





(C) 2011 Nesrine Akkari. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Surface structures have rarely been the subject of a comprehensive study in Polydesmida despite their tremendous variety within this order. A number of these peripheral structures are here studied in most families of the suborders Polydesmidea and Dalodesmidea (sensu
surfacemicrosculpture, prozonite, limbus, cuticular micro-scutes, ozopore, metazonital outgrowth, taxonomy
Whereas the gonopods have hitherto been acknowledged to be the most reliable source of characters for millipede identification, the details of external morphology have in most cases remained under-prospected. This is also true for the order Polydesmida although it is by far the most diverse millipede order in terms of non-gonopodal morphology. The taxonomy of several polydesmidan families, notably Fuhrmannodesmidae and Pyrgodesmidae, is in a deplorable state, and new taxonomic characters are badly needed.
Scanning electron microscopy has, in many contemporary works, significantly helped to illustrate fine surface structures in millipedes.
The limbus, or posterior margin of the metazonites, was investigated by
Some polydesmidans of the families Cryptodesmidae, Haplodesmidae, and Pyrgodesmidae are earth-incrusted, i.e., adult specimens bear a coat of dirt.
During the study of a new pyrgodesmid species from Tunisia using scanning electron microscopy (
The studied species and the states of the three main characters examined.
| Species | Knobs on the posterior surface of the prozonites | Limbus | Intercalary micro-scutes <br/> on metazonites | |
|---|---|---|---|---|
| Suborder Polydesmidea | ||||
| Ammodesmidae | Elassystremma sp. | + | lobes and spikes | ‒ |
| Cryptodesmidae | Aporodesmus sp. | ‒ | jagged lobes and spikes | ‒ |
| Elythesmus enghoffi | + | jagged lobes and spikes | _ | |
| Cyrtodesmidae | cyrtodesmid gen. sp. | + | lobes and spikes | ‒ |
| Fuhrmannodesmidae | Fuhrmannodesmus lividus | – | reduced | + |
| fuhrmannodesmid sp. | ‒ | reduced | + | |
| Gyrophallus sp. | ‒ | reduced | + | |
| Haplodesmidae | Prosopodesmus jacobsoni | + | lobes | ‒ |
| Macrosternodesmidae | Ophiodesmus albonanus | ‒ | reduced | + |
| Nearctodesmidae | nearctodesmid sp. | ‒ | reduced | + |
| Oniscodesmidae | Amphitomeus attemsi | + | lobes and spikes | ‒ |
| Opisotretidae | Corypholophus sp. | ‒ | reduced | ?+ |
| Solaenaulus butteli | – | reduced | + | |
| Polydesmidae | Brachydesmus superus | ‒ | ramified spikes | + |
| Propolydesmus laevidentatus | – | ramified spikes | + | |
| Pyrgodesmidae | Cryptocorypha ornata | + | lobes | ‒ |
| Cynedesmus sp. | + | lobes | – | |
| Rharodesmus tabarkensis | + | lobes | – | |
| Tonodesmus sp. | + | lobes | – | |
| Trichopolydesmidae | Napocodesmus endogeus | ‒ | reduced | + |
| trichopolydesmid sp. | – | reduced | + | |
| Suborder Dalodesmidea | ||||
| Dalodesmidae | Icosidesmus sp. | ‒ | ramified spikes | + |
Our study was mainly focused on three sets of characters: a) surface sculpture of the prozonite (anterior, cylindrical part of body ring); b) the limbus (posterior margin of body rings); c) intercalary micro-scutes on the surface of the metazonite. A few additional structures such as the cuticular outgrowths in earth-incrusted species and the ozopores are briefly presented below and compared. Moreover, recent literature accounts of 29 species from relevant families (see Table 2), including SEM illustrations or descriptions were checked and compared with our results.
Literature records
| Species | Reference | |
|---|---|---|
| Suborder Polydesmidea | ||
| Ammodesmidae | Elassystremma laeve Vandespeigel and Golovatch, 2003. Elassystremma prolaeve VandenSpeigel and Golovatch, 2003 | VandenSpeigel and |
| Cryptodesmidae | Aporodesmus gabonicus (Lucas, 1858) |
|
| Tarmadesmus azucarensis Kraus, 1959 | ||
| Fuhrmannodesmidae | Fuhrmannodesmus carli Kraus, 1955 |
|
| Salvadoria sagittalis Kraus, 1954 | ||
| Cutervodesmus similis Kraus, 1959 | ||
| Haplodesmidae | Agathodesmus steeli Silvestri, 1910 |
|
| Cylindrodesmus hirsutus Pocock, 1889 |
|
|
| Eutrichodesmus armatocaudatus Golovatch et al., 2009 |
|
|
| Eutrichodesmus basalis Golovatch et al., 2009 | ||
| Eutrichodesmus communicans Golovatch et al., 2009 | ||
| Eutrichodesmus inciues Golovatch et al., 2009 | ||
| Eutrichodesmus similis Golovatch et al., 2009 | ||
| Oniscodesmidae | Oncodesmoides rectus Kraus, 1954 |
|
| Opisotretidae | Opisotretus kraepelini (Attems, 1907) |
|
| Polydesmidae | Polydesmus complanatus (Linneaus, 1871) |
|
| Pyrgodesmidae | Lobiferodesmus vanuatu Golovatch, et al., 2008 |
|
| Poratia digitata (Porat, 1889) |
|
|
| Muyudesmus obliteratus Kraus, 1960 | ||
|
Cryptocorypha hoffmani |
|
|
| Myrmecodesmus hastatus (Schubart, 1945) |
|
|
| Monachodesmus albus Kraus, 1958 |
|
|
| Suborder Dalodesmidea | ||
| Dalodesmidae |
Ginglymodesmus tasmanianus |
|
| Not assigned to any family | Asphalidesmus bellendenkerensis Mesibov, 2011 |
|
| Asphalidesmus golovatchi Mesibov, 2009 | ||
| Noteremus infimus Mesibov, 2009 |
|
|
| Noteremus summus Mesibov, 2009 | ||
| Procophorella innupta Mesibov, 2003 |
|
|
The studied material is preserved in 70 % ethanol and deposited in the Natural History Museum of Denmark (Zoological Museum, University of Copenhagen, ZMUC). Earth-incrusted specimens were cleaned for scanning electron microscopy by soaking in a solution of commercial detergent (©Biotex) and/or by ultrasound then air dried. SEM pictures were made with a JEOL JSM-6335F scanning electron microscope, then processed and assembled with Adobe Photoshop CS5 software.
When not otherwise indicated, we have followed the classification proposed by
Suborder Polydesmidea Pocock, 1887
Ammodesmidae Cook, 1896
Elassystremma sp., Tanzania, Unzungwa Mts, Iringa Region, Uzungwa Scarp Forest Res., above Chita village, 1600–1650m, 8–13.ix.1984, pitfall Traps in Montane Rain Forest, N. Scharff leg. (ZMUC 00020487).
Cryptodesmidae Karsch, 1879
Aporodesmus sp., female, Cameroun, Northwest Province, Menchum Div. Near L. Oku forest, in litter, ca. 2150m, N6°12', E10°27', 7–13.ii.1992, C. Griswold, S. Larcher, N. Scharff and C. Wanzie leg. (ZMUC 00020478).
