





(C) 2011 Anh D. Nguyen. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
The East Asian millipede genus Riukiupeltis Verhoeff, 1939 is revised, and is restricted to a single species, Riukiupeltis jamashinai Verhoeff, 1939. Examination of the type specimens and freshly collected material from the Ryukyu Archipelago and Vietnam show that both subsequently allocated species, Riukiupeltis uenoi Murakami, 1975, and Riukiupeltis falcatus (originally Haplogonosoma falcatum Attems, 1953, reallocated by
Revision, Riukiupeltis, Paradoxosomatidae, Polydesmida, Vietnam, Japan
The millipede genus Riukiupeltis was established for a single species Riukiupeltis jamashinai by Verhoeff, 1939 from the Ryukyu island Miyako, Japan (Riukiu and Mijako, in German).
Following several discussions on the genus in the past (
Fresh material of Riukiupeltis falcatus (Attems, 1953) and Riukiupeltis uenoi Murakami, 1975 was collected from Bi Doup National Park, Lam Dong province, Vietnam, and Iriomote-jima and Ishigaki-jima islands, the Ryukyu Archipelago, Japan, respectively. The type specimen of Riukiupeltis jamashinai was studied as a loan from the Bavarian State Collection of Zoology, Munich, Germany (BSCZ), whereas the holotype of Riukiupeltis uenoi was borrowed from the National Museum of Nature and Science, Tokyo (NMNS). Further material is shared between the University Museum (Fujukan) of the University of the Ryukyus, Okinawa (RUMF), the Hungarian Natural History Museum, Budapest (HNHM), and the Institute of Ecology and Biological Resources (IEBR), Hanoi, Vietnam. In addition, new material of Riukiupeltis jamashinai was identified in the collection by M. Shimojana, acquired in 1979 on Miyako-jima island.
Line drawings were made by using an Olympus SZX10 (ADN), and a Leica M125 (ZK) stereo microscope with drawing tube attached. SEM images were made by using a Hitachi S4800 scanning electron microscope. Colour photographs were taken by ZK using a Nikon D90 digital camera with macro lens and Leica microscope photo tube attached. The distribution map was generated using the software DIVA-GIS version 7.0.
Taxonomic accounthttp://species-id.net/wiki/Riukiupeltis
