





(C) 2010 Traudl Krapp-Schickel. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
This is the second part of a revision of the most plesiomorphic genera in the amphipod family Stenothoidae sensu lato (see
Stenothoidae sensu lato, systematics, phylogenetic analysis, gen. Proboloides, Malvinometopa gen. n., Victometopa gen. n., Victometopa rorida sp. n.
As many species as possible of this group were studied and redescribed, in order to replace the (initially numerous) question marks in the start-up matrix. Species were borrowed from different Museums.
Acronyms for Museums
AMS Australian Museum Sydney
BMNH British Museum (Natural History), London
MNVCr Museo civico di Storia Naturale Verona
NMV Museum of Victoria, Melbourne, Australia
ZMUC Zoological Museum, University of Copenhagen or Købnhavn
Abbreviations in taxonomical descriptions as well as figures
A1, 2 antenna 1, 2
acc. accessory
art article
Cx coxal plate
Ep epimeral plate
flag flagellum
Gn1, 2 gnathopod 1, 2
IP inner plate
LL lower lip
Md mandible
Mx1, 2 maxilla 1, 2
Mxp maxilliped
OP outer plate
P3-7 peraeopod 3–7
ped peduncle
T telson
U1–3 uropod 1–3
UL upper lip
Us urosome
Character matrixThe chosen characters were as follows:
Head
(1.) A1 length (0) > A2; (1) ≤ A2
(2.) A2 peduncle article 1 ratio length: breadth (0) ≤ 3; (1) > 3
(3.) ratio A1: body length (0) ≥ 0.66% body; (1) < 0.66% body
(4.) A1 flagellum acc. (0) many articles; (1) 2–1 articles; ; (2) lacking
(5.) A1 peduncle art 2 (0) < article 1; (1) ≥ article 1
(6.) A1 peduncle art 1 (0) < ceph.; (1) = ceph.; (2) > ceph.
(7.) A1 peduncle art 3 (0) ≤0.3 art 1 ; (1) 0.3–0.5 art 1; (2) ≥ 0.5 art 1
(8.) A1 flagellum arts (0) <10; (1) 11–20; (2) 21–30; (3) >30
(9.) A2 peduncle art 5 (0) < flag, ; (1) = flag.; (2) > flag.
(10.) A2 peduncle art 4 (0) >art 5; (1) =art 5; (2) <art 5
(11.) A2 nr. flagellum arts (0) <9; (1) 10–15; (2) >15
(12.) Lateral cephalic lobes (0) rounded; (1) subacute, blunt
(13.) Eyes (0) medium; (1) small to absent; (2) large
Mouthparts
(14.) Mandible palp art 3 (0) ≥ half art 2; (1) < half 2 or lacking
(15.) Mandible palp art 2 (0) ≥3 setae; (1) 3–1 setae; (2) lacking
(16.) Mandible palp art 3 (0) ≥2 setae; (1) one distally; (2) lacking
(17.) Maxilliped outer plate : merus (0) ≥ 0.5; (1) 0.5–0.2; (2) <0.2
Coxal plates
(18.) Ratio length Cx2 : Cx1 (0) <2; (1) 2–2.5; (2) 2.5–3; (3) >3
(19.) Cx2 ratio length : breadth (0); ≥1.5; (1) < 1.5
(20.) Cx4 ratio length : breadth (0) l>b; (1) l=b; (2) l<b
(21.) Cx4 distally excavated (0) no; (1) yes
Gnathopods
(22.) Gnathopod 1 dactylus (0) ordinary; (1) spoon-shaped
(23.) Gn1 palm (0) < half propodus length; (1) ≥ half propodus
(24.) Gn1 palm angle about (0) no one = 180°; (1) blunt = 180–150° ; (2) acute = 120°; (3) transverse = 90°
(25.) Gn1, 2 propodus shape (0) similar; (1) different
(26.) Gn 1 propodus shape (0) rounded; (1) linear-rectangular; (2) triangular
(27.) Gn1 carpus (0) short, length< 2 breadth; (1) l=2b; (2) l<2b
(28.) Gn1 merus (0) short; (1) elongate; (2) freeprojecting
(29.) Gn1 ratio carpus : propodus (0) < prop.; (1) = prop.; (2) > prop.
(30.) Ratio length propodus Gn1 : Gn2 (0) ≥0.75; (1) 0.75–0.33; (2) <0.33
(31.) Gn2 ratio propodus : coxa male (0) ≥1; (1) 1–0.66; (2) <0.66
(32.) Gn2 ratio propodus:basis male (0) ≥1; (1) 1–0.66; (2) <0.66
(33.) Gn2 ratio propodus:basis female (0) ) ≥1; (1) 1–0.66; (2) <0.66
(34.) Gn2 ratio propodus : coxa female (0) ≥1; (1) 1–0.66; (2) <0.66
(35.) Gn2 palm male (0) smooth; (1) toothed-serrated; (2) incision(s)
(36.) Gn2 palm female (0) smooth; (1) toothed-serrated; (2) incision(s)
(37.) Gn2 carpus shape (0) short, l<b; (1) ; elongate, l≥b
(38.) Gn2 merus shape (0) not lobate; (1) lobate
Peraeopods
(39.) P4 ratio posterior margin of merus : propodus (0) ≤1.33; (1) >1.33
(40.) P5 basis (0) ovoid widened; (1) rectangularly widened; (2) slim like basis P4
(41.) P5 ratio anterior margin of merus : propodus (0) ≤1.25; (1) >1.25
(42.) P5 merus tip reaching (0) no carpus; (1) 0.25–0.75 carpus; (2) full carpus
(43.) P5 basis posterodistally (0) no lobe; (1) small lobe; (2) medium lobe; (3) lobe wide and deep, reaching merus
(44.) P5 basis width ratio maximum : minimum (0); 1–1.1 (1) 1.1–1.4; (2) 1.4–1.6; (3) 1.6–1.8; (4) >1.8
(45.) P6 basis (0) ovoid widened; (1) narrow like P5; (2) rectangularly widened
(46.) P6 basis hindmargin (0); harmonically rounded (1) straight
(47.) P6 basis posterodistal corner (0) rounded lobe; (1) no lobe
(48.) P6 merus length anterior: posterior margin (0) =1; (1) >1
(49.) P7 basis shape (0) rounded; (1) narrow like P5; (2) rectangularly widened
(50.) P7 merus reaching (0) no carpus; (1) <0.5 carpus; (2) >0.5 carpus; (3) all carpus
(51.) P7 ratio dactylus: propodus (0) <0.5; (1) ≥0.5
(52.) P7 basis posterior margin (0); convex; (1) concave; (2) straight Epimeral plates, Uropods + Telson
(53.) Ep3 posterodistally (0) rectangular corner; (1) acute, 60–70°
(54.) U1 rami (0) equal; (1) very different
(55.) U1 ratio peduncle : longer ramus (0) ≤1; (1) 1–1.33; (2) >1.33
(56.) U2 ratio of rami (0) > 66%; (1) ≤ 66%;
(57.) U2 ratio peduncle : longer ramus (0) < 1; (1) > 1
(58.) U2 peduncle spination (0) no to weak; (1) strong
(59.) U3 ramus ratio art 1 : art 2 (0) <1; (1) =1; (2) 1.1–1.9; (3) >2
(60.) U3 ramus spination (0) no; (1) 1–3 spines; (2) many
(61.) Telson (0); length : breadth ≤2 (1) ; length : breadth >2 (2) 3-dimensional
It is remarkable that all mouthparts are always unusually long and narrow and it could be imagined that they all together function as a sucking device (it is said that the name steno-thoids stems from the narrow mouthparts, stenos meaning narrow in Greek). Mxp has reduced plates and Md has more or less reduced molars and palps, while pars incisivus and lacinia mobilis are very well developed with acute and robust “teeth”; also both maxillae have robust setae, which could help to divide the food parts already cut by the mandible.
In many stenothoids P3, 4 are longer but weaker than P5–7, and all are kept parallel to the coxae and never twisted. Interesting are the quite often acutely lengthened meri (in P3, 4 anteriorly, in P5–7 posteriorly) which warrant an additional capacity against fallling off the substrate.
TaxonomyMetopa gregaria Sars, 1882: 93, t. 4, fig. 6
Proboloides mainly occurs in the Atlantic, but nominal Proboloides species have been reported also from the Pacific, Indian and Antarctic oceans. Its species are often found living in deep waters and show a clear sexual dimorphism, usually their gnathopods are quite different in size and shape, often with a strongly incised Gn2 palm, with palmar corner well defined in females, but not defined in males, and robust peraeopods.
A1 peduncle art 1 usually short, length < 3× width, subequal to cephalon; A1 usually shorter than 2/3 body length, A1 accessory flagellum lacking. Md palp with a very short or lacking art3, poorly setose; Mx1 palp 2 arts; Mx2 inner plate ordinary; Mxp inner plates well separated, outer plates usually reduced (less than 0.2 of merus length). Ratio Cx2:Cx1 > 3. Cx2 length equal or more than 1.5 x the width. Gn1, 2 different in size and shape; Gn1 small, almost simple, rarely subchelate; carpus length equal to propodus; length of propodus Gn1 about half or less than half length of propodus Gn2; Gn2 palm has serrations or teeth, usually no incisions; Gn2 propodus is in males often, in females always smaller than Cx2; carpus shorter than wide, merus elongate. P5 basis linear, without posterodistal lobe; merus anterior margin shorter than 1.25 length of propodus anterior margin. P6, 7 basis expanded and lobate, merus tip reaching half to full length of carpus. Ep3 with acute posterodistal corner. U1 peduncle is longer than longer ramus. T length is shorter to equal the double width, triangular, laminar.
At the beginning of this study 16 species were known:
11 from the Atlantic, Pacific and Indian Ocean: Proboloides anophthalmus Ledoyer, 1986, Proboloides calcaratus (Sars, 1882), Proboloides clypeatus (Stimpson, 1853), Proboloides grandimanus (Bonnier, 1896), Proboloides gregarius (Sars, 1895), Proboloides holmesi Bousfield, 1973, Proboloides pacificus (Holmes, 1908), Proboloides schokalskii Gurjanova, 1946, Proboloides schuleikini Gurjanova, 1946, Proboloides tundus Barnard, 1962, Proboloides zubovi Gurjanva, 1951.
5 members from Antarctic-Subantarctic region: Proboloides porcellanus KH Barnard, 1932, Proboloides rotundus (Stebbing, 1917), Proboloides stephenseni
The differences between the current diagnoses of Metopoides, Proboloides, Scaphodactylus and Torometopa are still quite small and not satisfactory:
Metopoides. Mouthparts ordinary. Long antennae with 2-articulate flag. acc.; unspecialized gnathopods; short coxal plates; basis P6, 7 with weakly lengthened merus.
Proboloides. Md palp may have a shortened third article, the inner plates of Mxp may be fused. Antennae robust with 0–1 articulate acc. flag.; gnathopods with sexual dimorphism, Gn1 much smaller than Gn2; coxal plates enlarged; basis P6, 7 with strongly lengthened and widened merus.
Scaphodactylus. Mouthparts ordinary. Antennae with 2- articulate acc. flag.; gnathopods without or with sexual dimorphism (there were two groups within the genus); Gn1 dactylus spoon-shaped excavated; coxal plates small; basis P5 rectolinear with posterodistal lobe lengthened and widened; P6, 7 merus very weakly lengthened and widened.
Torometopa. Mouthparts ordinary. Antennae with 0–2- articulate acc. flag.; gnathopods without or with sexual dimorphism; coxal plates small or large; basis P5 rectolinear with posterodistal lobe lengthened and widened to varying degrees; P6, 7 merus weakly to strongly lengthened and widened. In short, characters of Metopoides and Proboloides together, but P5 basis with posterodistal lobe, which might have evolved independently. Thus this genus was the least convincing one.