>Elythesmus enghoffi Hoffman, 1978, female, Tanzania, W. Usambara Mts, Mazumbai Forest Reserve, 19–29.ix.1992, M. Andersen leg. (ZMUC 00020477).
Cyrtodesmidae Cook, 1896
Cyrtodesmid sp., female, Colombia, Páramo de Sumapaz, soil under grasses, 3600m, 5.x.1978, H. Sturm leg. (ZMUC 00020494).
Fuhrmannodesmidae Brölemann, 1916
Fuhrmannodesmus lividus Carl, 1914, male, Colombia, Par de Monserrate, near Bogotá, 3250m, dead leaves of Espeletia grandifolia, 18.iv.1969, H. Sturm (ZMUC 00020483).
fuhrmannodesmid sp. (
Gyrophallus sp., female, Colombia, 1991, H. Sturm leg. (ZMUC 00020484).
Haplodesmidae Cook, 1895
Prosopodesmus jacobsoni Silvestri, 1910, female, Fiji Isl, Viti Levu Suva, in garden, 2–3.ix.1995, A van Hart leg. (ZMUC 00020476).
Macrosternodesmidae Brölemann, 1916
Ophiodesmus albonanus (Latzel, 1895), male, Denmark, NE Zaland, Copenhagen, Utterslev Mose, 22.iv.1973, H. Enghoff leg. (ZMUC 00020491).
Nearctodesmidae Chamberlin and Hoffman, 1950
nearctodesmid sp., male, Calif. Humboldt Co. Jolly Giant Canyon, 300–650m, Arcata Comm. For 13.i.1979, A.K. Johnson, R. M. Shelley leg. (ZMUC 00020482).
Oniscodesmidae de Saussure, 1860
Amphitomeus attemsi (Schubart, 1934), female, Dania: Nez UB47, Copenhagen, Botanical garden, væksthus, 16.iv.1986, H. Enghoff and Z. Korsòs leg. (ZMUC 0002046).
Opisotretidae Hoffman, 1980
Corypholophus sp., female, Thailand, Chieng Mai Province, Doi Inthanon N. P., Mae Ya, 6–700m (ZMUC 00020479).
Solaenaulus butteli (Carl, 1922), male, Fiji Isl. Viti Levu Suva, in garden, 2–3.ix.1995, A van Harten leg. (ZMUC 00020480).
Polydesmidae Leach, 1815
Brachydesmus superus (Latzel, 1884), female, Tunisia, NW, Jendouba Governorate, 9km of Hammam Bourguiba (West of Aïn Draham), N36°48.046, E08°39.544, 379m, Pine Forest, close to river, under stones, logs and leaf litter, 22.iii.2008, P. Stoev and N. Akkari leg. (ZMUC 00020496).
Propolydesmus laevidentatus (Loksa, 1967), male, Madeira, Faja da Nogueria, N. Side, ca. 800m. a.s.l. Laurisilva with Ocoteas, 20.xi.1980, H. Enghoff and O. Martin leg. (ZMUC 00020475).
Pyrgodesmidae Cook, 1895
Cryptocorypha ornata (Attems, 1938), unsexed fragment, Hawaii Isl., Kauai, in a grotto, moss and Adiantum, 20.x.1962, M. Hammer leg. (ZMUC 00020493)
Cynedesmus sp., female, La Gomera, Valle Gran Rey, litter, in banana plantation, 10m, 2.xii.1987, A. Fjellberg leg. (ZMUC 00020485).
Rharodesmus tabarkensis Akkari and Enghoff, 2011, male, Tunisia, NW, Jendouba Governorate, Tabarka, N36°58'10.5", E8°45'35.6", alt. < 40m, coastal slope below the Genoese fort, under stones, 9.iii.2009, N. Akkari and H. Enghoff leg. (ZMUC 00020532).
Tonodesmus sp., male, Spain, Almeria Sima terminal, T. M. Eidijo, 28.v.2000, M. Piquer and J.G. Pedro leg. (ZMUC-00020495).
Trichopolydesmidae Verhoeff, 1910
Napocodesmus endogeus Ceuca, 1974, female, Moldavian SSR, Tiraspol, deep in orchard soil, 1985, S. Golovatch leg. (ZMUC 00020481).
trichopolydesmid sp. Slovakia, Slovak Karst, Ardouská Cave A-04-47, 5.x.2004, A. Mock leg. (ZMUC 00020490).
Suborder Dalodesmidea Hoffman, 1980
Dalodesmidae Cook, 1896
Icosidesmus sp., male, New Zealand, South Isl, Christchurch Banks Peninsula, Hinewai Reserve, Big Kanuka Trail, 3 iii.2010, S43°48'38.0", E173°01'15.6", 508m, sifted leaf litter and mosses, N. Scharff and G. Hormiga leg. (ZMUC 00020488).
Results Fine sculpture of the prozoniteFigs 1–9
The prozonite of the studied species is divided into two main parts separated by a transverse ridge. While the anterior part is rather uniform, showing a scaly aspect, the posterior surface displays varied patterns within the studied families.

Fine sculpture of the prozonite in the families Ammodesmidae, Cryptodesmidae, Cyrtodesmidae, Haplodesmidae, Oniscodesmidae and Pyrgodesmidae 1 Elassystremma sp. 2 Elythesmus enghoffi, 3 cyrtodesmid sp. 4 Prosopodesmus jacobsoni 5 Amphitomeus attemsi 6 Rharodesmus tabarkensis. Abbreviations: a anterior part of the prozonite, b posterior part of the prozonite, r ridge, s spherical knobs.
