterior margin of the pronotum, 1: extending beyond the anterior margin of the pronotum. |
| Character 5 | Area between the antennal insertions and mandibles. 0: less than or equal to length of the mandibles, 1: greater than the length of the mandibles. |
| Character 6 | Male frons with median tubercule(s). 0: absent, 1: one 2: > one. |
| Character 7 | Male frons with lateral carina. 0: absent, 1: present. |
| Character 8 | Male head sulcate from the apex of the eyes to the vertex. 0: absent, 1: present. |
| Character 9 | Declivitous anterior edge of the pronotum. 0: absent, 1: present. |
| Character 10 | Dorsum of the pronotum. 0: with dense, minute, oppressed setae, 1: with scattered longer erect setae. |
| Character 11 | Anterior and lateral margins of the pronotum. 0: without asperities, 1: with asperities. |
| Character 12 | Basal margin of the elytra. 0: crenulate, 1: carinate. |
| Character 13 | Elytral striae. 0: deeply impressed, 1: shallowly impressed. |
| Character 14 | First and second interstriae on elytral declivity. 0: raised above striae, 1: flush with striae. |
| Character 15 | Third, fifth and seventh elytral interstriae on declivity. 0: with tubercules, 1: smooth. |
| Character 16 | Scales on elytral declivity. 0: absent, 1: present. |
| Character 17 | Tubercles on protibia (male). 0:3, 1:5, 2:6, 3:7, 4:8, 5:9. |
| Character 18 | Protibiae. 0: without socketed teeth, 1: with socketed teeth. |
| Character 19 | Metatibae. 0: widest at distal end, 1: widest near the middle. |
| Character 20 | Metatibae. 0: with less than five denticles, 1: with more than five denticles. |
Using this data matrix (Table 2), most parsimonious trees (mpts) were reconstructed by a branch and bound search in PAUP* 4.0 b10 PPC using default settings (
Character states used for the reconstruction of the Phloeotribini phylogeny (Fig. 1). Characters and states are in Table 1.
| Characters | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | |
| Aricerus sp. | 1 | 3 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Phloeotribus championi | 1 | 2 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 5 | 1 | 1 | 1 |
| Phloeotribus sp. Costa Rica | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | ? | 1 | 0 | 1 |
| Phloeotribus sp. Ecuador | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 2 | 1 | 1 | 1 |
| Phloeotribus frontalis | 1 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 1 |
| Phloeotribus liminaris | 1 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 2 | 1 | 0 | 1 |
| Phloeotribus ovatus | 1 | 1 | 0 | 0 | 0 | ? | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 3 | 1 | 0 | ? |
| Phloeotribus sp. Panama | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 2 | 1 | 0 | 1 |
| Dryotomicus oenophillis sp. n. | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 2 | 1 | 1 | 1 |
| Dryotomicus woodrex sp. n. | 1 | 2 | 1 | 1 | 1 | 2 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 1 | 2 | 1 | 1 | 1 |
| Phloeotribus puberulus | 1 | 2 | 1 | 1 | 1 | ? | ? | ? | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 2 | 1 | 1 | 1 |
| Phloeotribus tuberculatus | ? | ? | ? | ? | ? | ? | ? | ? | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | ? | 1 | 1 | 1 |
| Chramesus sp. | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 4 | 1 | 0 | 1 |
One hundred-eighty mpts where reconstructed for the 13
taxa. The strict consensus tree of the mpts was mostly unresolved except
for the monophyly of Dryotomicus species (Fig. 1).
This clade has a high bootstrap value (100), is supported by a
relatively high Bremer value (4) and has several diagnostic characters (Fig. 1).