To fill the gaps with question marks in the first matrix (see also
This species has a small posterodistal lobe on P5 basis (like Torometopa and different from all other Proboloides), but P6, 7 bases are not rounded (like in all Torometopa), but narrowing distad. Gn 1,2 show a striking similarity with those of Mesometopa sinuata Shoemaker, 1964 (female) from the American west coast, but P5–7 in that species are totally different, and the other Mesometopa species are Pacific arctic-boreal (southernmost locality is S. California).
This species thus fits in no existing genus, and a new one is erected:
urn:lsid:zoobank.org:act:17A26AB9-5D20-46FE-B5BA-4075DF82B8BD
Proboloides porcellanus KH Barnard, 1932
Md palp with 3 arts, Mxp IP separated; Mx1, 2 unknown; A1, 2 peduncle strong, flagellum reduced to 4–6 arts. Cx4 not much wider than Cx2+3. P5 basis rectolinear, with small posterodistal lobe. P6, 7 basis narrowing distad, both with posterodistal small lobe reaching along half of ischium.
The type species was collected from the pharynx of a large ascidian in the Falkland Islands, in Spanish Islas Malvinas.
This genus has a posterodistal lobe on P5 basis like the members of Torometopa; itis different from all other known stenothoid genera by the rectangularly widened, distally narrowing basis of P6, 7. In Metopella and Mesoproboloides P5 is rectolinear without a posterodistal lobe, P6, 7 are differently widened; in Hardametopa P5–7 all have a slender basis. The genera Metopelloides, Stenothoides, Vonimetopa and Zaikometopa differ in having a 1-articulate Md palp, while the palp is absent in Parametopella.
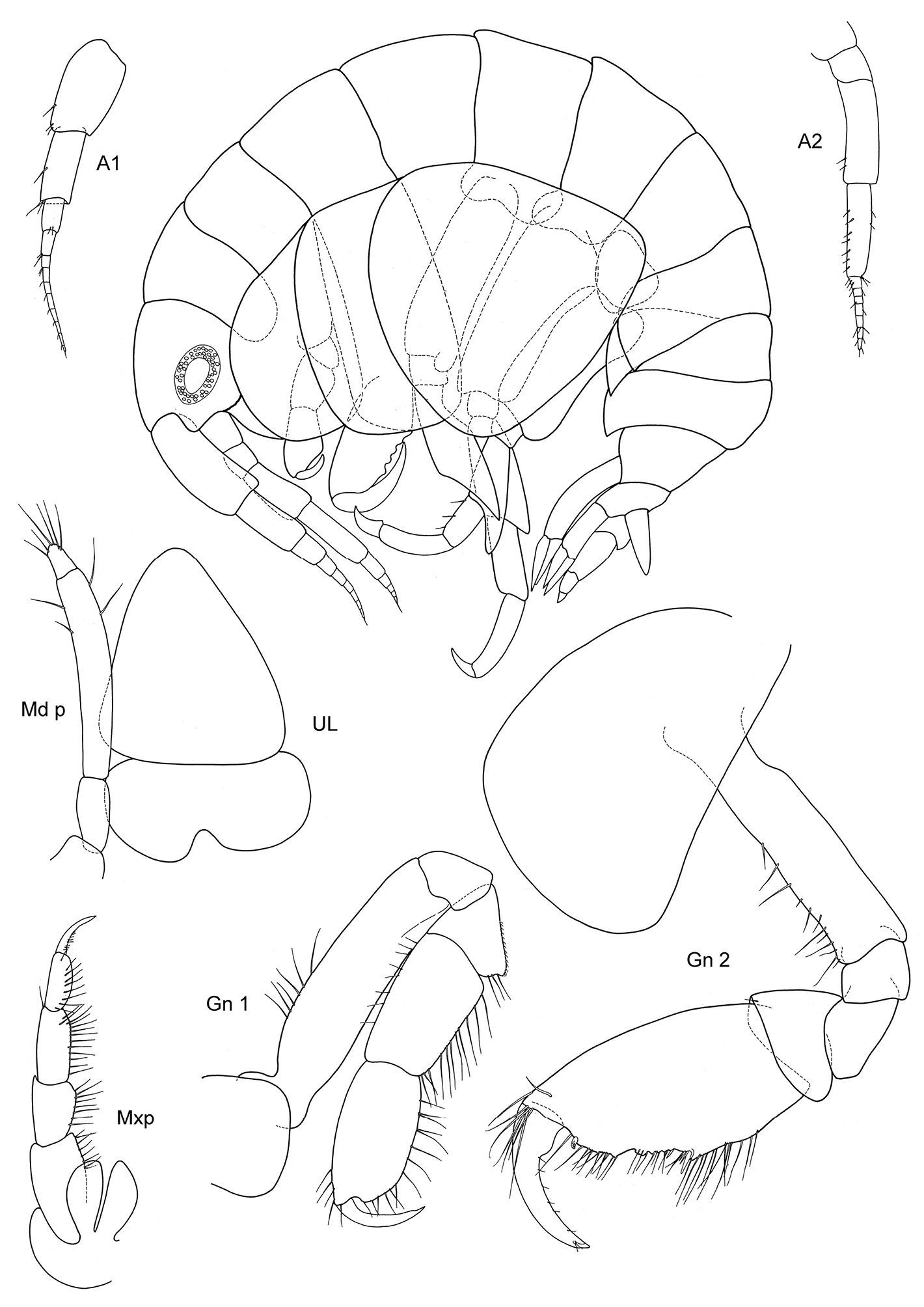
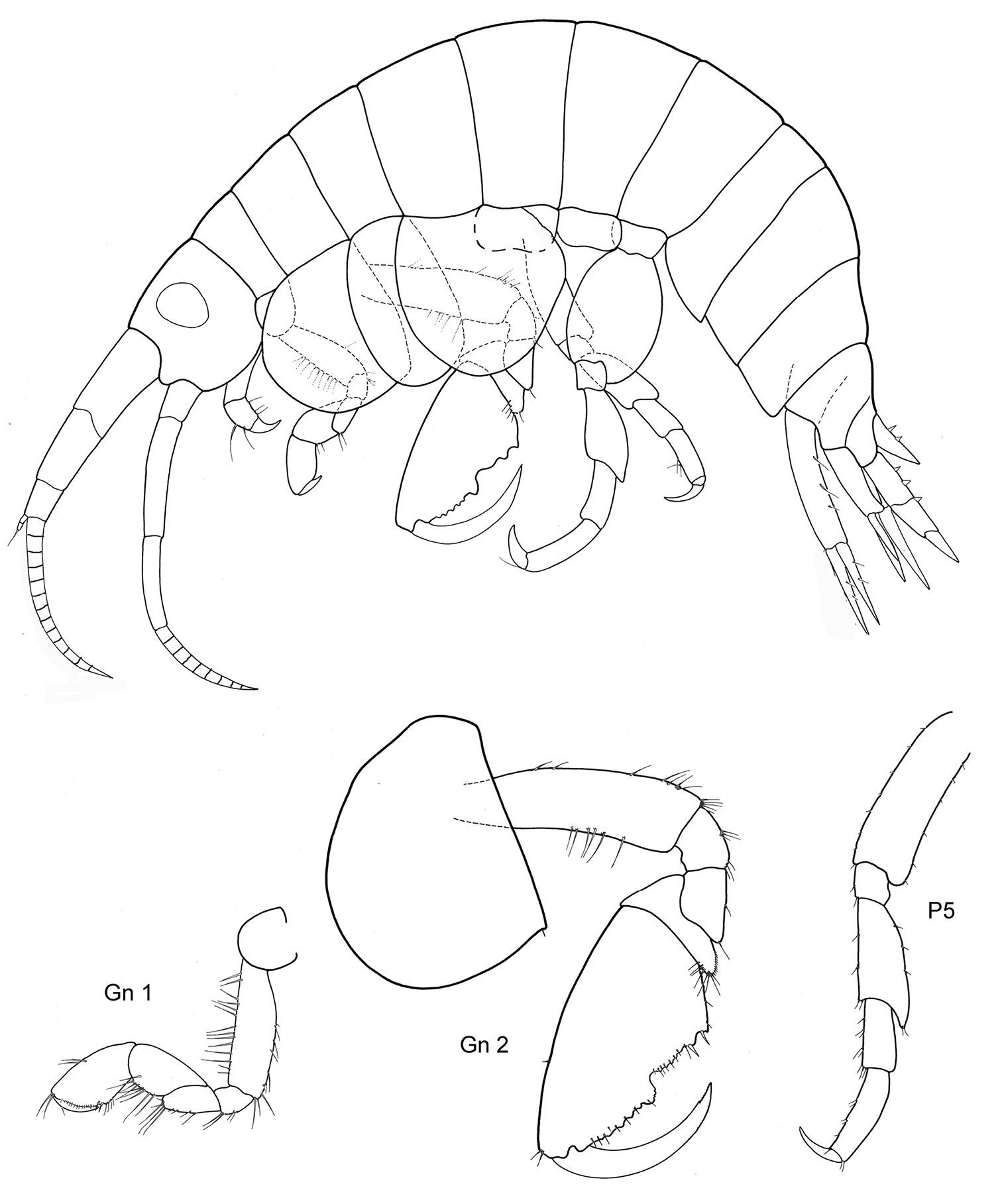
Figs 1, 2
Type material BMNH.
Body smooth. Eyes rounded, large.
Length: 5–9 mm.
Antennae. A1 less than half of body length, peduncle robust, art 1 length about three times the width; acc. flag. absent, flagellum 6–7 arts. A2 subequal in length to A1, peduncle robust, art 4 < art 5, flagellum about as long as or shorter than peduncle art 5, with 7 arts.
Mouthparts. Md palp with 3 arts, art 1 and 2 unusually long, art 3 much shorter than 1/3 length of art 2, with many distal setae. Mx 1, 2 unknown; Mxp IP not fused, 2/3 length of ischium; OP vanishing; dactylus long, subequal to propodus.
Coxae. Cx2 with rounded anterior margin, straight behind, angle rounded without tooth; Cx3 trapezoid-shaped, distally widening, Cx4 not excavated, anterior and posterior margin rounded, about as long as wide.
Gnathopods. Gn1, 2 propodi similar in shape, different in size. Gn1 dactylus ordinary; propodus with parallel margins, palm not defined, about twice as long as wide; carpus longer than propodus, subtriangular, longer than wide, proximally wider than distally; merus incipiently chelate; all articles densely beset with setae. Gn2 length of propodus more than 2/3 of Cx2; propodus subelliptical, twice the size of propodus Gn1; hind margin subequal to length of palm which has shallow incisions, palmar corner well defined by small tooth-shaped prolongation but no U-shaped incision. Dactylus same length as palm. Gn2 carpus shorter than wide, cup-shaped, merus not lobate.
Peraeopods. P4 merus anterodistal margin somewhat lengthened. P5 dactylus half length of slim propodus; merus posterodistal margin not reaching half of carpus length, basis rectolinear with short posterodistal lobe. P6 basis hind margin straight, with posterodistal lobe similar to P5, merus posterodistal corner acutely lengthened and somewhat widened, not reaching to half of carpus length. P7 basis proximally widened with lobe, distad narrowing with small posterodistal lobe, hind margin crenulate and excavated; merus lengthened and widened, reaching about half carpus length.
Uropods. U1 peduncle with many short robust setae, nearly twice as long as subequal rami; U2 peduncle also beset with many small robust setae, longer than longer ramus, rami somewhat unequal; U3 totally unarmed, peduncle longer than ramus, art 1 of ramus longer than art 2.
Telson. Not reaching end of peduncle U3; less than twice as long as wide; distally tongue-shaped rounded, naked.
Sexual differences. Small.
Distribution. Falkland Islands.
Ecology. From pharynx of a large ascidian.
Remarks. Malvinometopa porcellanus has extremely shortened A1, 2, no Mxp OP, a stout Gn1 and aberrant P6, 7: is this an adaptation to the life inside the pharynx of ascidians, where they certainly cannot swim but only crawl? All we know is that space there is at a premium.