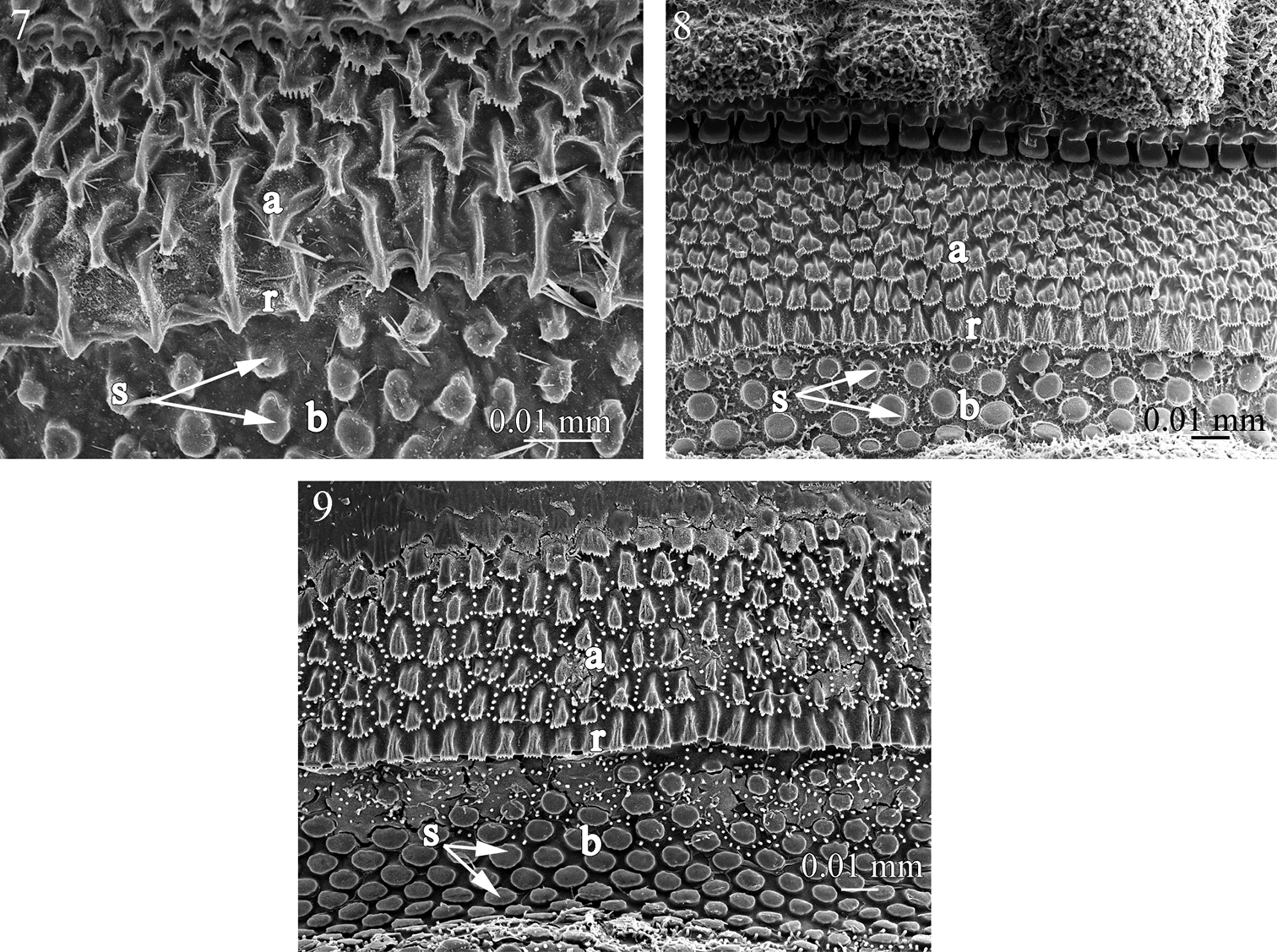
Fine sculpture of the prozonite in pyrgodesmid species. 7 Tonodesmus sp. 8 Cynedesmus sp. 9 Cryptocorypha ornata. Abbreviations: a anterior part of the prozonite, b posterior part of the prozonite, r ridge, s spherical knobs.
In the examined species of Ammodesmidae, Cryptodesmidae, Cyrtodesmidae, Haplodesmidae, Oniscodesmidae and Pyrgodesmidae the anterior part of the prozonite (a) displays a covering of small scaly units, juxtaposed in series of irregular transverse rows, becoming elongated in the posteriormost row to form a transverse ridge (r) marking the border of this part (Figs 1–9). In front of the ridge, the general aspect is quite uniform in all the above cited families, with lozenge-shaped units. These units could sometimes be star-like, furrowed, and marginally jagged (cyrtodesmid sp.) (Fig. 3). In Amphitomeus attemsi (Oniscodesmidae), the units are more elongated and strongly prominent, interconnected with parallel cuticular ridges (Fig. 5). On the other hand, the microsculpture of the anterior part of the prozonite in Prosopodesmus jacobsoni (Haplodesmidae) takes the shape of hollow chambers separated by walls of “microvilli-like” structures (Fig. 4). The posterior surface of the prozonite (b) is, in all examined species of these six families, characterized by a regular covering of sub-spherical knobs (s) placed on a smooth to microgranulated background. The cover of knobs is regularly dense in most cases (Figs 1, 3–9)though fairly dispersed in Elythesmus enghoffi (Cryptodesmidae) (Fig. 2). These knobs are generally uniformly smooth (Figs 1–3, 6–9) but sometimes exhibit special configurations: lobed in Prosopodesmus jacobsoni and spiky in Amphitomeus attemsi (Figs 4, 5).
The examined species of the families Fuhrmannodesmidae, Polydesmidae, Dalodesmidae, Macrosternodesmidae, Nearctodesmidae, Opisotretidae and Trichopolydesmidae show an anterior surface of the prozonite with polygonal units serrated marginally. However, these units are much less conspicuous and prominent than in the species of the first set of families (Figs 10–17). The posterior border of the anterior part of the prozonite is similarly marked by a transverse ridge but its units are nevertheless only slightly modified. The posterior surface of the prozonite remarkably differs by the complete absence of the sub-spherical knobs described above; instead the surface is smooth to scaly (Figs 10–17).
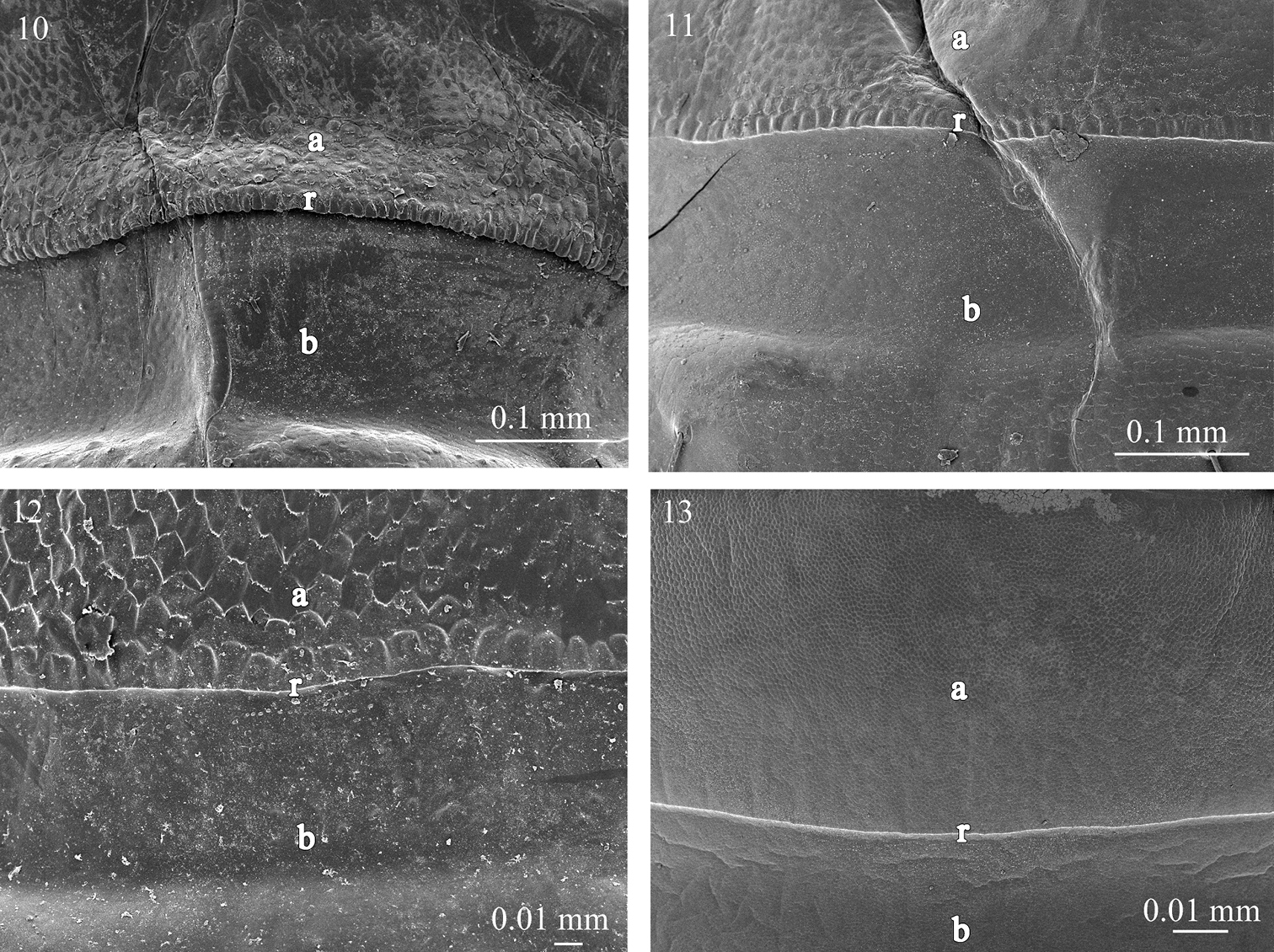
Fine sculpture of the prozonite in 10 Fuhrmannodesmus lividus (Fuhrmannodesmidae) 11 Gyrophallus sp. (Fuhrmannodesmidae) 12 Propolydesmus laevidentatus (Polydesmidae) 13 Icosidesmus sp. (Dalodesmidae). Abbreviations: a anterior part of the prozonite, b posterior part of the prozonite, r ridge.