Gonofemorite strongly curved, distal part somewhat swollen and membraneous. Postfemoral region demarcated from femorite by obvious cingulum, and bent continuously forming almost a complete circle with femorite. Postfemoral regions consisting of a thick, strong and free solenomere, and an extremely short solenophore (= tibiotarsus).
Riukiupeltis jamashinai Verhoeff, 1939
This genus is relatively close to the genus Chamberlinius Wang, 1956, however, it definitely differs in gonopod conformation: femorite without lamina; solenophore very short, thick, and without any basal processes.
http://species-id.net/wiki/Riukiupeltis_jamashinai
Figs 1–2, 6A, 7A, 8A, Map 1Holotype male, in fragments – only 11 segments in 5 pieces – with segments around gonopods missing, Reg.-Nr. ZSMA20052252, and two slides with gonopods, Reg.-Nr. ZSMA20035204, and legpairs 1–7, Reg.-Nr. ZSMA20035205 (all BSCZ).
New records: 2 males, 2 females, Japan, Ryukyu Archipelago, Miyako-jima Island, Rinko-abu (cave), 21 August 1979; 8 males, 2 females, Japan, Ryukyu Archipelago, Miyako-jima Island, Fukumine-no-kara (cave), 25 August 1979; 1 male, 1 female, 1 juv., 1 fragment, Japan, Ryukyu Archipelago, Miyako-jima Island, Nishibe zuzaga (cave), 26 August 1979, all leg. M. Shimojana (in the collection of M. Shimojana, Okinawa).
Japan, Ryukyu Archipelago, Miyako-jima island.
Although after the description of Riukiupeltis jamashinai in 1939,
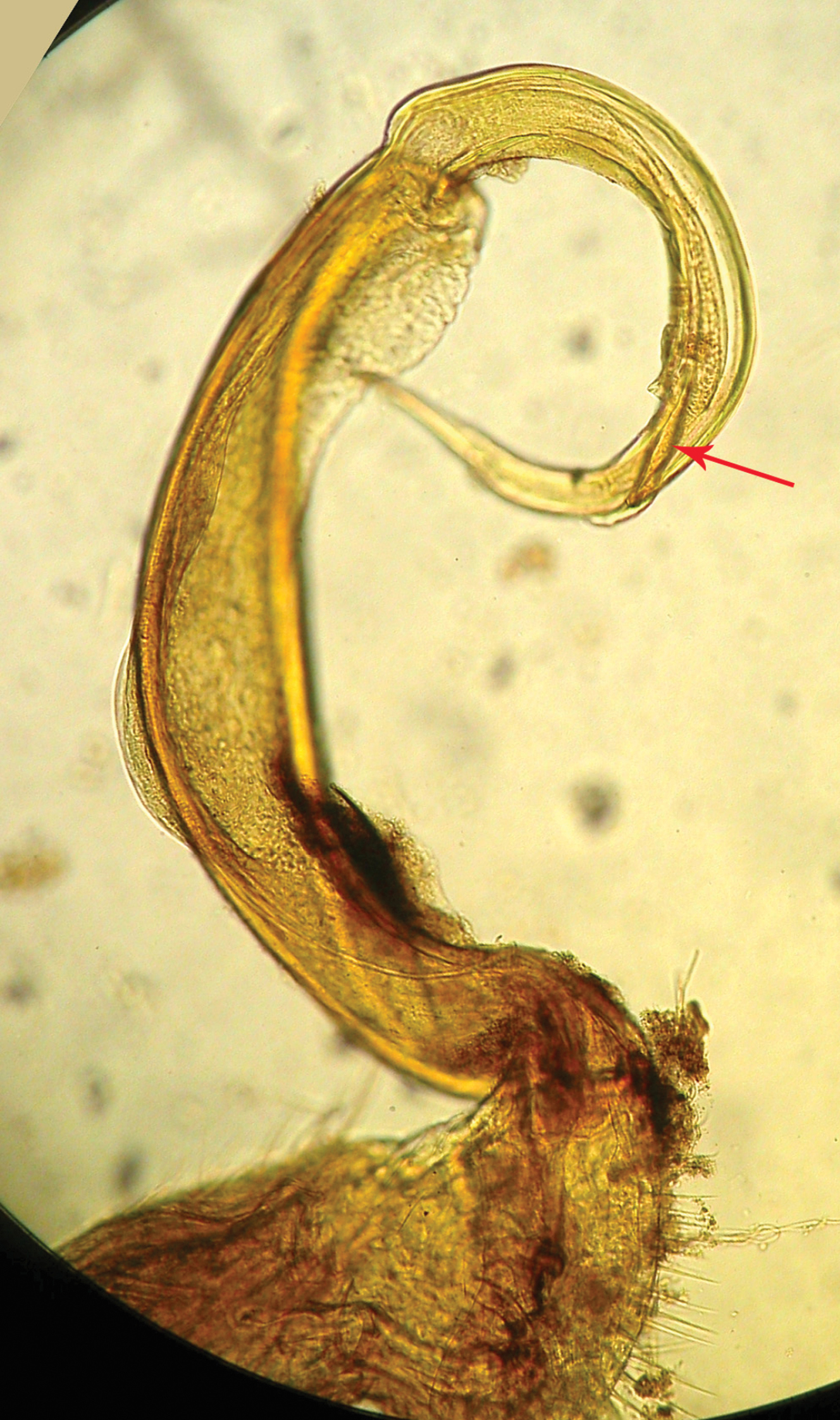
Riukiupeltis jamashinai Verhoeff, 1939, right gonopod preparation of holotype (slide Reg.-Nr. ZSMA20035204) (red arrow marks solenophore)
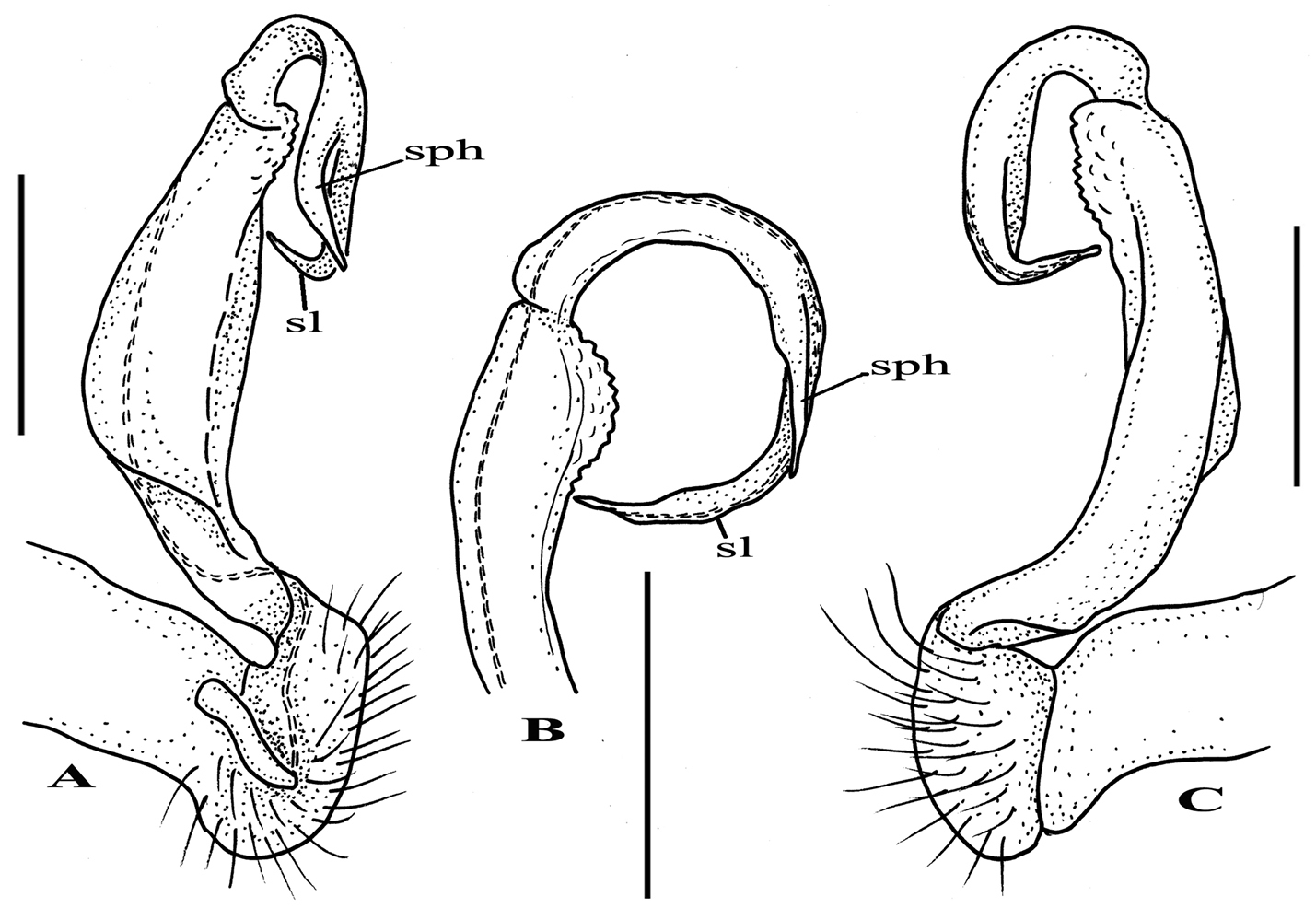
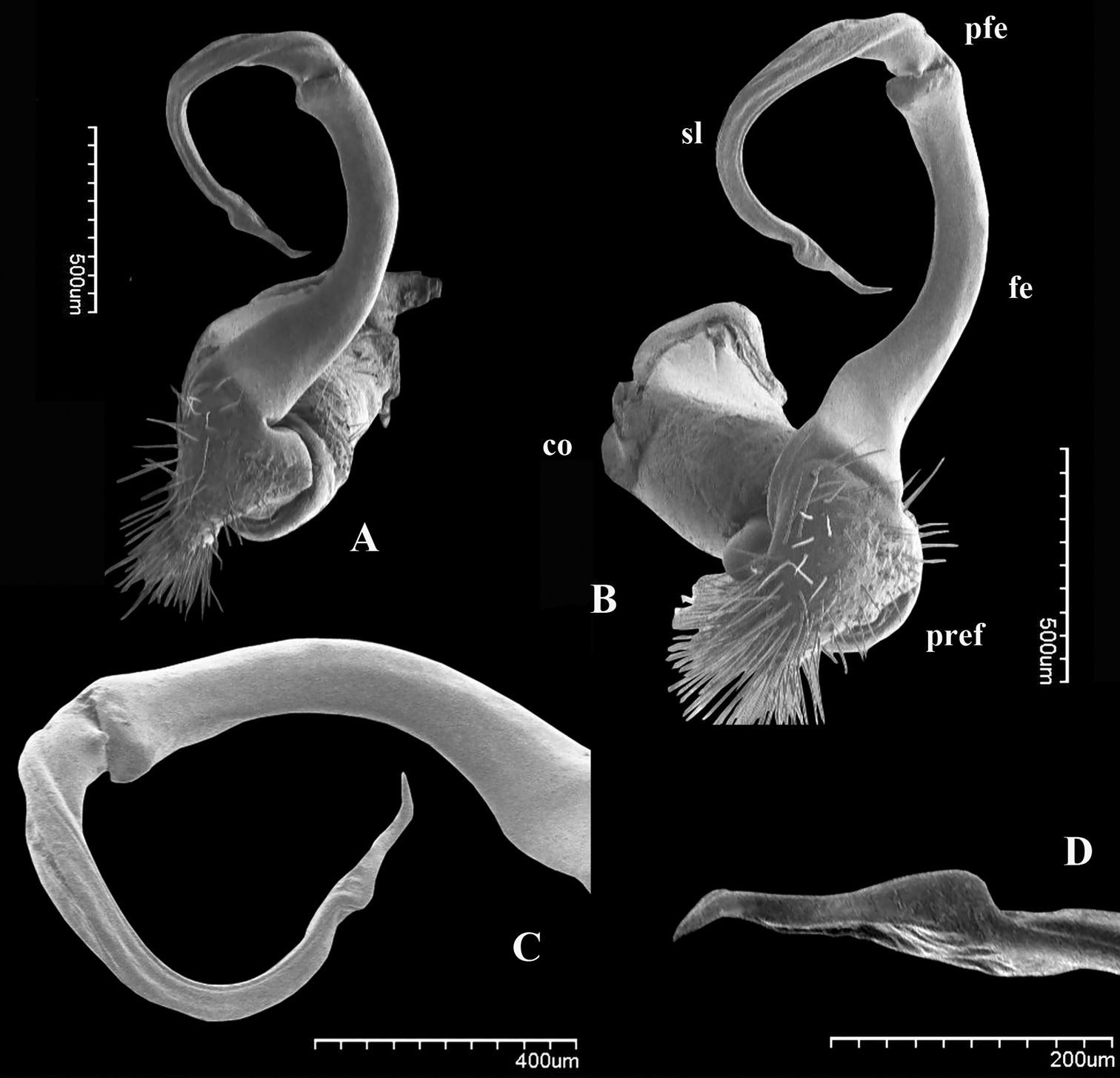
Riukiupeltis jamashinai Verhoeff, 1939, left gonopod of male from Fukumine-no-kara cave, Miyako-jima Island, 25 August 1979, leg. M. Shimojana A mesal view B dorsal view C ventro-lateral view (sl = solenomere, sph = solenophore or tibiotarsus). Scale bars = 0.5 mm.
http://species-id.net/wiki/Chamberlinius_uenoi
Figs 3, 6C, 7C, 8C, Map 1Holotype male, NSMT-My 358, “Japan, Okinawa Pref., Is. Ishigaki-jima, Ibaruma, Sabichi-go Cave, 31 July 1973, coll. by S. Uéno” (NMNS).