Malvinometopa porcellana (K.H. Barnard, 1932): Discovery Reports St. 51, Falklands.
Malvinometopa porcellana (K.H. Barnard, 1932): Discovery Reports St. 51, Falklands.
Also this species is very sparsely described and figured. I found material at the Australian Museum Sydney, and compared it with one specimen deposited at the Verona Museum by Bellan-Santini. Both fit the written description by Schellenberg well, and this species clearly belongs in Metopoides:
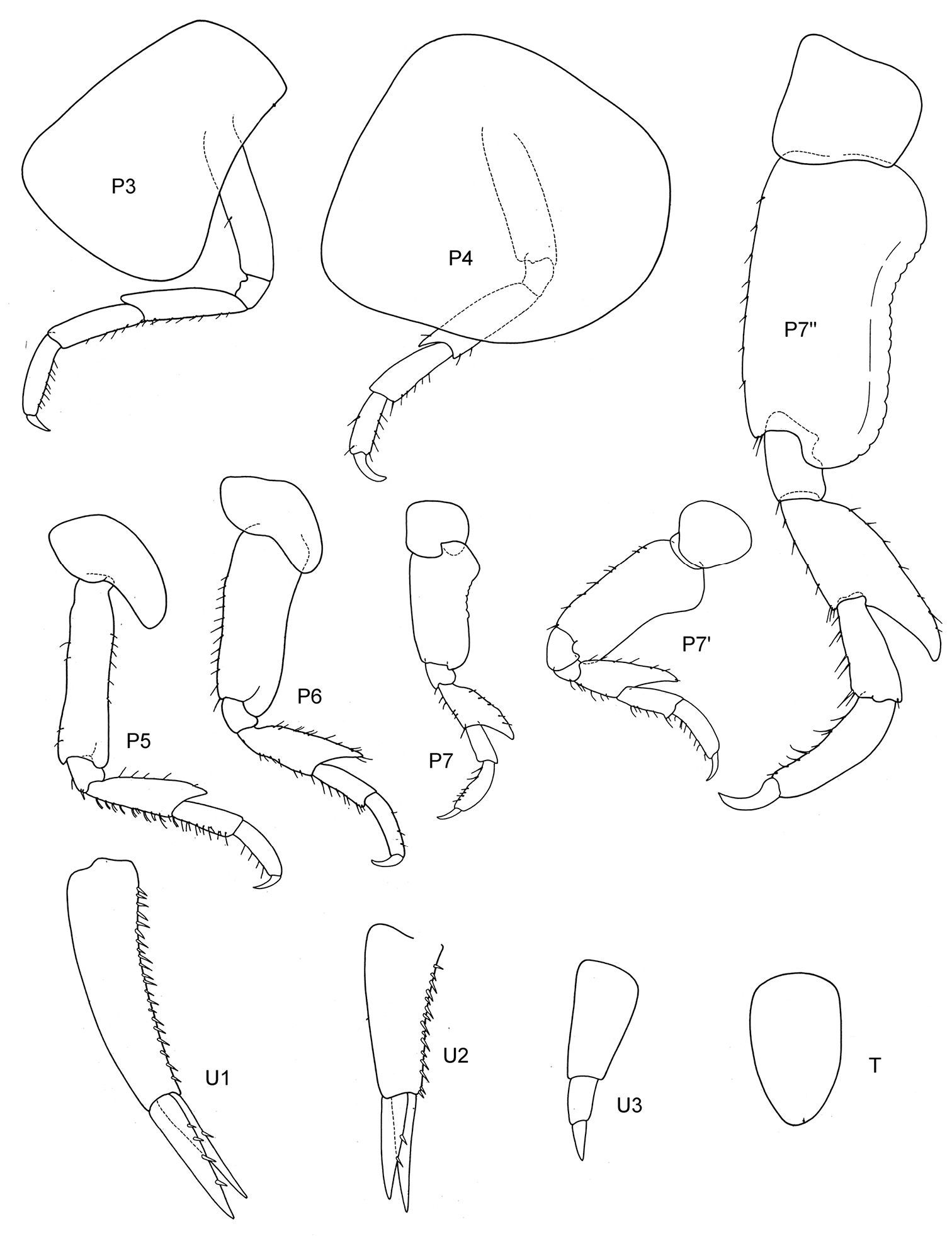
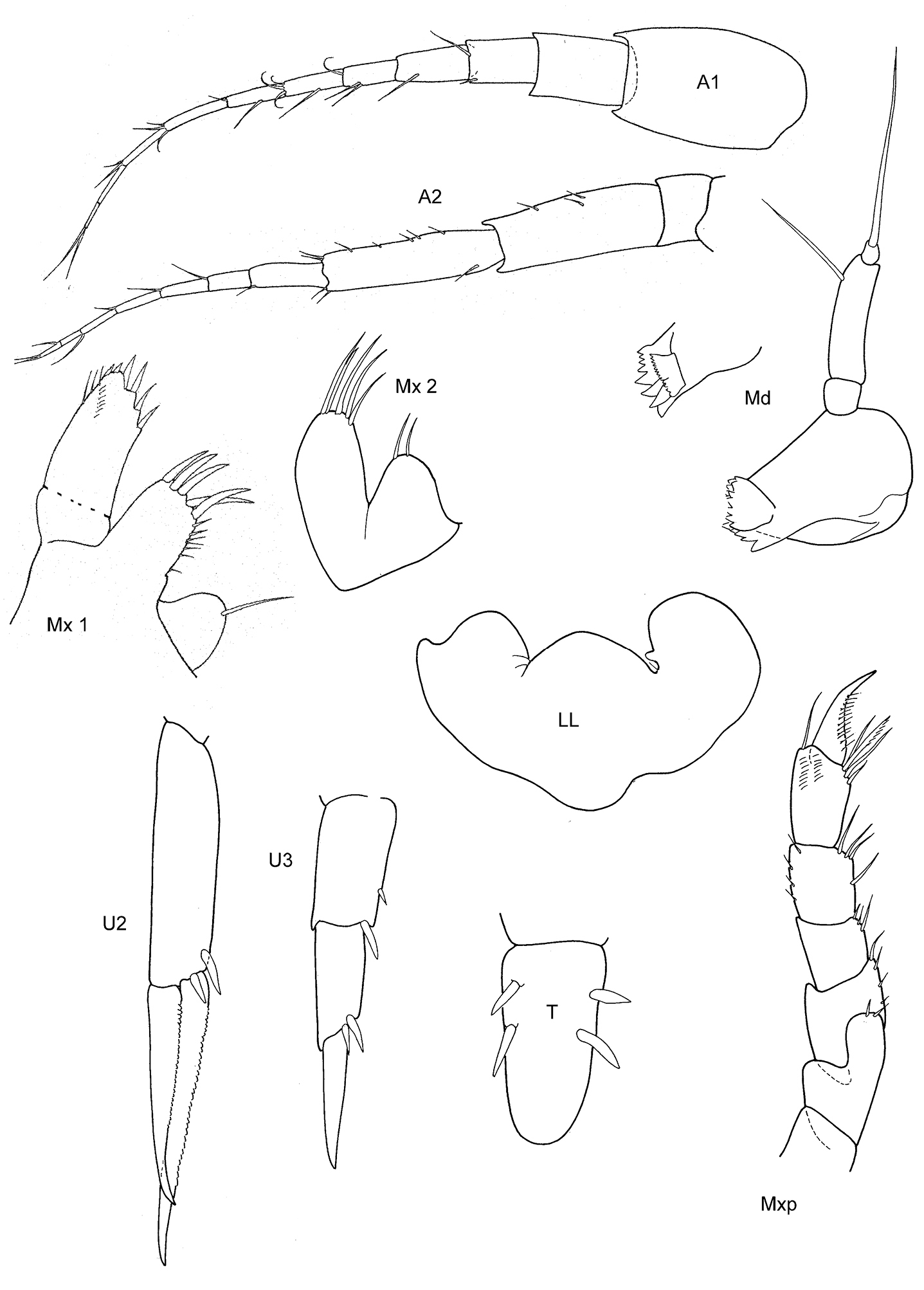
Figs 3, 4
Cape Bird, EBS, C3-C4, 70–100m, 14.12. 1971, several spec.; tide crack, near Cape Spencer, White Island, Ross Ice Shelf, 78°01'0"S, 167°20'0"E, 28.XII. 1976, coll. P. Ensor (AMS P 25504); Southern Rookery, Cape Bird, Ross Island, Antarctica (approx. 77°13'0"S, 166°27'0"E) AM P.80875 (1 slide); slide of “Metopoides sp. ”, Kerguelen, MNVCR.
Body smooth. Eyes rounded, medium size.
Length. 3–3, 5 mm.
Antennae. A1 less than 2/3 of body length, peduncle robust, art 1 shorter than three times wide; acc. flag. with 2 (very small) articles, flagellum 10 arts. A2 subequal in length to A1, peduncle robust, art 4 somewhat > art 5, flagellum about as long as peduncle art 5, with 7 arts (Walker: without acc. flag., A1 reaching to the middle of the flagellum of A2).
Mouthparts. Md incisor and raker spine row well developed; no clear molar cusp; palp with 3 arts, art 3 about 1/3–1/2 length of art 2, with 3 distal long setae (Walker: Md palp lacking third art, therefore creating a new genus Proboliella; but Schellenberg already noticed 1926: 323 fig. 41, that there is a well-developed third article). Mx 1 IP with 1 distal seta, OP with 6 strong robust setae, palp with 2 arts; Mx 2 inner plate ordinary, shorter than outer; Mxp IP not fused, 2/3 length of ischium; OP narrow, well developed, reaching more than half of merus length; dactylus long, subequal to propodus.
Coxae. Cx2 with rounded anterior margin straight behind, angle rounded with small tooth; Cx3 narrow with parallel margins, Cx4 not excavated, inferior and posterior margin rounded, about as long as wide.
Gnathopods. Gn1, 2 propodi different in size and shape. Gn1 dactylus ordinary; propodus with parallel margins, palm well defined (corner about 120°), somewhat longer than half length of propodus, about twice as long as wide; carpus shorter than propodus, triangular, longer than wide, merus incipiently chelate. Gn2 length of propodus more than 2/3 of basis in male, less in female; propodus subelliptical, twice the size of propodus Gn1; hind margin half length of palm which is in male and female with incisions, palmar corner well defined by acute tooth-shaped prolongation and U-shaped incision. Dactylus clearly shorter than palm, probably working together with robust setae of palmar corner. Gn2 carpus shorter than wide, cup-shaped, merus not lobate.
Peraeopods. P4 merus anterodistal margin somewhat lengthened. P5 dactylus long, weak, much longer than half of slim propodus; merus posterodistal margin not reaching half of carpus length, basis slender without lobe. P6 basis hind margin harmonically rounded, clearly longer than wide, merus posterodistal corner acutely lengthened but not widened, not reaching to half of carpus length. P7 basis and merus similar to P6.
Epimeral plates. Ep3 posterodistally lengthened to triangular corner.
Uropods. U1 peduncle slightly longer than subequal rami, with many robust setae; U2 peduncle longer than shorter ramus, rami clearly unequal; U3 peduncle shorter than ramus, first article of ramus shorter than peduncle, ramus art 2 about ¾ of art 1.
Telson. Not reaching end of peduncle U3; about twice as long as wide; distally triangulary pointed, medio-laterally with 2–3 robust setae.
Sexual differences. Probably small.
Distribution. Antarctica, Hut Point near Mc Murdo, 77.47°S (
Ecology. Steeply sloping rock bottom with encrusted bryozoans and hydroids.
Remarks: As this species clearly has an accessory flagellum (although tiny), unspecialized gnathopod propodi and neither much lengthened nor widened merus on P5–7, it has to be placed in the genus Metopoides, and even is a very “typical” representative of that genus.
Metopoides typicus (Walker, 1906): Cape Bird, Southern Rookery; AMS.