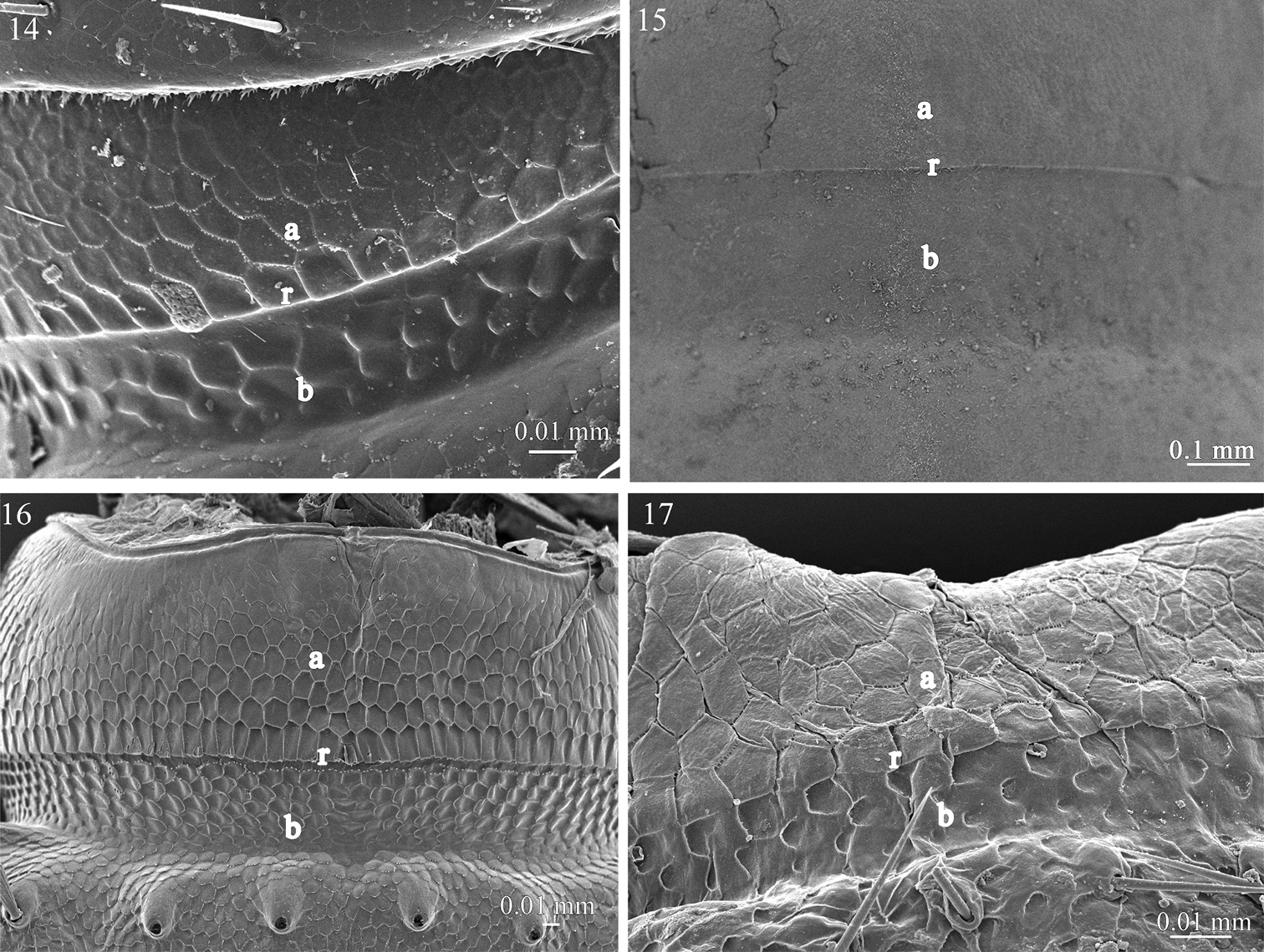
Fine sculpture of the prozonite in 14 Ophiodesmus albonanus (Macrosternodesmidae) 15 nearctodesmid sp. (Nearctodesmidae) 16 Solaenaulus butteli (Opisotretidae) 17 Napocodesmus endogeus (Trichopolydesmidae). Abbreviations: a anterior part of the prozonite, b posterior part of the prozonite, r ridge.
Figs 18–34

The structure of the limbus in 18 Prosopodesmus jacobsoni (Haplodesmidae) 19 Rharodesmus tabarkensis (Pyrgodesmidae) 20 Cynedesmus sp. (Pyrgodesmidae) 21 Tonodesmus sp. (Pyrgodesmidae) 22 Cryptocorypha ornata (Pyrgodesmidae). Abbreviations: L limbus, lo lobe, pa palette-like lobe, sp spike, t tooth-like lobe.