New records: 2 males, 1 female, Japan, Ryukyu Archipelago, Yaeyama Island Group, Ishigaki-jima Island, Banna-dake, secondary forest, N24.3859°, E124.1651°, 30 August 2009, leg. Z. Korsós & Y. Nakamura (RUMF); 1 male, 2 females, Japan, Southern Ryukyus, Yaeyama Group, Iriomote-jima Island, Funaura, around university research station, N24.3929°, E123.7913°, secondary forest, 18 January 2011, leg. Z. Korsós (RUMF); 1 male, 1 female, Japan, Southern Ryukyus, Yaeyama Group, Iriomote-jima Island, Mihara, along Aira river, primary forest, N24.3400°, E123.9137°, in decaying log, 12 m a.s.l., 19 January 2011, leg. Z. Korsós (HNHM).
. Japan, Ryukyu Archipelago, Yaeyama Group, Ishigaki-jima and Iriomote-jima islands.
After studying the type and freshly collected specimens, it became clear that the species uenoi is strongly different from Riukiupeltis jamashinai in its gonofemorite having a long lamina l, and a longer solenophore with basal processes pp and lp (Fig 3). We found that this species is more similar to Chamberlinius hualienensis Wang, 1956. Both Chamberlinius hualienensis and Chamberlinius uenoi comb. n. have well-developed paraterga (Figs 7A–B), large, slender and strongly concave gonofemorite, with a lamina at the mesal side (l in Fig. 3). Postfemoral region is demarcated from femorite by obvious cingulum (c in Fig. 3), and includes a long and large solenomere (sl) reaching femur, and a shorter solenophore (sph) with a basal lobe. However, the two species differ from each other in the length of the postfemoral processes, by the shape of the small basal processes on the solenophore, and by live colouration. The dark brown, transversal metatergal bands in Chamberlinius uenoi comb. n. are not divided by a median light brown longitudinal line (Figs 7C, 8C) as in Chamberlinius hualienensis (Figs 7B, 8B). Moreover, Chamberlinius uenoi comb. n. is strictly confined to undisturbed, natural evergreen broadleaf forests, and can only be found deep in decaying dead wood, whereas Chamberlinius hualienensis has a strong tendency for being synanthropic, and dispersed in large numbers onto many islands (especially in the southern part of Japan) by human activities.