Metopoides typicus (Walker, 1906): Cape Bird, Southern Rookery; AMS.
Shortly after the war the possibility to check foreign literature was restricted. As only few characters were illustrated, the character states included in the matrix most probably were not always appropriate.
Until type material of Metopa rotunda can be checked, this species is synonymized with question mark to Stebbing’s species “Metopa rotunda”. In any case it should belong to Metopoides for many plesiomorphic character states.
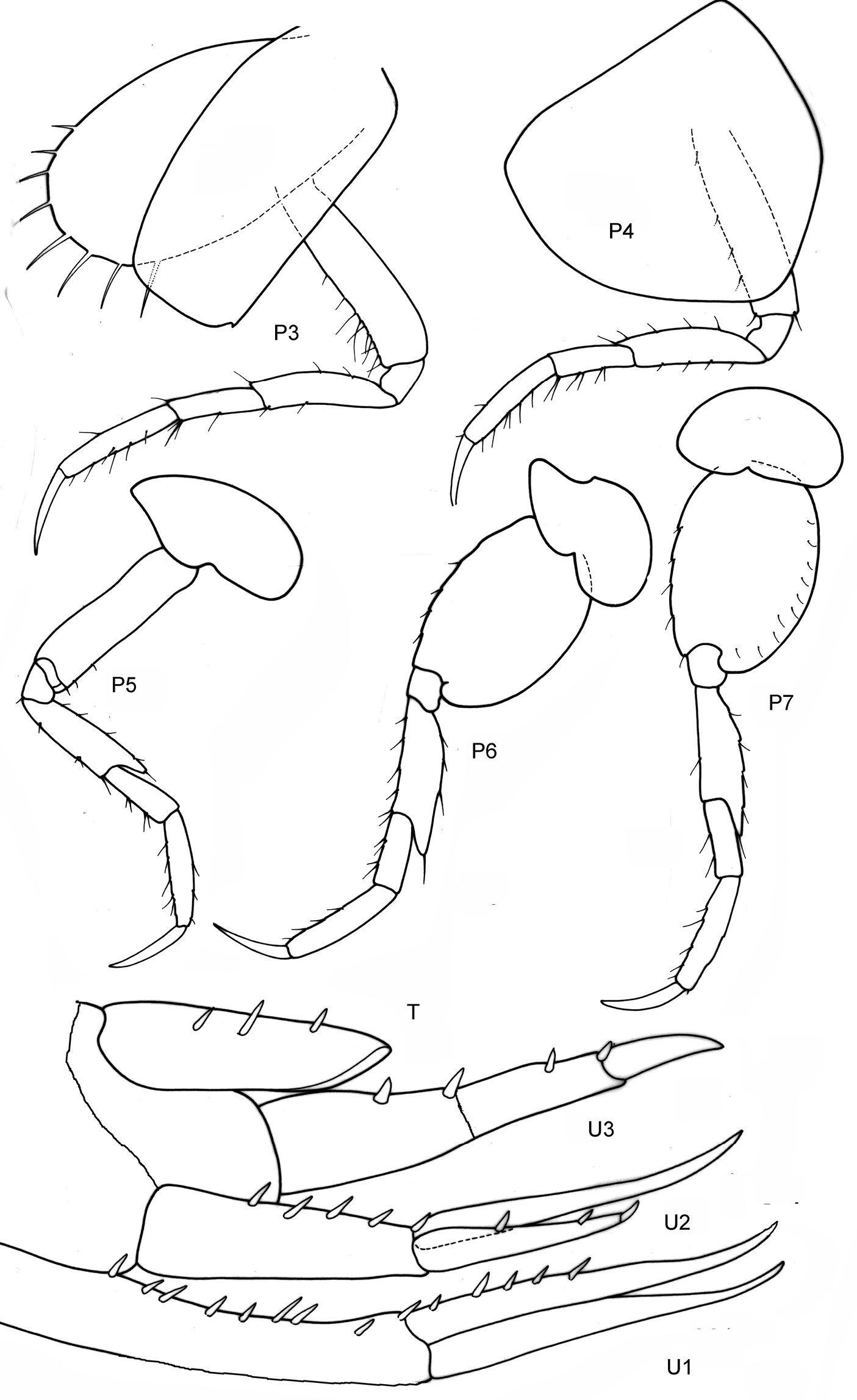
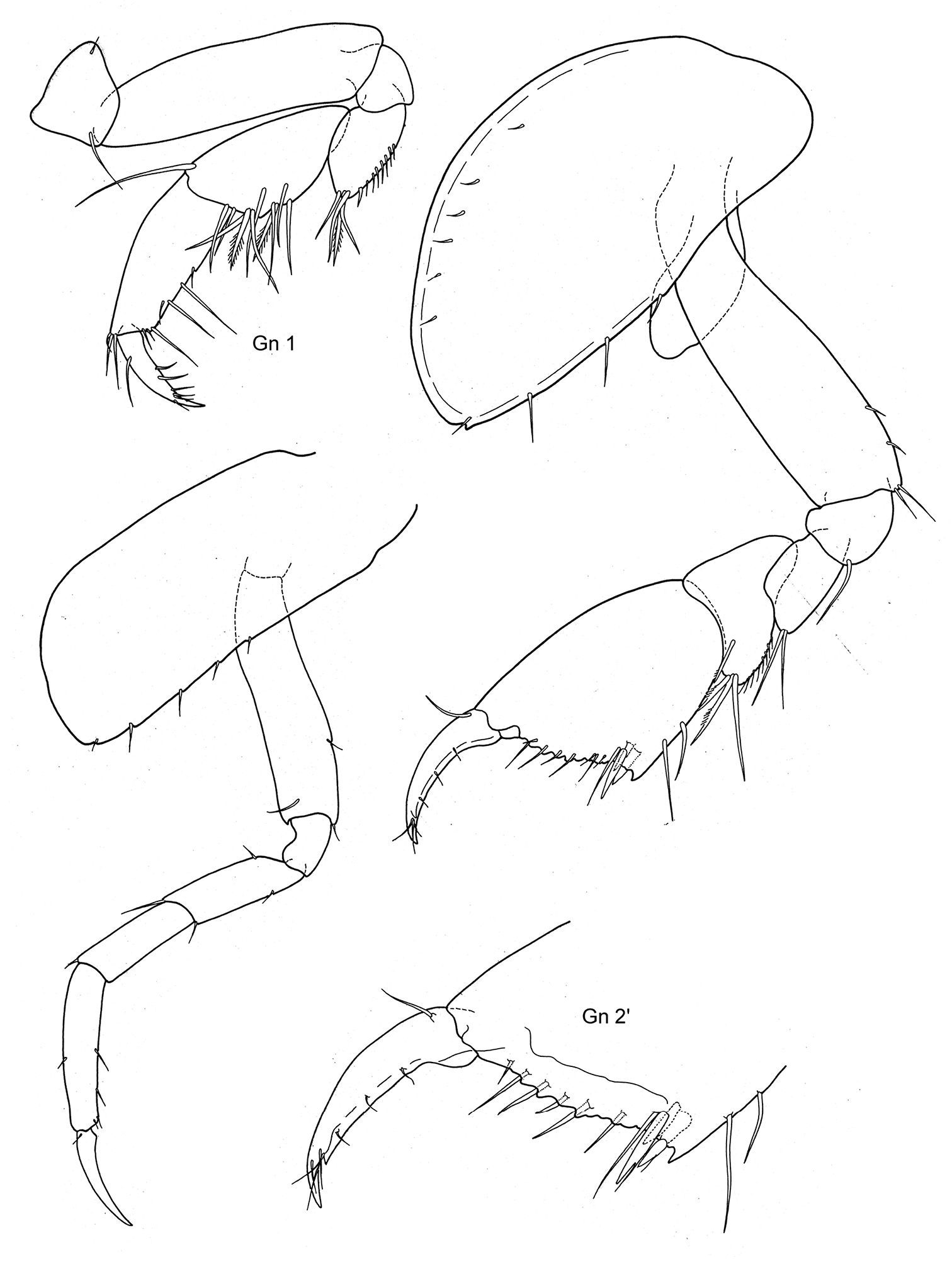
Fig. 5
Type material of Proboloides stephenseni MNVCr.
Body smooth. Eyes rounded, large.
Length. Male 3, 5 mm.
Antennae. A1 = A2, A1 less than half of body length, peduncle robust, art 1 length about three times the width; acc. flag. with 2 arts, flagellum 12 arts. A2 subequal in length to A1, peduncle robust, art 4 ≥ art 5, flagellum about as long as peduncle art 5, with 9 arts.
Mouthparts. Md palp with 3 arts, art 1 and art 3 subequal, art 3 longer than 1/3 art 2, with 2 long distal setae. Mx 1 palp with 2 arts. Mxp IP separated, OP longer than half ischium; dactylus as long as propodus.
Coxae. Cx2 with rounded anterior margin, straight posterior one, angle rounded with small tooth; Cx3 with parallel margins, Cx4 not excavated, anterior and posterior margin rounded, about as long as wide.
Gnathopods. Gn1, 2 propodi similar in shape, different in size. Gn1 dactylus ordinary; propodus triangular, palm well defined, about twice as long as wide, about as long as hind margin; carpus shorter than propodus, trapezoid, longer than wide, with parallel margins; merus with very short distal free margin. Gn2 length of propodus = Cx2; propodus more than twice the size of propodus Gn1; hind margin much shorter than length of palm which has shallow incisions and crenulations, palmar corner well defined by small tooth-shaped prolongation but no U-shaped incision. Dactylus shorter than palm. Gn2 carpus shorter than wide, cup-shaped, merus not lobate.
Peraeopods. P5 dactylus half length of slim propodus; merus posterodistal margin not reaching half of carpus length, basis rectolinear, width proximally and distally subequal, posterodistally rounded, but not lobed. P6, 7 similar, basis hind margin rounded, merus posterodistal corner shortly lengthened and somewhat widened, not reaching to half of carpus length.
Epimeral plates. Ep3 posterodistal corner rectangular, but rounded.
Uropods. U3 peduncle shorter than ramus, art 1 of ramus longer than art 2; peduncle with one short robust seta distally, ramus art 1 with 2 robust setae.
Telson. not reaching end of peduncle U3; less than twice as long as wide; distally pointed, marginally two robust setae.
Sexual differences. Females unknown.
Distribution. Antarctica, 70°23'0"S, 82°47'0"W (Proboloides stephenseni Ruffo, 1949). South Africa (Proboloides rotunda Stebbing, 1917).
Depth. 42 fathoms = 76, 8 m (Proboloides rotunda,
After
Proboloides stephenseni Ruffo, 1949: Antarctica (70°23'0"S, 82°47'0"W); MCV.
Although the shape of the gnathopods (especially the simple Gn1) creates doubt if it could not belong in Metopa or Stenula,
checking of the mouthparts showed at least that this species has a Md
palp with 3 arts and a palp of Mx1 with ? 2 arts (although the
articulation is not clear, see
At the Verona Museum I found a tiny specimen called “Metopa sp.”(1, 5 mm) which is extremely similar to Proboloides holmesi, except the rounded palmar corner (see Fig. 6, 7 and compare to
?Proboloides holmesi Bousfield, 1973: Raunefjorden near Bergen; MCV.
?Proboloides holmesi Bousfield, 1973: Raunefjorden near Bergen; MCV.
S of Cape Cod; Vineyard Sound, Elizabeth Islands, Buzzard Bay (
Mainly on sandy and shelly sand bottoms, among hydroids and bryozoans, in depth of 5–30 m (
During a stay at the Copenhagen Museum I checked Stephensen’s type material of this species, as
Proboloides aequicornis (Stephensen, 1931): after typical material from Atlantic, between Faroes and Iceland.