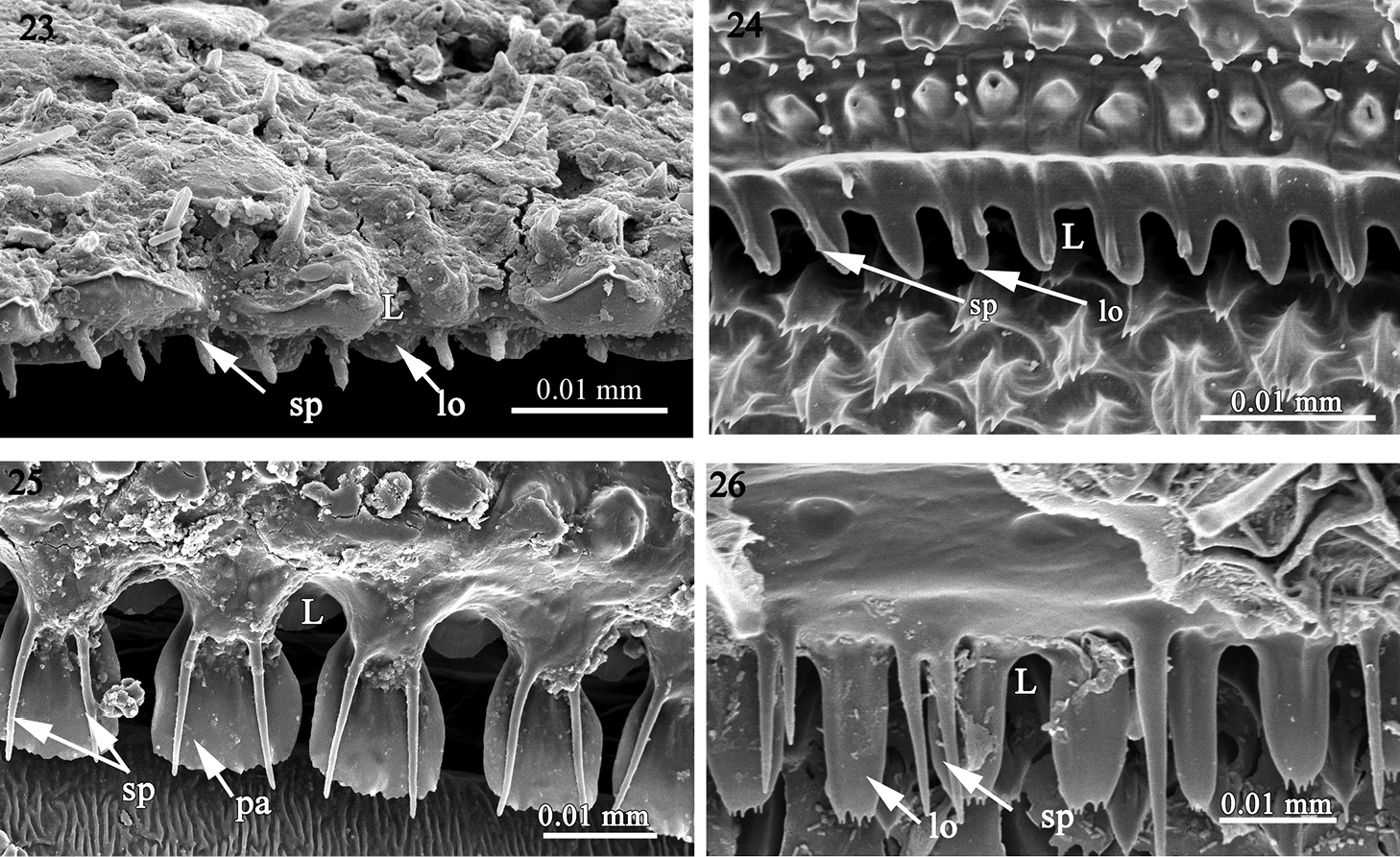
The structure of the limbus in 23 Amphitomeus attemsi (Oniscodesmidae) 24 cyrtodesmid sp. (Cyrtodesmidae) 25 Elassystremma sp. (Ammodesmidae) 26 Elythesmus enghoffi (Cryptodesmidae). Abbreviations: L limbus, lo lobe, sp spike, pa: palette-like lobe.
The limbus (L) displays three major patterns of shapes in the studied families:
1) A regular set of rounded lobes (lo) placed in one row as in Prosopodesmus jacobsoni(Haplodesmidae), Rharodesmus tabarkensis(Pyrgodesmidae) (Figs 18, 19) or two superposed rows of 'palette-shaped' lobes (pa) separated by spikes (sp) as in Cynesdesmus sp. (Pyrgodesmidae) (Fig. 20) although in some species of the latter family (e.g. Tonodesmus sp. and Cryptocorypha ornata) the lobes are more tooth-like (t) (Figs 21, 22). The lobes are also surmounted by fine spikes as in Amphitomeus attemsi (Oniscodesmidae), cyrtodesmid sp. (Figs 23, 24), Elassystremma sp. (Ammodesmidae) in which the lobes are moreover stocky or ‘palette-like’(pa) and serrated (Fig. 25) and in Elythesmus enghoffi(Cryptodesmidae) where the spikes are more elongate and inserted between the jagged lobes (Fig. 26).
2) A series of ramified spikes (rs) in Icosidesmus sp. (Dalodesmidae) (Fig. 27) or “icicles” in Ophiodesmus albonanus (Macrosternodesmidae), Propolydesmus laevidentatus and Brachydesmus superus (Polydesmidae) (Figs 28–30).

The structure of the limbus in 27 Icosidesmus sp. (Dalodesmidae), 28 Ophiodesmus albonanus (Macrosternodesmidae) 29 Propolydesmus laevidentatus (Polydesmidae) 30 Brachydesmus superus (Polydesmidae). Abbreviations: L limbus, rs ramified spike.
3) Reduced. In Solaenaulus butteli (Opisotretidae), Napocodesmus endogeus (Trichopolydesmidae) and Fuhrmannodesmus lividus (Fuhrmannodesmidae) the limbus is hardly developed, taking the shape of a regular edge bearing a few scattered bulges (bu) which could be isolated or grouped, e.g. sets of three bulges in furhmannodesmid sp. (Figs 31–34).