The antennal funicle, in which the length of the first two funicular
segments equals the last three segments, and the scape, which is twice
the length of the funicle, are the most striking features. Antennal
morphological variation is taxonomically important because these
features diagnose Aricerus as well as species of Phloeotribus (
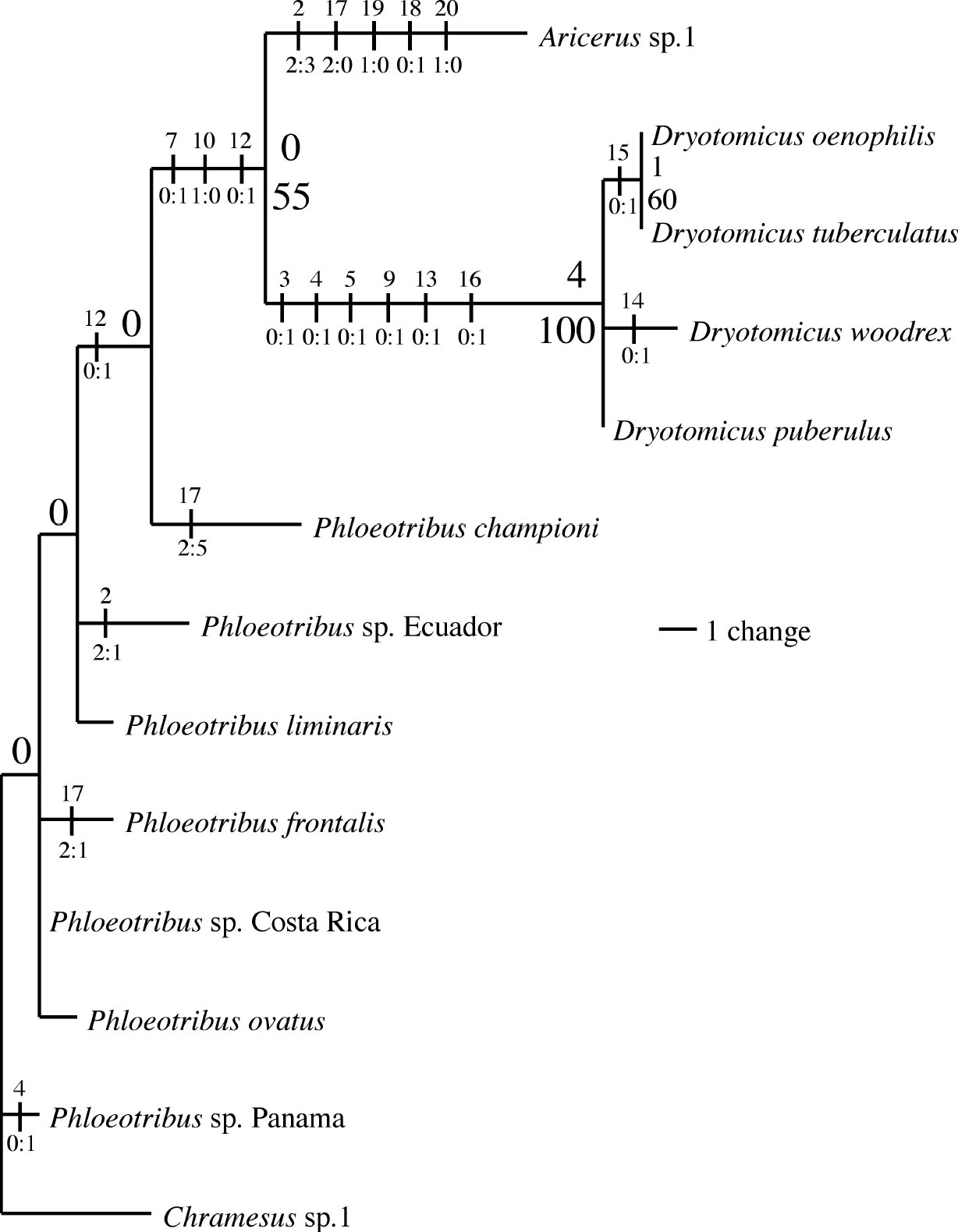
Phylogeny of Phloeotribini species, 1 of 180 most parsimonious trees (tree length=35, RC=0.6429). Numbers above branches are Bremer support and numbers below are bootstrap values (< 50 not shown). Nodes with zero Bremer support were unresolved in a strict consensus of the 180 mpts. Vertical lines represent unambiguous character state changes inferred using accelerated transformations. Numbers above vertical lines are the characters and numbers below are the character state changes.
The relationship of the four Dryotomicus species is mostly unresolved (Fig. 1). Low support values indicate a potential sister- relationship between Dryotomicus oenophilis and Dryotomicus tuberculatus. However this is a likely spurious result caused by missing character data for Dryotomicus tuberculatus; the only known specimen representing this species is missing its head. However, the distinct morphology of the male frons and elytral declivity distinguish the new species from Dryotomicus tuberculatus (Fig. 1).