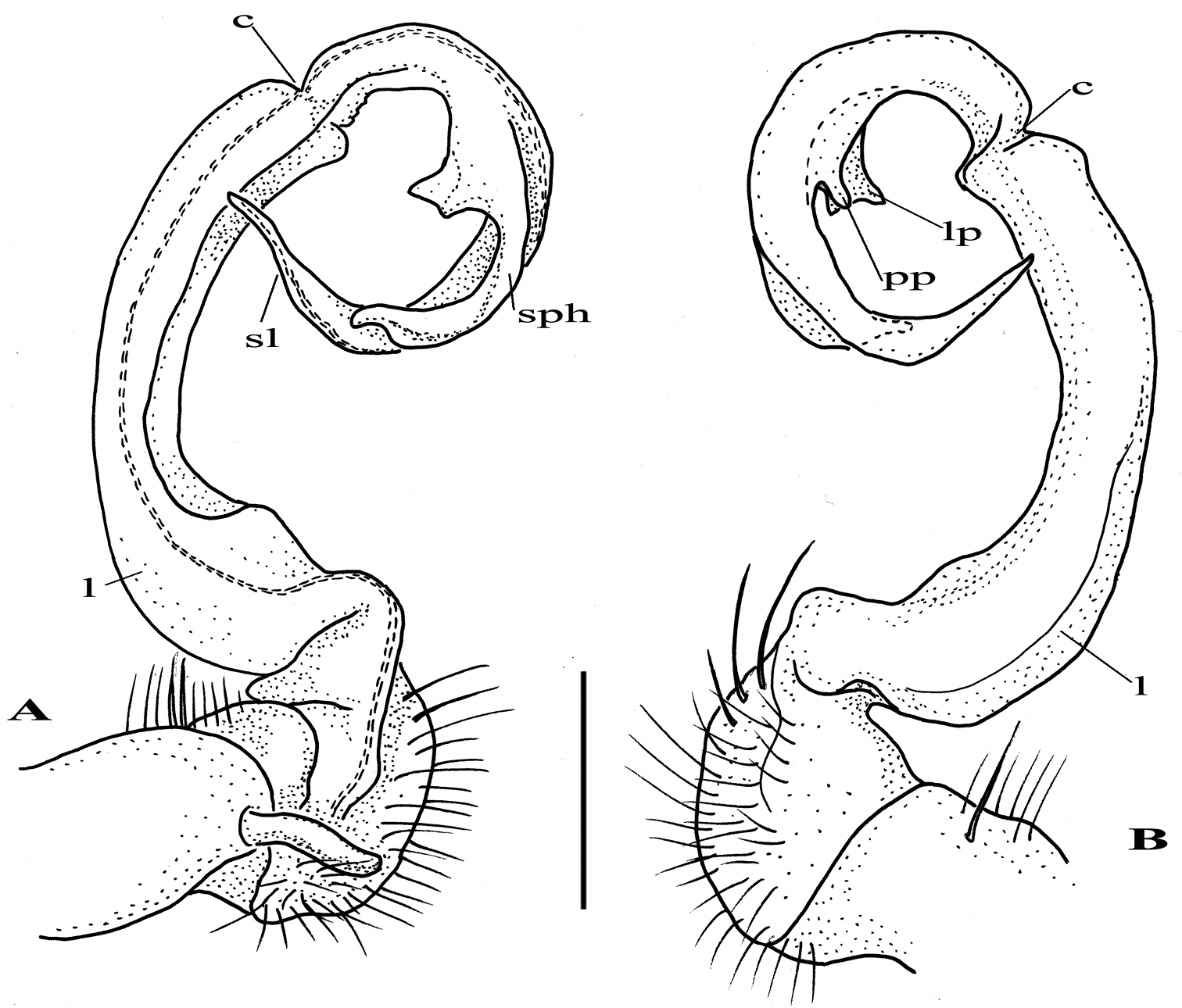
Chamberlinius uenoi (Murakami, 1975) comb. n., left gonopod of male Mt. Banna-dake, Ishigaki-jima Island A dorso-mesal view B lateral view. (l = lamina, c = cingulum, sl = solenomere, sph = solenophore or tibiotarsal process, lp = laminar process, pp = pointed process)). Scale bar = 0.5 mm.
urn:lsid:zoobank.org:act:878B9422-C9BB-462E-87A5-4A2610BB5B52
Haplogonosoma falcatum Attems, 1953, by present designation
This genus, Simplogonomorpha gen. n. can be distinguished from other genera within the tribe Chamberlinini Wang, 1956 (as defined by
A feminine noun to emphasize the simple gonopod conformation.
In fact, the solenophore of Riukiupeltis jamashinai still exists, although short, and somewhat hidden next to the solenomere, whereas solenophore of Simplogonomorpha gen. n. is totally missing. A comparison of genera in the tribe Chamberlinini is provided in Table 1.