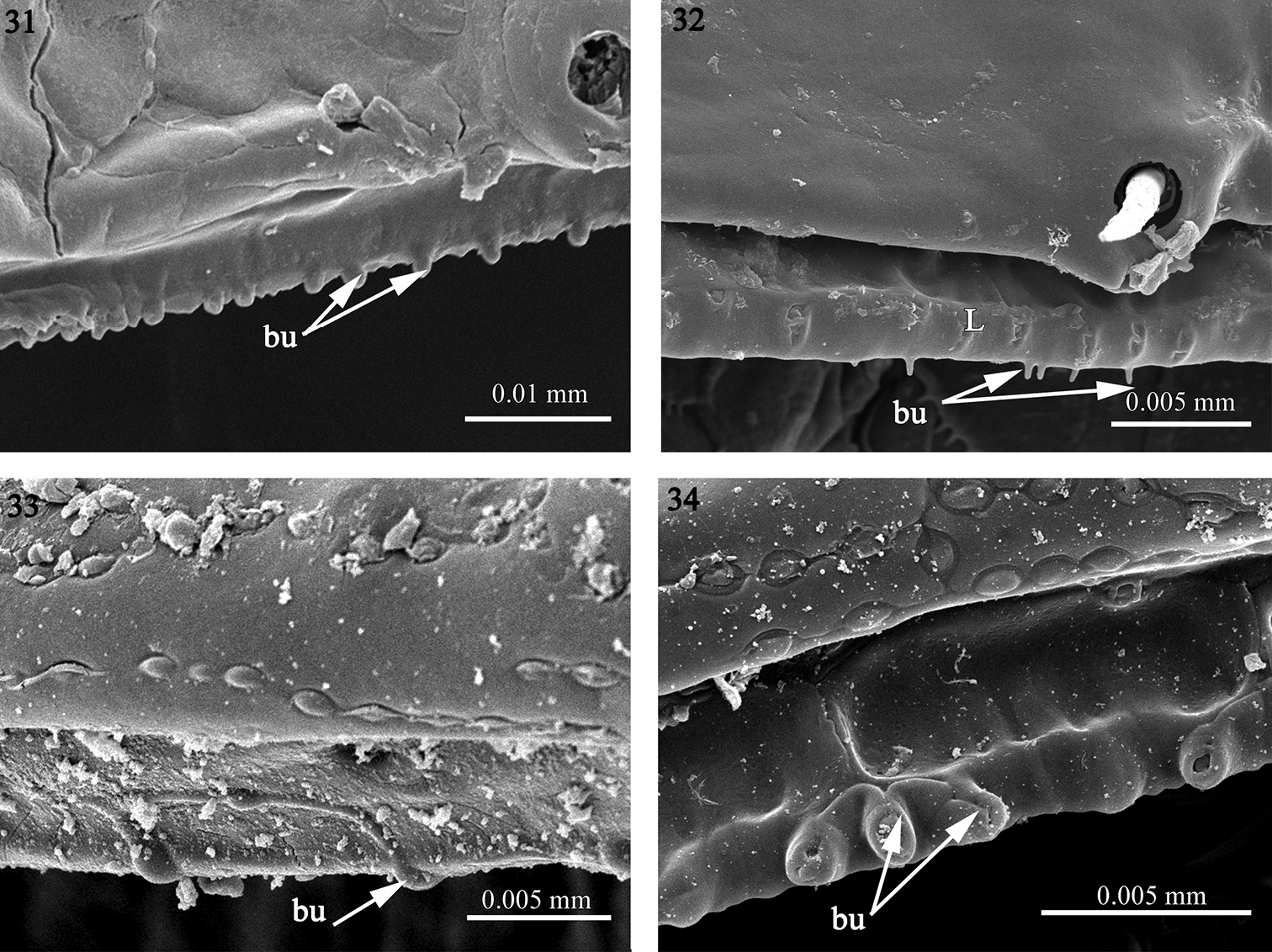
The structure of the limbus 31 Solaenaulus butteli (Opisotretidae) 32 Napocodesmus endogeus (Trichopolydesmidae), 33 Fuhrmannodesmus lividus (Fuhrmannodesmidae) 34 fuhrmannodesmid sp. (Fuhrmannodesmidae). Abbreviations: bu bulges, L limbus.
Figs 35–41

Intercalary micro-scutes on the metazonitesof 35 Fuhrmannodesmus lividus (Fuhrmannodesmidae) 36 Propolydesmus laevidentatus (Polydesmidae) 37 Icosidesmus sp. (Dalodesmidae) 38 Ophiodesmus albonanus (Macrosternodesmidae) 39 nearctodesmid sp. (Nearctodesmidae) 40 Solaenaulus butteli (Opisotretidae) 41 Napocodesmus endogeus (Trichopolydesmidae). Arrows point to the micro-scutes.
The studied species of the families Fuhrmannodesmidae, Polydesmidae, Dalodesmidae, Macrosternodesmidae, Nearctodesmidae, Opisotretidae and Trichopolydesmidae (cf. Table 1) present a peculiar structure on the metazonital surface: between the normal polygonal cuticular scutes which cover the metazonital surface there are rows of small ovoid “intercalary scutes”. The placement of these structures is unlikely to be accidental or indicating any kind of bacterial infection as they seem well arranged in a regular pattern, appearing like spaced nodes or pearls aligned on strings crossing the surface of the metazonites (Figs 35–41). These structures have never been documented. However, they are visible on an illustration in
Figs 42–49

Cuticular outgrowths 42 Rharodesmus tabarkensis (Pyrgodesmidae) 43 Elassystremma sp. (Ammodesmidae).
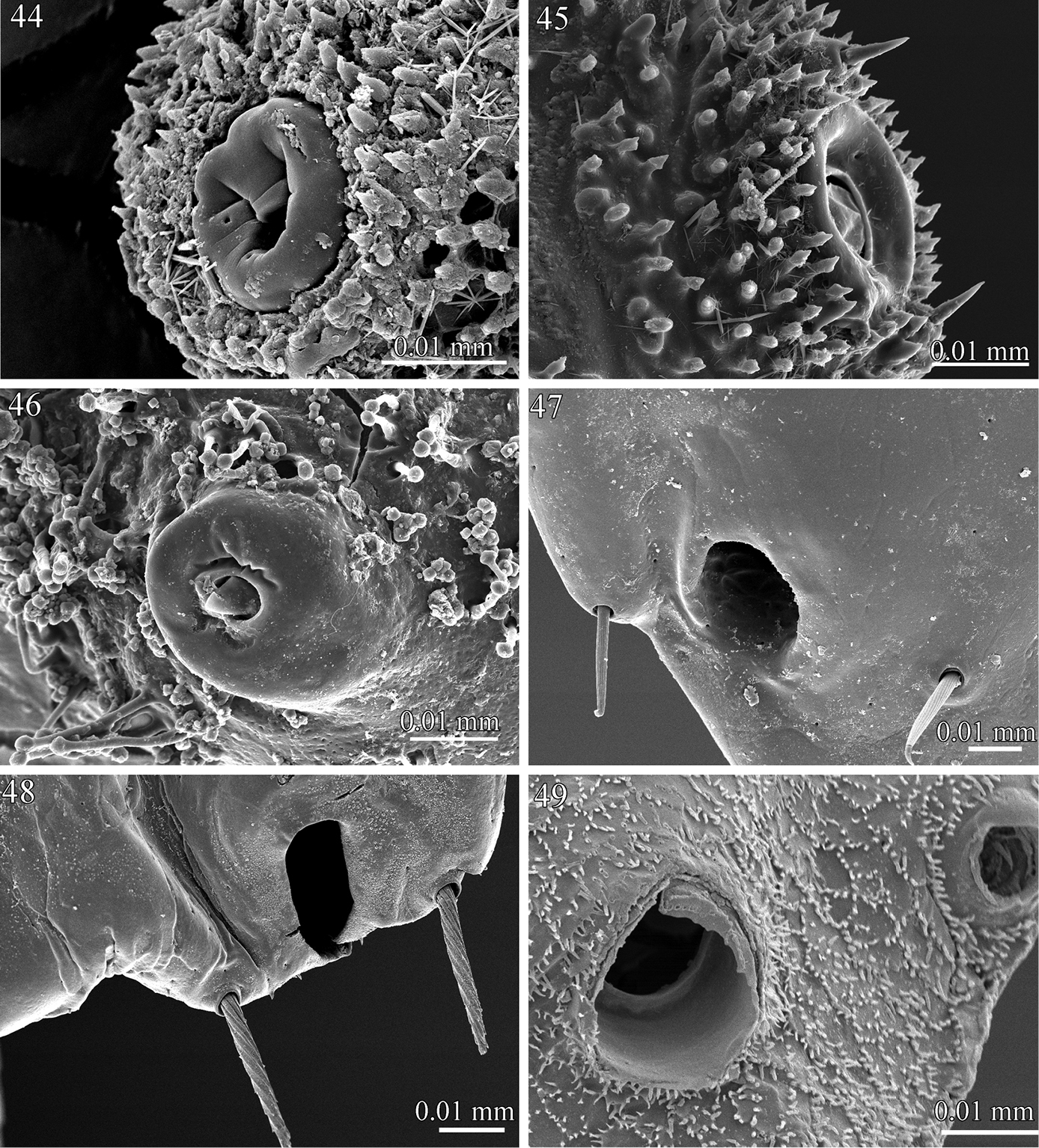
Ozopores 44 Rharodesmus tabarkensis (Pyrgodesmidae) 45 Tonodesmus sp. (Pyrgodesmidae) 46 Elassystremma sp. (Ammodesmidae) 47 Propolydesmus laevidentatus (Polydesmidae) 48 Gyrophallus sp. (Fuhrmannodesmidae) 49 Corypholophus sp. (Opisotretidae).