SystematicsThe asymmetrical first segment of antennal club, socketed teeth on the protibae, and rounded lateral margins of the pronotum distinguish this genus from Aricerus. The usual median tubercle(s) on the male frons, the longer second antennal funicular segment, declivitous anterior edge of the pronotum, the shallowly impressed elytral striae, and elytral declivity with scales and long setae distinguishes Dryotomicus from Phloeotribus.
urn:lsid:zoobank.org:act:A2174B38-3EBD-4463-AB45-718C449F858D
Figs 2– 5Dryotomicus oenophilis is distinguished from the other Dryotomicus species by a large medial tubercle and lateral carina with acute proximal tips on the male frons, interstriae 2 without long uniserial setae, and by raised interstriae of the elytral declivity having tubercles on interstriae 3, 5, and 7 (Fig. 3B).
Holotype, male, total length 4.5 mm (3.8–4.5 mm, n=7), 2× longer than wide, color reddish-black (Fig. 2).
Head. Frons shagreen with setae as long as or longer than the large median tubercle, longest setae close to epistoma and frontal margins; a large median tubercle between antennal insertions and dorsal margin of eye; lateral carinae from epistoma to dorsal end of eye thicker at antennal insertion and ending acutely (Fig. 3A). Vertex, shagreen with setae approximately as long as or longer than large median tubercle; slightly concave with distinct slightly carinate lateral margins; obtuse median carina from median frontal tubercle to epistoma. Antennae, scape expanded distally and curved proximally beyond the anterior edge of pronotum, funicle five segmented, segments 1 and 2 about equal length and each as long as the combination of segments 3, 4, 5, club pseudo-lamellate, asymmetric, basal segment 1 expanded at base (j- shaped). Eyes oval, ventrally acute (Fig. 3A).

Dryotomicus oenophilis sp. n. male. Habitus, A Dorsal B Lateral.

Dryotomicus oenophilis sp. n. male, A Frons B Elytral declivity.
Pronotal width 2.2 mm (1.7–2.2 mm, n=7), 0.64× longer than wide; quadrate flat summit not apparent, densely punctured with appressed minute pubescence and scattered longer setae approximately as long as the funicle concentrated anteriorly and laterally.
Elytra 1.2× longer than wide, 2× longer than pronotum, striae on disk impressed, punctures only evident near declivity; striae 4–9 marked by shallow, uniserial punctures; interstriae on disk 2–3× width of striae, confused scales from base to apex on interstriae 1 and 2, interstriae 1 and 3–9 with long uniserial setae approximately as long as the funicle; interstriae 3–9 minutely punctured. (Figs 2, 3B). Elytral declivity with densely placed scales and scattered long setae; striae impressed; interstriae 3, 5, 7 each with 3 tubercles (Fig. 3B).
Male genitalia. Aedeagal body (median lobe) conical, apex acute, lateral margins heavily sclerotized medially on apical half, apophyses (struts) as long as body, attached ventrally; internal sac central area lightly sclerotized, lateral margins heavily sclerotized appearing as ventral apophyses (struts) directed apically , seminal trough at proximal end comprised of two lobes that curve medially (Fig. 4). Tegmen circular, weakly sclerotised on dorsal side. Spiculum gastrale nearly as long as adeagus, crescent-shaped with small knob near the apicalend.