Comparison of genera of Chamberlinini
| No. | Characters | Riukiupeltis | Chamberlinius | Simplogonomorpha | Aponedyopus | Haplogonosoma | Geniculodesmus |
|---|---|---|---|---|---|---|---|
| 1 | Paraterga | Well-developed | Well-developed | Modestly-developed | Poorly-developed | Poorly-developed | Poorly-developed |
| 2 | Sternal process between coxae 4 | Missing | Missing | Two separate cones | A single or bifid process | A single process | Linguiform |
| Gonopod conformation | |||||||
| 3 | Coxa | Long, cylindrical but not thick | Long, thick and subcylindrical | Thick and subcylindrical | Long and subcylindrical | Long and subcylindrical | Long and subcylindrical |
| 4 | Femorite | Long and slender, with a rugose membranous lamina at distal part of lateral side | Especially long, slender, simple without any outgrowths, however, sometimes, with a lamina at mesal side | Especially long, but simple and slender, devoid of any lobes or lamina. | Long, but broadened parabasally at dorsal side, without any lamina or processes | Long, slender and erected, a little bit broadened distally, devoid of any modifications | Long, slightly curved, not broadened at base |
| 5 | Postfemoral part | Demarcated by cingulum | Demarcated by cingulum | Demarcated by cingulum | Demarcated by obvious sulcus at lateral side | Set off by oblique sulcus at lateral side, apical part with a big tooth. | absent |
| 6 | Solenomere | Long, thick and simple, not sheathed by solenophore | Long, thick and simple, not sheathed by solenophore | Thick, but slender, and strongly curved downwards | Flagiliform, almost completely sheathed by solenophore, sometimes, only tip exposed | Long, slender and coiled, subfiliform | Long, flagiliform, almost sheathed by solenophore |
| 7 | Solenophore | Very short, simple, without any additional processes | As long as solenomere, with a basal dentiform outgrowth | Completely missing | Base with a small, obvious lobe. Tip with bifid lobe | Laminate, coiled, and as long as solenomere | Long and coiled, with a process at base |
http://species-id.net/wiki/Simplogonomorpha_falcata
Figs 4–5, 7D, Map. 13 males, 2 females, (IEBR-166), Vietnam, Lam Dong prov., Bi Doup-Nui Ba National Park, corn field, 1400m a.s.l., pitfall traps, 2–9 April 2008, leg. Anh. D. Nguyen; 1 male, 1 female, 1 juvenile, (IEBR-167), same locality, grasslands, 1400m a.s.l., pitfall traps, 2–9 April 2008, leg. Anh. D. Nguyen; 1 female, (IEBR-168), same locality, bushes near stream, 1400m a.s.l., pitfall traps, 25 April 2008, leg. Anh. D. Nguyen; 4 males, 2 females, (IEBR-169), same locality, evergreen forest, 1800m a.s.l., 25 March–23 April 2008, leg. Anh. D. Nguyen;1 male (IEBR-125), Vietnam, Khanh Hoa Prov., Hon Ba Mts., 1300–1500m a.s.l., primary forest, 15–24 April 2006, leg. Anh. D. Nguyen; 1 male, 1 female, (HNHM), same data as sample IEBR-125.
Head: yellowish-brown to blackish brown, a slightly paler toward labrum. Epicranial suture distinct, obviously deep.
Antennae: short and stout, yellowish-brown to blackish brown. Length of antennomere 2 subequal to that of antennomere 3, 4 or 5. Antennomere 6 shorter and claviform.
Body: yellowish-brown to blackish brown on terga, paler on paraterga and pleura. Body parallel-sided on somites 5–17, thereafter gradually tapering.
Surface of metaterga general fine in posterior part, but with small oblique or longitudinal rugulose in anteriormost part. Stricture dividing pro- and metazona deep, obvious and beaded. Prozona surface shagreened with fine microgranulation.
Transverse sulcus on metaterga starting from somite 5 and more evident on subsequent somites. Metaterga with a row of 2+2 setae in pre-sulcus part. Axial line vague.
Paraterga not well-developed, small, look like small keels in poreless segments, but more developed in pore-bearing somites. Ozopore located on lateral side, near tip of angular paraterga of segments 5, 7, 9–10, 12–13 and 15–19.
Pleura shagreened with fine microgranulation. Pleurosternal carinae rather well-developed in anteriormost segments, gradually decreasing posteriorly.
Epiproct truncated and curved down ventrad, with 4 strong setae on tip. Anal valves sub-semicircular with 1+1 long setae, and a deep emargination inbetween. Hypoproct trapeziform, with 1+1 setae.
Legs: yellow to yellowish brown, short and stout. Tarsal brushes present until legpair 10, sparsely until legpair 16, and completely missing thereafter.
Sterna: normal, sparsely setose, with two large tubercles between coxae 4.
Male gonopod: very simple, hook-like in dorsal view. Coxa subcylindrical, distoventral part sparsely setose. Prefemoral part usually densely setose, with evident demarcation from both femorite and coxa. Femorite slender, much longer than coxite and a little curved down distally, separated from postfemoral part by an evident, subtransverse sulcus laterally and mesally. Solenomere simple, strongly curved down, slender, and tapering at tip. Tibiotarsus totally absent.
Prostatic groove runs mesally along femorite, distolaterad, and turns to lateral side, then running mesally, and ending at tip of solenomere.