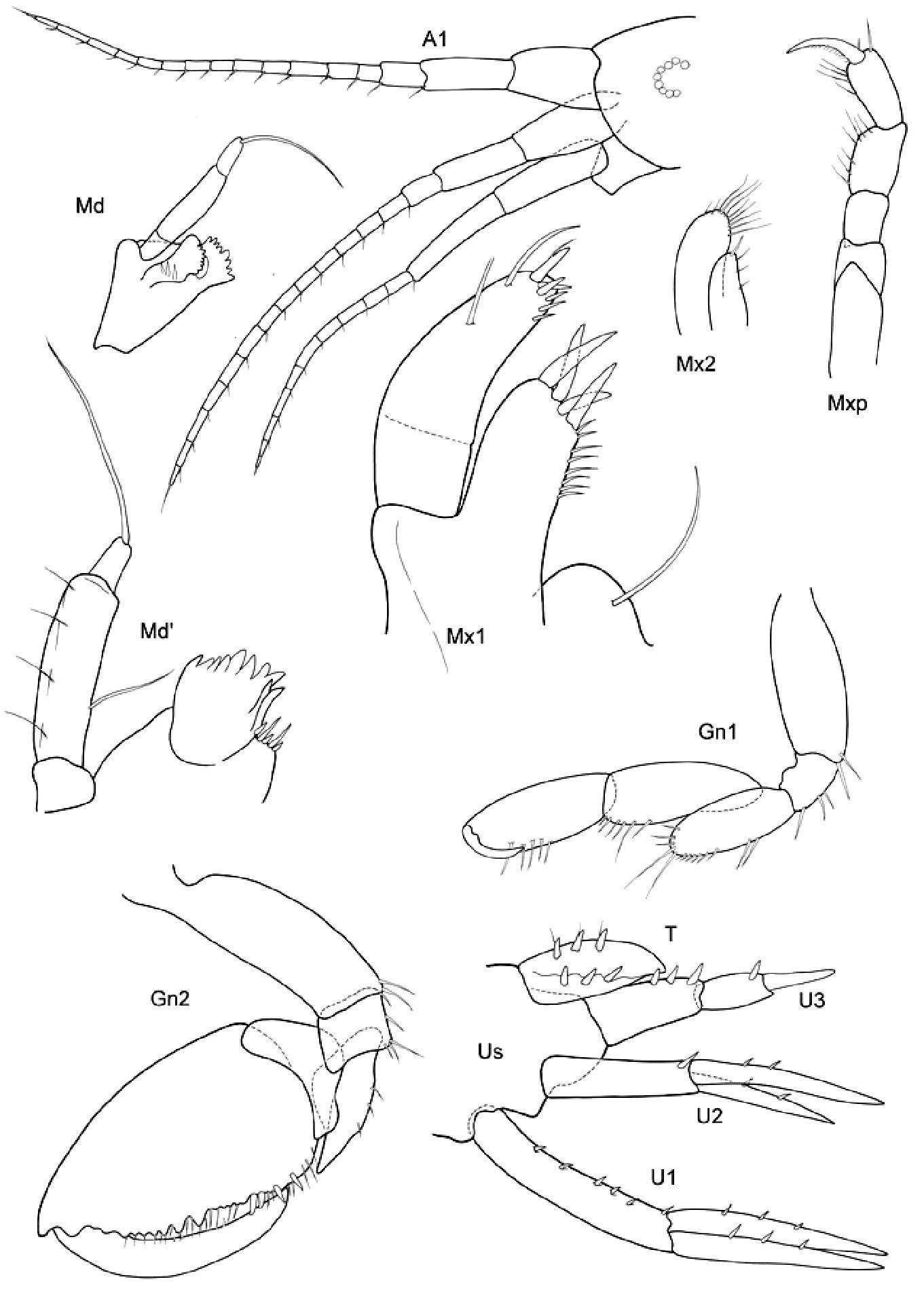
Figs 8–10
Type material of Stenothoe aequicornis ZMUC.
Body smooth. Eyes rounded, small.
Length. Male 5 mm.
Head. Lateral cephalic lobes bluntly angular.
Antennae. A1 subequal A2 or A1 scarcely > A2. A1 peduncle robust, art 1 length about 2–3 x the width; art 3 only 1/3 of art 1 length; acc. flag. with 2 arts, flagellum about 1, 5 x length of peduncle, 12–14 arts. A2 subequal in length to A1, peduncle subequal to flagellum, peduncle robust, art 4 ≥ art 5, flagellum with 9–11 arts.
Mouthparts. Md palp with 3 arts, art 1 and art 3 subequal, art 3 longer than 1/3 art 2, with 1 long distal seta. Mx 1 palp with 2 arts (but articulation not easy to see, cf. fig. 8, 9). Mxp IP separated, OP longer very short; dactylus as long as propodus.
Coxae. Cx2 with rounded anterior margin, straight or even somewhat concave behind, front angle rounded without tooth; Cx3 with trapezoid-shaped margins, Cx4 not excavated, anterior and posterior margin rounded, wider than long.
Gnathopods. Gn1, 2 propodi different in size and shape. Gn1 dactylus ordinary; propodus elongate, about 3× as long as wide, palm well defined, much shorter than hind margin; carpus longer than propodus, triangular, nearly 3× longer than wide, with parallel margins; merus with very long distal free margin. Gn2 length of propodus > Cx2; propodus about 3× the size of propodus Gn1; hind margin much shorter than length of palm which has shallow incisions and crenulations, palmar corner scarcely defined by group of robust setae, no U-shaped incision. Dactylus subequal to palm. Gn2 carpus very short, cup-shaped, merus acutely lobate.
Peraeopods. P5 dactylus > half length of slim propodus; merus posterodistal margin not reaching end of carpus length, basis rectolinear, width proximally and distally subequal, posterodistally rounded, but not lobed. P6 basis hind margin straight, merus posterodistal corner acutely lengthened and widened, reaching to end of carpus length; P7 similar to P6, but basis hind margin regularly rounded.
Epimeral plates. Ep3 posterodistal corner acute, but rounded at the apex.
Uropods. U3 peduncle shorter than ramus, art 1 of ramus subequal to art 2; peduncle with 3 robust setae distomarginally, ramus art 1 with 1 robust seta.
Telson Not reaching end of peduncle U3; less than twice as long as wide; distally pointed, marginally 3 robust setae.
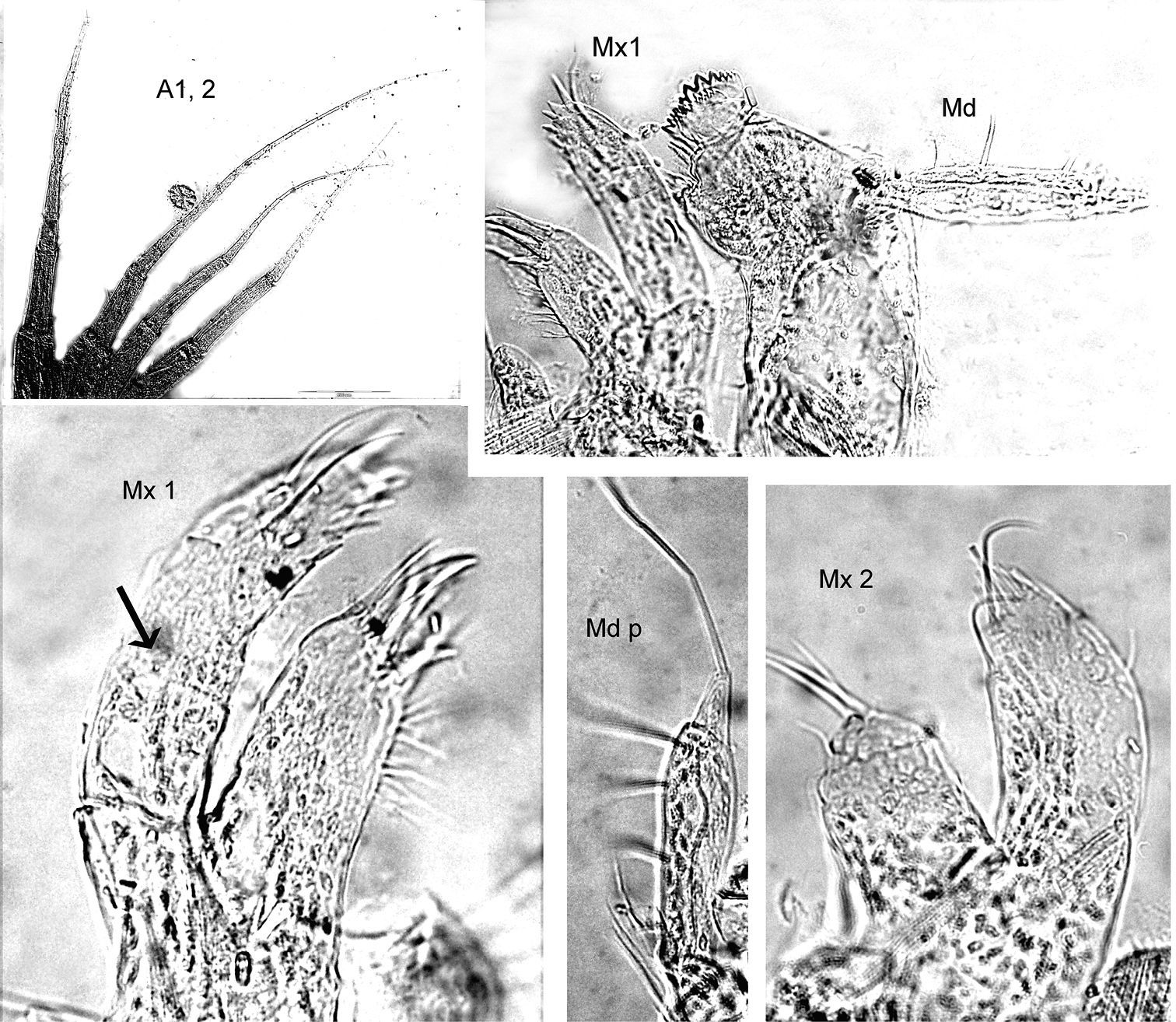
Proboloides aequicornis (Stephensen, 1931): as above, photographs of the material taken with Olympus BX51 with cell imaging software.