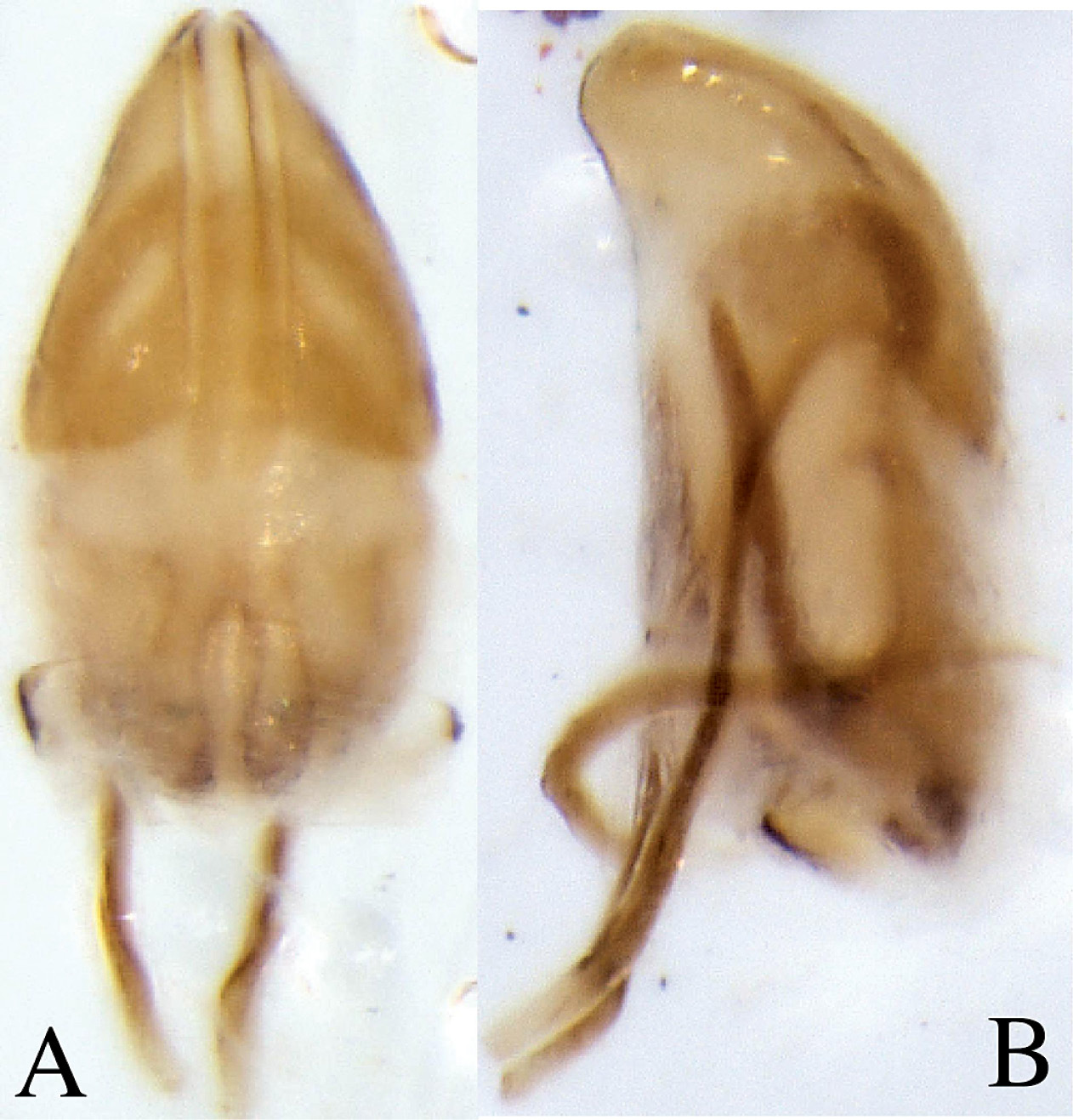
Dryotomicus oenophilis sp. n. male genitalia, A Dorsal B Lateral. Spiculum gastrale not shown.
Female similar to male in most features, except frons flat to slightly convex, densely punctured, without median tubercles and carinae (Fig. 5A). Strial punctures on elytra more distinct, interstrial tubercles smaller (Fig. 5B).

Dryotomicus oenophilis sp. n. female, A Lateral habitus B Frons.
Holotype and 6 paratypes (3 males and 3 females) bear two collection data labels, First: “Guyana: Iwokrama Forest, GPS N 04, 40.486’, W 58.41.028’, 4–9 March 2007, Cognato, Hulcr, Smith, Dole, McCall Colls”; Second: “Collected with ethanol trap”. The holotype is deposited in the Biodiversity Center at the University of Guyana and 4 paratypes are deposited in the A. J. Cook Arthropod Research Collection, Michigan State University, East Lansing; 2 paratypes are in the U.S. National Museum of Natural History, Washington D.C.
In Guyana, we collected all specimens from 20 plastic cups filled with 100 ml of 90% ethanol and nailed to trees 1.5 meters above ground.
oeno (G) = wine, philis (G) = lover. The name “wine-lover” signifies the collection of all specimens from ethanol traps.
Dryotomicus woodrex is distinguished from the other Dryotomicus species by three medial tubercles arranged transversely on a tumescence on the male frons; the interstriae flush with striae on the elytral declivity.