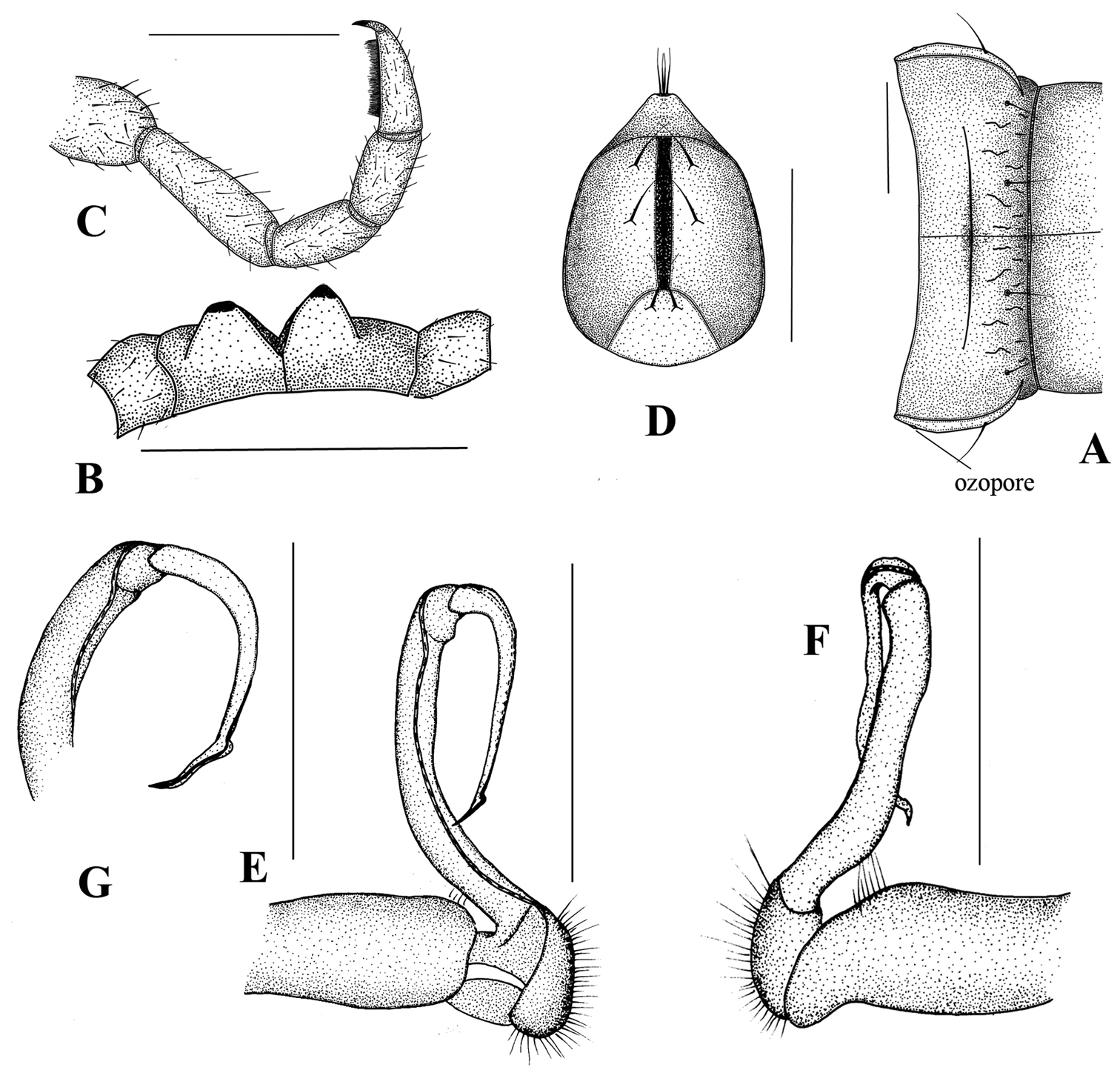
Simplogonomorpha falcata (Attems, 1953) comb. n. from Vietnam, BiDoup National Park A 10th body segment, dorsal view B sternal processes between 4th coxae, posteriovenral view C leg 10, lateral view D telson, ventral view E–G right gonopod E mesal F lateral, and G subdorsal view. Scale bar = 1 mm.
Simplogonomorpha falcata (Attems, 1953) comb. n. from Vietnam, BiDoup National Park, right gonopod: ventral A and mesoventral view B, C Tip of gonopod, ventral view D.
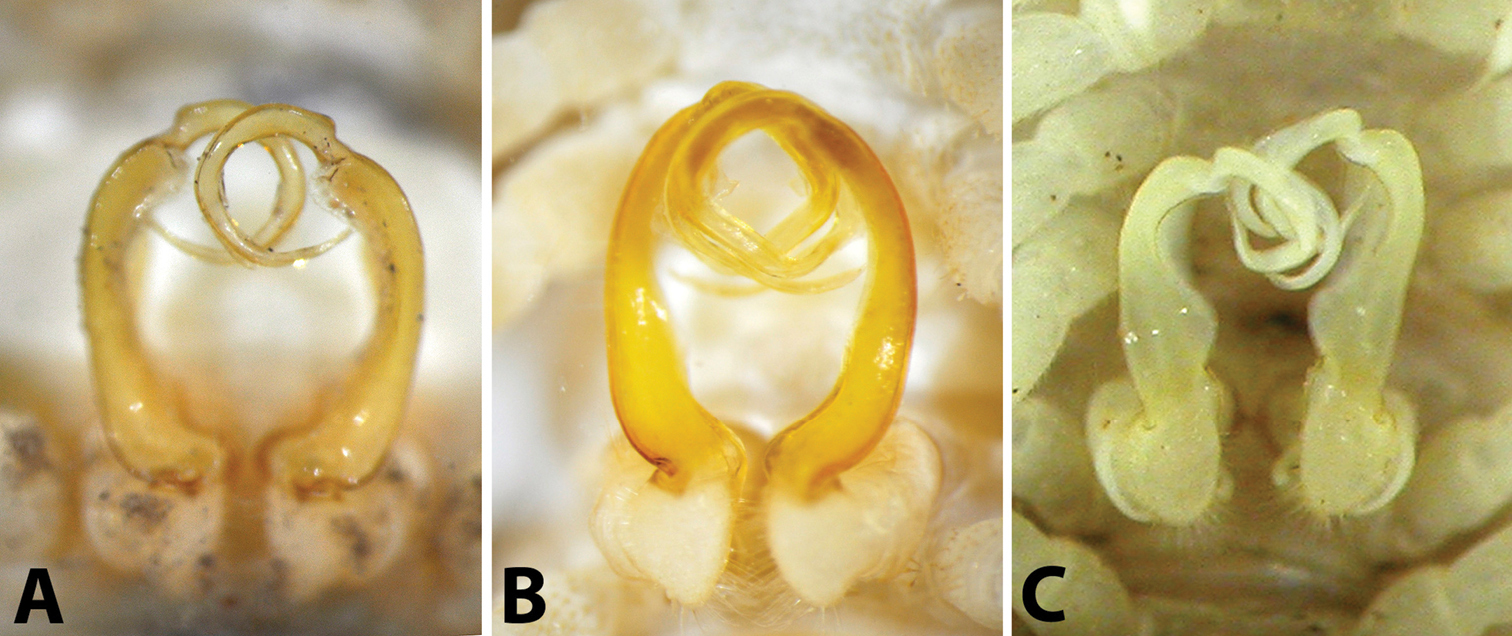
Male gonopods in situ, ventral view: A Riukiupeltis jamashinai (Miyako-jima Island) B Chamberlinius hualienensis (Okinawa-jima Island) C Chamberlinius uenoi comb. n.(Ishigaki-jima Island).