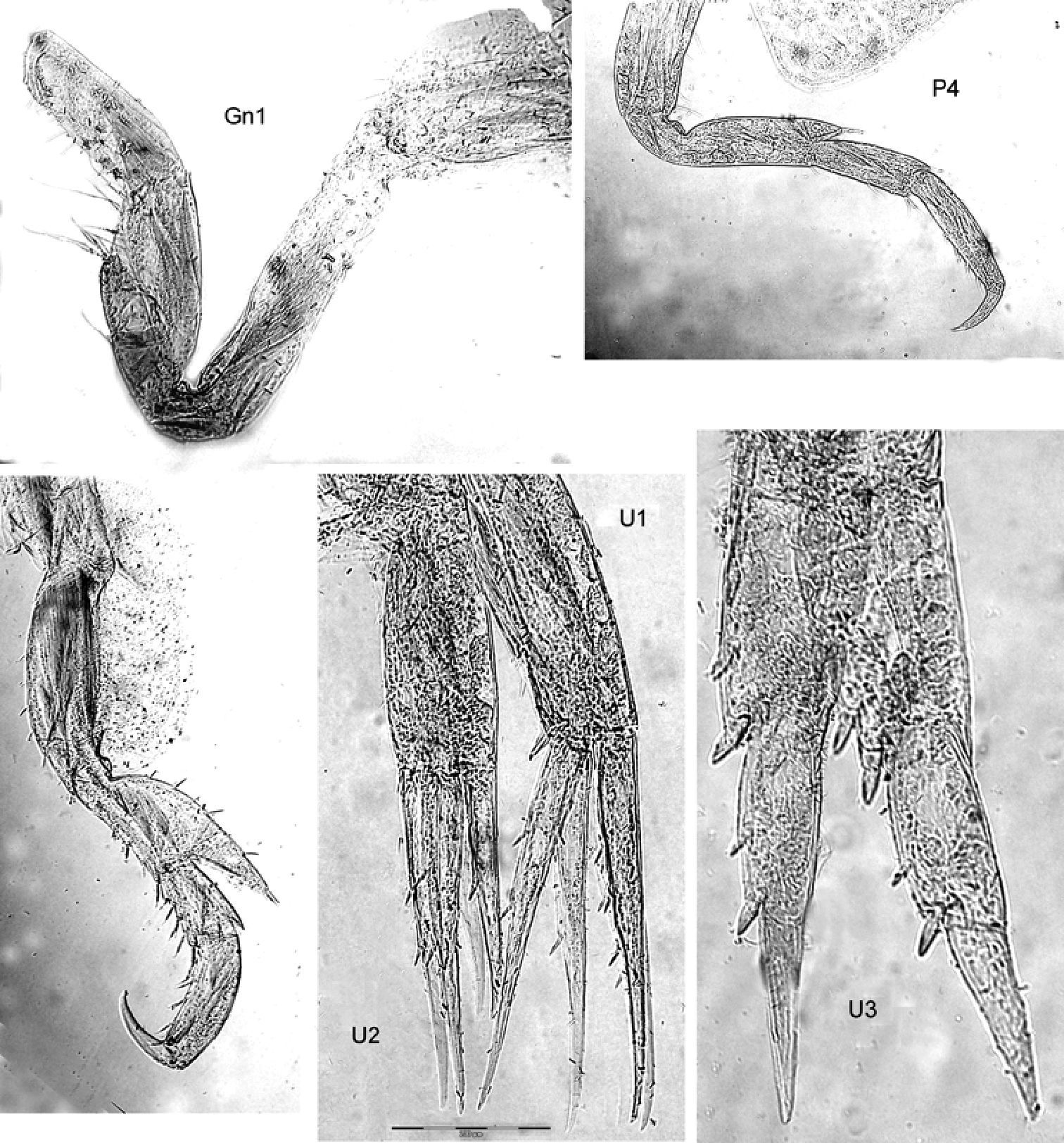
Proboloides aequicornis (Stephensen, 1931): as above, photographs of the material taken with Olympus BX51 with cell imaging software.
Females unknown.
Between Faroes and Iceland, 375 m depth.
At the Verona Museum I looked for the only species of the genus Torometopa cited in
Torometopa? armata Barnard & Karaman, 1991: 700
It is the unique (female) type specimen from Îles Glorieuses N of Madagascar, from 3718 m depth.
Unfortunately I could not examine the slide and confirm the drawings of the sub-rectangularly widened Gn1 propodus and the 1-articulated Md palp, both very unusual characters in our treated group, as this type must be deposited elsewhere.
But at the Victoria Museum Melbourne I found a species from the Bass Strait from 770 m clearly belonging also to this basic species-complex of stenothoids, having a posterodistal lobe on P5 basis. To my big surprise it turned out that this species too had a 1-articulate Md palp.
As this character- combination does not fit any of the extant stenothoid genera, a new one was erected:
urn:lsid:zoobank.org:act:5DA70786-2BC5-415E-9953-E2FA6850218F
Victometopa rorida sp. n.
Victometopa armata (Ledoyer, 1986), comb. n.
Md palp with 1 art, Mxp OP reduced. P5 basis rectolinear, with posterodistal lobe. P6, 7 basis widening.
The stem -metopa combined with the first letters of “Victoria”, for expressing admiration for the rich collection at the Victoria Museum Melbourne (Australia).
Male 4.4mm. Cruise 79-K-1, Stn 34, 30°38.7'0"S, 148°49.4'0"E, Flinders Canyon, eastern Bass Strait, 770 m, 27.3.1979. Sediment: shell/sand, gear: dredge. MVM J 39597
Male 4.2mm. Same locality.
The Latin adjective roridus means “set with dew” and should stress the “pearls” on the Cx3 in this species.
Length. 4.2 - 4.4 mm
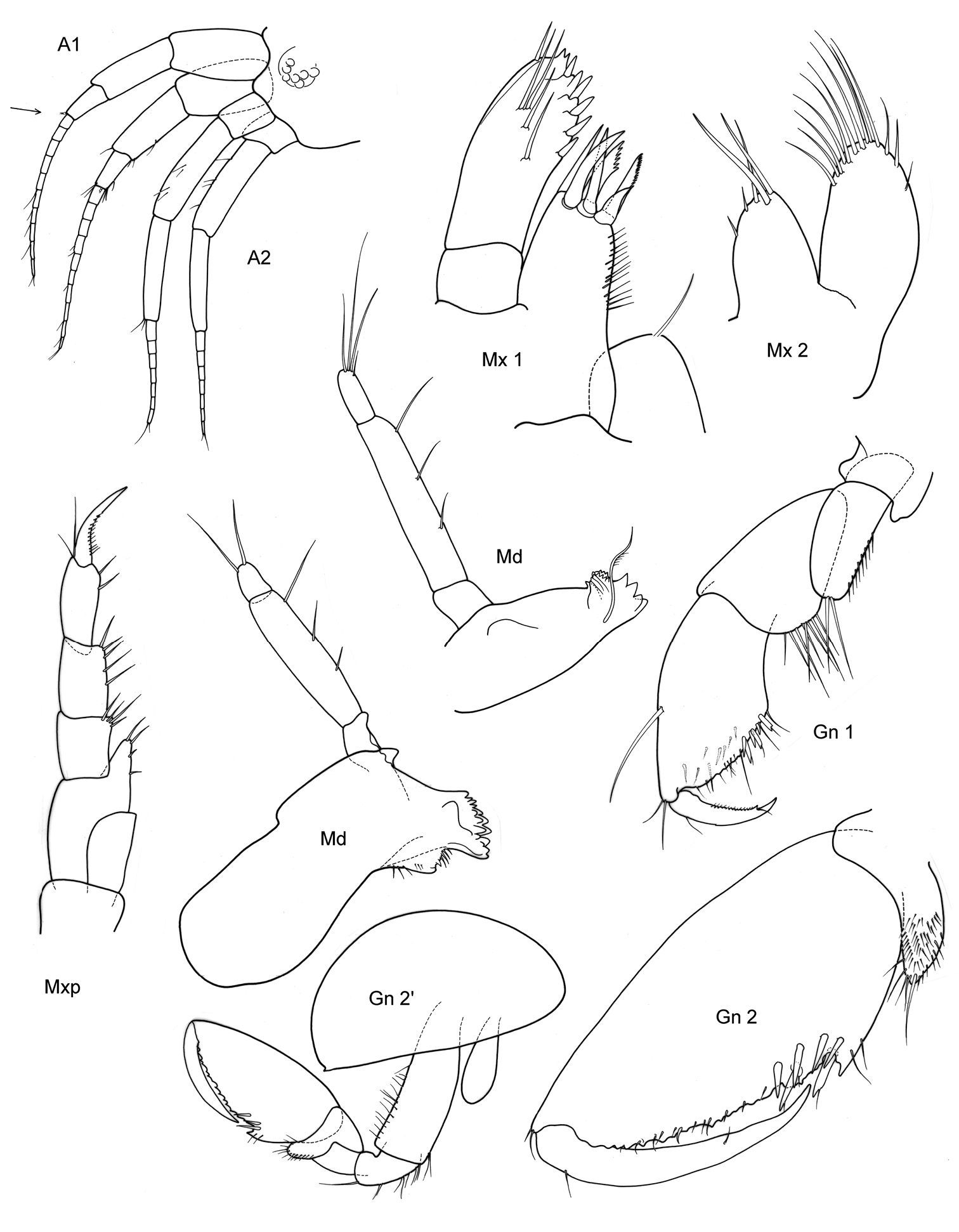
Body.Smooth. P3–7 all clearly prehensile, with falcate-concave, strikingly long merus and strong dactylus opposing with spinose propodus.
Head. Lateral cephalic lobes subacute or blunt, triangular. Eyes rounded, medium.
Antennae. A1: body length ≥ 0.66 body, ped. art 1 l:b >3; ped. art 2 ≥art 1; art 1 =cephalon; ped. art 3 ≤0.3 art 1; acc. flag. absent; flagellum arts 11–20. A2 ped .art 5> flag., ped. art 4 =art 5, nr. flag. arts ≤ 9 (A2 broken into pieces, thus indications not totally sure).
Mouthparts. Mdb palp one long article, on tip a fine articulation-line visible, marginally no setation, distally 1 long and 1 shorter seta. Mxp outer plate reduced.
Coxae. Cx2:Cx1 ratio of length >3. Cx2 l:b (l=parallel post. margin) < 1.5. Cx3 unusually widened, nearly as long as wide, distoposterior margin with string “pearls” or stridulation ridges. Cx4 l<b, distally not excavated.
Gnathopods. Gn1 dactylus ordinary. Gn1 palm subequal to half propodus length; propodus palm angle 180–150°, blunt; propodus shape rounded, l≤ 2b; carpus l>2b; merus free projecting; carpus longer than propodus. Gn1 propodus < 0.33 Gn2 propodus. Gn1, 2 propodus shape different. Gn2 propodus ≥coxa and basis in male, palm in male smooth, only at dactylus-insertion some serrations; carpus very short, merus small, subquadrangular, not lobate.
Peraeopods. In all dactylus clearly longer than propodus. P3, 4 merus long, falcate curved, nearly twice the length of propodus. P5 basis distally somewhat widened but strongly lengthened to lobe maximal to minimal breadth 1.4–1.6; merus also nearly twice the length of propodus, posterodistal tip neither lengthened nor widened. P6 basis hind margin with straight margins, merus anterior and posterior margin subequal, distally not lengthened, reaching no carpus. P7 basis wider than in P6, but posterior margin also rather straight.
Epimeral plates. Ep3 posterodistally rectangular corner.
Urosome. U1 rami equal. U1 peduncle longer than ramus. U2 rami different, the shorter is longer than 0.66 % of the longer one, peduncle is longer than rami, spination weak. U3 with very long peduncle, much longer than ramus; ramus art 1:2 <1, spination poor.
Telson. l:b ≤2, distally rounded, marginally with two strong robust setae.