Holotype, male, total length 4.6 mm, 2× longer than wide, antennae reddish-black, head, legs, thorax, and elytra tannish (perhaps teneral). Pronotum tannish with dark diamond pattern (Fig. 6).

Dryotomicus woodrex sp. n. male. Habitus, A Dorsal B Lateral.
Head. Frons shagreen with setae approximately as long as or longer than funicle; three medial tubercles arranged transversely on a tumescence between antennal insertion and dorsal margin of eye; lateral carinae from epistoma to dorsal end of eye thicker at antennal insertions (Fig. 7A). Vertex shagreen with setae approximately as long or longer than funicle; slightly concave. Antennae, scape expanded distally and curved proximally beyond the anterior edge of pronotum, funicle 5-segmented, segments 1 and 2 about equal length and each as long as segments 3–5 combined, club pseudo-lamellate, asymmetric, segment 1 expanded at base (j- shaped). Eyes oval, ventrally acute (Fig. 3A).

Dryotomicus woodrex sp. n. male, A Frons B Elytral declivity.
Pronotal width 2.3 mm, 0.65× longer than wide; quadrate disk, summit not distinct, surface densely punctured with appressed pubescence and scattered longer setae approximately as long as funicle concentrated anteriorly and laterally.
Elytra 1.2× longer than wide, 2× longer than pronotum, striae on disk not impressed, punctures distinct; interstriae 3–4× width of striae, long uniserial setae approximately as long as funicle arising from granules (Fig. 6). Elytral declivity densely scaled with scattered long setae concentrated along the lateral margin (Fig. 7B).
Male genitalia. Aedeagal body (median lobe) conical, apex acute, lateral margins heavily sclerotized medially on apical half, apophyses (struts), as long as body, attached ventrally; internal sac central area lightly sclerotized, lateral margins heavily sclerotized appearing as ventral struts directed apically, seminal trough proximal end comprised of two parallel lobes (Fig. 8). Tegmen circular, weakly sclerotised on dorsal side. Spiculum gastrale destroyed by dissection.
Female is unknown.
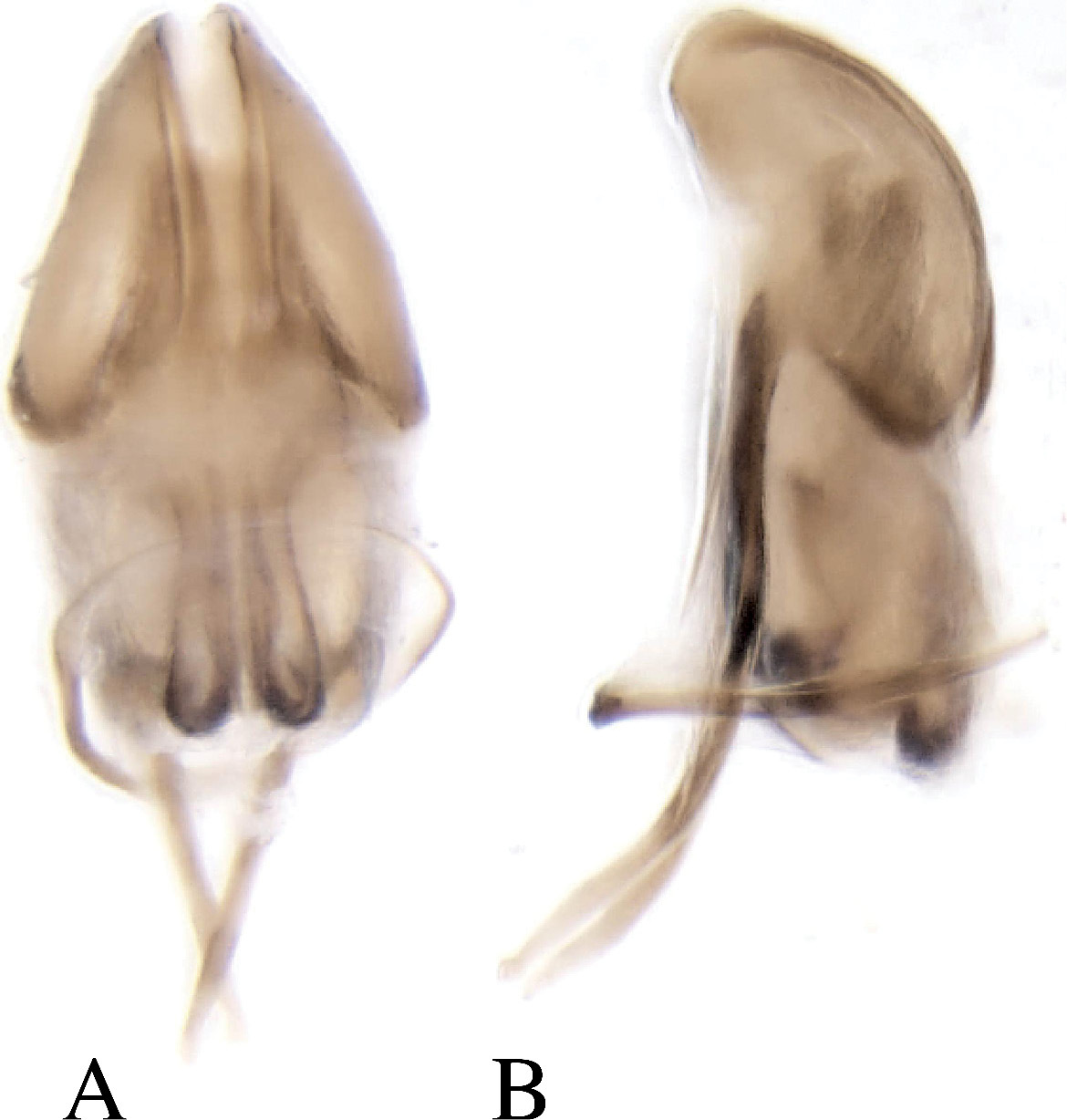
Dryotomicus woodrex sp. n. male genitalia, A Dorsal B Lateral.
Holotype bears the collection data label: “PERU: Dept. Loreto, 1.5km N Teniente Lopez, 4°35.66’S; 76°06.92’W, 22 July 1993, 210–240 m, Richard Leschen #164, ex: flight interception trap”. The holotype is deposited in the Snow Museum, University of Kansas [SMEC].