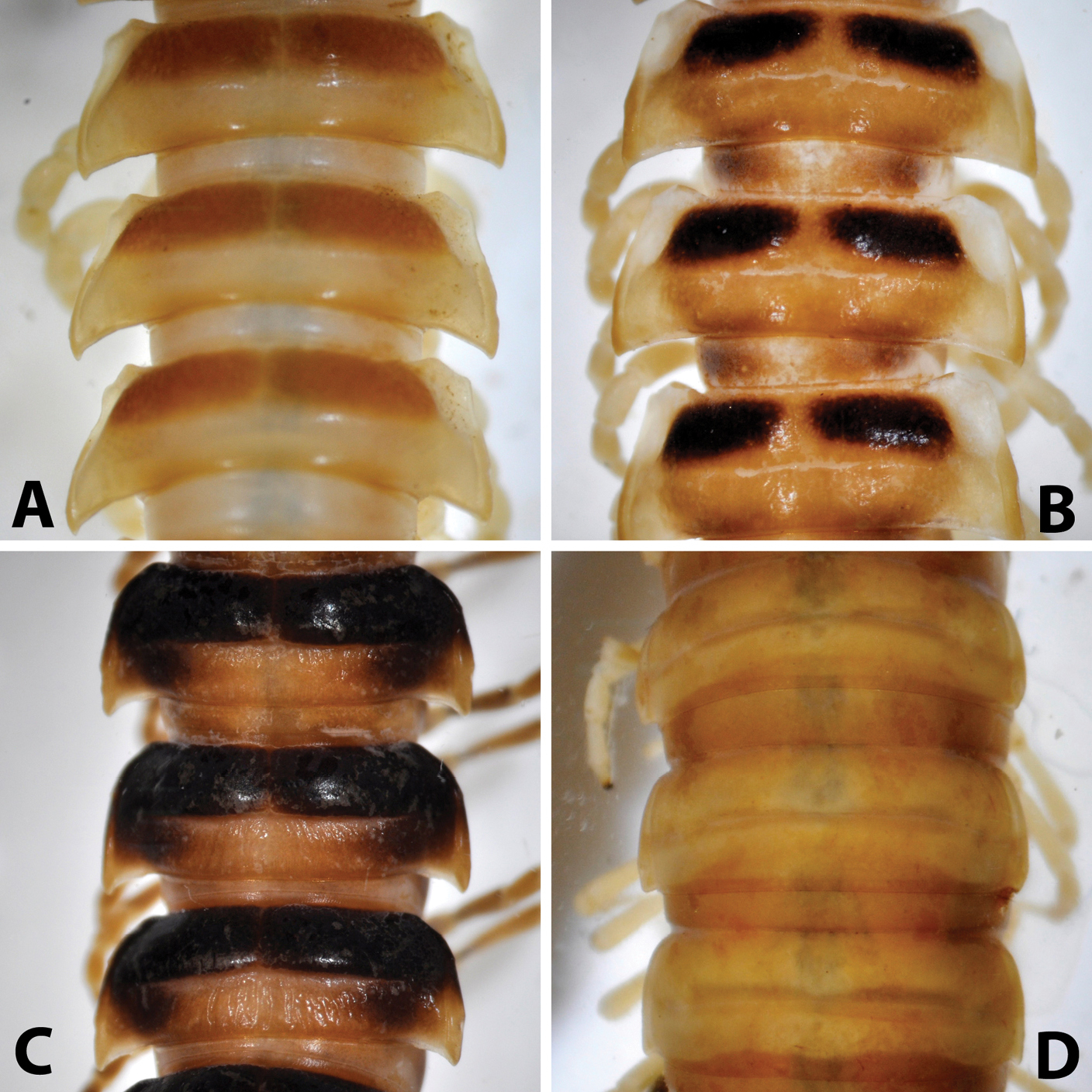
Midbody (11th-13th) segments, dorsal view: A Riukiupeltis jamashinai (Miyako-jima Island) B Chamberlinius hualienensis (Okinawa-jima Island) C Chamberlinius uenoi comb. n.(Ishigaki-jima Island) and D Simplogonomorpha falcata comb. n. (Vietnam, Hon Ba Mts.)

Habitus of millipedes: A Riukiupeltis jamashinai (live from Miyako-jima Island) B Chamberlinius hualienensis (live on Okinawa-jima Island) and C Chamberlinius uenoi comb. n.(live on Iriomote-jima Island).