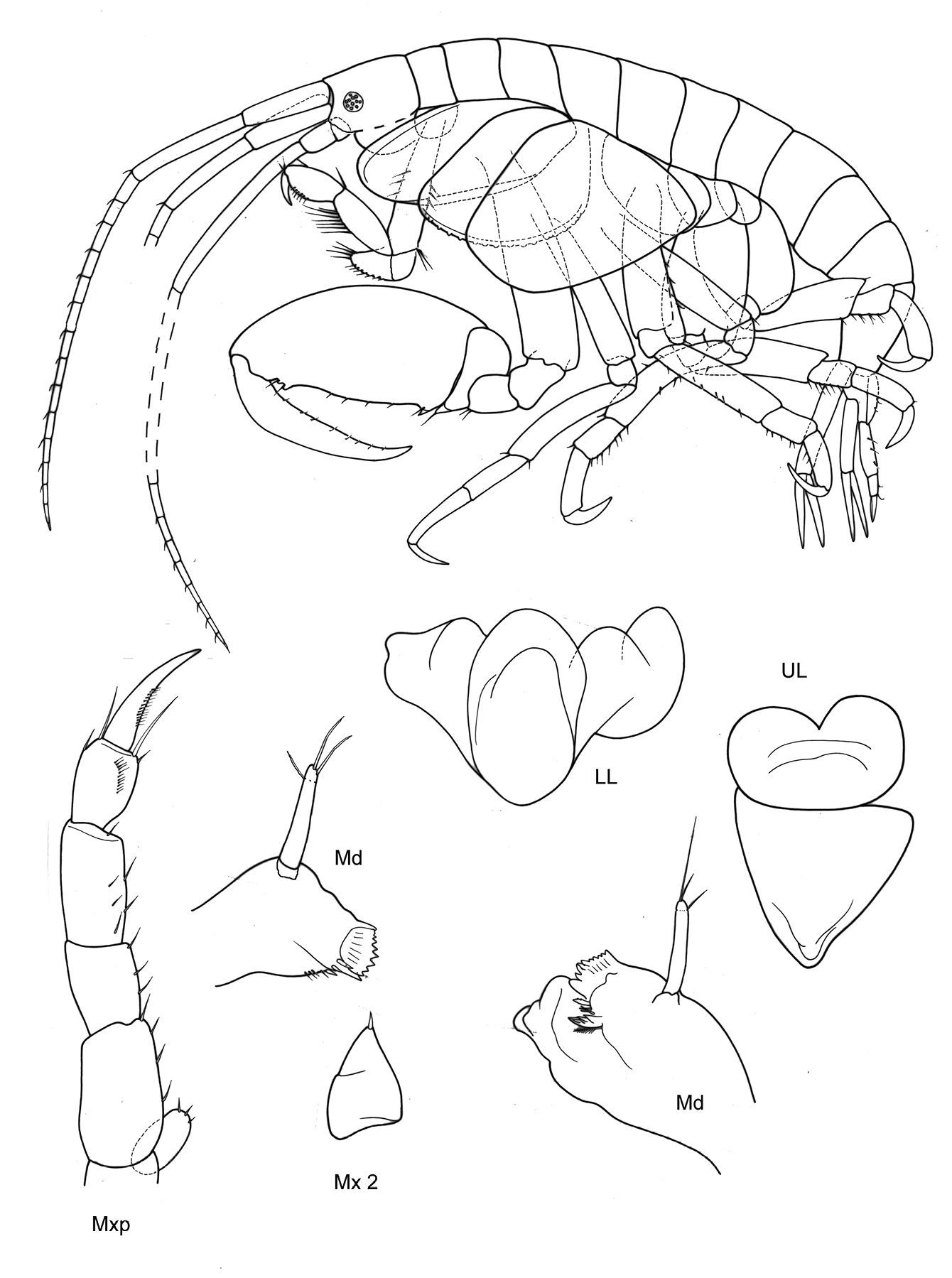
Victometopa rorida gen. n. sp. n.: Habitus ? male 4.4 mm; mouthparts UL, Mx1, 2 Md, LL, Mxp.
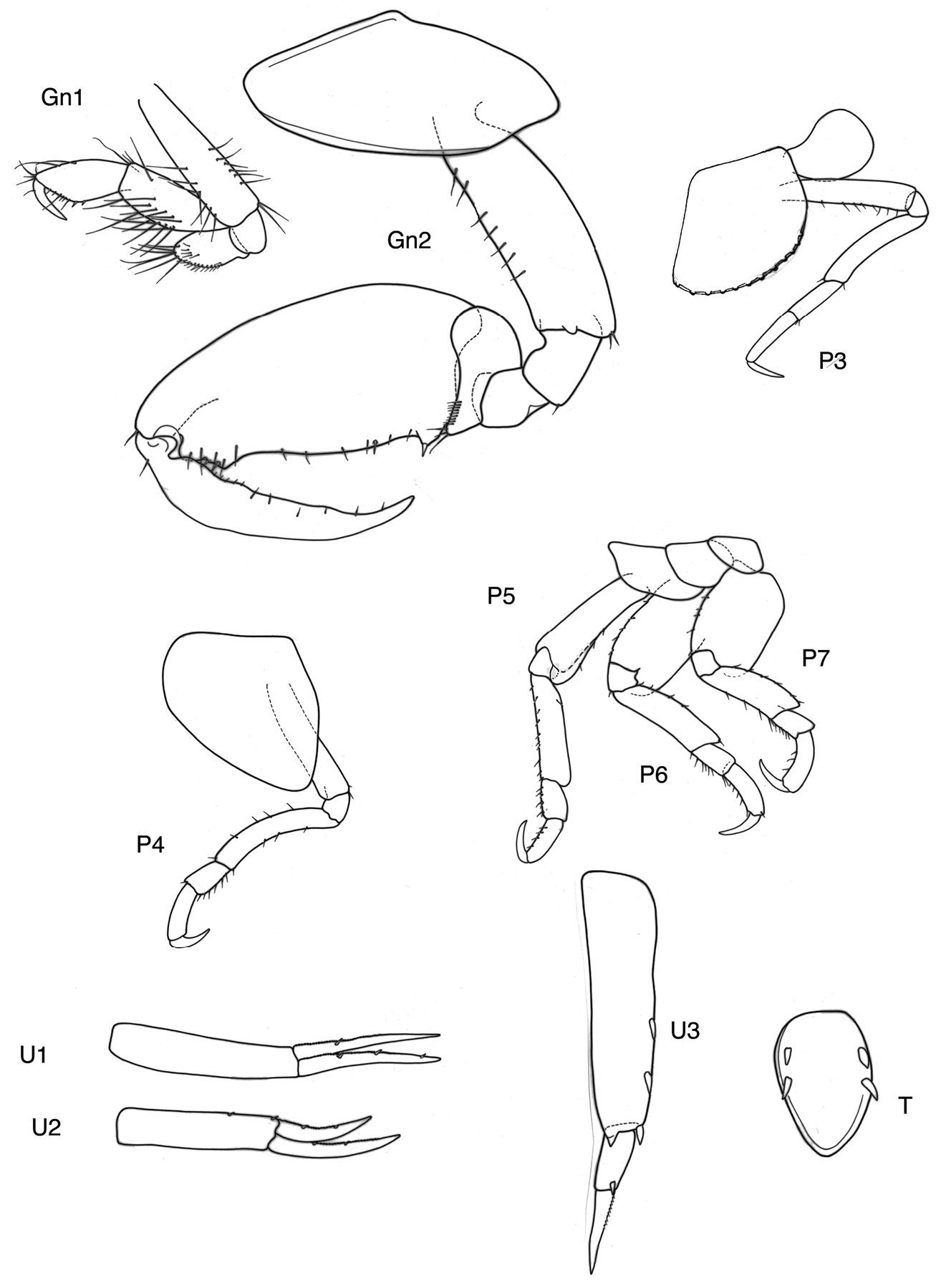
Victometopa rorida gen. n. sp. n.: Gn1, 2 = gnathopod 1, 2; P3-7 = peraeopods 3-7; U1-3 = uropods 1-3; T = telson.
Figure 13, 14.
A matrix of 38 species and 61 characters was built (Fig. 13): all presently known species in the genera Proboloides, Torometopa and Scaphodactylus were included. A hypothetical Gammarus species was chosen as out-group (see also

Matrix for 38 taxa and 61 characters (see description for character states in the text).
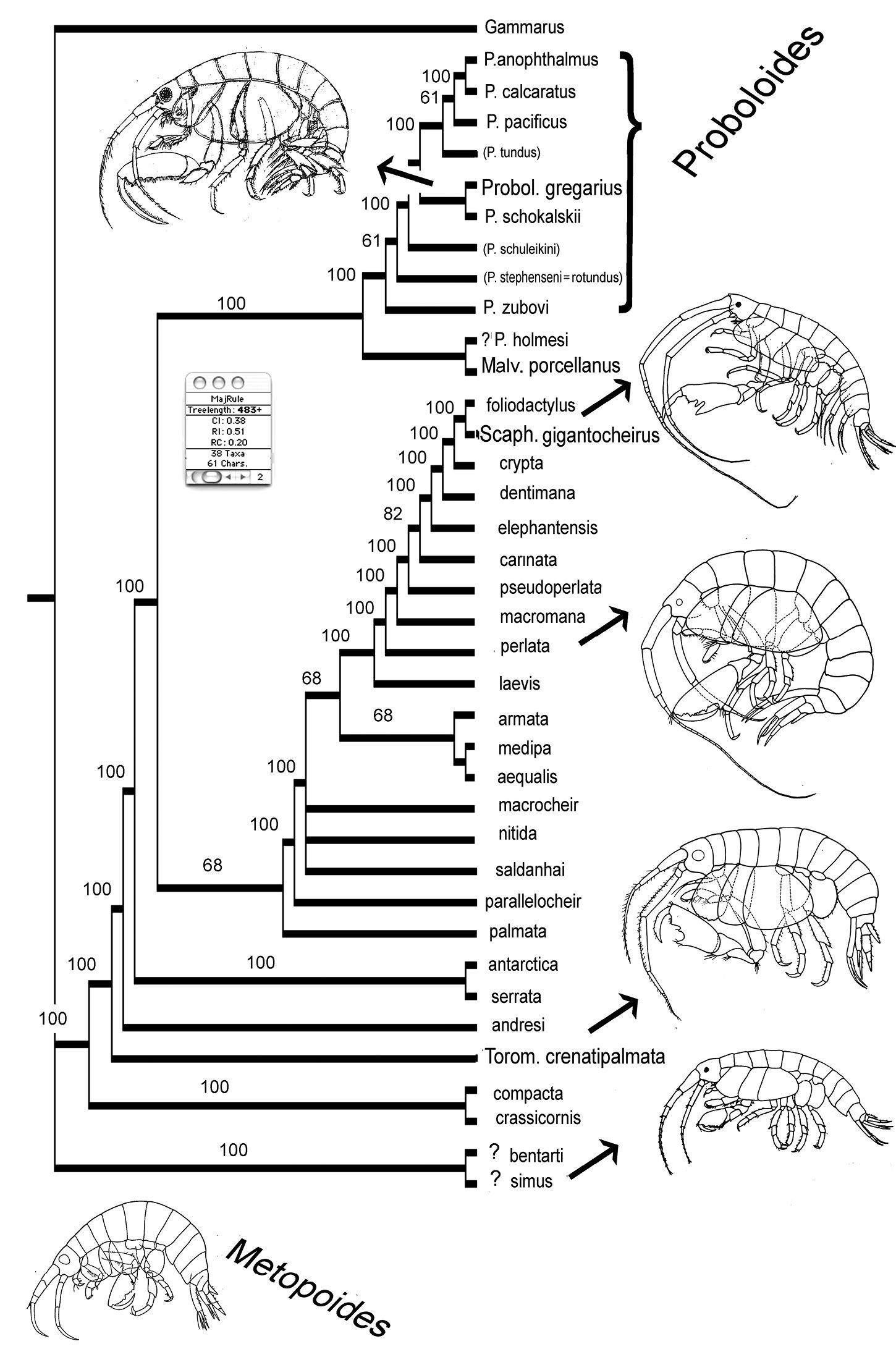
Heuristic analysis using 38 taxa and 61 characters: majority rule tree of 28 trees with length = 483. Genera abbreviated with more than one letter indicate type species; names written with small letters indicate that species are put in synonymy.
The programs MacClade 4.06 (
Heuristic search settings:
Optimal criterion = parsimony
Characters were unweighted and unordered
Gaps are treated as “missing”
Multistate taxa interpreted as polymorphism
Starting tree(s) obtained via stepwise addition
Addition sequence: random
Number of replicates = 50
Starting seed = 1191736759
Number of trees held at each step during stepwise addition = 7
Branch-swapping algorithm: tree-bisection-reconnection (TBR)
Steppest descent option not in effect
Initial “MaxTrees” setting = 200 (will be auto-increased by 100)
Branches collapsed (creating polytomies) if maximum branch length is zero
‘MulTrees’ option in effect
Topological constraints not enforced
Trees are unrooted
Total number of rearrangements tried = 46863460
Score of best tree(s) found = 486
Number of trees retained = 28
Tree length = 483
CI = 0, 38
RI = 0, 51
RC = 0, 20
ResultsAt the beginning of the present analysis 16 species were cited for Proboloides:
Eight species were reported from the Atlantic or Arctic Ocean: Proboloides calcaratus, Proboloides clypeatus, Proboloides grandimanus, Proboloides gregarius, Proboloides holmesi, Proboloides schokalskii, Proboloides schuleikini, Proboloides zubovi. In most species at least the females have a pronounced palmar corner in Gn2, while Gn1 is weak and slender. In the deep-water species Proboloides calcaratus and Proboloides gregarius the males have Gn2 propodus + dactylus very much lengthened and the eyes large, Cx3 has no parallel margins, but becomes wider distally and is not much narrower than Cx 4, while Proboloides holmesi has a narrow Cx3 with parallel margins, very different from Cx4.