The surface structure of most earth-incrusted species is characterized by the presence of papilla-like cuticular outgrowths which are particularly abundant in Ammodesmidae, Cyrtodesmidae and Pyrgodesmidae. These papillae are boletiform (mushroom-shaped) and are presumed to keep the cuticular secretions and adhering soil particles in place. Displaying variation in length, shape of the apex, and density on the surface, the papillae are short, with a rounded apex in Rharodesmus tabarkensis (Fig. 42) and Cynedesmus sp., elongate and slender in Elassystremma sp. (Fig. 43) and in cyrtodesmid sp. The same structures have been mentioned in previous works, generally quoted as “papillis” (
The ozopores (defense gland openings) display a notable variation within the examined families. In Prosopodesmus jacobsoni (Haplodesmidae), Tonodesmus sp., Rharodesmus tabarkensis, Cynedesmus sp. (Pyrgodesmidae) and Elassystremma sp. (Ammodesmidae), they open on small rounded discs, situated on the surface of the paratergites or on porosteles. The discs are of variable thickness and diameter, bear an apparent internal closing mechanism and are externally bordered with several whorls of papillae (Figs 44–46). On the other hand, the ozopores appear as simple sub-circular openings on the surface of the paratergites in Icosidesmus sp. (Dalodesmidae), Propolydesmus laevidentatus (Polydesmidae), Fuhrmannodesmus lividus and Gyrophallus sp. (Fuhrmannodesmidae), and Corypholophus sp. (Opisotretidae) (Figs 47–49).
DiscussionPolydesmida is the most speciose millipede order, and despite the fact that it has remained quite stable in terms of number of families (
Taken together, the fine sculpture of the prozonite, the structure of the limbus and the presence/absence of metazonital micro-scutes indicate two main groupings of families within the suborders Polydesmidea and Dalodesmidea. The first group (A) comprises the families Ammodesmidae, Cryptodesmidae, Cyrtodesmidae, Haplodesmidae, Oniscodesmidae and Pyrgodesmidae. These six families have a cover of sub-spherical knobs on the posterior surface of the prozonites (Figs 1–9) and a toothed limbus constituted of a series of lobes which may or not be surmounted by spikes (Figs 18–26).
The second group (B) encompasses the families Fuhrmannodesmidae, Polydesmidae, Dalodesmidae Macrosternodesmidae, Nearctodesmidae, Opisotretidae and Trichopolydesmidae, and is characterized by 1) an absence of knobs on the posterior surface of the prozonite which is smooth to scaly (Figs 10–17), 2) an absence of lobes on the limbus which has instead a jagged margin or spikes with varied complexity (Figs 27–34), and 3) intercalary micro-scutes on the surface of the metazonites (Figs 35–41), absent in the first set of families.
An assessment of several recent species descriptions and SEM illustrations shows in most cases similar structural arrangements of the prozonites. The presence of sub-spherical knobs on the posterior surface of the prozonite has been verified in a few additional genera and species of Pyrgodesmidae, such as Lobiferodesmus vanuatu (
The fine structure of the prozonite in both species of the Tasmanian dalodesmid genus Noteremus, Noteremus summus and Noteremus infimus (
Although assigned to Dalodesmidea (
Several literature records clearly support the patterns described above for the limbus. For example, the limbus shows a series of lobes with or without additional spikes in the pyrgodesmid species Lobiferodesmus vanuatu (
Though we are aware that the present data alone do not warrant a strict cladistic analysis, we have compared our findings (Fig. 51) with the only existing phylogenetic work on the order Polydesmida (
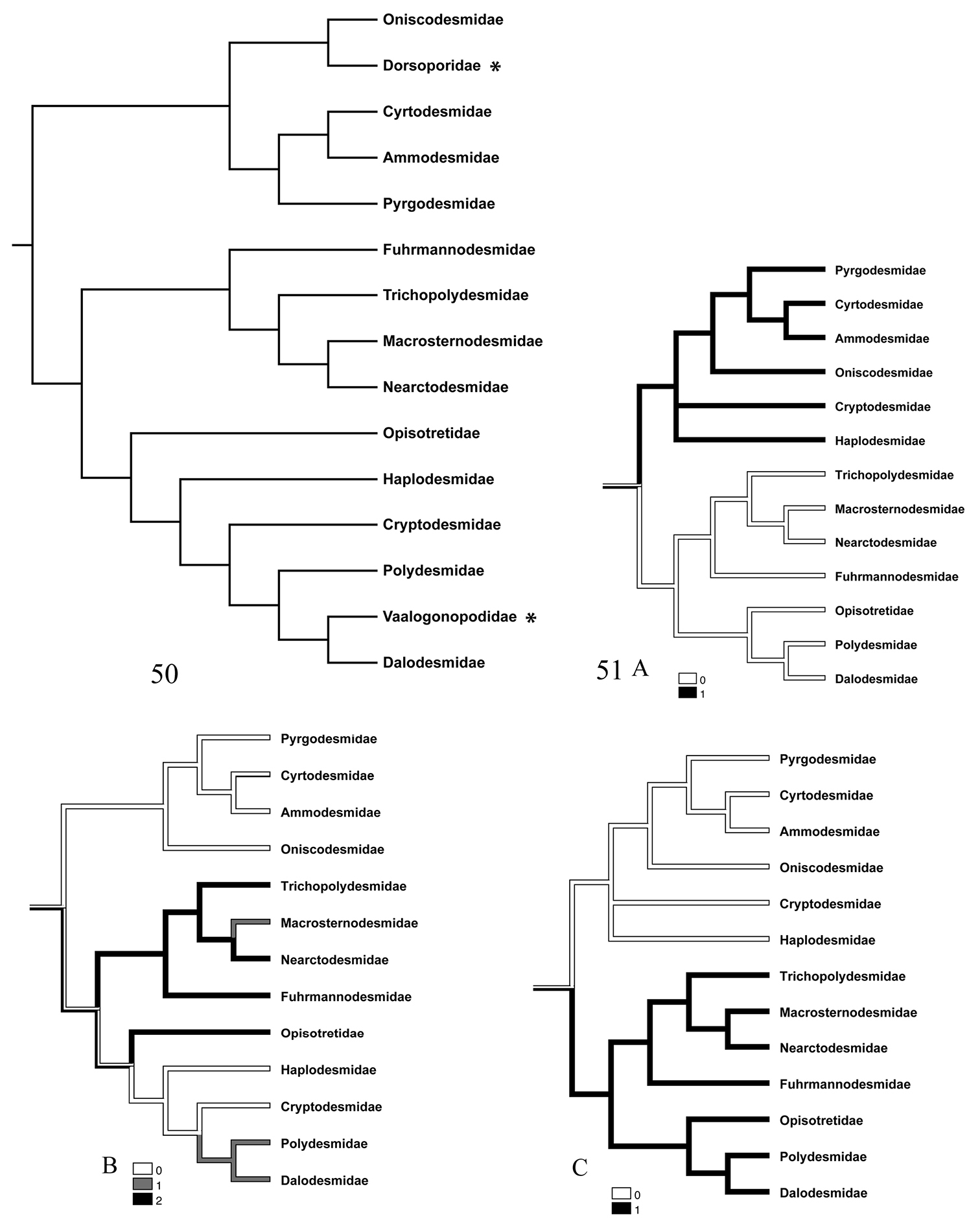
50 Family-level cladogram of suborders Polydesmidea + Dalodesmidea according to
The Cryptodesmidae studied by us alos present a complication as Elythesmus has knobs on the posterior part of the prozonites (Fig. 2) and a lobed limbus (Fig. 26) as in families group A, whereas Aporodesmus has a posterior prozonite surface free of knobs (Fig. 52) and dentate leaf-shaped (le) elements and spikes (sp) on the limbus (Fig. 53).