The name “woodrex” honors Dr. Stephen L. Wood’s kingly contribution to the knowledge of scolytine and platypodine taxonomy. It is used as a noun in apposition.
This species differs from other Dryotomicus spp. by the absence of tubercules from the third, fifth and seventh interstriae and the raised first and second interstriae of elytral declivity.
See
This species differs from other Dryotomicus spp. by the presence of tubercules on the third, fifth and seventh interstriae of the elytral declivity and rugose interstriae of elytral disk and with more than 3 tubercles (Fig. 10)
See
| 1. | First segment of antennal club symmetrical chevron-shaped (Fig. 9); protibia without socketed teeth; lateral margin of pronotum marked by asperites; Australia to New Guinea | Aricerus |
| – | First segment of antennal club asymmetrical; protiba with socketed teeth; lateral margin of pronotum rounded, without asperites | 2 |
| 2. | Second antennal funicular segment longer than total length of funicular segments 3–5; anterior edge of the pronotum declivitous; elytral declivity with scales and long setae | Dryotomicus |
| – | Second antennal funicular shorter than total length of funicular segments 3–5; anterior edge of the pronotum flat; elytral declivity with long setae only | Phloeotribus |
| 1. | Elytral declivity with tubercules on the third, fifth and seventh interstriae | 2 |
| – | Elytral declivity without tubercules on the third, fifth and seventh interstriae | 3 |
| 2. | Interstriae of elytral disk smooth and with 3 or fewer tubercles | Dryotomicus oenophilis sp.n. |
| – | Interstriae of elytral disk rugose and with more than 3 tubercles (Fig. 10) | Dryotomicus tuberculatus (Eggers) |
| 3. | First and second interstriae on elytral declivity flush with striae | Dryotomicus woodrex sp. n. |
| – | First and second interstriae on elytral declivity raised above striae (Fig. 11) | Dryotomicus puberulus (Chapuis) |

Aricerus sp. antenna.

Dryotomicus tuberculatus elytra.
Dryotomicus puberulus elytral declivity.
We thank Robin McCall and the faculty of University of Guyana who helped with the acquisition of collecting and export permits for Guyanese specimens. We also thank Max Barclay (BMNH), Natalia Vanderburg (USNM), Zack Falin (SEMC) and Peter Grootaert (IRSNB) for the loans of specimens. This research was supported by NSF-PEET grant (DEB-0328920).