Proboloides clypeatus must remain a species dubia, as it is too poorly described.
The species Proboloides gregarius and Proboloides schuleikini (originally only subspecies of Proboloides gregarius) show differences only in the even more elongated Gn1 in the latter, and I think Proboloides schuleikini is a big female of Proboloides gregarius. I have also my doubts about the description of Proboloides grandimana (Bonnier), where all details match Proboloides gregarius except the big and triangular Cx1 which should be even larger than Cx2, an extremely unusual character in stenothoids; it seems quite probable that this is an error and that Cx2 is repeated. - Branch et al. (1991) illustrate a Proboloides sp. with U3 with 2 rami, which undoubtedly is also an error of the drawing.
Thus the only certain Atlantic-Arctic members are Proboloides calcaratus, Proboloides gregarius, Proboloides schokalskii and Proboloides zubovi.
There are 5 nominal Proboloides species from S-Africa and the Antarctic-Subantarctic region: Proboloides porcellanus, Proboloides rotundus, Proboloides stephenseni, Proboloides typicamimus, Proboloides typicus. The species Proboloides stephenseni and Proboloides rotundus are morphologically similar and may be synonymized; Proboloides typicus is redescribed and both could be transferred to Metopoides, as they have more plesiomorphic character states than members of Proboloides. - Proboloides porcellanus is redescribed and is the type of a new monotypic genus Malvinometopa.
The remaining species Proboloides typicamimus would then be the only member of the genus Proboloides living in the Antarctic, but it seems quite probable that also this species does not belong to this genus. But it is incompletely described though, based on a single specimen and knowledge about its character states is still very inadequate.
There are two nominal Pacific species of Proboloides remaining, Proboloides tundus and pacificus,
which may well be synonymous: the shape of Gn2 matches (the only
illustrated detail of the first), and the written description of the
shape of P5-7 merus in Proboloides tundus “narrow, scarcely produced” matches the description by
Proboloides anophthalmus is the only species living in the Indian Ocean (Madagascar). It is very similar to the Atlantic species Proboloides? holmesi, however there are some differences in the shape of T, the presence of pearls on the Cx3 margin and the shape of A1, 2. This deep-sea species has no eyes.
Two species can be added here:
“Metopa nordmanni” sensu
Proboloides aequicornis (Stephensen, 1931), as explained above.
After the thorough check of 16 species, eight (plus one doubtful member) remain belonging to Proboloides and show that the title in this series of papers is no longer appropriate, as none of them was found in the Austral-Antarctic region:
aequicornis (Stephensen, 1931: 198 fig. 59); between Faroes and Iceland, N-Atlantic.
anophthalmus Ledoyer, 1986: 965–66 (Madagascar, 335–450 m); Indian Ocean.
calcaratus (Sars, 1882: 92, t.4 sub Metopa calcaratus, 1895: 247, t. 85 sub Probolium. c.; 1992: 247 t. 85), Atlantic.
gregarius (Sars, 1882: 93; 1895: 245 pl. 84), Atlantic; = probably grandimanus (
? holmesi
pacificus (Holmes, 1908: 524), Pacific O, = ? tundus JL
schokalskii
sp. Shoemaker 1955: 128, Arctic, Point Barrow
zubovi
Species dubia.
Proboloides clypeatus (Stimpson, 1853: 51 sub Stenothoe clypeata), Atlantic.
Species incertae sedis.
? Metopoides typicamimus
? Proboloides holmesi
Species transferred to Metopoides.
Metopoides rotundus (Stebbing, 1917) (= ? Proboloides stephenseni Ruffo, 1949)
Metopoides typicus (Walker, 1906)
Key to 15 members of Metopoides species (including ?Metopoides typicamimus)| 1 | P7 basis posterodistally regularly rounded | 2 |
| – | P7 basis distally clearly narrowing | 13 |
| 2 | Gn2 male palm with incisions, excavations and teeth; palmar corner about 90° | 3 |
| – | Gn2 male palm smooth or serrated | 4 |
| 3 | Gn1 carpus < propodus; P5–7 merus distoposteriorly lengthened, clearly reaching more than half length of carpus, in P7 even more than full carpus length; U3 ramus art 1 = art 2, peduncle and ramus art 1 with 1 robust seta each | Metopoides pollex Krapp-Schickel, 2008 (3–4 mm) |
| – | Gn1 carpus > propodus; P5–7 merus posterodistal tip not reaching half length of carpus; U3 ramus art 1 > art 2, peduncle and ramus art 1 with many robust setae | ? Metopoides typicamimus Andres, 1995 (3–3, 5 mm) |
| 4 | Gn1 carpus twice as long as wide; propodus widening distad, with concave palm and upturned palmar corner | Metopoides clavatus Schellenberg (5.5–8 mm) |
| – | Gn1 carpus not as long; propodus not as above | 5 |
| 5 | P6 basis widened, but anterior and posterior margin parallel, not convex | 6 |
| – | P6 basis posterior margin convex, rounded as P7 | 7 |
| 6 | Gn1 short and wide, propodus and carpus l < 2b; Gn2 palm 1/3 of total length of propodus; U3 ramus art 1 subequal length of peduncle | Metopoides sarsii (Pfeffer) (2.8–6.5 mm?!) |
| – | Gn1 elongate, propodus and carpus l > 2b; Gn2 palm > 1/3 of total length of propodus; U3 ramus art 1 < length of peduncle | Metopoides lanceolatus Rauschert (3–4 mm) |
| 7 | Cx4 inferior margin distinctly excavated, concave | 8 |
| – | Cx4 inferior margin convex or only slightly excavate | 9 |
| 8 | Gn2 fem. propodus with parallel margins, palmar corner about 150°, width < half length of anterior margin | Metopoides cf. heterostylis (3 mm), Metopoides heterostylis Schellenberg (2.8–3.3 mm) |
| – | Gn2 fem. propodus widest at palmar corner, which is < 120°, width > half length of anterior margin | Metopoides latus Rauschert (2.8–3.4 mm) |
| 9 | Gn 1, 2 propodus with parallel margins, shape very similar; P7 merus twice as wide as carpus, reaching half length of carpus | Metopoides curvipes Schellenberg(juv. fem. 2 mm) |
| – | Gn2 propodus widening | 10 |
| 10 | Gn2 crenulated, in the middle of the long palm a semicircular excavation | Metopoides rotundus (Stebbing, 1917) , ?= Metopoides stephenseni (Ruffo, 1949) (3.5 mm) |
| – | Gn2 palm serrated or smooth | 11 |
| 11 | Gn2 palmar corner well defined by short and acute tooth as well as shallow excavation;P6, 7 merus distoposterior tip not reaching half length of carpus | Metopoides typicus (Walker, 1906) (3–3.5 mm) |
| – | Gn2 palmar corner smooth | 12 |
| 12 | U3 peduncle = ramus, strongly spinose; P7 basis posteriorly semicircularly rounded | Metopoides bellansantiniae (Bushueva) (3 mm) |
| – | U3 peduncle < ramus, naked; P7 basis oval | Metopoides magellanicus (Stebbing) (2.8–3 mm) |
| 13 | Gn2 propodus twice as long as wide; A1>A2 | 14 |
| – | Gn2 propodus clearly much more than twice as long as wide; A1< A2 | 15 |
| 14 | Gn1 length of propodus = carpus length; P7 basis proximally twice as wide as distally and about twice as long as the distal width | Metopoides longicornis Schellenberg (2–3 mm) |
| – | Gn1 length of propodus < carpus length; P7 basis proximally only a little wider than distally, about three times as long as the distal width | Metopoides angustus Rauschert (3.2 mm) |
| 15 | P6 basis elliptical, distally and proximally about the same width; P7 posterior margin regularly convex, but posterodistally no lobe | Metopoides ellipticus Schellenberg (4.5 mm) |
| – | P6, 7 basis trapezoid shaped, distally distinctly narrower than proximally | Metopoides leptomanus Rauschert (3.6–3.9 mm) |
| 1 | Gn2 male palm identical to propodus length; propodus longer than 2× width | 2 |
| – | Gn2 male palm not identical to propodus length; propodus shorter than 2× width | 3 |
| 2 | Gn2 male propodus with short serrated margin near dactylus-insertion, then rectangular corner, two humps along the remaining palmar margin; Gn2 female palm with similar serrated part near dactylus insertion, remaining palmar margin somewhat excavated | Proboloides gregarius (Sars, 1882) (5–6 mm, N-Atlantic, Arctic) |
| – | Gn2 male propodus with short serrated margin near dactylus insertion, defined by blunt corner, remaining palmar margin smooth; Gn2 female propodus rounded, smooth, palm = hind margin | Proboloides calcaratus (Sars, 1882) (5–6 mm, N-Atlantic) |
| 3 | Gn2 male palmar margin not defined by acute tooth | 4 |
| – | Gn2 male palmar margin defined by acute tooth | 5 |
| 4 | Gn2 propodus l:b about 3:2, palm about the length of remaining hind margin, corner rounded; Gn1 simple, triangular carpus < propodus | ? Proboloides holmesi Bousfield, 1973 (2, 5 mm, Pacific) |
| – | Gn2 propodus l:b about 2:1, palm not defined, dactylus reaching along 2/3 of palm which is unregularly serrated; Gn1 subchelate, elongate carpus with parallel margins, > propodus | Proboloides aequicornis (Stephensen, 1931) 5 mm, Faroes, N-Atlantic |
| 5 | Gn2 male, female palm longer than remaining hind margin of propodus; Gn1 propodus triangular, palmar corner about 120° | Proboloides schokalskii Gurjanova (5 mm, Kara Sea) |
| – | Gn2 male, female palm not longer than remaining hind margin of propodus | 6 |
| 6 | Gn2 male next to dactylus insertion strongly serrate; besides the palm-defining strong and acute tooth in the middle of the palm another one, followed by a small narrow incision; in male, female P7 basis oval | Proboloides pacificus (Holmes, 1908) (6 mm, Pacific) |
| – | Gn2 palm not divided by additional tooth, regularly serrated from dactylus-insertion to corner | 7 |
| 7 | Gn1 carpus clearly longer than propodus, more than 4× longer than wide |
Proboloides sp. (see “Proboloides nordmanni” in |
| – | Gn1 carpus subequal to propodus | 8 |
| 8 | Gn1 propodus l:b = 2; P6 basis with regularly rounded margin, merus reaching end of carpus | Proboloides anophthalmus Ledoyer, 1986 (2, 5 mm, Madagascar) |
| – | Gn1 propodus l> 2b; P6 with straight, parallel margins, merus not reaching end of carpus | Proboloides zubovi Gurjanova, 1951 (5 mm, Kara Sea) |
The habitus sketches added to the resulting tree in Fig. 14 (from above: Proboloides, a large Scaphodactylus, Torometopa with A1> A2, Torometopa with A1 < A2, small Scaphodactylus and Metopoides) may give an idea about the differences in the body shapes in this basic group of stenothoids: e.g. members of Proboloides have a disto-posteriorly lengthened and widened merus on the last three peraeopods, which is much less the case in all other groups. Ed Bousfield (in litteris) opines that this must have an important hydrodynamic function, which could well be imagined. Also the relative length and width of Cx 4 (in Scaphodactylus gigantocheirus strikingly small) or the relation of the antennae (in the group near Torometopa antarctica and Torometopa crenatipalmata with the second one always being longer and stronger) could tell us something about the swimming (or even digging?) ability, if we would know more about their life style. However, it seems probable that Proboloides members are mainly free-living and do not live in association with, or at least not inside of, other animals; they ought therefore to be good swimmers and have a rather strong sexual dimorphism.
The remaining species in Fig. 14., not belonging to the genus Proboloides, will be treated in the following and final part.
As already written in part 1 (