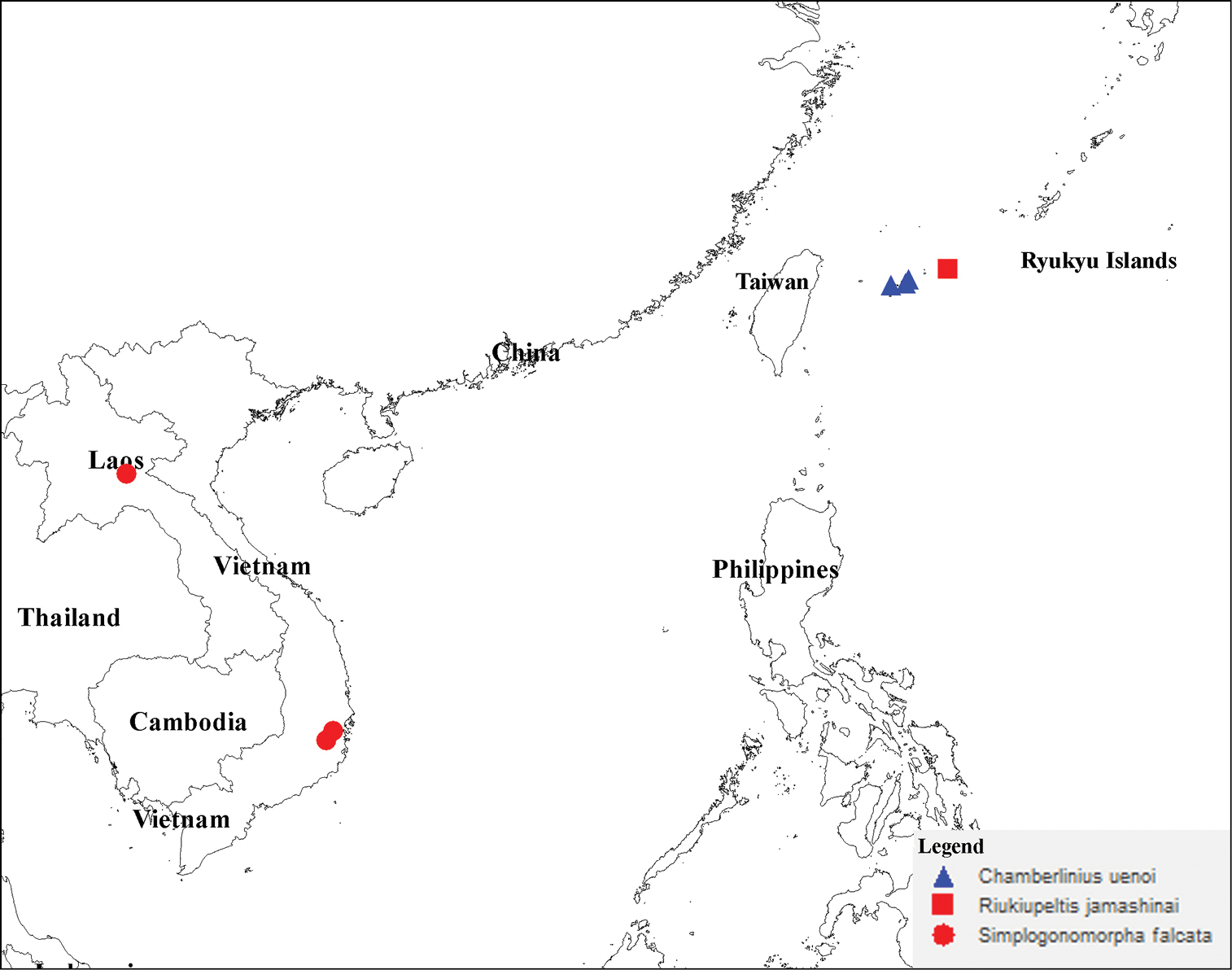
Distribution of three species Chamberlinius uenoi (Murakami, 1975) comb. n., Riukiupeltis jamashinai Verhoeff, 1939, and Simplogonomorpha falcata (Attems, 1953) comb. n.
Vietnam, Lam Dong province, Bi Doup-Nui Ba National Park; Khanh Hoa province, Hon Ba Mountain; Laos PDR, Xieng Khoang
New material does not much differ from Attems’s description. Only minor difference is the presence of two separate cones between coxae 4 instead of only one small conal process in Attems’s description. Recently,
Key to representatives to all three genera (based on male characters)
| 1 | Paraterga very weak, body looks almost cylindrical, colour uniformly light brown-yellowish. Gonofemorite very long and slender, without any modifications or processes. Postfemoral region consisting only a simple, strong, long solenomere. Solenophore totally absent (Figs 4–5) | Simplogonomorpha |
| – | Paraterga well-developed with strong, triangular, posterio-lateral processes. Dorsal metazona colouration divided into dark brown anterior and much lighter posterior half, transversely separated by a deep sulcus | 2 |
| 2 | Gonofemorite large, slender and strongly concave, with a lamina on the mesal side. Postfemoral region with a long, large solenomere and a shorter solenophore, the latter basally with two processes, a laminar mesal (lp) and a more pointed lateral one (pp) (Fig. 3B) | Chamberlinius |
| – | Gonofemorite only slightly curved, with a weak dorsal lamina, distal part swollen and membraneous. Postfemoral region consisting a thick, strong, free solenomere, and a short, somewhat hidden solenophore (Fig. 2) | Riukiupeltis |
In the key above, Riukiupeltis and Simplogonomorpha are represented by only one species each (jamashinai and falcata, respectively). Chamberlinius, on the other hand, includes at present five species: Chamberlinius hualienensis, Chamberlinius piceofasciatus, Chamberlinius pessior, Chamberlinius sublaevus (all keyed already by
As a result of our character comparisons, the taxonomic status of the following three species: Riukiupeltis jamashinai Verhoeff, 1939, Riukiupeltis uenoi Murakami, 1975 and Riukiupeltis falcatus (Attems, 1953) has been clarified. Based on significant morphological differences in body shape and gonopod structure, they all belong in three different genera. Riukiupeltis jamashinai having priority is maintained as such, whereas uenoi is transferred to Chamberlinius Wang, 1956, and a new genus, Simplogonomorpha gen. n., is erected to accommodate falcatum.
We would like to express our deepest thanks to the Vietnamese–Russian Tropical Centre (Vietnam), and the Bi Doup-Nui Ba National Park for supporting the first author during his field expeditions in Vietnam. Special thanks also to the AusAID program of the Australian government for providing financial support through its International Seminar Support Scheme for the first author to participate and present the paper at the 15th International Congress of Myriapodology.
The second author would like to express his sincere gratitude to Prof. H. Ota and M. Toda (Tropical Biosphere Research Center, University of the Ryukyus, Okinawa) for inviting him to study millipedes in the Ryukyus. His study is also supported by the Hungarian Research Fund (OTKA No. 69235). H. Ono (National Museum of Nature and Science, Tokyo) and S. Friedrich (Bavarian State Collection of Zoology, Munich) are deeply acknowledged for arranging the loans of appropriate type material. We also thank M. Shimojana (Urasoe, Okinawa) for letting us study his valuable unidentified material collected in Miyako-jima Island, and Y. Nakamura (Okinawa) for his help in the field and in literature search.