Prozonite and limbus in Aporodesmus sp. (Cryptodesmidae) 52 Prozonite fine sculpture 53 The limbus. Abbreviations: a anterior part of the prozonite, b posterior part of the prozonite, L limbus, le dentate leaf-shaped element of the limbus, r ridge, sp spike.
As stated above, the second set of families (B) have no lobes on the limbus. Instead, ramified spikes and “icicles” (Figs 27–30) can be observed in the families Dalodesmidae, Macrosternodesmidae and Polydesmidae, which clearly separates them from the Fuhrmannodesmidae, Trichopolydesmidae, Nearctodesmidae and Opisotretidae in which the limbus is hardly developed and bears but few scattered bulges (Figs 31–34). However, this separation fits neither with Hoffman’s (1980) classification in which the Macrosternodesmidae, Nearctodesmidae, Trichopolydesmidae and Fuhrmannodesmidae form the superfamily Trichopolydesmoidea (see also
In the present work, we do not pretend to offer a new subordinal classification of Polydesmida, or to solve any of the taxonomic problems related to families – a colossal task which definitely will require a lot more sampling and character scoring, including the gonopod structures which haven’t been considered here. However, documenting these remarkable surface structures and trying to compare them between the different families will perhaps contribute to bringing new insights, leading towards a better understanding of polydesmidean and dalodesmidean relationships.
We are grateful to Bill Shear and Robert Mesibov for sharing useful information. Many thanks are also extended to Jadranka Rota, who kindly helped with the diagrams, and two anonymous reviewers for their constructive comments.
List of characters and tentative character states
(1) Knobs on posterior surface of the prozonites: (0) absent; (1) present
(2) Shape of the limbus: (0) with one or two rows lobes additionally bearing simple spikes; (1) with ramified spikes and icicles; (2) reduced
(3) Rows of small intercalary scutes on metazonites: (0) absent; (1): present
Only the species studied by us have been coded.
| Char. no. | 1 | 2 | 3 | ||
|---|---|---|---|---|---|
| Suborder Polydesmidea Pocock, 1887 | |||||
| Ammodesmidae Cook, 1896 | Elassystremma sp. | 1 | 0 | 0 | |
| Cryptodesmidae Karsch, 1879 | Elythesmus enghoffi Hoffman, 1978 | 1 | 0 | 0 | |
| Aporodesmus sp. | 0 | 0 | 0 | ||
| Cyrtodesmidae Cook, 1896 | cyrtodesmid sp. | 1 | 0 | 0 | |
| Fuhrmannodesmidae Brölemann, 1916 | Fuhrmannodesmus lividus Carl, 1914 | 0 | 2 | 1 | |
| fuhrmannodesmid sp. | 0 | 2 | 1 | ||
| Gyrophallus sp. | 0 | 2 | 1 | ||
| Haplodesmidae Cook, 1895 | Prosopodesmus jacobsoni Silvestri, 1910 | 1 | 0 | 0 | |
| Macrosternodesmidae Brölemann, 1916 | Ophiodesmus albonanus (Latzel, 1895) | 0 | 1 | 1 | |
| Nearctodesmidae Chamberlin and Hoffman, 1950 | nearctodesmid sp. | 0 | 2 | 1 | |
| Oniscodesmidae de Saussure, 1860 | Amphitomeus attemsi (Schubart, 1934) | 1 | 0 | 0 | |
| Opisotretidae Hoffman, 1980 | Corypholophus sp. | 0 | 2 | ?+ | |
| Solaenaulus butteli (Carl, 1922) | 0 | 2 | + | ||
| Polydesmidae Leach, 1815 | Brachydesmus superus (Latzel, 1884) | 0 | 1 | 1 | |
| Propolydesmus laevidentatus (Loksa, 1967) | 0 | 1 | 1 | ||
| Pyrgodesmidae Cook, 1895 | Rharodesmus tabarkensis Akkari and Enghoff, 2011 | 1 | 0 | 0 | |
| Tonodesmus sp. | 1 | 0 | 0 | ||
| Cynedesmus sp. | 1 | 0 | 0 | ||
| Cryptocorypha ornata (Attems, 1938) | 1 | 0 | 0 | ||
| Trichopolydesmidae Verhoeff, 1910 | trichopolydesmid. sp. | 0 | 2 | 1 | |
| Napocodesmus endogeus Ceuca, 1974 | 0 | 2 | 1 | ||
| Suborder Dalodesmidea Hoffman, 1980 | |||||
| Dalodesmidae Cook, 1896 | Icosidesmus sp. | 0 | 1 | 1 | |
| Char. no.→ | 1 | 2 | 3 |
|---|---|---|---|
| Ammodesmidae | 1 | 0 | 0 |
| Cryptodesmidae | 1 | 0 | 0 |
| Cyrtodesmidae | 1 | 0 | 0 |
| Fuhrmannodesmidae | 0 | 2 | 1 |
| Haplodesmidae | 1 | 0 | 0 |
| Macrosternodesmidae | 0 | 1 | 1 |
| Nearctodesmidae | 0 | 2 | 1 |
| Oniscodesmidae | 1 | 0 | 0 |
| Opisotretidae | 0 | 2 | 1 |
| Polydesmidae | 0 | 1 | 1 |
| Pyrgodesmidae | 1 | 0 | 0 |
| Trichopolydesmidae | 0 | 2 | 1 |
| Dalodesmidae | 0 | 1 | 1 |